


I. Introduction
Based on structural and evolutionary criteria, neuropeptide Y (NPY)b, peptide YY (PYY) and pancreatic polypetide (PP) are closely related polypeptides (Larhammar, 1996a). They are composed of 36 amino acids each and share considerable amino acid homology, amidated C-terminal ends, and the presence of a large number of tyrosine residues including both ends of the molecule (fig.1). The tertiary structure of turkey PP has been elucidated by crystallography (Larhammar, 1996a). This tertiary structure appears to be characteristic for the whole family of peptides and has been termed the “PP-fold.” The PP-fold is U-shaped and consists of an extended polyproline helix and an α helix connected by a β turn.

Alignment of human neuropeptide Y (NPY), peptide YY (PYY) and pancreatic polypeptide (PP). Amino acid identities between peptides are indicated by a *.
NPY, PYY, and PP can be found in different locations (Sundler et al., 1993). Thus, NPY is primarily synthesized and released by neurons, which in the peripheral nervous system are predominantly sympathetic neurons. PYY is predominantly synthesized and released by intestinal endocrine cells and can also coexist with glucagon in pancreatic acini and enteroglucagon in endocrine cells of the lower bowel. PP is mainly found in pancreatic cells distinct from those storing insulin, glucagon, or somatostatin. However, in some cases, other cell types can also express NPY, PYY, and PP. While NPY acts as a neurotransmitter, PYY and PP act as hormones. Physiological effects attributed to NPY, PYY, and PP include stimulation of food intake and inhibition of anxiety in the central nervous system (CNS) (Colmers and Bleakman, 1994; Wettstein et al., 1995), presynaptic inhibition of neurotransmitter release in the CNS and the periphery (Lundberg, 1996), vasoconstriction (Michel and Rascher, 1995), inhibition of insulin release (Wang et al., 1994), regulation of gut motility (Sheikh, 1991), and gastrointestinal and renal epithelial secretion (Playford and Cox, 1996).
Although PP was discovered first and NPY last, evolutionary analysis shows that PP is actually the newest member of the family (Larhammar, 1996a). Both NPY and PYY are found in representatives of all major vertebrate groups. NPY is the most highly conserved; even the sequence in Torpedo marmorata is identical with mammalian NPY in 33 of 36 positions. PYY and NPY presumably evolved by duplication from a common ancestral gene in an early vertebrate ancestor. The corresponding genes are located on different chromosomes. The PP gene probably arose by duplication of the PYY gene; both genes are located close to one another in the same chromosomal segment (Larhammar, 1996a). Based on these evolutionary considerations, it is recommended that the family is denoted by the NPY family.
The members of the NPY family act upon the same family of receptors (see below). Therefore, it is recommended that the receptors for NPY, PYY, and PP are classified together as NPY receptors. Because the members of the NPY family contain many tyrosine residues, which are abbreviated by the letter Y in the single letter amino acid code, the NPY receptors are designated by a capital Y. The various NPY receptors within the family are designated by subscript numbers, e.g. Y1, Y2, etc. Although nonmammalian NPY receptor types have been identified as distinct from all those described below (see Section V.), they are not included in this classification until mammalian homologs have been identified.
II. Signal Transduction of Neuropeptide Y Receptors
All known NPY receptors belong to the large superfamily of G-protein-coupled, heptahelical receptors. They appear to use similar signal transduction pathways, and no clear and consistent alignment of a specific receptor type with a distinct transduction pathway has been identified. In almost every cell type studied, NPY receptors act via pertussis toxin-sensitive G-proteins, i.e., members of the Gi and Go family. Although responses to NPY have been found to be pertussis toxin-insensitive in a small number of cases, mostly involving presynaptic receptors (Colmers and Pittman, 1989; Foucart and Majewski, 1989; Millar et al., 1991;Hexum et al., 1994; Lemos and Takeda, 1995), it remains unclear whether this indeed relates to a distinct signaling mechanism or to the failure of pertussis toxin to fully inactivate its substrates in some cell types. The typical signaling responses of NPY receptors are similar to those of other Gi/Go-coupled receptors. Thus, inhibition of adenylyl cyclase is found in almost every tissue and cell type investigated; this is also observed with all cloned NPY receptor subtypes upon heterologous expression (Gerald et al., 1996;Weinberg et al., 1996). However, adenylyl cyclase inhibition probably cannot explain many of the functional responses after stimulation of NPY receptors. Additional signaling responses that are restricted to certain cell types include inhibition of Ca2+channels, e.g. in neurons (Ewald et al., 1988), and activation and inhibition of K+ channels, e.g. in cardiomyocytes (Millar et al., 1991) and vascular smooth muscle cells (Xiong and Cheung, 1995), respectively. Based on experiments with Ca2+ entry blockers, it has been postulated that NPY stimulates Ca2+ channels in the vasculature (Michel and Rascher, 1995). In some cell types, members of the NPY family can mobilize Ca2+ from intracellular stores; although this appears to involve inositol phosphates in some cells (Perney and Miller, 1989), inositol phosphate-independent Ca2+ mobilization has been postulated in other cell types (Motulsky and Michel, 1988). A sensitivity of certain responses to NPY to the cyclooxygenase inhibitor, indomethacin, indicates possible activation of a phospholipase A2 by NPY receptors (Martin and Patterson, 1989), but this has yet to be demonstrated definitively. Activation of a phospholipase D or of a tyrosine kinase, which can occur with some Gi/Go-coupled receptors, also has not been clearly demonstrated to date. Thus, in general, Y receptors demonstrate a preferential coupling to pertussis toxin-sensitive G-proteins, i.e., the Gi and Go family, which is followed by the responses typically under the control of these G-proteins (Limbird, 1988).
III. Historical Aspects of Neuropeptide Y Receptor Subdivision
Historically, the subdivision of NPY receptors comes from the observation that C-terminal fragments of NPY or PYY, e.g. NPY13–36, can mimic some responses to NPY, e.g. prejunctional inhibition of twitch responses in the rat vas deferens, but not others, e.g. vasoconstriction in guinea pig iliac vein (Wahlestedt et al., 1986). Thus, it has been proposed that receptors that are only activated by the holopeptides are designated Y1, while those that are activated by the holopeptides and the C-terminal fragment are designated Y2. Although short C-terminal fragments, e.g. PYY13–36 and NPY18–36, have primarily been generated synthetically, it has now been recognized that some long C-terminal fragments of PYY and NPY may exist endogenously, i.e., PYY3–36 (Eberlein et al., 1989) and NPY3–36 (Grandt et al., 1996). These do not appear to be nonspecific degradation products of their parent peptides but rather are formed by the dipeptidyl peptidase IV, also known as CD26 (Mentlein et al., 1993). The occurrence of PYY3–36 in plasma appears to be regulated by mechanisms that may be distinct from those regulating release of PYY because physiological stimuli such as food intake may differentially alter the plasma levels of both peptides in humans (Grandt et al., 1994b). Although the endogenous formation of PYY3–36 and NPY3–36 does not give them a specific importance for receptor classification, it may be physiologically relevant because NPY3–36 and PYY3–36 are highly selective for Y2 over Y1 receptors (Grandt et al., 1992, 1996). Numerous other C-terminal fragments have been synthesized but will not be discussed further because they have been investigated much less frequently than the 3–36, 13–36, and 18–36 C-terminal fragments.
The principal subdivision of NPY receptors into the Y1 and Y2 subtypes has survived the test of time. Meanwhile it is supported by three additional sets of data. First, analogs of NPY and PYY have been synthesized that contain a proline residue in position 34 of the molecule, i.e., [Leu31, Pro34]NPY (Fuhlendorff et al., 1990) or [Pro34]PYY (Grandt et al., 1994b), and are much more potent at Y1 than Y2receptors. The introduction of an additional [Leu31]substitution into [Pro34]substituted analogs does not appear to be important for Y1 selectivity (Grandt et al., 1994a; Wieland et al., 1995a). Second, nonpeptide antagonists have been synthesized (fig. 2), which competitively block Y1 but not Y2receptors, e.g. BIBP 3226 (Wieland et al., 1995b; Doods et al., 1996;Gerald et al., 1996), SR 120819A (Serradeil-Le Gal et al., 1995), or GR 231118 (also known as GW1229 or 1229U91) (Daniels et al., 1995; Hedge et al., 1995). Third, and most important, complement deoxyribonucleic acids (cDNAs) and genes have been cloned that encode Y1 and Y2 receptors from several species (see below).

Structures of neuropeptide Y receptor antagonists.
IV. Present Definition of Neuropeptide Y Receptors
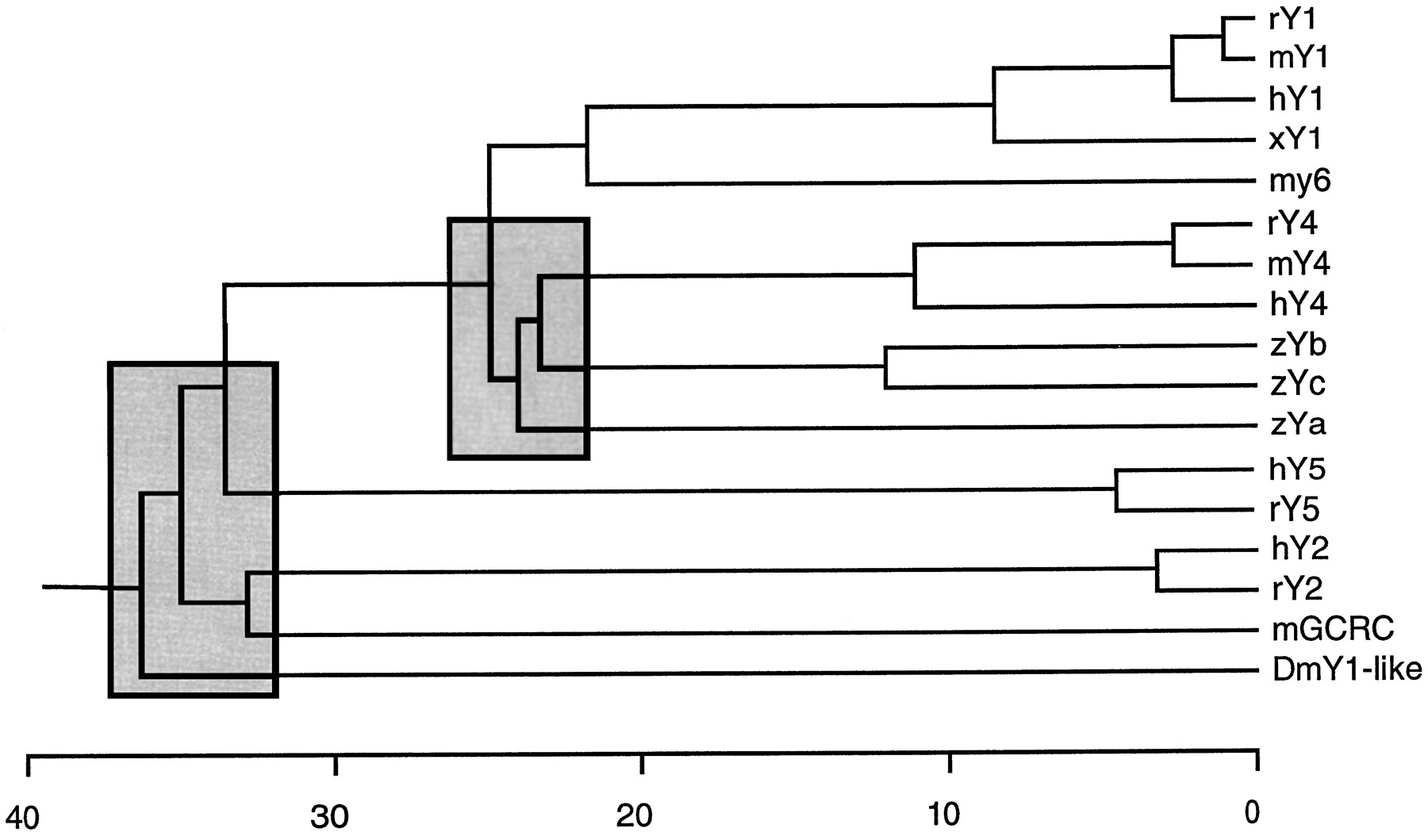
Five distinct NPY receptors have been cloned (fig.3). Sequence comparisons show that receptors Y1, Y4, and y6 are more closely related to each other than to the receptors Y2 and Y5(fig. 4; Larhammar, 1996b). This is apparent not only from sequence identity but also from other features, such as cysteines, believed to form disulfide bonds and the size of the third cytoplasmic loop, which is large in Y5. The receptors Y2 and Y5 are equally distantly related to one another as to the Y1/Y4/y6group. In fact the Y1/Y4/y6group, the Y2, and the Y5receptor are more distantly related to one another than any other G-protein-coupled receptors that bind the same endogenous ligand, despite the fact that Y1, Y2, and Y5 each bind two distinct endogenous ligands, namely NPY and PYY. Two explanations are possible for the great differences between Y1, Y2, and Y5. The first possibility is that they evolve rapidly and, hence, have accumulated all these differences in a short period of time, but this is unlikely because Y1 is highly conserved between the frogXenopus laevis and mammals. The second possibility is that the three genes arose very long ago and, hence, have had a long time to diverge from one another. Although these data could indicate the existence of three subfamilies of NPY receptors, which contain the Y1/Y4/y6, the Y2, and the Y5receptor, respectively, the overall homologies between the NPY receptor types are small and not all receptor types have been cloned yet. Moreover, based on pharmacological recognition profiles, the receptors Y1 and Y5 are more similar to one another than to the receptor Y4 (see Sections V.A., V.D., and V.F.). Therefore, the present evidence is insufficient to recommend a formal division of NPY receptors into subfamilies.

Alignment of predicted human (h) Y1, Y2, Y4, and Y5 and mouse (m)Y6 receptor amino acid sequence. The putative transmembrane (TM) domains are indicated by boxes. Amino acid identities are indicated by dots.

Distance tree for receptors of the NPY family. Branch lengths correspond to sequence divergence calculated from an alignment starting at the cysteine preceding the first transmembrane-spanning domain (see fig. 3) or the equivalent position and extending through the termination codon. The tree was calculated with the neighbor joining method with the Lasergene DNASTAR Megalignn software. The branching order within the shaded boxes is uncertain and may vary depending on which sequences are included in the analysis. The human neurokinin 3 receptor was used as outgroup. Species abbreviations are h for humans, m for mouse, r for rat, x for Xenopus laevis, z for zebrafish, and Dm for Drosophila melanogaster. mGCRC is a murine orphan receptor. Figure provided by Dan Larhammar (Uppsala, Sweden).
Among the cloned receptors, the Y1, Y2, Y4, and Y5 receptors represent fully defined subtypes, while no functional correlate of the cloned y6receptor has been reported to date. The subtypes Y1, Y2, and Y5 preferentially bind NPY and PYY, whereas the subtype Y4 preferentially binds PP; the pharmacological profile of the Y6 receptor is controversial (see below). The pharmacologically defined and cloned Y1 and Y2 receptors correspond to each other. Although the cloned Y1and Y5 receptors can clearly be distinguished by long C-terminal fragments of NPY and PYY (e.g. PYY3–36) and by the antagonist BIBP 3226, their pharmacological profile with regard to NPY, PYY, [Pro34]substituted analogs and short C-terminal fragments (e.g. NPY13–36) is similar (see below). Thus, it cannot be excluded that some physiological responses, which have been attributed to Y1 receptors based on incomplete pharmacological characterization, are actually Y5–mediated.
V. Characteristics of Receptor Subtypes (table1)
A. Y1 Receptors
In 1990, the cloning of a cDNA from rats was described (Eva et al., 1990) that was later found to encode a Y1receptor (Krause et al., 1992; Petitto et al., 1994). Thereafter, species homologs from mice (Eva et al., 1992), humans (Larhammar et al., 1992) and Xenopus laevis (Blomqvist et al., 1995) have been identified. Moreover, in humans and mice, the genomic organization of the Y1 subtype gene has been determined and the gene has been located on human chromosome 4q(31.3–32) (Eva et al., 1992; Herzog et al., 1993a). Three splice variants have been identified in the 5′ region of the human Y1 receptor that yield multiple promoters with tissue-specific expression patterns (Ball et al., 1995). Two splice variants of the murine Y1 receptor have been described; although both variants bind NPY, the form with a shortened seventh transmembrane-spanning region and a lacking C-terminal tail does not appear to couple to signal transduction as efficiently as the full length form (Nakamura et al., 1995). An order of potency of NPY ≥ PYY ≥ [Pro34]substituted analog ≫ C-terminal fragment > PP is characteristic for the Y1 subtype (Krause et al., 1992; Larhammar et al., 1992; Wieland et al., 1995a; Gerald et al., 1996). C-terminal fragments may act as partial agonists at Y1receptors and in some cell lines even as antagonists (Michel et al., 1990); whether such partial antagonism also occurs with intact tissues or in vivo remains to be determined. The nonpeptide BIBP 3226 (fig. 2) is a high potency antagonist at the Y1 receptor (Ki or KB 1 to 10 nm) although its affinity at other NPY receptor types exceeds 10 μm (Wieland et al., 1995b; Gerald et al., 1996). Other antagonists of Y1 receptors, e.g. SR 120819A or GR 231118 (fig. 2), have also been described, but their selectivity, in particular toward Y5 and y6 receptors, is less well-established (Daniels et al., 1995; Hedge et al., 1995; Serradeil-Le Gal et al., 1995). GR 231118 has been shown to have a high affinity for Y4 receptors (Gehlert et al., 1996a), but whether it is an antagonist at these receptors is yet to be determined. Messenger ribonucleic acid (mRNA) for the Y1receptor has been detected in a variety of human, rat, and murine tissues including brain, heart, kidney, and gastrointestinal tract (Larsen et al., 1993; Wharton et al., 1993; Nakamura et al., 1995). Prototypical responses for the Y1 subtype include vasoconstriction in most vascular beds, e.g. in the isolated perfused kidney, but vasoconstriction via other NPY receptor subtypes has also been observed (Michel and Rascher, 1995). Based on antisense oligodeoxynucleotide studies, Y1 receptors also appear to be involved in the anxiolytic effects of NPY (Wahlestedt et al., 1993). SK-N-MC (Fuhlendorff et al., 1990; Wieland et al., 1995a) and human erythroleukemia (HEL) cells (Michel et al., 1990; Feth et al., 1992) are human cell lines containing apparently homogeneous populations of Y1 receptors.
Characterization of neuropeptide Y receptor subtypes
B. Y2 Receptors
A cDNA for Y2 receptors was first cloned from human SMS-KAN cells (Rose et al., 1995) and subsequently from human brain cDNA libraries (Gehlert et al., 1996b; Gerald et al., 1995) and the human neuroblastoma cell line KAN-TX (Rimland et al., 1996). An order of potency of NPY ≈ PYY ≥ C-terminal fragment ≫ [Pro34]substituted analog >PP is characteristic for the Y2 subtype (Rose et al., 1995; Gehlert et al., 1996b; Gerald et al., 1996). Messenger RNA for the Y2 receptor has been detected in various parts of the CNS, although apparently low levels of Y2 mRNA were found in human peripheral tissues (Rose et al., 1995; Gehlert et al., 1996b; Zhang et al., 1997). The prototypical response for the Y2 subtype is the presynaptic inhibition of neurotransmitter release, e.g. of noradrenaline release in the prostatic half of the rat vas deferens (Wahlestedt et al., 1986), but postjunctional Y2receptors also exist in a variety of tissues. SMS-KAN cells are a human cell line containing an apparently homogeneous population of Y2 receptors, and the rabbit kidney is also a rich source for this receptor type (Wieland et al., 1995a).
C. Putative Y3 Receptors
It has been reported that PYY is considerably less active than NPY, i.e., is less than one-tenth as potent as NPY in several model systems including rat CNS (Grundemar et al., 1991), rat colon (Dumont et al., 1994), rat lung (Hirabayashi et al., 1996), rat and bovine adrenals (Bernet et al., 1994; Nörenberg et al., 1995), and in the adrenal-derived PC12 cell line (McCullough and Westfall, 1995). This site of action of NPY has been referred to as a “Y3 receptor.” However, at present, the evidence for the existence of such a site is circumstantial. It has not been cloned and no specific agonists or antagonists have been described. An early report on the cloning of a rat NPY receptor subtype with greater potency for NPY than for PYY (Rimland et al., 1991) was later shown to result from an artifact (Herzog et al., 1993b; Jazin et al., 1993). Therefore, the present evidence is not sufficient to grant the Y3 site receptor status. Because this designation has already been used by various investigators, we propose to leave the number three spot in the NPY receptor series vacant for the time being and refer to binding sites and responses where NPY is considerably (at least ten-fold) more potent than PYY as “putative Y3” receptors.
D. Y4 Receptors
The gene for Y4 receptors was initially cloned from a human genomic library and the receptor derived from this clone was originally designated “PP1” (Lundell et al., 1995). A rat homolog (Gerald et al., 1996; Lundell et al., 1996; Yan et al., 1996) and a murine homolog (Gregor et al., 1996a) have also been cloned. The principal feature of the Y4 receptor is its very high affinity (<100 pm) for PP of the same species. PP homologs from other species may have 50- to 100-fold lower affinities, e.g. rat PP at the human Y4 receptor (Gehlert et al., 1996a), although this has not been found in all cases (Gerald et al., 1996; Gregor et al., 1996a). Human PP appears to have very high affinity (<100 pm) for human (Bard et al., 1995; Lundell et al., 1995; Gehlert et al., 1996a), rat (Gerald et al., 1996), and murine Y4 receptors (Gregor et al., 1996a). PYY, NPY, and [Pro34]substituted analogs were reported to have affinities in the low nanomolar range for the human Y4 receptor in most (Bard et al., 1995; Lundell et al., 1995; Gehlert et al., 1996a), but not all, studies (Yan et al., 1996). In contrast, except for [Leu31, Pro34]NPY, all related peptides were reported to have negligible affinity for the rat Y4 receptor (Gerald et al., 1996; Lundell et al., 1996; Yan et al., 1996). Among the NPY antagonists, BIBP 3226 has very low affinity for the human Y4 receptor (>10 μm), although GR 231118 has an affinity in the picomolar range (Gehlert et al., 1996a). These studies indicate that PP is the primary endogenous ligand for the Y4 receptor, but activation of the human, but not rat, homolog by circulating PYY can also be envisioned when these conflicting data indeed represent species’ differences. Human Y4 mRNA is mainly expressed in the colon, small intestine, and prostate, although other peripheral tissues appear to lack it, and various CNS regions display low expression levels (Lundell et al., 1995). Rat Y4 mRNA was mainly detected in testis and lung with much weaker signals in colon (Lundell et al., 1996).
E. Y5 Receptors
Very recently, the cloning of additional cDNAs has been reported from rats and humans that encode the proteins of 456 amino acids, according to one report (Gerald et al., 1996) and 445 amino acids, according to another report (Hu et al., 1996). The corresponding gene appears to reside on human chromosome 4q, i.e., the same location as the human Y1 receptor gene but, apparently, in an opposite orientation (Gerald et al., 1996; Hu et al., 1996). Messenger RNA for that receptor was detected by Northern blotting and in situ hybridization in several rat brain areas, including those believed to be important for the regulation of food intake, as well as in testis (Gerald et al., 1996; Hu et al., 1996). Upon expression in 293 cells, the cloned subtype couples to inhibition of cAMP accumulation (Gerald et al., 1996). For this response, an order of potency of NPY ≥ PYY ≈ [Pro34]substituted analog ≈ NPY2–36 ≈ PYY3–36 ≫ NPY13–36 has been observed; rat PP had very low potency at the rat and human Y5 receptor, although human and bovine PP had affinities similar to those of NPY and PYY (Gerald et al., 1996). A similar order of potency was observed in binding studies with rat and human Y5 receptors expressed in COS-7 cells (Hu et al., 1996). This is similar to the order of potency observed to mimic NPY-stimulated enhancements of food intake (Gerald et al., 1996) or enhancements of renal sodium excretion (Bischoff et al., 1997). This clone may, therefore, represent a subtype previously referred to as “Y1-like” or “food intake” receptor.
F. y6 Receptors
An additional receptor subtype has been cloned from mouse genomic DNA; the intronless gene encodes a 371 amino acid protein and, at the time, was designated “Y5” (Weinberg et al., 1996). Homologs have also been cloned from rabbit, monkey, and human libraries and designated either “Y2B” (Matsumoto et al., 1996) or “PP2” (Gregor et al., 1996b). In agreement with the authors of the respective publications, the use of these designations is no longer encouraged, and, instead, the designation “y6” is recommended. Fluorescent in situ hybridization has localized the y6 gene to the human chromosome 5 in the 5q31 region (Gregor et al., 1996b). Messenger RNA for this subtype is found in areas of murine brain by in situ hybridization (Weinberg et al., 1996) but not in mouse tissues as assessed by Northern blotting (Gregor et al., 1996b). Similarly, mRNA for the y6receptor was not detected in rabbit tissues by Northern blotting but readily seen using RT-PCR in rabbit brain areas including hypothalamus and hippocampus and in the small intestine and adrenals (Matsumoto et al., 1996). In contrast, Northern blotting has detected mRNA for the y6 receptor in human tissues, including heart and skeletal muscle (Gregor et al., 1996b; Matsumoto et al., 1996). The monkey and human sequences differ from those in mice and rabbits by a frame shift mutation located in the putative third intracellular loop of the receptor that results in a stop codon and a predicted truncated protein of only 290 amino acids (Gregor et al., 1996b; Matsumoto et al., 1996). Although expression of the mouse and rabbit clones results in functional proteins (Gregor et al., 1996b; Matsumoto et al., 1996;Weinberg et al., 1996), expression of the monkey or human proteins has not been successful under a variety of conditions despite the presence of its mRNA in a variety of human tissues (Gregor et al., 1996b;Matsumoto et al., 1996). Thus, the y6 gene of primates may have become nonfunctional during evolution. The pharmacological profile of the expressed y6receptor has remained controversial: Although one study has reported an order of potency of NPY ≈ PYY ≈ [Pro34]substituted analog >NPY13–36 ≫ PP (>1000 nm) for the murine receptor (Weinberg et al., 1996), another study has described an order of potency of PP ≫ [Pro34]substituted analog ≫ PYY ≈ NPY >C-terminal fragment (NPY3–36, NPY13–36, each >1000 nm) at the murine receptor (Gregor et al., 1996b); expression of the rabbit clone exhibits an order of potency of PYY≈ NPY13–36≈ NPY >[Pro34]substituted analog >PP (Matsumoto et al., 1996). A physiological correlate of the cloned y6 receptor has not yet been described.
G. Additional Sites
Several reports have used the term “PYY-preferring receptor. ” In most cases, it has been applied to describe a receptor where PYY was three to five times more potent than NPY. However, a PYY preference of this small magnitude is observed for many Y1 receptor-mediated responses and may be a general feature of this subtype rather than the hallmark of an additional subtype. Thus, convincing evidence for the existence of such a subtype is lacking. We recommend that the term “PYY-preferring receptor” is not used unless a potency difference of at least twenty-fold between PYY and NPY is observed.
A receptor belonging to the NPY receptor family has also been cloned from Drosophila melanogaster (Li et al., 1992). Moreover, multiple NPY receptor types have been cloned from zebrafish (Danio rerio; Larhammar et al., unpublished). Although the zebrafish subtypes appear to be quite distinct from all known mammalian subtypes with regard to primary sequence data, it remains to be determined whether mammalian homologs exist. Until the identification of such homologs, it is not recommended to extend the Y receptor designation to the zebrafish clones but rather propose to specifically identify them as zebrafish NPY receptors.
In addition to the above signaling responses, NPY and related peptides can induce histamine release from mast cells, but it is questionable whether this is a receptor-mediated event (Mousli and Landry, 1994). Finally, it has been claimed that NPY might act via so-called “sigma sites” (Roman et al., 1989), but this could not be confirmed by others (Tam and Mitchell, 1991).
VI. Recommendations for Classification of Neuropeptide Y/Peptide YY/Pancreatic Polypeptide-Mediated Responses
The pharmacological profiles of the Y1and Y2 receptors have now been clearly established, but those of the Y4, Y5, and y6 receptors certainly require a more extensive investigation, particularly with regard to the endogenously expressed receptors. The classification of NPY/PYY/PP-induced functional responses still relies largely upon the use of agonists because (a) few antagonists of NPY receptors have been described, (b) most of them have not systematically been evaluated against all types and (c) most are not yet readily available for widespread use. A clear classification of a functional response should employ at least the endogenous agonists NPY, PYY, and PP and one each of the [Pro34]substituted analogs and C-terminal fragments. In this context, it should be noted that in some cases the species of origin of the agonist may be important for its potency, and this is particularly true of species variants of PP. The development of other selective antagonists, particularly with selectivities for other Y receptor types, is an urgent prerequisite to the full pharmacological characterization of the newer Y receptors. Although none of the antagonists reported so far appears to have specific advantages, BIBP 3226 has been the best investigated. Moreover, it is the only drug that has been evaluated in cells expressing one each of the five receptors cloned thus far and an inactive stereoisomer is available as a control.
Finally, we suggest that investigators cloning additional types of the NPY/PYY/PP family consult as soon as possible with the nomenclature committee before designation of novel names to minimize further confusion in this burgeoning pharmacological field. The committee assures these parties that any information received in this way will be handled in the strictest confidence.
Footnotes
-
↵FNa Address for correspondence: Martin C. Michel, Nephrology Laboratory IG 1, Klinikum, 45122 Essen, Germany.
- Abbreviations:
- cAMP
- cyclic adenosine 3′,5′-cyclic monophosphate
- cDNA
- complementary deoxyribonucleic acid
- CNS
- central nervous system
- HEL
- human erythroleukemia cells
- mRNA
- messenger ribonucleic acid
- NPY
- neuropeptide Y
- PP
- pancreatic polypeptide
- PYY
- peptide YY
- The American Society for Pharmacology and Experimental Therapeutics
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- I. Introduction
- II. Signal Transduction of Neuropeptide Y Receptors
- III. Historical Aspects of Neuropeptide Y Receptor Subdivision
- IV. Present Definition of Neuropeptide Y Receptors
- V. Characteristics of Receptor Subtypes (table1)
- VI. Recommendations for Classification of Neuropeptide Y/Peptide YY/Pancreatic Polypeptide-Mediated Responses
- Footnotes
- References
- Figures & Data
- Info & Metrics
- eLetters