


Abstract
Receptors for corticotropin-releasing factor (CRF) are members of a family of G protein-coupled receptors (“Family B”) that respond to a variety of structurally dissimilar releasing factors, neuropeptides, and hormones (including secretin, growth hormone-releasing factor, calcitonin, parathyroid hormone, pituitary adenylate cyclase-activating polypeptide, and vasoactive intestinal polypeptide) and signal through the cyclic AMP and/or calcium pathways. To date, three genes encoding additional CRF-like peptides (urocortins) have been identified in mammals. The urocortins and CRF bind with differential ligand selectivity at the two mammalian CRF receptors. This report was prepared by the International Union of Pharmacology Subcommittee on CRF Receptors, to summarize the current state of CRF receptor biology and to propose changes in the classification and nomenclature of CRF ligands and receptors.
I. Introduction
In 1981, the 41-amino acid corticotropin-releasing factor (CRF1) peptide was isolated from ovine hypothalamus (Vale et al., 1981). Initially, the action of CRF appeared to be restricted to regulating ACTH secretion by pituitary corticotrope cells (Vale et al., 1981; Dunn and Berridge 1990; Hauger and Dautzenberg, 1999; Smagin et al., 2001). The central distribution of CRF pathways, however, suggested that CRF's function extended far beyond the classical action of a hormone (Swanson et al., 1983; De Souza et al., 1984; Wynn et al., 1984; Vale et al., 1997; Arborelius et al., 1999; Heinrichs and De Souza 1999;Dautzenberg et al., 2001a). Because CRF was shown to generate neuroendocrine, autonomic, and behavioral stress responses, it was hypothesized that CRF contributed to the development of stress and affective disorders by over-activating its receptors expressed in various brain neurons in neocortex, the amygdala and its extended neurocircuits, and brainstem nuclei.
Urocortin, a second mammalian CRF-like peptide was identified after two novel CRF-like peptides, urotensin I (Lederis et al., 1982) and sauvagine (Montecucchi and Henschen, 1981), were discovered in fish and amphibian species in addition to CRF (Okawara et al., 1988;Stenzel-Poore et al., 1992). Human, sheep, rat, and mouse urocortin possess a 40-amino acid sequence (Vaughan et al., 1995; Donaldson et al., 1996; Zhao et al., 1998; Cepoi et al., 1999) and a high degree of homology with fish urotensin I. Although the Edinger-Westphal locus, the hypothalamus, and a small population of forebrain neurons express substantial levels of urocortin (Bittencourt et al., 1999), urocortin is more broadly expressed in the periphery, especially in the pituitary, gastrointestinal tract, testis, cardiac myocytes, thymus, spleen, and kidney (Kageyama et al., 1999).
Very recently, two novel isoforms of urocortin, urocortin 2 and urocortin 3 (Lewis et al., 2001; Reyes et al., 2001), were cloned at the Salk Institute from human and mouse cDNA libraries. At the same time, another group identified two similar peptides, which they named stresscopin (which is homologous with urocortin 3) and stresscopin-related peptide (which is homologous with urocortin 2) (Hsu and Hsueh, 2001). Urocortin 2 (stresscopin-related peptide) and urocortin 3 (stresscopin) are discretely expressed in the central nervous system with a pattern distinct from the known CRF and urocortin pathways (Hsu and Hsueh, 2001; Lewis et al., 2001; Reyes et al., 2001). Furthermore, the central distribution of urocortin 1-, urocortin 3-, and CRF2 receptor-expressing neurons suggests that urocortin 1 may serve as the major CRF2receptor ligand in the hindbrain whereas urocortin 3 may serve as the major CRF2 receptor ligand in the forebrain. Urocortin 2 or a novel endogenous ligand may signal at CRF2 receptors expressed in certain brain regions lacking urocortin 1 or urocortin 3 innervations (e.g., the hippocampus and certain regions of the cerebral cortex). Because neither CRF1 nor CRF2 receptors have yet been identified in the locus coeruleus, central nucleus of the amygdala, and several other stress-sensitive brain structures, it is possible that a novel CRF receptor may eventually be cloned (Hsu and Hsueh, 2001; Li et al., 2002). In the periphery, urocortin 2 mRNA is detected in the heart, adrenal gland, and peripheral blood cells (Hsu and Hsueh, 2001; Reyes et al., 2001). The highest peripheral levels of urocortin 3 mRNA expression have been detected in the gastrointestinal tract, muscle, adrenal gland, and skin (Hsu and Hsueh, 2001; Lewis et al., 2001). Because urocortin 2 and urocortin 3 have been identified by molecular cloning strategies (Hsu and Hsueh, 2001; Lewis et al., 2001;Reyes et al., 2001), their exact size has not been established yet. The structure of both precursor genes appears to predict 38-amino acid mature peptides (Lewis et al., 2001; Reyes et al., 2001) although one group (Hsu and Hsueh, 2001) postulated the existence of N-terminally extended peptides 40 (urocortin 3) and 43 (urocortin 2) amino acids in length. However, because the human and mouse homologues of both precursors are less conserved in the extended N terminus than in the remaining sequence, it seems more likely that urocortin 2 and urocortin 3 exist as 38-amino acid peptides. Furthermore, the 38- and the 40-/43-amino acid versions of both ligands possess similar pharmacological potencies. Because CRF ligands rapidly lose agonistic potency upon N-terminal truncation (Rivier et al., 1984; Brauns et al., 2002) the pharmacological data also speak in favor of the 38-amino acid variants for urocortin 2 and urocortin 3. This issue will not be conclusively resolved until endogenous forms of these two urocortin peptides are isolated in various species. Like CRF (Vale et al., 1981) and urocortin 1 (Vaughan et al., 1995), urocortin 2 and urocortin 3 possess biological activity only when they are C-terminally amidated (Hsu and Hsueh, 2001). Only 4 amino acids are completely conserved among CRF peptides (Fig. 1). Therefore, secondary structure rather than linear sequence homology most likely determines differences in biological activity of the members of the CRF peptide family (Hsu and Hsueh, 2001; Lewis et al., 2001; Reyes et al., 2001; Dautzenberg and Hauger, 2002).

Alignment of the members of the CRF peptide family. The amino acids that are homologous between the CRF peptides are boxed. h, human; m, mouse; o, ovine; UCN 1, urocortin 1; UCN 2, urocortin 2; UCN 3, urocortin 3.
II. The Corticotropin-Releasing Factor Type 1 Receptor
The CRF1 receptor, a 415–446 amino acid polypeptide, has been cloned from a variety of species including man (Chen et al., 1993; Vita et al., 1993), tree shrew (Palchaudhuri et al., 1998), mouse (Vita et al., 1993), rat (Chang et al., 1993; Perrin et al., 1993), sheep (Myers et al., 1998), chicken (Yu et al., 1996), frog (Dautzenberg et al., 1997), and fish (Arai et al., 2001;Pohl et al., 2001). Although a larger number of splice variants of the CRF1 receptor cDNA have been identified (Chang et al., 1993; Chen et al., 1993; Ross et al., 1994; Myers et al., 1998;Grammatopoulos et al., 1999) (Fig. 2), they have not been shown to encode functional receptors in vivo due to their low binding affinity or lack of activation in recombinant systems (reviewed in Dautzenberg et al., 2001a). The CRF1receptor is abundantly expressed in the central nervous system with major expression sites in cortex, cerebellum, hippocampus, amygdala, olfactory bulb, and pituitary (Potter et al., 1994; Chalmers et al., 1996; Palchaudhuri et al., 1998). In the periphery, CRF1 receptor mRNA is expressed at low levels in the skin, ovary, testis, and adrenal gland (Vita et al., 1993; Nappi and Rivest, 1995; Palchaudhuri et al., 1998).

Two-dimensional structure of the human CRF1 and CRF2 receptors. Identical amino acids between both receptors are represented as filled circles whereas divergent residues are shown as open circles. The arrows indicate sites for insertion or deletion of exons in nonfunctional variants of the CRF1 and CRF2 receptor (Chen et al., 1993;Myers et al., 1998; Grammatopoulos et al., 1999; Miyata et al., 1999). The symbols { and } indicate the deletion of a 40-amino acid exon in a nonfunctional splice variant of the human CRF1receptor (Ross et al., 1994) whereas the symbol ] represents the common splice site for the three CRF2 variants CRF2(a), CRF2(b), and CRF2(c).
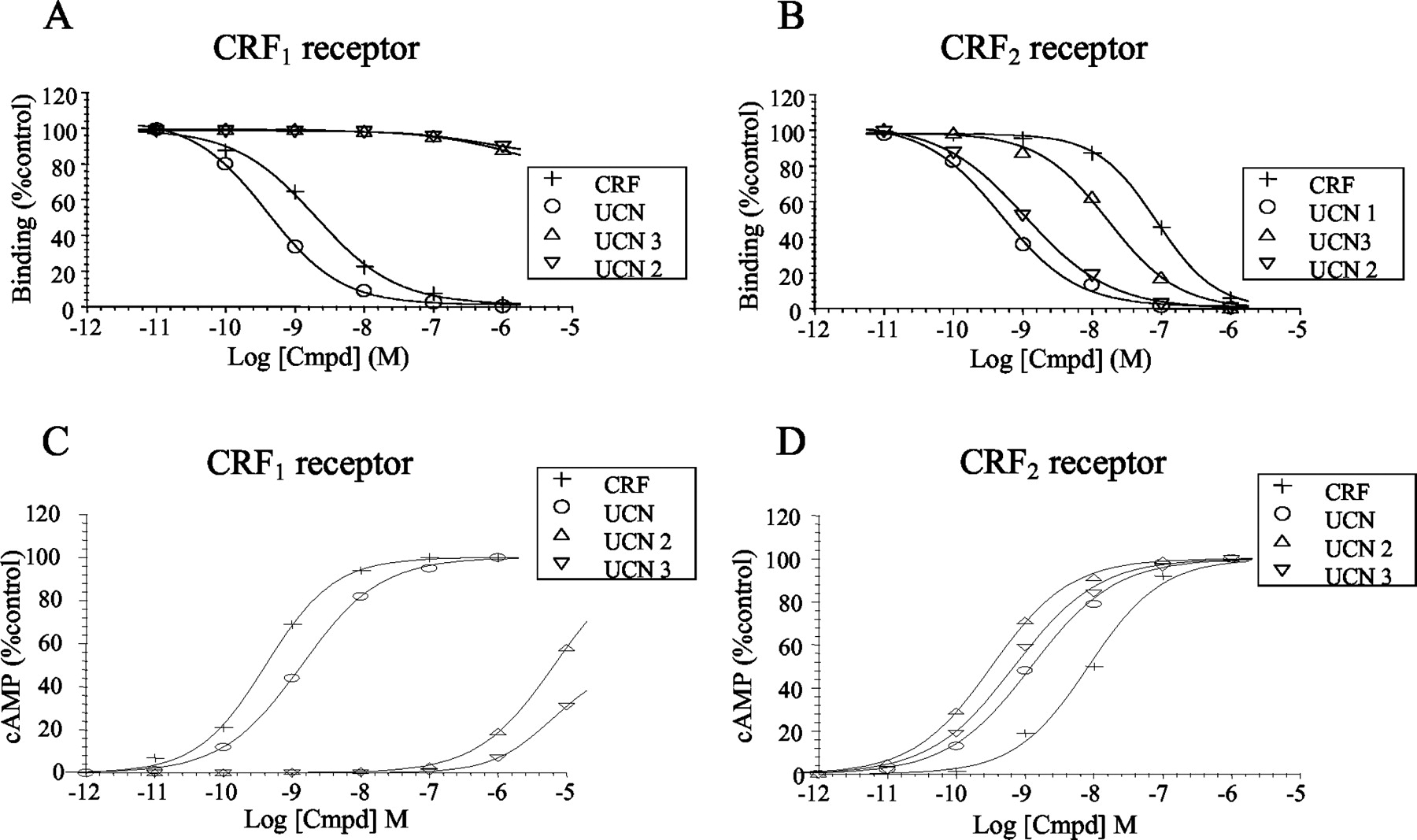
Binding and functional studies using cell lines recombinantly or endogenously expressing CRF1 receptors revealed a distinct ligand-selective profile whereby human and ovine CRF, urocortin 1, urotensin I, and sauvagine all bind with high affinity to the mammalian CRF1 receptor and activate the cyclic AMP signaling pathway (Donaldson et al., 1996; Dautzenberg et al., 1997, 2001b; Perrin et al., 1999). In contrast, urocortin 2 and urocortin 3 do not bind to or activate CRF1receptors (Hsu and Hsueh, 2001; Lewis et al., 2001; Reyes et al., 2001). Therefore, CRF and urocortin 1 can be classified as the endogenous ligands for mammalian CRF1 receptors (Fig. 3).

Pharmacological properties of the four mammalian CRF agonists CRF, urocortin 1, urocortin 2, and urocortin 3 at the human CRF1 and CRF2 receptors. Binding of the four natural agonists to the CRF1 (A) and CRF2(B) receptors reveals a distinct profile for each ligand with urocortin 1 being a high-affinity ligand to both receptors and CRF being a high affinity ligand to the CRF1 and a low-affinity ligand to the CRF2 receptor. Urocortin 2 and urocortin 3, however, only bind to the CRF2 receptor. Similar findings are observed in cAMP stimulation studies with recombinant receptors (C and D). In contrast to the binding studies, however, a very weak agonistic potency (>1 μM) is observed for urocortin 2 and urocortin 3 at the CRF1 receptor (C).
III. The Corticotropin-Releasing Factor Type 2 Receptor
Complementary DNAs for the CRF2 receptor have been isolated from man (Liaw et al., 1996; Valdenaire et al., 1997; Kostich et al., 1998), tree shrew (Palchaudhuri et al., 1999), rat (Lovenberg et al., 1995b), mouse (Kishimoto et al., 1995; Perrin et al., 1995; Stenzel et al., 1995), frog (Dautzenberg et al., 1997), and fish (Arai et al., 2001; Pohl et al., 2001). Three functional splice variants (Lovenberg et al., 1995b; Kostich et al., 1998) and a truncation variant (Miyata et al., 1999) have been identified for the mammalian CRF2 receptor. The CRF2(a) receptor variant is only expressed in nonmammalian species (Dautzenberg et al., 1997; Arai et al., 2001;Pohl et al., 2001), whereas the 430–438 amino acid CRF2(b) receptor and the CRF2(a) receptor are both expressed in mammals (Kishimoto et al., 1995; Lovenberg et al., 1995b; Perrin et al., 1995;Stenzel et al., 1995; Valdenaire et al., 1997; Palchaudhuri et al., 1999). Expression of the 397-amino acid CRF2(c)receptor has only been detected in limbic regions of the human central nervous system (Kostich et al., 1998). Splicing of the CRF2 receptor variants occurs at the extreme 5′ terminus of the receptor gene and reflects usage of different promoters in man. The hCRF2 gene, which is located on chromosome 7p14-15, is ∼50 kb in size and contains 15 exons (Dautzenberg et al., 2000). The first four exons give rise to the different 5′-ends of the splice variants hCRF2(a), hCRF2(b) and hCRF2(c), respectively; exons 5 to 15 form the common parts of the various hCRF2 splice variants. Exons 1 and 2, which are separated by ∼10.5 kb intronic sequence encode the CRF2(b)-specific part, followed by the CRF2(c) and CRF2(a) exons (Dautzenberg et al., 2000).
In rodents, CRF2(a) receptor mRNA is expressed primarily in brain neurons whereas CRF2(b)receptor mRNA is detected in non-neuronal brain structures such as the choroid plexus cerebral arterioles and peripheral tissues (Lovenberg et al., 1995a). The CRF2(a) receptor is the dominant CRF2 receptor splice variant expressed in the mammalian brain. Central CRF2(a)receptors are expressed in a discrete pattern with highest densities in the lateral septum, ventromedial hypothalamus, cortical nucleus of the amygdala, dorsal raphe, nucleus of the solitary tract, and the choroid plexus; whereas, a widespread but discrete expression of CRF1 receptors is found in neocortex and the amygdala and its extended neurocircuits (Bittencourt et al., 1999; Li et al., 2002). In humans and tree shrews, CRF2(a)receptor mRNA has also been detected in the brainstem, olfactory bulbs, cortex, cerebellum, retina, and pituitary (Palchaudhuri et al., 1999;Sanchez et al., 1999). Because CRF2(b) receptor mRNA can be detected in neuronal structures such as retina and cerebellum, the distribution of these two splice variants may overlap in primates and primate-like animals. In the periphery, substantial expression of CRF2 receptor can be found in the heart, skeletal muscle, vasculature, and gastrointestinal tract. The CRF2(a) receptor is the major splice variant found in the peripheral tissues of humans (e.g., heart and skeletal muscle) (Valdenaire et al., 1997; Kostich et al., 1998), whereas the CRF2(b) splice variant is the CRF2 receptor peripherally expressed in rodents. The main expression sites for the rodent CRF2(b)receptor have been reported for heart, lung, skeletal muscle, gastrointestinal tract, testis, and ovaries (Lovenberg et al., 1995a;Palchaudhuri et al., 1999).
Pharmacological characterization of the CRF2receptor splice variants revealed no major differences between CRF2(a), CRF2(b), and CRF2(c) receptors (Donaldson et al., 1996;Kostich et al., 1998; Palchaudhuri et al., 1999). However, the binding profiles of these three CRF2 receptors strongly diverge from the binding profile of the CRF1receptor (Donaldson et al., 1996; Perrin et al., 1999; Dautzenberg et al., 2001b; Hsu and Hsueh, 2001; Lewis et al., 2001; Reyes et al., 2001). The nonmammalian CRF peptides urotensin I and sauvagine and the mammalian peptides urocortin 1, urocortin 2, and urocortin 3 generally bind with up to 100-fold higher affinities to the CRF2 receptor than species homologues of CRF (Fig. 3B). In agreement with the binding data, a similar rank order of potency is typically observed when stimulation of intracellular cyclic AMP accumulation is measured (Donaldson et al., 1996; Dautzenberg et al., 2001b; Hsu and Hsueh, 2001; Lewis et al., 2001; Reyes et al., 2001) (Fig. 3D). Therefore, currently, urocortin 2 and urocortin 3 are generally considered to represent the endogenous ligands for mammalian CRF2 receptor variants, whereas urocortin 1 is thought to be an endogenous ligand for both the CRF1 and CRF2 receptors.
IV. Proposed Nomenclature
Early clinical neuroendocrine studies, particularly those directed at investigating the role of CRF in regulating the hypothalamic-pituitary-adrenal axis, defined this peptide's function to be a stimulator of ACTH secretion. Accordingly, the term “corticotropin-releasing hormone” gained acceptance in the neuroendocrine literature. However, ample evidence has accumulated over the past two decades demonstrating that CRF's physiological function extends far beyond the biology of a hormone. In fact, CRF most likely plays a role in immune, neurodegenerative, neuropsychiatric, and gastrointestinal disorders. Thus, the term “corticotropin-releasing factor” more appropriately designates the function of this peptide. It is recommended that CRF rather than corticotropin-releasing hormone (CRH) should be used in keeping with the nomenclature for other important factors (i.e., transforming growth factor and brain-derived neurotrophic factor) that exhibit a multiplicity of biological actions.
The three-dimensional structure, CRF2 receptor binding profile, and biological actions of urocortin 2 and urocortin 3 are similar to each other and to urocortin 1 (see Hsu and Hsueh, 2001;Lewis et al., 2001; Reyes et al., 2001; Dautzenberg and Hauger, 2002). These novel ligands should therefore retain the following designations: urocortin 2 and urocortin 3. Stresscopin, which has recently been used to designate the peptide with urocortin 2's sequence, implies its role is to mediate an organism's “coping” with stress. Activation of central CRF2 receptors can produce anxiogenesis or anxiolysis depending on the brain site (Reul and Holsboer, 2002). Thus, the term “stresscopin” (Hsu and Hsueh, 2001) only covers one aspect of the behavioral response mediated by brain CRF2 receptor signaling. Therefore, the urocortin 2 and urocortin 3 should be used in place of the terms stresscopin and stresscopin-related peptide, respectively. Finally, since the urocortin family contains three members, urocortin 1 will replace the earlier generic term “urocortin”. Table 1summarizes all of the recommended changes for the nomenclature of CRF peptides.
Nomenclature of the members of the CRF ligand and receptor family
A revised nomenclature for CRF receptors is summarized in Table2. Although several alternative terminologies such as CRH1, CRFR1 or CRF-R1 have been used to designate the CRF1 receptor, the term CRF1 receptor should be utilized. Because the CRF2 receptor is highly selective for urocortin 1 over CRF, renaming the CRF2 receptor as the urocortin receptor was considered. However, urocortin 1 binds equally well to the CRF1 and CRF2receptors. Therefore, redesignating the CRF2receptor as the urocortin receptor fails to distinguish the biology of the two receptors. Therefore, the term CRF2receptor should be used for the time being to maintain clarity (Table2). As new insight into the normal and pathophysiological actions of the CRF2 receptor subtypes is gained, and as species variations and the behavioral roles of the CRF2 receptor subtypes become understood, the naming of this CRF receptor may require future revisions. In addition, in accordance with IUPHAR convention, CRF receptor splice variants should be listed alphabetically using Roman rather than Greek letters as have appeared in the current literature (Humphrey and Barnard, 1998). Thus, the three CRF2 receptor splice variants will be designated as CRF2(a), CRF2(b), and CRF2(c). Finally, as novel CRF1 and CRF2 splice variants become identified, they will only be classified as formal CRF receptors if they encode physiologically functional receptor proteins but not simply polymerase chain reaction products. Numerous low-affinity or nonfunctional CRF receptor variants possess an incomplete structure due to either exon deletions or insertions, which mostly occur in a single species (reviewed in Dautzenberg et al., 2001a).
Nomenclature of CRF receptors
V. Unresolved Issues
Recently a third CRF receptor, termed CRF3, was cloned from catfish (Arai et al., 2001). The catfish CRF3 receptor was not detected in salmon (Pohl et al., 2001). The catfish CRF1 and CRF3 receptors are highly homologous. Interestingly, CRF3 receptor expression is restricted to the catfish pituitary, a tissue that normally expresses CRF1 receptors in other species (Arai et al., 2001). Species homologues for this receptor may not exist.
Human urocortin 2 lacks the standard consensus site required for proteolytic cleavage and C-terminal amidation, which is a prerequisite for biological potency (reviewed in Dautzenberg and Hauger, 2002). Therefore, human urocortin 2 may not be processed into a biologically active peptide in vivo (Hsu and Hsueh, 2001; Reyes et al., 2001). The isolation of the urocortin 2 peptide from native human tissues would resolve this issue.
Acknowledgments
We thank Profs. A. J. Harmar and M. Spedding for liaison with the IUPHAR Committee on Receptor Nomenclature and Drug Classification (NC-IUPHAR).
Footnotes
-
Address correspondence to: Dr. Frank M. Dautzenberg, c./o. Axovan Ltd., Gewerbestrasse 16, CH-4070 Allschwil, Switzerland. E-mail:frank.dautzenberg{at}t-online.de
-
↵1 CRF, corticotropin-releasing factor; IUPHAR, International Union of Pharmacology; ACTH, adrenocorticotropin; CRH, corticotropin-releasing hormone; kb, kilobase(s).
-
Article, publication date, and citastion information can be found athttp://pharmrev.asptjournals.org
-
DOI: 10.1124/pr.55.1.3
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}