


Abstract
The γ-aminobutyric acidB(GABAB) receptor was first demonstrated on presynaptic terminals where it serves as an autoreceptor and also as a heteroreceptor to influence transmitter release by suppressing neuronal Ca2+ conductance. Subsequent studies showed the presence of the receptor on postsynaptic neurones where activation produces an increase in membrane K+ conductance and associated neuronal hyperpolarization. (−)-Baclofen is a highly selective agonist for GABAB receptors, whereas the established GABAAreceptor antagonists, bicuculline and picrotoxin, do not block GABAB receptors. The receptor is Gi/Go protein-coupled with mixed effects on adenylate cyclase activity. The receptor comprises a heterodimer with similar subunits currently designated 1 and 2. These subunits are coupled via coiled-coil domains at their C termini. The evidence for splice variants is critically reviewed. Thus far, no unique pharmacological or functional properties have been assigned to either subunit or the variants. The emergence of high-affinity antagonists for GABAB receptors has enabled a synaptic role to be established. However, the antagonists have generally failed to establish the existence of pharmacologically distinct receptor types within the GABAB receptor class. The advent of GABAB1 knockout mice has also failed to provide support for multiple receptor types.
I. Introduction
The GABAB 1receptor was originally defined on the basis of pharmacological responses to GABA and related agonists, including baclofen (Bowery et al., 1981). In studies focusing on control of transmitter release, it was noted that a GABA receptor responsible for modulating evoked release in a variety of isolated tissue preparations differed pharmacologically from the receptor responsible for the Cl−-dependent actions of GABA. Thus, the ability of GABA to inhibit neurotransmitter release from these preparations was not blocked by bicuculline, was not mimicked by isoguvacine, and was not dependent on Cl−, all of which are characteristic of the classical GABA receptor. Most striking was the finding that baclofen (β-parachlorophenyl GABA), a clinically employed spasmolytic (Bein, 1972; Keberle and Faigle, 1972), mimicked, in a stereoselective manner, the effect of GABA in these systems. Furthermore, ligand-binding studies provided direct evidence of distinct attachment sites for baclofen on central neuronal membranes (Hill and Bowery, 1981). The term GABAB was coined to distinguish this site from the bicuculline-insensitive receptor, which was, in turn, designated GABAA(Hill and Bowery, 1981).
A major distinction between GABAA and GABAB receptors is that the former are ligand-gated ion channels, whereas the latter are coupled to G proteins (Wojcik and Neff, 1984; Hill, 1985; Karbon and Enna, 1985). Hence, GABAB receptors can be defined as metabotropic, whereas GABAA receptors form part of the ionotropic receptor superfamily. Characterization of the GABAB receptor has led to new insights into the structural and functional properties of seven-transmembrane receptors in general. Contained in this report is an overview of selected studies on the properties of GABAB receptors. Readers desiring additional information on particular aspects of this topic are urged to consult other sources (e.g., Enna, 1997; Enna and Bowery, 1997; Marshall et al., 1999a; Bowery and Enna, 2000; Enna, 2000). Characteristics of the GABAB receptor, which are described in detail in this review, are summarized in Table1.
GABAB receptor characteristics
II. γ-Aminobutyric AcidB Receptor Structure
Functional G protein-coupled receptors are expressed in cell membranes in different ways. In some cases they may be present as a single protein, whereas in others they may form homodimers (Bouvier, 2001). The structural characterization of the metabotropic GABAB receptor revealed a third possibility. In this case, the receptor exists as a heterodimer with the subunits designated GABAB1 and GABAB2 (Jones et al., 1998; Kaupmann et al., 1998a; White et al., 1998; Kuner et al., 1999; Martin et al., 1999; Ng et al., 1999). The heterodimeric nature of the GABAB receptor was not initially appreciated when the GABAB1 subunit was first cloned (Kaupmann et al., 1997). Although shown to be a high molecular weight, seven-transmembrane spanning protein with homology to metabotropic glutamate receptors, the recombinant GABAB1protein exhibited binding affinities for agonists that were 1000-fold lower than those for wild-type GABAB receptors. Moreover, the coupling to presumed GABAB effector systems in heterologous cells was surprisingly inefficient (Kaupmann et al., 1997, 1998b). Subsequent studies revealed that the GABAB1 protein is not transported to the plasma membrane but remains associated with the endoplasmic reticulum (Couve et al., 1998). This, and the inefficient coupling to effector systems, led to the hypothesis that a trafficking protein, such as a RAMP (receptor activity modifying protein) (McLatchie et al., 1998), might be required for the efficient functional expression of the GABAB site. Ultimately, the discovery of a second GABAB receptor subunit, GABAB2, provided the necessary explanation (seeMarshall et al., 1999a ). The GABAB2protein has 54% similarity and 35% homology to GABAB1 and has many of the structural features of the GABAB1 subunit, including a high molecular weight (110 kDa), seven-transmembrane domains, and a long extracellular chain at the N terminus. The GABAB2 protein not only serves to escort GABAB1 to the cell surface, it appears to be the receptor component that links to the G protein, whereas the GABAB1 subunit is necessary for agonist activation (Margeta-Mitrovic et al., 2000; Calver et al., 2001;Galvez et al., 2001; Pagano et al., 2001). It appears, therefore, that the agonist binds to a component of the GABAB1subunit, producing a conformational change in the protein complex that allows GABAB2 to engage and activate the G protein-coupled signaling system. In support of this model, it has been shown that GABAB2 must remain linked to GABAB1 after the dimer is inserted into the cell membrane to maintain receptor function ( Margeta-Mitrovic et al., 2000; Calver et al., 2001; Pagano et al., 2001 ). Recombinant GABAB2 is expressed at the cell surface in the absence of GABAB1, and early reports suggested that it could display some functionality under this condition (Kaupmann et al., 1998a; Kuner et al., 1999; Martin et al., 1999). Although it now appears unlikely that wild-type GABAB2 subunits can function alone in this way (Prosser et al., 2001; Schuler et al., 2001), there is no doubt that the coupling of GABAB2 with GABAB1 yields a fully functional GABAB receptor, with the GABAB1, rather than the GABAB2 component, displaying a high affinity for radiolabeled ligands (Kaupmann et al., 1998a; White et al., 1998). Indeed, the GABAB1 isoform, when expressed as part of the heterodimer, has increased agonist affinity similar to that of the wild-type receptor (Kaupmann et al., 1998a; White et al., 1998).
Several proteins other than GABAB2 have been shown to interact with GABAB1 (Nehring et al., 2000; White et al., 2000; Couve et al., 2001), but none of these complexes yields a functional receptor. It is possible that the interaction of GABAB1 or GABAB2 with transcription factors, such as activating transcription factor-4, may serve to regulate gene expression through a novel signal transduction pathway (Nehring et al., 2000; White et al., 2000).
The interaction of GABAB1 and GABAB2 within the cells appears crucial for the correct assembly of the heterodimer on the membrane surface. This has been demonstrated for both recombinant and wild-type GABAB receptors (Marshall et al., 1999b; Filippov et al., 2000; Chronwall et al., 2001). The interaction of the C-terminal coiled-coil domains, by masking the action of the retention motif RXRR present in the C terminus of GABAB1, ensures that only correctly assembled receptor complexes traffic to the cell surface (Margeta-Mitrovic et al., 2000; Pagano et al., 2001). Expression of the coupled heterodimer in cell membranes can occur even when the GABAB1 and/or GABAB2 C-terminal domains are missing (Calver et al., 2001; Pagano et al., 2001), suggesting that the coiled-coil structures are not essential for heterodimerization per se. Although it has been proposed that mGlu4R can associate with GABAB1 and traffic it to the cell surface (Sullivan et al., 2000), this finding could not be replicated in a subsequent study using a different experimental approach (Pagano et al., 2001). The critical importance of the GABAB1subunit is supported by the finding that tissue from mice lacking the gene for this protein fails to respond to GABABagonists and shows a loss of detectable pre- and postsynaptic responses (Prosser et al., 2001; Schuler et al., 2001). Importantly, the GABAB2 subunit is heavily down-regulated in GABAB1 null-mutant mice. This requirement of GABAB1 for stable GABAB2expression supports the notion that in wild-type mice virtually all GABAB2 protein is associated with GABAB1, in agreement with previous biochemical studies (Benke et al., 1999). The null-mutant mice generated on the 129Sv background only survive for 3 to 4 weeks postnatally, apparently due to recurrent seizures (Prosser et al., 2001), whereas those generated on the BALB/c background survive through adulthood even though they exhibit spontaneous seizures, hyperalgesia, hyperlocomotor activity, and memory impairment (Schuler et al., 2001). The viability of BALB/c mice lacking the GABAB1 subunit has allowed their characterization in GABAB receptor paradigms. GABAB agonist administration to BALB/c null-mutant mice failed to produce the typical muscle relaxation, hypothermia, or delta electroencephalogram waves observed in wild-type animals. These behavioral findings were paralleled by a loss of all biochemical and electrophysiological GABABresponses in the null-mutant mice. This demonstrates that GABAB1 is an essential component of pre- and postsynaptic GABAB receptors and indicates that most, probably all, brain GABAB receptors incorporate the GABAB1 subunit. Moreover, from the analysis of the GABAB1 null-mutant mice it follows that GABAB2 is unlikely to function as an autonomous receptor. Although these results are in line with previous work that failed to find any evidence for pharmacologically distinct GABAB receptor subtypes (Waldmeier et al., 1994), there remains the possibility that unidentified splice variants or GABAB1-associated proteins generate diversity.
Numerous splice variants of the GABAB1 subunit have been identified (Kaupmann et al., 1997; Isomoto et al., 1998;Pfaff et al., 1999; Calver et al., 2000; Schwarz et al., 2000; Wei et al., 2001a,b) with sometimes different names in rat and human. A comprehensive description of these variants is made possible by the complete sequence of the human and mouse GABAB1genes, which are contained within GenBank accession numbers AL031983and AL078630, respectively, and the nearly complete rat gene (Pfaff et al., 1999). The 1a splice variant in all three species contains all 23 conserved exons of the gene, with the first exon being untranslated and the transmembrane domains being encoded by exons 15 to 21. It should be noted that this number of exons differs from much of the literature because Pfaff et al. (1999), apparently through assembly errors in their rat gene sequence (GenBank accession numbers AF110796 andAF110797), failed to recognize introns that split exons 7 and 11 each into two exons. The existence of these exons can be confirmed using sequence from the rat genome sequencing project (http://www.ncbi.nlm.nih.gov/genome/seq/RnBlast.html). In addition,Pfaff et al. (1999) did not use any 5′ untranslated cDNA sequence and, thus, did not identify the first exon. The 1b splice variant initiates 5′ of exon 6, thereby producing an extended exon 6, which contains a new initiation codon, giving rise to an alternative amino-terminal sequence for the 1b protein. The amino-terminal sequence unique to the 1b variant is 47 versus 162 amino acids for the sequence unique to the 1a variant. Isoform-specific antibodies have shown both variants to be expressed in rat brain with 1a predominating before birth and 1b predominating in adults (Fritschy et al., 1999). A third variant, called 1e in both rat and human, skips exon 15, which leads to premature termination prior to the first transmembrane domain. Although this isoform can heterodimerize with GABAB2subunits, it appears to be unable to activate G protein-coupled, inwardly rectifying potassium channels or to inhibit cAMP production when coexpressed with GABAB1 subunits (Schwarz et al., 2000). Several variants have been observed in only one species. In humans, a variant called 1c is similar to the 1a variant but skips exon 4, resulting in the deletion of 63 amino acids. It is expressed at much higher levels in fetal brain than in adult brain (Calver et al., 2000;Martin et al., 2001). A rat variant, also called 1c, corresponds to an insertion of a 93-base exon located between exons 19 and 20, which results in the insertion of 31 amino acids into the beginning of transmembrane domain 5. Although the homologous region can be identified in the mouse gene, it is too poorly conserved to be functional due to the insertion of two bases, which disrupts the reading frame. No homologous exon is evident in the human gene. Thus, although the rat variant has been reported to be functional in vitro (Pfaff et al., 1999), it is unclear if it is functional in vivo. Rat variant 1d has a 567-base insertion corresponding to the failure to splice out intron 22. Rat variant 1f skips exon 5, resulting in the deletion of seven amino acids. Rat variant 1g has a 124-base insertion that extends the 5′ end of exon 5 by using an alternative splice acceptor. This insertion shifts the reading frame and results in a severely truncated protein. Aside from variants 1a and 1b, it is presently unknown whether any of these variants act as a subunit of physiological receptors. The 1a and 1b variants are not, strictly speaking, splice variants but instead appear to be transcription start site variants that originate in high guanosine-cytosine content (∼80%) regions of the gene separated by about 5 kilobases. Although such high GC content makes it difficult to make full-length cDNA, to map the transcription start sites, and could easily cause artifacts, variant-specific antibodies have provided critical evidence that both proteins are physiologically expressed at significant levels. Thus, it is appropriate to use the IUPHAR nomenclature reserved for significant splice variants, i.e., GABAB1(a) and GABAB1(b), for these two variants.
Partial cDNAs corresponding to two potential splice variants of the human GABAB2 subunit, called 2b and 2c, which delete 81 and 78 bases of the carboxyl-terminal encoding portion of the cDNA have been reported (Clark et al., 2000). Subsequent analysis of the human GABAB2 gene (Martin et al., 2001) has demonstrated that neither of the deleted regions correspond to an independent exon but instead reside within the last exon of the gene. There are no appropriate splice donor or acceptor consensus sequences that might act as alternative splice sites. The lack of such sites and the presence of short (4–5 bases) repeated sequences at the ends of the deletion regions suggest that they are polymerase chain reaction artifacts. Thus, there is currently no good evidence for splice variants of the GABAB2 subunit.
Although there is a 1:1 stoichiometry between GABAB1 and GABAB2 subunits in the functional receptor, production of the subunits appears to be regulated, at least in part, independent of one another (McCarson and Enna, 1999). Thus, whereas expression of GABAB1and GABAB2 mRNA increases in rat dorsal spinal cord following 24 h of hind paw inflammation, the increase in GABAB2 mRNA is significantly greater than for GABAB1. This supports the notion that GABAB receptor subunits may serve a variety of functions in the cell and could indicate that other, as yet unidentified, proteins may form functional heterodimers with GABAB1 subunits to form a functional receptor.
III. γ-Aminobutyric AcidB Receptor Effector Mechanisms
Effector mechanisms associated with neural GABAB receptors are the adenylate cyclase system and Ca2+ and K+ ion channels (Hill et al., 1984; Karbon et al., 1984; Hill, 1985; Inoue et al., 1985; Andrade et al., 1986; Xu and Wojcik, 1986; Dolphin et al., 1990; Bindokas and Ishida, 1991; Gage, 1992). GABAB receptor activation is mediated by G proteins that are members of the pertussis toxin-sensitive family Giα/Goα, in particular Gi2α (Odagaki et al., 2000; Odagaki and Koyama, 2001). However, some pertussis toxin-insensitive effects of baclofen have been noted in, for example, the magnocellular neurons of the paraventricular and supraoptic nuclei of the rat (Noguchi and Yamashita, 1999; Cui et al., 2000). In particular, it has been reported that presynaptic, compared with postsynaptic, GABAB receptor mechanisms are insensitive to pertussis toxin (Harrison et al., 1990). It has also been found that whereas exposure of spinal cord membranes to baclofen results in an increase in guanosine 5′-3-O-(thio)triphosphate binding in young rats, no such response can be obtained in membranes from animals older than 21 days. This would suggest that there may be a developmental change in the coupling of GABABreceptors and G proteins in the cord (Moran et al., 2001).
A. Adenylate Cyclase
GABAB agonists inhibit basal and forskolin-stimulated neuronal adenylate cyclase in brain slices (Xu and Wojcik, 1986; Knight and Bowery, 1996) through a G protein-dependent mechanism that results in a reduced level of intracellular cAMP. Activation of the GABAB receptor can also enhance cAMP formation in response to Gs-coupled receptor agonists, such as isoprenaline, in brain slices but not in isolated neuronal membranes, suggesting it entails activation of cytoplasmic cyclases (Enna, 2000). The physiological relevance of these effects on cAMP production has been confirmed by in vivo microdialysis experiments in the cerebral cortex of freely moving rats (Hashimoto and Kuriyama, 1997). Both baclofen and GABA reduced the increase in cAMP generated by an infusion of forskolin, and this was blocked by CGP54626, a selective GABAB receptor antagonist, substantiating the role of GABAB receptors in this response. Baclofen was also able to potentiate the increase in cAMP produced by isoprenaline in this in vivo preparation.
A direct GABAB-mediated increase in basal adenylate cyclase activity has been detected in membranes of rat olfactory bulb (Olianas and Onali, 1999). Interestingly, this effect is blocked by pertussis toxin, suggesting an involvement of Gi/Go rather than Gs protein.
B. Ion Channels
When activated, GABAB receptors decrease Ca2+ and increase K+conductance in neuronal membranes. The effect on Ca2+ conductance appears to be primarily associated with presynaptic P/Q- and N-type currents (Santos et al., 1995; Lambert and Wilson, 1996; Chen and van den Pol, 1998; Takahashi et al., 1998; Bussieres and El Manira, 1999; Barral et al., 2000), although facilitation of an L-type current in non-mammalian retina has also been described (Shen and Slaughter, 1999).
Modulation of K+ conductance appears to be linked primarily with postsynaptic GABAB sites and with perhaps multiple types of K+ channels (Wagner and Dekin, 1993, 1997; Lüscher et al., 1997; Harayama et al., 1998). Whereas a K+(A) current is thought to be coupled to GABAB receptors on presynaptic terminals in hippocampal cultures, changes in membrane K+ flux appear to be due to postsynaptic GABAB receptor activation (Saint et al., 1990).
Although suppression of Ca2+ influx is probably the most frequently observed response associated with presynaptic GABAB receptors (Doze et al., 1995; Wu and Saggau, 1995; Isaacson, 1997, 1998; Isaacson and Hille, 1997), a process independent of Ca2+ or K+ channels but perhaps linked with protein kinase C activation, has been reported in rodent CA1 hippocampal pyramidal cells (Jarolimek and Misgeld, 1997). This had been demonstrated previously but was then only apparent in rat hippocampal slices obtained in early postnatal life (Tremblay et al., 1995).
Low threshold Ca2+ T-currents, which are inactivated at normal resting membrane potentials, may also be involved in the response to GABAB receptor activation, at least within the thalamus (Scott et al., 1990). This postsynaptic hyperpolarization of long duration, which initiates Ca2+ spiking activity in thalamocortical cells, could contribute to the generation of spike and wave discharges associated with absence seizures (Crunelli and Leresche, 1991).
IV. γ-Aminobutyric AcidB Receptor Subtypes
Although GABAB receptors appear structurally heterogenous in the sense that several splice variants exist for the two subunits known, evidence for functionally distinct receptor subtypes is limited. Transmitter release studies with native and rat brain GABAB receptors suggest pharmacological differences between autoreceptors and heteroreceptors and even within heteroreceptors (Bonanno and Raiteri, 1993a; Banerjee and Snead, 1995;Teoh et al., 1996; Bonanno et al., 1997; Ong et al., 1998b; Phelan, 1999). Similarly, the dual action of GABABagonists on adenylate cyclase in brain slices would support the concept of receptor subtypes (Cunningham and Enna, 1996). The existence of pharmacologically distinct subtypes of a native receptor has traditionally been considered likely when the affinities of one antagonist for the hypothetical subtypes differed by at least one order of magnitude. In the case of GABAB receptors sited on terminals releasing GABA, glutamate, cholecystokinin, or somatostatin, not only do the antagonist affinities differ in some cases by more than two orders of magnitude but the orders of potency of some antagonists differ between receptors. Qualitatively similar results were obtained when GABAB receptor antagonists were tested on nerve endings isolated from human cerebrocortex (Fassio et al., 1994; Bonanno et al., 1996, 1997, 1999;Raiteri et al., 1996). The results with rat and human nerve endings are summarized in Table 2. Pharmacological differences also seem to exist between GABABautoreceptors inhibiting GABA release in rat cerebral cortex and spinal cord (Raiteri et al., 1989; Bonanno and Raiteri, 1992, 1993b; Bonanno et al., 1998). Thus, the evidence for pharmacologically distinct subtypes of the GABAB receptor derived from release studies appears to at least equate with some other receptor systems, which can boast the chrism of molecular biology. Also it would be quite surprising if the GABABreceptor was the only example of a metabotropic receptor without subtypes. Comparative data obtained with wild-type GABAB receptors and recombinant GABABl(b)/B2 receptors expressed in CHO cells indicate that the recombinant receptor, unlike the wild-type, is insensitive to the antagonists, phaclofen, saclofen, and CGP35348 (Wood et al., 2000). However, a comparison of GABABreceptors containing different isoforms of GABAB1in combination with GABAB2 in CHO cells indicate that these heterodimers are pharmacologically indistinguishable (Kaupmann et al., 1998a; Green et al., 2000).
GABABreceptor subtypes IC50values of antagonists (μM). The drugs were tested against (−)-baclofen.
In a recent report, Ng et al. (2001) presented data suggesting gabapentin, an anticonvulsant/analgesic agent, is a selective agonist at the GABAB1(a)/GABAB2site, compared with the GABAB1(b)/B2 receptors, expressed in oocytes. A similar conclusion was drawn from studies on wild-type GABAB receptors in mIL-tsA58 cells (Bertrand et al., 2001). Questions remain, however, whether this selectivity is generally detectable when testing wild-type receptors in nontransformed mammalian central nervous system tissue. In fact, the possible relationship between gabapentin and GABAb receptors has been examined independently by Lanneau et al. (2001). These authors believe that gabapentin is not a GABAB receptor agonist. In another study the antihyperalgesic effects of the GABAB agonists, baclofen and CGP35024, but not those produced by gabapentin, were blocked by CGP56433A, a GABAB receptor antagonist (Patel et al., 2001). Thus, the extent to which the actions of gabapentin are mediated in vivo by effects on GABAB receptors, remains to be conclusively demonstrated. Nevertheless, the possibility that under certain conditions gabapentin can activate a particular form of the GABAB receptor is an interesting observation.
Electrophysiological studies in mammalian brain suggest subtle distinctions between pre- and postsynaptic receptors (Colmers and Williams, 1988; Dutar and Nicoll, 1988b; Harrison et al., 1990;Thompson and Gähwiler, 1992; Deisz et al., 1997; Chan et al., 1998; Yamada et al., 1999). For example, the GABAB receptor agonist CGP44533 failed to induce an increase in postsynaptic membrane conductance whereas (−)-baclofen and CGP35024 did (Yamada et al., 1999) and, on comparing the effects of six GABAB receptor antagonists, it was found that 5- to 10-fold higher concentrations were required to block presynaptic as opposed to postsynaptic receptors in the rat hippocampus (Pozza et al., 1999). However, in general, the receptor ligands currently available do not reliably distinguish between potential subtypes. Unfortunately, studies in GABAB1 null-mice have also failed to provide any positive evidence for subtyping of the GABAB receptors (Prosser et al., 2001; Schuler et al., 2001).
V. γ-Aminobutyric AcidB Receptor Distribution
A. Central Nervous System
Within the mammalian brain, the highest density of GABAB binding sites is in the thalamic nuclei, the molecular layer of the cerebellum, the cerebral cortex, the interpeduncular nucleus, and the dorsal horn of the spinal cord (Bowery et al., 1987; Chu et al., 1990). In situ hybridization studies of mRNA for the GABAB1(a) and GABAB1(b) splice variants reveal they are distributed differentially in brain (Liang et al., 2000). Studies with rat and human cerebellum and spinal cord indicate that GABAB1(a) is associated with presynaptic receptors, whereas GABAB1b is located predominantly at postsynaptic sites, at least in cerebellum (Kaupmann et al., 1998b; Billinton et al., 1999; Bischoff et al., 1999;Princivalle et al., 2000; Towers et al., 2000). Elsewhere in the brain, however, the GABAB1(b) protein is in presynaptic terminals and the GABAB1(a) at postsynaptic sites (Benke et al., 1999; Princivalle et al., 2001). In the dorsal horn of the rat spinal cord, the density of GABAB1(a) is low, whereas in the dorsal root ganglia, which contain cell bodies of the primary afferent fibers, >90% of the GABAB1subunit mRNA is GABAB1(a), with GABAB1(b) comprising less than 10% of the total GABAB1 mRNA (Towers et al., 2000). Immunocytochemical studies provide support for this in revealing that the level of GABAB1(a) protein appears to be higher than of GABAB1(b) in the dorsal horn of the rat spinal cord (A. P. Princivalle and N. G. Bowery, unpublished observation). Similarly, in rat and human cerebellum, GABAB1(a) mRNA is detected over the granule cells, which send their excitatory fibers into the molecular layer to innervate the Purkinje cell dendrites (Kaupmann et al., 1998b;Billinton et al., 1999; Bischoff et al., 1999). Presumably, the GABAB receptors on the granule cell terminals modulate the output of the excitatory transmitter. In contrast, GABAB1(b) mRNA is associated with the Purkinje cell bodies, which express GABAB receptors on their dendrites in the molecular layer postsynaptic to the GABA-ergic stellate cells. However, the contrary arrangement has also been observed elsewhere in the brain. For example, GABAB1(a) subunits appear to be postsynaptic on cell bodies in the thalamocortical circuits (Princivalle et al., 2001). Thus, it would seem that a functional role, or cellular location, cannot be generally assigned to specific GABABreceptor subunit splice variants (Poorkhalkali et al., 2000;Princivalle et al., 2001).
The regional distribution of individual GABAB1and GABAB2 protein subunits is similar to that of the wild-type receptor, but in some brain areas such as the caudate-putamen, GABAB2 is not detectable, even though GABAB1 and the native receptor are present (Durkin et al., 1999; Margeta-Mitrovic et al., 1999; Clark et al., 2000). In addition, there appears to be very little GABAB2 mRNA, relative to GABAB1 mRNA, in the hypothalamus (Jones et al., 1998; Clark et al., 2000). These findings, along with those suggesting that GABAB1 and GABAB2subunit expression is not regulated in tandem (McCarson and Enna, 1999), support the existence of additional, as yet unidentified, GABAB receptor subunits.
B. Peripheral Organs and Tissues
Functional GABAB receptors are not restricted to the central nervous system. Thus, GABA has been known for some time to play an important role in modulating autonomic inputs to the intestine, and GABAB receptors are capable of mediating responses in other organs (Ong and Kerr, 1990). Moreover, GABAB receptor agonists inhibit relaxation of the lower esophageal sphincter in dogs, ferrets, and humans, and attenuate esophageal reflux by an inhibitory action on the vagus nerve (Blackshaw et al., 1999, 2000; Lehmann et al., 1999, 2000; Lidums et al., 2000;Smid and Blackshaw, 2000).
Studies monitoring functional GABAB responses suggest their presence in peripheral organs (see Bowery, 1993). More recently, Northern blot analysis and receptor protein immunoblotting has provided direct evidence for GABAB1 isoforms and GABAB receptors throughout the periphery of the rat (Castelli et al., 1999; Calver et al., 2000). However, the GABAB2 subunit was not always present with GABAB1, such as in uterus and spleen (Calver et al., 2000).
Western blotting revealed the presence of GABAB1and GABAB2 proteins in rat heart myocytes, supporting the observation that baclofen influences inwardly rectifying K+ currents in these cells (Lorente et al., 2000). Moreover, photoaffinity-labeling studies suggest that GABAB1(a) and GABAB1(b) are differentially distributed in the periphery as well as in the central nervous system (Belley et al., 1999). Thus, GABAB1(a) is present in the adrenals, pituitary, spleen, and prostate, whereas GABAB1(b), but not GABAB1(a), is found in the rat kidney and liver.
VI. γ-Aminobutyric AcidB Receptor-Mediated Responses
A. γ-Aminobutyric AcidB Receptor Agonists
The observation that β-[4-chorophenyl] GABA (baclofen; Fig.1), is a stereospecifically active agonist at the GABAB receptor (Bowery et al., 1980, 1981) provided part of the original evidence for the existence of a distinct receptor. Since then 3-aminopropyl-phosphinic acid (3-APPA, CGP27492; Bittiger et al., 1988) and its methyl homolog (3-APMPA, CGP35024 identical with SK&F 97541; Froestl et al., 1992, 1993; Howson et al., 1993) have emerged and are reported to be 3- to 7-fold more potent than the active isomer of baclofen [IC50values, i.e., inhibition of binding of [3H]CGP27492 to GABABreceptors on rat cerebral cortex membranes: baclofen: 107 nM, (R)-(−)-baclofen: 32 nM, 3-APPA (CGP27492): 5 nM, 3-APMPA (CGP35024): 16 nM]. The latter compounds are also available as tritiated radioligands (Bittiger et al., 1988; Hall et al., 1995). Other methyl phosphinic acid-based agonists have been produced (Froestl et al., 1995a), such as CGP44532 (IC50 = 45 nM) and its (R)-(+)-enantiomer CGP44533 (IC50 = 152 nM; racemate CGP34938: IC50 = 77 nM; Fig. 1), which differ only by a factor of 3 in binding to GABAB receptors and show comparative activities in biochemical paradigms (Ong et al., 2001) but show significant differences in electrophysiological experiments (Yamada et al., 1999).

Structures of high-affinity GABABreceptor agonists.
Interestingly, ethyl (and higher homolog) phosphinic acids derivatives (e.g., CGP36216, IC50 = 2 μM; Fig.2) show effects of GABAB receptor antagonists (Froestl et al., 1995b). The difluoromethyl phosphinic acid derivative CGP47656 (IC50 = 89 nM; Fig. 1) with a substituent, the size of which is between a methyl and an ethyl group, showed properties of a partial GABAB receptor agonist (Froestl et al., 1995a). GABAB receptor agonists display a number of pharmacological effects, including central muscle relaxation, antitussive action, bronchiolar relaxation, inhibition of urinary bladder contraction, an increase in gastrointestinal motility, epileptogenesis, suppression of cocaine, nicotine, and opioid self-administration, antinociception, yawning, hypotension, brown fat thermogenesis, cognitive impairment, inhibition of gastric acid secretion, and inhibition of hormone release.

Structures of low-affinity GABABreceptor antagonists.
1. Antispasticity.
The centrally mediated muscle relaxant effect of baclofen is the most widely exploited clinical response to this agent. This action appears due to a baclofen-induced reduction in neurotransmitter release onto motoneurons in the ventral horn of the spinal cord. There is also a suggestion that the antispastic effect is due to post- rather than presynaptic action on motoneurons (Orsnes et al., 2000a). Regardless of the site of action, the efficacy of baclofen in alleviating spasticity has made it a drug of choice for this condition, although side effects, principally sedation, limit its utility. Baclofen is effective in treating spasticity associated with tardive dystonia, brain and spinal cord injury, cerebral palsy, tetanus, multiple sclerosis, and stiff-man syndrome (Ochs et al., 1989,1999; Penn et al., 1989; Penn and Mangieri, 1993; Becker et al., 1995,1997, 2000; Campbell et al., 1995; Seitz et al., 1995; Albright et al., 1996; Azouvi et al., 1996; Dressnandt and Conrad, 1996; Ford et al., 1996; Paret et al., 1996; Armstrong et al., 1997; Dressler et al., 1997; Dressnandt et al., 1997; François et al., 1997, 2001;Meythaler et al., 1997; Gerszten et al., 1998; Auer et al., 1999;Orsnes et al., 2000b; Trampitsch et al., 2000; Krach, 2001).
2. Antinociceptive.
Even though baclofen is used to treat migraine headache, musculoskeletal pain, and the pain associated with trigeminal neuralgia, stroke, and spinal cord injury, its general effectiveness as an analgesic is limited (Fromm, 1994; Taira et al., 1995; Hansson and Kinnman, 1996; Loubser and Akman, 1996; Hering-Hanit, 1999; Idänpään-Heikkilä and Guilbaud, 1999;Becker et al., 2000). Although the reason for this is unknown, it may be due to a rapid desensitization of GABABreceptors.
Laboratory animal studies have established the antinociceptive action of GABAB receptor agonists and suggest it is mediated by effects in both spinal cord and brain (Cutting and Jordan, 1975; Levy and Proudfit, 1979; Liebman and Pastor, 1980; Kendall et al., 1982; Sawynok and Dickson, 1985; Vaught et al., 1985; Serrano et al., 1992; Hammond and Washington, 1993; Smith et al., 1994; Dirig and Yaksh, 1995; Thomas et al., 1995; McCarson and Enna, 1996; Thomas et al., 1996; Wiesenfeld-Hallin et al., 1997; Cui et al., 1998; Przesmycki et al., 1998). Further evidence supporting the analgesic potential of GABAB agonists is provided by the finding that tiagabine, a GABA uptake inhibitor, displays antinociceptive activity in rodents that is blocked by CGP35348, a GABABreceptor antagonist (Ipponi et al., 1999). Inasmuch as intrathecal administration of a GABA-producing neuronal cell line permanently reverses neuropathic pain, it has been suggested that altered spinal GABA levels may contribute to the induction phase of chronic pain (Eaton et al., 1999).
In the dorsal horn of the rat spinal cord, GABABreceptors are located on small diameter afferent fiber terminals, with activation of these sites decreasing the evoked release of sensory transmitters, such as substance P and glutamate (Price et al., 1987;Malcangio and Bowery, 1996; Ataka et al., 2000; Iyadomi et al., 2000;Riley et al., 2001). It has also been suggested that modulation of voltage-dependent, nifedipine-sensitive calcium channels in dorsal horn neurons may contribute to the antinociceptive effects of GABAB agonists (Voisin and Nagy, 2001).
Baclofen and CGP35024 reversed neuropathic mechanical hyperalgesia following single s.c. or intrathecal administration but did not affect inflammatory mechanical hyperalgesia (Patel et al., 2001). GABAB receptor agonists, such as baclofen and CGP44532, inhibit the formalin-induced increase of the expression of neurokinin-1 receptor mRNA (Enna et al., 1998).
The hot-plate, tail-flick as well as the paw pressure techniques were used to characterize acute pain behaviors in GABAB1 null-mutant mice (Schuler et al., 2001). The tail-flick is a reflex response to a noxious thermal stimulus applied to the tail and is generally taken to represent a spinal reflex response, whereas the hot-plate response to a noxious thermal stimulus to the plantar surface of the paws is thought to involve supraspinal sites. In these nociceptive tests, GABAB1null-mutant mice showed pronounced hyperalgesia to noxious heat in the hot-plate and tail-flick tests and reduced paw withdrawal thresholds to mechanical pressure. From these data it is likely that GABAB-mediated effects do indeed exert a tonic control of nociceptive processes in the naive animal. The sites for this action are expected to be both spinal and supraspinal, although further experiments are needed to confirm this.
3. Suppression of Drug Craving.
Preliminary data suggest that baclofen reduces the craving for cocaine in humans (Ling et al., 1998). In rats, baclofen suppresses cocaine self-administration at doses that do not affect responding for food reinforcement (Roberts and Andrews, 1997; Shoaib et al., 1998; Campbell et al., 1999; Munzar et al., 2000). Moreover, the selective GABAB receptor agonist CGP44532 mimics this action of baclofen without disrupting the response for food (Brebner et al., 1999, 2000a,b). Similar results are obtained whether laboratory animals are administered the GABAB agonist either systemically or directly into select brain regions (Corrigall et al., 2000).
The importance of these observations in the possible treatment of drug abuse is reinforced by the finding that baclofen reduces craving for a host of addictive substances, including heroin, alcohol, and nicotine (Xi and Stein, 1999; Corrigall et al., 2000; Lobina et al., 2000). Thus, elevation of endogenous GABA levels in the mesolimbic area by administration of vigabatrin, an inhibitor of GABA metabolism, or the GABA uptake inhibitor NO-711, attenuates heroin and cocaine self-administration in rats and prevents cocaine-induced increases in dopamine in this brain region (Ashby et al., 1999; Xi and Stein, 2000). It has also been reported that gabapentin, a putative GABAB receptor agonist (Bertrand et al., 2001; Ng et al., 2001), reduces the craving for cocaine in dependent adults (Myrick et al., 2001).
There is preclinical and preliminary clinical evidence that baclofen is effective in reducing alcohol craving and intake (Addolorato et al., 2000; Colombo et al., 2000). Baclofen blocks the rapid tolerance to ethanol, an effect that can be blocked by GABABantagonists such as CGP36742 and CGP56433A (Zaleski et al., 2001).
4. Miscellaneous Actions.
GABABreceptors in the hypothalamus and nucleus tractus solitarius modulate sympathetic nerve activity, resulting in an elevation in blood pressure (Takenaka et al., 1995). GABAB receptor activation in the hypothalamus also leads to an increase in metabolic rate and brown fat thermogenesis (Addae et al., 1986). In addition to the above, other centrally mediated effects of GABAB agonists include alterations in epileptogenesis, cognition (Tang and Hasselmo, 1996), yawning (Doger et al., 1989), and micturition (Kontani et al., 1988).
Baclofen has been shown to have anti-bronchoconstrictor activity through activation of presynaptic receptors on parasympathetic nerve terminals (Chapman et al., 1991). In addition, through an action on sensory nerves in the lung, baclofen is reported to inhibit nonadrenergic, noncholinergic bronchoconstriction (Belvisi et al., 1989).
Baclofen has also displayed antitussive activity in humans and laboratory animals (Bolser et al., 1993, 1994; Dicpinigaitis and Dobkin, 1997). This effect is mediated through both a direct action on peripheral nerves in the lung as well as receptors in the brain stem controlling the cough reflex. Through a similar mechanism, possibly involving GABA-ergic inputs from the nucleus raphe magnus (Oshima et al., 1998), baclofen is effective in the management of intractable hiccoughs (Guelaud et al., 1995; Nickerson et al., 1997; Kumar and Dromerick, 1998; Marino, 1998). Baclofen has also been reported to inhibit the growth of mammary cancer cells in mice and humans, and there appears to be a correlation between glandular GABA levels and mammary pathology (Opolski et al., 2000).
B. γ-Aminobutyric AcidB Receptor Antagonists
The design and development of selective, high-affinity GABAB receptor antagonists have been important in establishing the significance and isolation of the GABAB receptor genes. Kerr, Ong and their colleagues (1987, 1988; Fig. 2) described phaclofen, saclofen, and 2-hydroxysaclofen, the first selective antagonists. Although these agents have low affinities (IC50 values, i.e., inhibition of binding of [3H]CGP27492 to GABAB receptors on rat cerebral cortex membranes: 130, 26, and 11 μM, respectively) for GABABbinding sites in rat brain membranes, as the first antagonists they were important tools for defining the pharmacological and physiological relevance of GABAB receptors (Dutar and Nicoll, 1988a; Karlsson et al., 1988).
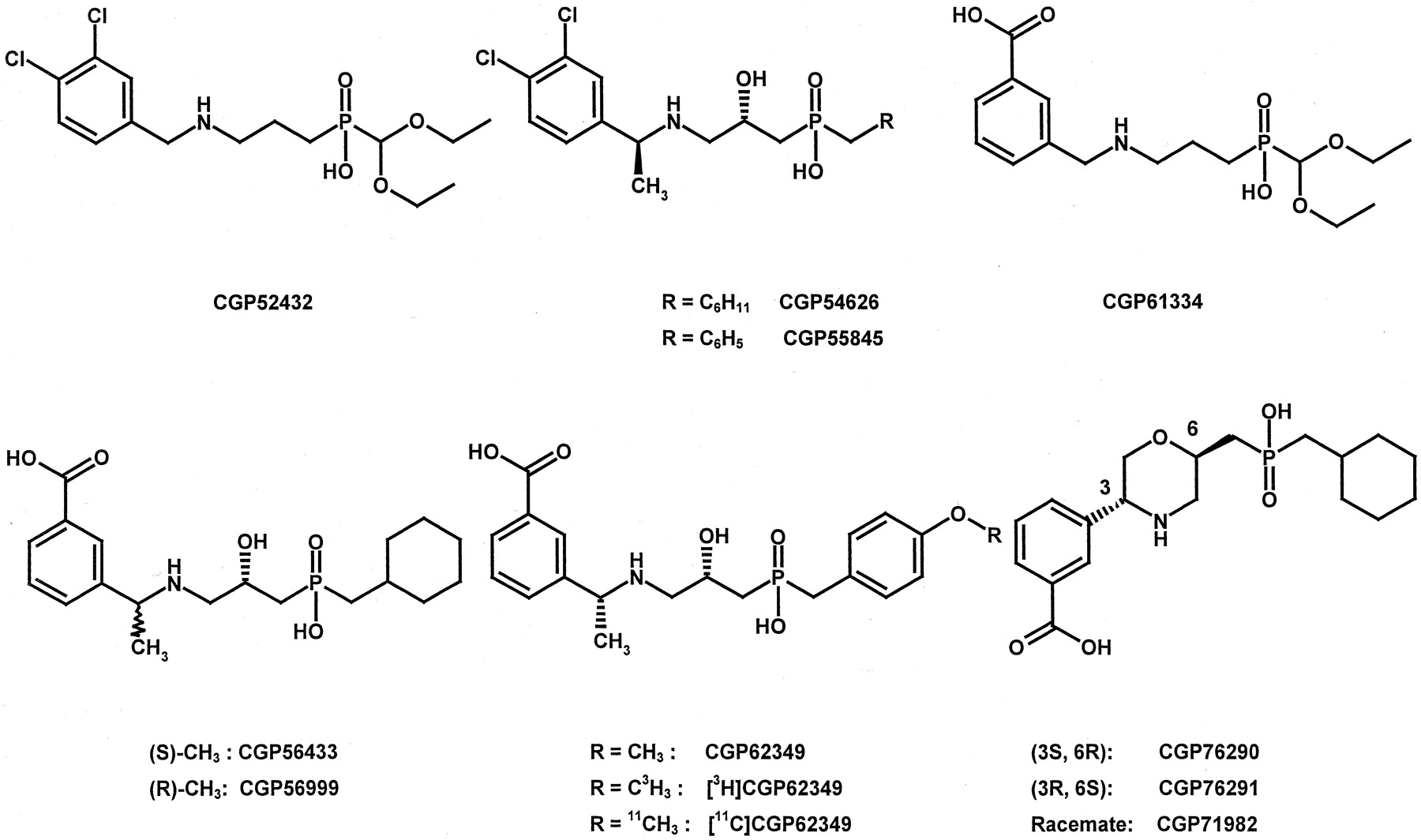
Subsequent discoveries of antagonists were derived largely by a group at Novartis in Basel, Switzerland (Froestl and Mickel, 1997). They developed the first GABAB receptor antagonist able to cross the blood-brain barrier, CGP35348, and the first orally active agents, CGP36742 (Olpe et al., 1990, 1993a) and CGP51176 (Froestl et al., 1995b). However, these compounds, and others in this chemical series, have affinities for the GABABreceptor in the same range as 2-hydroxysaclofen (IC50 values: 27, 38, and 6 μM, respectively). The same is true for SCH 50911 (IC50 = 3 μM; Fig. 2), a chemically distinct agent that is effective following systemic administration but which has a relatively low affinity for the receptor (Bolser et al., 1995; Frydenvang et al., 1997). The most crucial breakthrough in the discovery of antagonists came with the development of compounds with affinities about 10,000 times higher than previous antagonists. This major advance stemmed from the attachment of 3,4-dichlorobenzyl or 3-carboxybenzyl substituents to the existing molecules. This produced a profusion of compounds with affinities in the low nanomolar range (Froestl et al., 1996; Froestl and Mickel, 1997). Numerous investigations have been carried out with CGP52432 (IC50 = 55 nM), CGP54626A (IC50 = 4 nM), CGP55845A (IC50 = 6 nM), CGP56433A (IC50 = 80 nM), CGP56999A (IC50 = 2 nM), CGP61334 (IC50 = 36 nM), and CGP62349 (IC50 = 2 nM; Fig.3). Several compounds are also available as radioligands, such as [3H]CGP54626 (Bittiger et al., 1992; Green et al., 2000), [3H]CGP56999, and [3H]CGP62349 (Bittiger et al., 1996a;Ambardekar et al., 1999; Keir et al., 1999; Sloviter et al., 1999;Billinton et al., 2000). From the latter compound a radioligand containing the positron-emitting isotope 11C was prepared as a potential positron emission tomography ligand (Todde et al., 2000).

Structures of high-affinity GABABreceptor antagonists.
Introducing the phosphinic acid moiety into the Schering compound SCH 50911 led to a new class of very potent GABABreceptor antagonists, such as CGP76290A (Ong et al., 1998a; IC50 = 2 nM, enantiomer CGP76291: IC50 = 69 nM, racemate CGP71982, IC50 = 8 nM; Fig. 3).
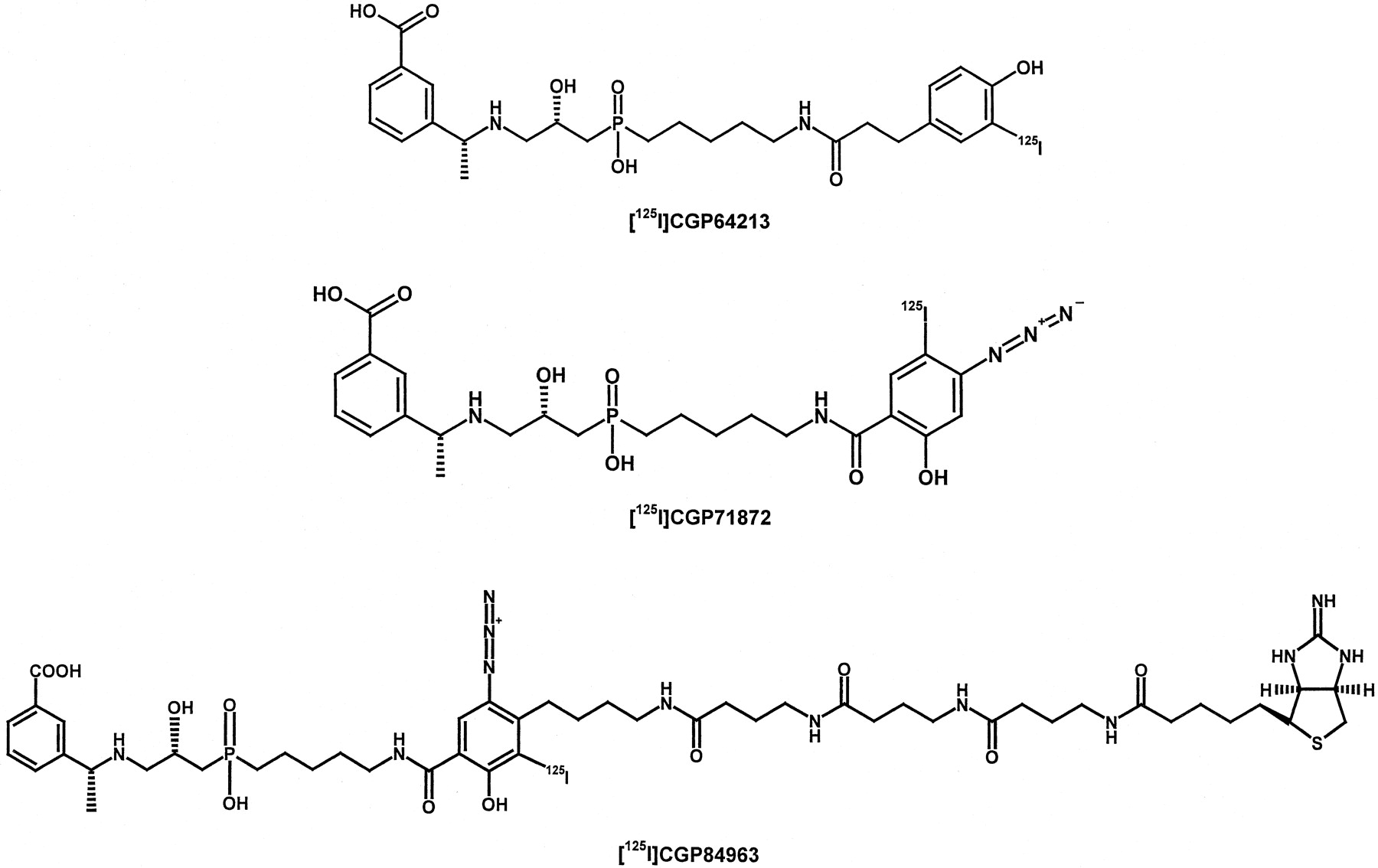
Finally, two iodinated high-affinity antagonists, i.e., [125I]CGP64213 (IC50 = 1.6 nM, i.e., inhibition of binding of [125I]CGP64213 to GABABreceptors on rat cerebral cortex membranes) and [125I]CGP71872 (IC50 = 2.4 nM), a photoaffinity ligand, both with high specific radioactivities of >2000 Ci/mmol were developed, which were used for the elucidation of the structure of GABAB1(Kaupmann et al., 1997; Belley et al., 1999; Calon et al., 2000;Froestl et al., 2001; Fig. 4). The ligand [125I]CGP84963 (IC50 = 6 nM, i.e., inhibition of binding of [125I]CGP64213 to GABABreceptors on rat cerebral cortex membranes; Fig. 4) combines in one molecule a GABAB receptor-binding part, an azidosalicylic acid as a photoaffinity moiety separated by a spacer of three GABA molecules from 2-iminobiotin, which binds to avidin in a reversible, pH dependent fashion. This compound was prepared to facilitate isolation and purification of the extracellular N-terminal GABAB1 receptor fragment for crystallization and X-ray studies of the GABAB1 binding site (Froestl et al., 1999).

Structures of high-affinity125I-labeled GABAB receptor antagonist.
Although GABAB receptor antagonists have yet to be studied in humans, results of animal studies suggest that they may have clinical utility. Thus, GABAB receptor antagonists suppress absence seizures in a variety of animal models (Marescaux et al., 1992). When administered either systemically or directly into the thalamus, GABAB receptor antagonists prevent spike and wave discharges in the electroencephalograms of genetic absence rats. Similar results are obtained with the lethargic mouse and in rats injected with γ-hydroxybutyrate (GHB), which produces absence-like seizure activity (Hosford et al., 1992; Snead, 1992). In all cases, GABAB receptor antagonists dose dependently reduce seizure activity. In the genetic absence rats, the spontaneous seizures are blocked by bilateral administration of pertussis toxin, supporting the involvement of Gi/Go coupling in generating and maintaining the seizures (Bowery et al., 1999). These results suggest that GABAB receptor activation may contribute to the absence syndrome, possibly through Ca2+ spike generation in the thalamus (Crunelli and Leresche, 1991; Charpier et al., 1999).
At high doses, GABAB receptor antagonists induce convulsions in rats (Vergnes et al., 1997). Although the mechanism(s) underlying this action is unknown, the response is blocked by GABAB receptor agonists. Importantly, not all GABAB receptor antagonists cause seizures. For example, SCH 50911 fails to cause convulsions at doses 10- to 100-fold higher than those that completely block seizures in the genetic absence rat (Richards and Bowery, 1996).
The GHB-induced absence-like seizures in rats appear due, at least in part, to a weak partial agonist action at GABABreceptors (Bernasconi et al., 1999; Lingenhoehl et al., 1999). However, GHB also acts through sites distinct from GABABreceptors (Snead, 2000).
Several GABAB receptor antagonists have been found to improve cognitive performance in a variety of animal paradigms, such as the low-affinity compounds, CGP35348 (Bianchi and Panerai, 1993; Castellano et al., 1993; Saha et al., 1993;Stäubli et al., 1999) and CGP36742 (Carletti et al., 1993;Mondadori et al., 1993, 1994, 1996a,b; Nakagawa and Takashima, 1997; Yu et al., 1997; Bonanno et al., 1999; Genkova-Papazova et al., 2000; Farr et al., 2000; Pittaluga et al., 2001), or the high-affinity compounds CGP55845A, CGP56433A, CGP61334, CGP62349, and CGP71872 (Getova et al., 1997; Getova and Bowery, 1998). Olpe et al. (1993b) observed a very pronounced facilitation of long-term potentiation in vivo with doses of 100 mg/kg i.v. CGP35348 on eliciting long-term potentiation by nonprimed tetanic stimulation in the CA1 region of the hippocampus of rats. Brucato et al. (1996) reported a suppression of long-term potentiation with CGP46381 using θ-like stimulus trains to the dentate gyrus. However, the latter GABAB receptor antagonist did not show effects on working memory in the radial maze in rats (Brucato et al., 1996) nor did it improve learning and memory in mice in a step-down passive avoidance paradigm (C. Mondadori and W. Froestl, unpublished observations).
Perhaps not surprisingly, therefore, GABABagonists impair learning behavior in animal models (Soubrie et al., 1976; Swartzwelder et al., 1987; Nabeshima et al., 1988a,b; Sidel et al., 1988; Castellano et al., 1989; Sharma and Kulkarni, 1990 and 1993;Castellano and McGaugh, 1991; Saha et al., 1993; DeSousa et al., 1994;Stackman and Walsh, 1994; Nakagawa et al., 1995; McNamara and Skelton, 1996; Tang and Hasselmo, 1996; Arolfo et al., 1998). This induced amnesia appears to be mediated via G protein-linked receptors because the impairment produced by baclofen in mice can be blocked by pertussis toxin administered intracerebroventricularly (Galeotti et al., 1998). Baclofen has occasionally produced memory deficits in patients (Sandyk and Gillman, 1985).
GABAB receptor antagonists improve cognitive performance in a variety of animal models (Carletti et al., 1993;Mondadori et al., 1993; Brucato et al., 1996; Getova et al., 1997;Nakagawa and Takashima, 1997; Yu et al., 1997; Getova and Bowery, 1998;Stäubli et al., 1999; Genkova-Papazova et al., 2000; Farr et al., 2000). Conversely, GABAB receptor agonists impair learning, an action that is blocked by pertussis toxin, supporting the involvement of Gi/Go in the action of these agents (Nakagawa et al., 1995; McNamara and Skelton, 1996; Tang and Hasselmo, 1996; Arolfo et al., 1998; Galeotti et al., 1998). In studies with mice lacking the GABAB1receptor subunit, a clear impairment of passive avoidance performance was observed, which was related to gene dosage (Schuler et al., 2001). These passive avoidance deficits are a reflection of impaired memory processes further linking GABAB receptors to memory performance. Reports suggest that both GABAB receptor antagonists and agonists are neuroprotective. Although baclofen is neuroprotective in a gerbil cerebral ischemia model, very high doses are required (Lal et al., 1995). Moreover, baclofen attenuates the neurotoxic effect of quinolinic acid on CA1 cells in rat hippocampus (Beskid et al., 1999). In contrast, studies with mouse cultured striatal neurons reveal that GABAB receptor activation enhances the neurotoxic effects of N-methyl-d-aspartate, reinforcing the concept that GABAB antagonists are more likely to be neuroprotective than agonists (Lafon-Cazal et al., 1999). In support of this, low doses of GABAB receptor antagonists increase levels of nerve growth factor and brain-derived neurotrophic factor in rat brain hippocampus, neocortex, and spinal cord, which could attenuate neurodegenerative processes (Heese et al., 2000).
The potential significance GABAB receptor mechanisms in depression was first suggested by Lloyd and colleagues (Pilc and Lloyd, 1984; Lloyd et al., 1985, 1989), but this was challenged by other groups. More recently, however, further suggestions that GABAB antagonists, e.g. CGP36742, are effective in animal models of depression have emerged (Nakagawa et al., 1999). Clear antidepressant effects were seen after 4 weeks of oral treatment with CGP51176 in the chronic mild stress model (Bittiger et al., 1996b). This might be supported in due course by the observations of Heese et al. (2000) who showed that GABABantagonists produce a rapid increase in nerve growth factor and brain-derived neurotrophic factor levels. Interestingly, antidepressants have been shown to produce the same increase in those growth factors but only after 2 to 3 weeks (Nibuya et al., 1995; Duman et al., 1997). Could there be a link between these phenomena?
VII. Conclusions
The G protein-coupled GABAB receptor was first described over 20 years ago but only recently has the site been cloned and with this has come the identification of its unique heterodimeric structure. Even though much is known about the formation and characteristics of this receptor, many important questions remain. Thus, it is crucial to determine whether GABABreceptor subtypes exist that can be exploited pharmacologically, to determine whether other proteins can link with GABAB1 to form a functional receptor, to establish whether GABAB receptor subunits serve other functions in the cell, and to assess the clinical value of GABAB receptor agonists and antagonists. Given the pace of discovery in this field, answers to these questions will be forthcoming. These results will not only have significant implications with regard to understanding the GABAB receptor system in particular but may allow novel drugs acting at this receptor to be developed.
It is evident that the presence of two distinct proteins (or a protein and an essential accessory protein), coupled to G proteins, forming a receptor for GABA, poses certain problems for nomenclature. The subcommittee considers that there is little evidence for distinct functional types of the receptor, but this is a rare, and usually short-lived, situation in pharmacology. The present proposition is to continue to call the receptor the GABAB receptor, and when distinct splice variants are studied, i.e. GABAB1(a)/2(a), using the NC-IUPHAR designation for splice variants. This nomenclature is provisional and may be changed when there is evidence of distinct functional GABAB receptor types or if NC-IUPHAR issues general guidelines for ligand-gated ion channels, which modify the GABAA/B terminology.
Footnotes
Address correspondence to: Dr. Norman Bowery, University of Birmingham Medical School, Edgbaston, Birmingham, B15 2TT UK. E-mail:n.g.bowery{at}bham.ac.uk
Abbreviations
- GABAB
- γ-aminobutyric acidB
- 3-APPA
- 3-aminopropyl-phosphinic acid
- 3-APMPA
- methyl homolog of 3-APPA
- GHB
- γ-hydroxybutyrate
- NC-IUPHAR
- International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification
- CHO
- Chinese hamster ovary
- U.S. Government
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- I. Introduction
- II. γ-Aminobutyric AcidB Receptor Structure
- III. γ-Aminobutyric AcidB Receptor Effector Mechanisms
- IV. γ-Aminobutyric AcidB Receptor Subtypes
- V. γ-Aminobutyric AcidB Receptor Distribution
- VI. γ-Aminobutyric AcidB Receptor-Mediated Responses
- VII. Conclusions
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters