


I. Introduction
Nicotinic acetylcholine receptors (nACh receptors2) in jawed vertebrates are prototypical members of the multisubunit, neurotransmitter-gated superfamily of ion channels (ionotropic neurotransmitter receptors; see Section IV for selected seminal papers, reviews, and collections). nACh receptors mediate some of the effects of the endogenous neurotransmitter, acetylcholine, and they are principal biological targets of the tobacco alkaloid, nicotine, which generally (although exceptions exist) mimics acute actions of acetylcholine at nACh receptors. nACh receptors play critical physiological roles throughout the brain and body, mediating excitatory neurotransmission at the vertebrate neuromuscular junction, across autonomic ganglia, and at selected synapses in the brain and spinal cord. Roles have also been suggested for nACh receptors in the modulation of neurotransmitter release as well as in neurotrophism.
nACh receptors exist as a variety of types. At least one nACh receptor type is found in developing skeletal muscle, and another is expressed at the mature neuromuscular junction. There is evidence that several nACh receptor types exist in peripheral neurons of autonomic and sensory ganglia and in the brain and spinal cord. Evidence also exists for expression of nACh receptor types in other tissues and cell types, including lymphocytes, fibroblasts, pulmonary neuroendocrine cells, spermatozoa, keratinocytes, granulocytes, chondrocytes, and placenta, as well as in several sensory organs.
Classical pharmacological distinctions between some nACh receptor types endure. For example, decamethonium is a selective inhibitor of nACh receptor types in muscle, and hexamethonium is a selective antagonist of nACh receptor types in autonomic ganglia. More contemporary studies are beginning to yield distinctive pharmacological profiles for some nACh receptor types. However, these findings are not yet clearly integrated with molecular definitions of those receptors. This precludes a pharmacological description of nACh receptor types in this report, which instead focuses on matters of nomenclature and on structural bases of nACh receptor diversity.
Heterogeneity or diversity in nACh receptor types is derived in part from diversity in genes that encode nACh receptor subunits. Diverse nACh receptor types are formed as pentamers from different combinations of genetically distinct subunits, but not from all possible combinations of subunits (Fig. 1 and Table 1). Subunits confer distinctive functional and structural properties to the nACh receptor types that they form. With few exceptions, subunit compositions, stoichiometries, and arrangements of naturally expressed nACh receptor types are not known with absolute certainty.
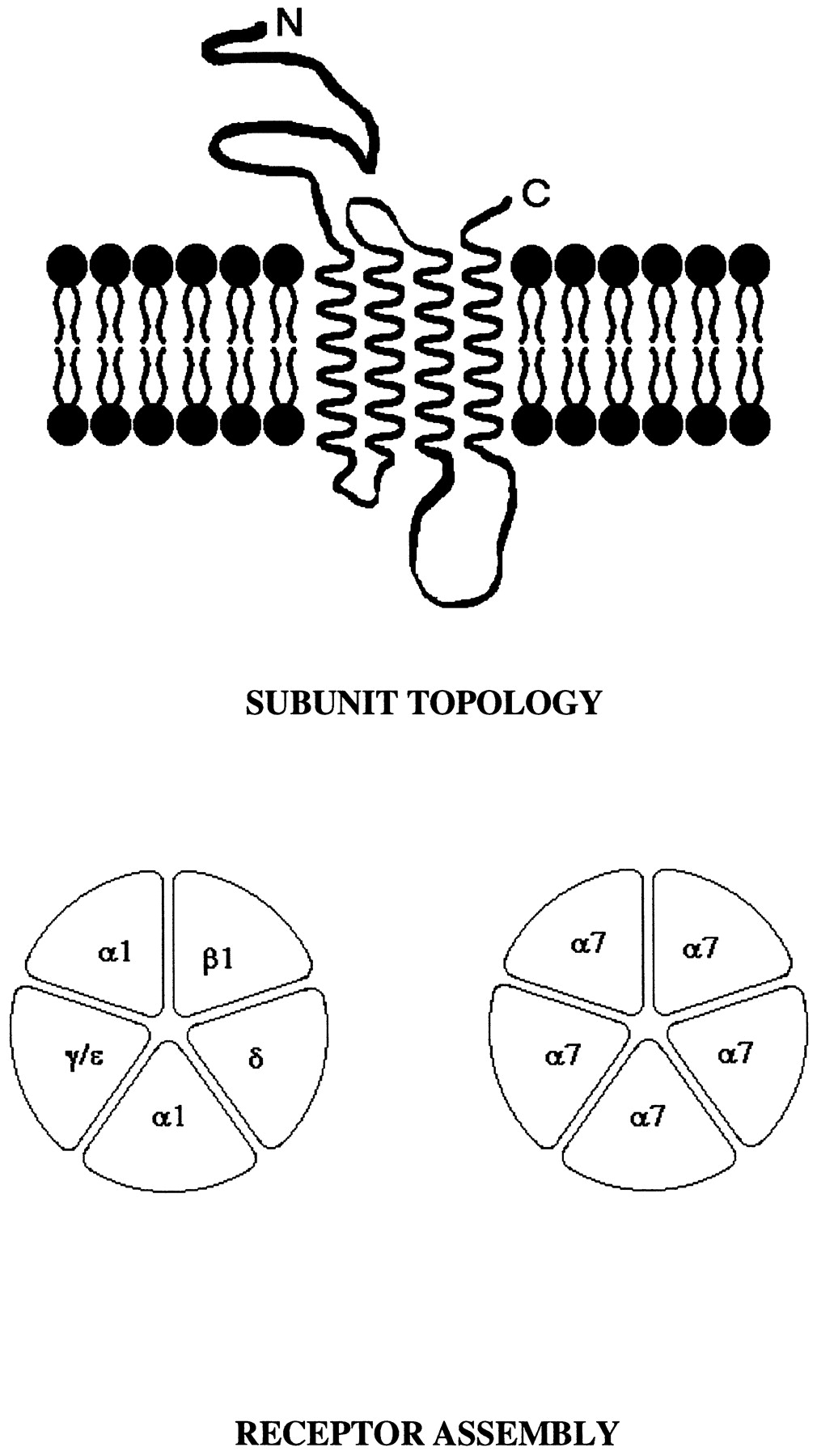
Schematic diagrams showing (upper) transmembrane topology of nAChR subunits and (lower) features of nAChR subunit assembly. Upper schematic illustrates the existence of an extensive, extracellular N-terminal region, four transmembrane segments, a cytoplasmic loop, and an extracellular C terminus for each nACh receptor subunit (not drawn to scale; surrounding lipid bilayer is also represented schematically). Lower schematic illustrates known arrangements of subunits constituting a nACh receptor type predominant in fetal skeletal muscle (left) and constituting a homooligomeric nACh receptor composed of α7 subunits (right). Assembly of subunits is thought to be regulated and limited by interactions between amino acid residues at subunit interfaces. Ligand binding pockets are thought to be formed at a subset of these assembly interfaces. For example, two distinct classes of ligand binding sites are formed at interfaces between α1 and γ subunits or between α1 and δ subunits of the nACh receptor from fetal muscle. Five largely similar ligand binding domains are thought to be formed at negative and positive faces of α7 subunits in homooligomeric α7-nACh receptors. All subunits contribute to channel formation, reflected by influences of every nACh receptor subunit identified to date on features of channel activity upon interaction with nicotinic ligand (see Table 1).
Subunit compositions of well established, naturally expressed and heterologously expressed nACh receptors
To date, 16 nACh receptor subunit genes have been cloned from vertebrates (Table 1). Each of the nACh receptor subunit proteins encoded by these genes is thought to have the same, general structural organization. The features of these integral membrane proteins include an extensive N-terminal region positioned extracellularly, four transmembrane segments (M1–M4), an extended cytoplasmic loop between the third and fourth transmembrane segments containing amino acid sequences absolutely unique to each subunit, and an extracellular C terminus (Fig. 1). All nACh receptor subunits also contain a loop of 13 amino acids between two cysteine residues forming a disulfide bridge in the N-terminal domain. These features are shared with families of subunits that constitute ionotropic glycine receptors, ionotropic γ-amino butyric acid receptors, invertebrate glutamate-gated chloride channels, and ionotropic serotonin receptors (5-HT3receptors). Invertebrate nACh receptor subunits can combine with vertebrate nACh receptor subunits to form functional receptors, suggesting that the former also belong to the superfamily. Comparisons between rat and human amino acid sequences typically reveal over 80% identity for a given nACh receptor subunit.
II. Current Status of nACh Receptor Subunit Nomenclature and Receptor Type Nomenclature and Classification
The IUPHAR Subcommittee on Nicotinic Acetylcholine Receptors of the International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification (NC-IUPHAR) recommends continued use of the present, Greek letter-based nomenclature for nACh receptor subunits, as is described in Section III, for the foreseeable future. The subcommittee also recommends using the suffix “gene” to distinguish the gene encoding a given subunit from the subunit protein itself (e.g., the δ subunit gene encodes the δ subunit). In addition, the subcommittee recommends for the foreseeable future a minimal nACh receptor type nomenclature/classification system based on the subunits that compose those receptor types to the extent that such information is available. Examples of such a nACh receptor type nomenclature and classification are also presented inSection III.
The NC-IUPHAR subcommittee consensus is that it is premature to create any alternative to the nomenclature for nACh receptor subunits or receptor types already in place or to generate a classification of nACh receptor types based on any of their properties other than their subunit composition. This consensus is based partly on the subcommittee’s perspectives that there are probably more nACh receptor subunit genes to be cloned, that there likely are more nACh receptor types to be identified, and that the current understanding is deficient about subunit compositions and other properties of many nACh receptor types. Although ligand binding properties, functional characteristics, and schemes for assembly of nACh receptor types differ, the subcommittee’s view is that more information is needed before these features can be exploited as criteria for an enduring system of nACh receptor type classification. The subcommittee’s consensus also recognizes the broad acceptance across the research community of the nACh receptor subunit and receptor type nomenclature/classification system already in place; there are very few deviations from this nomenclature in the literature. Moreover, the subcommittee’s opinion is that the nomenclature/classification system currently in use is reasonably sensible and understandable to the newly initiated, particularly given the inherent complexity of multisubunit, ionotropic neurotransmitter receptors relative to metabotropic neurotransmitter receptors composed of a single polypeptide. The subcommittee acknowledges limitations in the present system and that it is at variance with several of the guidelines established by NC-IUPHAR. For example, NC-IUPHAR prefers to avoid use of Greek letters but bows to historical precedence in this instance. However, the subcommittee does not see any obvious advantages to adoption of a new system at the present time, and the current system is viewed as having flexibility adequate to accommodate at least another series of advances in our understanding of the nACh receptor system. Nevertheless, it is anticipated that research progress over the next few years, including completion of the human genome project and identification of all nACh receptor subunit genes, will ultimately allow rational generation of enduring nACh receptor subunit nomenclature and nACh receptor type nomenclature/classification systems.
The NC-IUPHAR subcommittee recommends that commonly used prefixes such as “neuronal nACh receptor”, “muscle nACh receptor”, or “ganglionic/autonomic nACh receptor” not be adopted into nACh receptor nomenclature or classification. This is because there are many possible nACh receptor subunits or receptor types that can be found in any one of these tissues and because some nACh receptor subunits or receptor types may be found in more than one of these tissues. Such terminology applied to nACh receptor-directed, curaremimetic, snake neurotoxins (“neuronal-bungarotoxin”) instead of a neutral system (such as one based on Greek letters) has been misleading at best.
Similarly, it is recommended that use of the prefix “structural” not be propagated with regard to classification of nACh receptor subunits. This term currently has little use or meaning because evidence exists that each of the nACh receptor subunits identified to date participates in formation of ligand binding pockets and/or influences nACh receptor functional properties. Continued use of the term “non-α subunit” is also discouraged.
III. Present Definitions of nACh Receptor Subunits and Receptor Types
The 16 nACh receptor subunits identified to date are defined using a Greek letter sometimes followed by an Arabic numeral (neither subscripted nor superscripted). For example, there are nine alpha subunits (α1–α9; α8 subunits have only been derived from chick), four beta subunits (β1–β4), and one each gamma (γ), delta (δ), and epsilon (ɛ) subunit. All nACh receptor α subunits share expression of a pair of tandem cysteine residues in the putative, N-terminal, extracellular region. These residues are near to a site on the α1, α4, or α7 subunits known to engage in agonist binding. None of the other subunits (i.e., β1–β4, γ, δ, or ɛ) have these tandem cysteines. Several approaches have been taken in attempts to illuminate relationships between nACh receptor subunits and their genes. At a low level of resolution, these analyses make it clear that same nACh receptor subunits and their genes are most closely related (e.g., the β1, γ, δ, and ɛ grouping; uniqueness of the α7/α8 grouping and of the α9 subunit; grouping into an extended family of α2/α4 and α3/α6 pairs). However, each of these analyses is unique with respect to the range of amino acid or nucleic acid sequences examined and the analytical/computational tools applied, and each has produced somewhat different results, particularly with regard to placement of the α1 subunit and the β2/β4 and α5/β3 subunit pairs. The NC-IUPHAR subcommittee consensus is that these analyses are interesting and help guide those working in the field. However, further evolution in these approaches and identification and sequencing of nACh receptor subunits and their genes are needed to help guide the subcommittee and NC-IUPHAR in future efforts to derive nACh receptor subunit and receptor type classification/nomenclature.
It is useful to point out that the grouping of β1, γ, δ, and ɛ subunits is relevant to their contributions in formation of nACh receptor types in muscle, in combination with the α1 subunit. nACh receptors in vertebrate fetal muscle contain two α1 subunits and one each β1, δ, and γ subunit, whereas the ɛ subunit substitutes for the γ subunit in nACh receptors found in adult muscle. Thus, in vertebrates, one form of nACh receptor found in fetal muscle can be defined as the α1β1γδ-nACh receptor, and one form of nACh receptor found in adult muscle can be defined as the α1β1ɛδ-nACh receptor. Subscript and parenthetical notation is used to define subunit stoichiometries [e.g., (α1)2β1γδ-nACh receptors for nACh receptors containing two copies of the α1 subunit and single copies of β1, γ, and δ subunits]. Note the caveat here with respect to nACh receptor subunits and types found in muscle, as perhaps will be the case with regard to other nACh receptor subunits and types, that there is evidence for alternatively spliced products of α1 and γ subunit genes. Thus, there may be more than one kind of nACh receptor found in fetal or adult muscle, and a formal nACh receptor subunit and receptor type nomenclature will need to accommodate and define features of subunit variants and the nACh receptor types containing them.
The grouping of the α7/α8 subunit pair and the separation of the α9 subunit from the others is relevant in that nACh receptors containing these subunits happen to be capable of interaction with longer forms of curaremimetic, snake neurotoxins. These subunits also have the capacity to combine as homooligomers to create functional nACh receptor types. nACh receptors known to form only as homooligomers are defined, for example, as α7-nACh receptors, α8-nACh receptors, or α9-nACh receptors, with stoichiometries noted in subscript if known [e.g., (α7)5-nACh receptors]. nACh receptors containing both α7 and α8 subunits have been isolated from chick brain and/or retina and can be defined as α7α8-nACh receptors. Moreover, there is additional evidence that some nACh receptors might contain α7 plus other kinds of subunits. If, for example, nACh receptors known to contain α7 subunits possibly contain or are known to contain additional kinds of subunits as a pure or mixed population of receptors, the subcommittee recommends a notation employing an asterisk as a “wild card” to define those entities as α7*-nACh receptors.
There is strong evidence that α3 and β4 subunits coexist in naturally expressed nACh receptors found in autonomic ganglia and derived cell lines. There is additional evidence that these naturally expressed nACh receptors in autonomic neurons also contain α5 subunits, consistent with the fact that genes encoding α3, α5, and β4 subunits are clustered. Therefore, a more specific definition of these receptors would be α3α5β4-nACh receptors. Moreover, there is evidence that a subset of nACh receptors containing α3, α5, and β4 subunits also contain β2 subunits; these receptors can be defined as α3α5β2β4-nACh receptors. If a pure or mixed population of nACh receptors is known to contain α3 or α3 plus β4 subunits, but the identity of coassembled subunits is not certain, the subcommittee recommends use of α3*-nACh receptors or α3β4*-nACh receptors, respectively, to describe those entities.
A numerically abundant nACh receptor type in the central nervous system (CNS) is composed of α4 and β2 subunits, and data supporting a 2:3 stoichiometry exists. Thus, these receptors can be defined as α4β2-nACh receptors [or as (α4)2(β2)3-nACh receptors as information warrants]. However, minor populations of nACh receptors containing α4 and β2 subunits known to or possibly containing additional subunits can be defined employing the asterisk/wild card notation as α4β2*-nACh receptors.
The subunit compositions of nACh receptors naturally expressed at lower abundance in brain are not known, although there must be some roles for α2, α3, α5, α6, β3, and β4 subunits in formation of other nACh receptor types. Considerable evidence exists suggesting natural expression of functional nACh receptors in brain and spinal cord other than those containing just α7 or just α4 and β2 subunits, but further work is needed to determine the subunit compositions and stoichiometries of these entities. From heterologous expression studies, it is known that unique, ligand binding pockets can be formed at interfaces between α2, α3, α4, or α6 subunits and β2 or β4 subunits. Presence of α5 or β3 subunits has also been shown to influence ligand recognition of heterologously expressed nACh receptors containing other subunits. Heterologously expressed nACh receptors containing these subunits can be defined in a straightforward fashion, as knowledge about contributions of subunits to expressed nACh receptor warrants (e.g., α6β4-nACh receptors if composed only of α6 and β4 subunits). Similarly, at least provisional definitions of naturally expressed nACh receptors can be generated as information about the subunits constituting those nACh receptor is progressively obtained (e.g., α2*-nACh receptors for receptors known to contain α2 subunits).
Acknowledgments
We thank J. Brek Eaton of the Barrow Neurological Institute for composing Fig. 1.
Footnotes
-
↵1 Address for correspondence: Ronald J. Lukas, Division of Neurobiology, Barrow Neurological Institute, 350 West Thomas Rd., Phoenix, AZ 85013. E-mail: rlukas{at}mha.chw.edu
Abbreviations
- nACh receptor(s)
- nicotinic acetylcholine receptor(s)
- NC-IUPHAR
- International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification
- 5-HT
- 5-hydroxytryptamine (serotonin)
- CNS
- central nervous system
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}