


Abstract
A gene encoding a novel class a G-protein-coupled receptor was discovered in 1993 by homology cloning and was called APJ. It was designated an “orphan” receptor until 1998, when its endogenous ligand was identified and named apelin (for APJ endogenous ligand). Since this pairing, both apelin and its receptor have been found to have a widespread distribution in both the central nervous system and the periphery. A number of physiological and pathophysiological roles for the receptor have emerged, including regulation of cardiovascular function, fluid homeostasis, and the adipoinsular axis. This review outlines the official International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification nomenclature, designating the receptor protein as the apelin receptor, together with current knowledge of its pharmacology, distribution, and functions.
I. Introduction
Since the discovery of the gene encoding the apelin receptor, originally called APJ, and pairing of the protein with its cognate ligands, the apelin peptides, a number of physiological and pathophysiological roles for the receptor have emerged. These include cardiovascular regulation, fluid homeostasis, modulation of the adipoinsular axis, and HIV coreceptor function in vitro. Here we summarize the discovery, deorphanization, and recommended nomenclature with subsequent characterization of the physiological and pathophysiological roles of the apelin receptor. The primary aim of this review is to outline the recommended nomenclature for this receptor. For comprehensive overviews of the apelin field, see Kleinz and Davenport (2005), Masri et al. (2005), Carpéné et al. (2007), Davenport et al. (2007), and Ladeiras-Lopes et al. (2008). Details of the pairing of APJ with apelin are outlined in Hinuma et al. (1999). For more information on the roles of apelin and its receptor in cardiovascular function, see Lee et al. (2006), Sorli et al. (2006), Chandrasekaran et al. (2008), Japp and Newby (2008), Quazi et al. (2009), and Kalea and Batlle (2010). The involvement of apelin in fluid homeostasis is described by Brown et al. (2008), Bundzikova et al. (2008) and Llorens-Cortes and Moos (2008). Castan-Laurell et al. (2005), Bełtowski (2006), Lee et al. (2006), and Rayalam et al. (2008) specifically review the contribution of this system in the adipoinsular axis.
II. The Apelin Receptor: Recommendations for Nomenclature
In 1993, a gene encoding a novel class A G-protein-coupled receptor (GPCR1) was discovered by homology cloning. It was located on chromosome 11 band q12 and showed greatest sequence homology with the angiotensin AT1 receptor (54% in the transmembrane regions) but did not bind angiotensin II. It was therefore designated an “orphan” GPCR, having no known ligand, and was named APJ by O'Dowd et al., (1993). The approved Human Genome Organization (HUGO) gene symbol for APJ is now APLNR. The gene has a number of other aliases, including AGTRL1, APJ, APJR, and FLJ90771. The endogenous ligand for this receptor was later identified as apelin, which led the International Union of Pharmacology (IUPHAR) to recommend “apelin receptor” as the nomenclature for the receptor protein (Davenport and Kleinz, 2008) (Table 1). This follows the convention of naming the receptor protein after its endogenous ligand.
IUPHAR recommendations on receptor nomenclature
III. Receptor Structure
The human apelin receptor comprises 380 amino acid residues and has the typical 7-transmembrane domain structure of a class A GPCR. It contains consensus sites for phosphorylation by cAMP-dependent protein kinase, palmitoylation, and glycosylation (O'Dowd et al., 1993). The apelin receptor has been identified in a number of other species, including mouse, rat, cow, rhesus macaque, Xenopus laevis, and Danio rerio. The mouse apelin receptor has 377 amino acids and 91% sequence homology with the human receptor, whereas the rat apelin receptor has 377 amino acids and 89% sequence homology with the human receptor (Fig. 1).

Alignment of human, rat, and mouse apelin receptors. *, identical amino acids, :, conserved amino acid substitution; ., semiconserved amino acid substitution.
To date, there is no evidence for multiple receptor subtypes in mammals. During the initial receptor identification, a polymerase chain reaction strategy using oligonucleotides based on the apelin receptor yielded no closely related genes (O'Dowd et al., 1993). In addition, saturation binding experiments in human tissues gave Hill coefficients close to unity, indicating that the radioligand bound to a single receptor population (Katugampola et al., 2001), although this does not exclude the possibility of two receptor subtypes with the same affinity.
Activation of apelin receptors expressed in cell lines inhibited forskolin-stimulated cAMP production, suggesting that the receptor is coupled to inhibitory G-proteins (Gi) (Habata et al., 1999), which is supported by the finding that apelin actions are pertussis toxin-sensitive (Hosoya et al., 2000; Masri et al., 2002). A number of interactions between the apelin and angiotensin systems have been reported, including recent evidence that the apelin receptor forms heterodimers with the angiotensin AT1 receptor (Chun et al., 2008).
IV. Endogenous Agonists
In 1998, the endogenous ligand for APJ was identified as a 36-amino acid peptide named apelin (for APJ endogenous ligand), isolated from bovine stomach extracts. This peptide induced extracellular acidification in CHO cells expressing apelin receptors (Tatemoto et al., 1998). cDNA encoding a 77-amino acid prepropeptide (preproapelin) was identified in human and bovine tissue (Tatemoto et al., 1998), showing considerable sequence similarity across all species examined, with the last 23 residues of the C terminus being identical in mammals (Fig. 2).

Sequence alignment of mammalian, fish and amphibian apelin-36 amino acid sequences. *, identical amino acids; :, conserved amino acid substitution; ., semiconserved amino acid substitution. Residues that differ from the human sequence are highlighted in red.
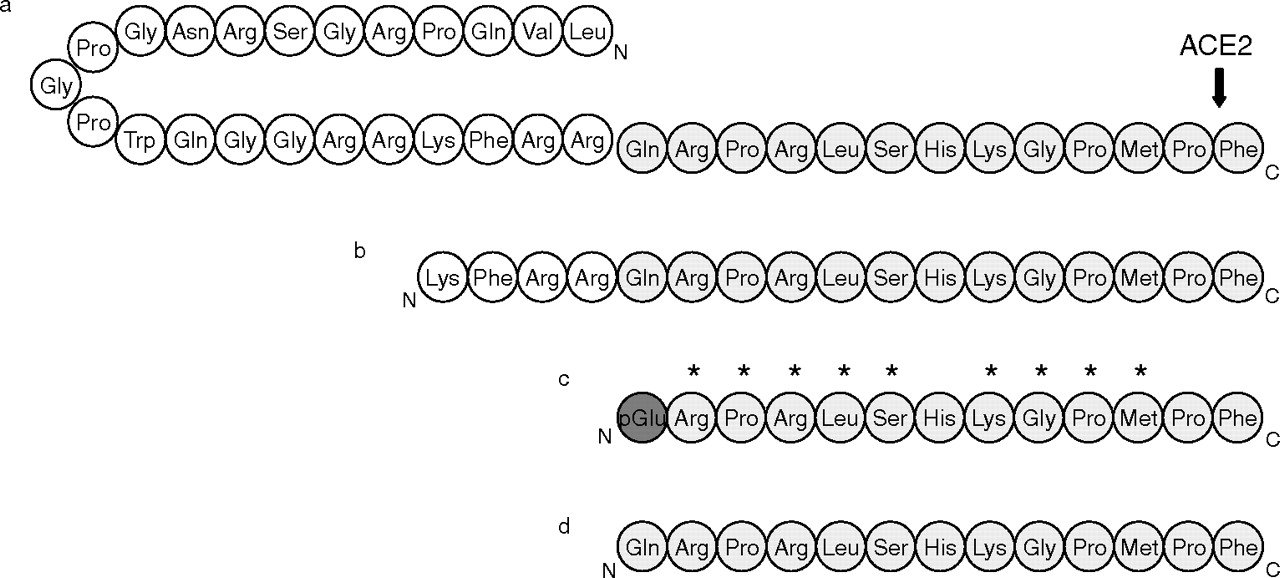
Native preproapelin exists as a dimer, stabilized by disulfide bridges, in mouse heart (Lee et al., 2005). Preproapelin contains a number of paired basic amino acids residues (Arg-Arg and Arg-Lys) that are possible cleavage sites for endopeptidases (Habata et al., 1999). Cleavage at these sites would produce a predicted family of C-terminal fragments, including apelin-36, apelin-17, apelin-13, and the post-translationally modified (Pyr1)apelin-13 (Fig. 3), which are all agonists at the apelin receptor (Table 2). The lack of cysteine residues in these C-terminal fragments suggests that the mature peptides are monomeric.

Amino acid sequence of the endogenous apelin receptor agonists apelin-36 (a), apelin-17 (b), (Pyr1)apelin-13 (c), and apelin-13 (d). Shaded residues are those identical in all peptides. The post-translational modification of the N-terminal glutamate of apelin-13 to pyroglutamate is shown in dark gray. ACE2 cleaves apelin-36 and apelin-13 at the position shown (Vickers et al., 2002). *, residues found to be important for binding and activation of the apelin receptor by apelin-13 (Fan et al., 2003; Medhurst et al., 2003; El Messari et al., 2004).
Actions and affinities of endogenous apelin receptor agonists
The degradative pathway for apelin peptides is unknown, but the recently identified angiotensin-converting enzyme 2 (ACE2) cleaves the C-terminal phenylalanine from apelin-13 and apelin-36 (Vickers et al., 2002). However, this cleavage is unlikely to inactivate the peptide, because an in vitro structure activity study showed that a fragment lacking this residue retained binding and functional activity at the rat apelin receptor expressed in CHO cells (El Messari et al., 2004). However, this fragment did not induce apelin receptor internalization in vitro and lost hypotensive activity in the rat in vivo, suggesting that cleavage by ACE2 may produce apelin fragments that induce a conformational state of the apelin receptor different from that induced by the full-length peptides (El Messari et al., 2004). It has recently been reported that (Pyr1)apelin-13, lacking the C-terminal phenylalanine, has comparable affinity and agonist activity at the native apelin receptor in human tissues in vitro (Pitkin et al., 2009), suggesting that cleavage of apelin by ACE2 is not an inactivating step in man.
The existence of multiple predicted apelin isoforms raises the question of which isoform is most important in vivo. All of these predicted isoforms have been shown to be present in vivo, but the predominant apelin isoform in human cardiac tissue is (Pyr1)apelin-13 (Maguire et al., 2009), which is not unexpected because the pyroglutamate moiety protects the N terminus of peptides from exopeptidase degradation (Van Coillie et al., 1998). The predominant isoforms in plasma are (Pyr1)apelin-13, apelin-13 and apelin-17 (De Mota et al., 2004; Miettinen et al., 2007; Azizi et al., 2008), whereas apelin-36 is the major isoform present in colostrom (Hosoya et al., 2000). Rat brain was found to contain both (Pyr1)apelin-13 and apelin-17 (De Mota et al., 2004).
The relative potency of the apelin peptides varies between experimental systems, (Pyr1)apelin-13 and apelin-13 being the most potent activators of apelin receptors expressed in cell lines (Tatemoto et al., 1998; Habata et al., 1999; Kawamata et al., 2001; Medhurst et al., 2003), whereas apelin-36 is the most potent inhibitor of HIV infection of cells in vitro (Zou et al., 2000). However, (Pyr1)apelin-13, apelin-13, and apelin-36 are equipotent mediators of vascular tone and cardiac contractility in human tissues in vitro, with pD2 (negative log10 of the molar concentration required to produce half-maximal response) values in the range of 8.8 to 10.4 (Maguire et al., 2009).
V. Receptor Distribution
A. Rat
Within the rat central nervous system (CNS), mRNA encoding the apelin receptor is found in a number of areas, including the supraoptic (SON) and paraventricular (PVN) nuclei of the hypothalamus, the pineal gland, and the pituitary (De Mota et al., 2000; Lee et al., 2000; O'Carroll et al., 2000). Apelin receptor protein also has a widespread distribution within the rat brain, including the cerebral cortex, basal ganglia, hypothalamus, thalamus, midbrain, and reticular formation (Medhurst et al., 2003). It is noteworthy that apelin receptor mRNA and protein colocalize with vasopressin in a subset of magnocellular neurons within the SON and PVN of the rat hypothalamus (Reaux et al., 2001; O'Carroll and Lolait, 2003; De Mota et al., 2004), suggesting a role for the receptor in water balance.
Apelin receptor transcripts are found in most peripheral rat tissues, the highest levels being found in the lung, heart, and kidney (Hosoya et al., 2000; O'Carroll et al., 2000; Medhurst et al., 2003). Receptor protein is present in rat lung and heart and at lower levels in the kidney cortex (Katugampola et al., 2001).
B. Human
mRNA encoding the apelin receptor is widely distributed within the human CNS but is highest in the spinal cord, corpus callosum, and medulla (Matsumoto et al., 1996; Edinger et al., 1998; Medhurst et al., 2003;). The distribution of receptor protein within the human brain has not yet been determined.
As in rats, apelin receptor mRNA is found in most human peripheral tissues. However, the highest levels are found in human spleen and placenta, whereas it is highest in lung in the rat. Receptor is also present in human heart, liver, lung, kidney, and stomach (Edinger et al., 1998; Medhurst et al., 2003). The widespread distribution of apelin receptor transcripts in the periphery was consistent with a vascular localization. Autoradiography revealed apelin binding sites within a range of human large-diameter blood vessels (Katugampola et al., 2001), which was confirmed by immunohistochemistry showing localization of apelin receptor to vascular endothelial and smooth muscle cells (Kleinz et al., 2005). The presence of the apelin receptor on both the vascular endothelium and smooth muscle suggested a role in endothelium-dependent and -independent modulation of vascular tone. Apelin receptors are also present on human cardiomyocytes (Kleinz et al., 2005), raising the possibility of effects on cardiac contractility.
VI. Apelin Peptide Distribution
A. Rat
Like its receptor, apelin peptide has a widespread distribution throughout the CNS and periphery, and there is a large amount of overlap in the expression profiles of the receptor and peptide. In the rat, mRNA encoding apelin is found within a number of CNS regions, including the nucleus accumbens, spinal cord, cortex, striatum, midbrain, hippocampus, cerebellum, and pituitary (Lee et al., 2000; Medhurst et al., 2003). Apelin peptide is expressed at highest levels in the rat pineal gland, pituitary, arcuate nucleus, and SON and PVN of the hypothalamus (Reaux et al., 2002; De Mota et al., 2004). Within the SON and PVN, the majority of apelin-positive neurons also express vasopressin (De Mota et al., 2004; Reaux-Le Goazigo et al., 2004).
In the rat periphery, apelin mRNA is highest in the lung, with high levels also found in the mammary gland and heart (Habata et al., 1999; O'Carroll et al., 2000; Kawamata et al., 2001; Medhurst et al., 2003). A similar distribution is shown by the peptide (Kawamata et al., 2001).
B. Human
Within the human CNS, apelin mRNA is present in all regions tested, with highest expression in the spinal cord, corpus callosum, amygdala, substantia nigra, and pituitary gland (Medhurst et al., 2003).
Peripheral apelin mRNA, like its receptor, is found at highest levels in the placenta, with lower levels in the heart, lung, and kidney (Medhurst et al., 2003). Apelin is present in a range of cell types within the placenta, lung, and kidney (De Falco et al., 2002) but has a restricted distribution within the cardiovascular system, being expressed only by vascular and endocardial endothelial cells (Kleinz and Davenport, 2004).
VII. Synthetic Agonists
A number of synthetic apelin analogs have biological activity. Apelin-12 is the smallest C-terminal fragment to bind and activate the apelin receptor (El Messari et al., 2004), and any apelin fragment containing this 12 amino acid core maintains agonist function. Cyclic apelin analogs (Fig. 4) have been found to have binding and functional activity at the apelin receptor (Hamada et al., 2008; Macaluso et al., 2009), giving important structural insights into the apelin binding site. The first nonpeptidic agonist at the apelin receptor, E339–3D6, was reported by Iturrioz et al. (2009).

Cyclic apelin analogs drawn from the sequences reported by Hamada et al. (2008) (a) and Macaluso et al. (2009) (b).
VIII. Antagonists
A small-molecule antagonist of the chemokine receptor CXCR4, ALX40-4C, has been shown to directly bind apelin receptors expressed in cell lines and to block ligand-induced receptor internalization and signaling. It also inhibits apelin receptor-mediated cell membrane fusion. This suggests that ALX40-4C is a nonspecific apelin receptor antagonist (Table 3) (Zhou et al., 2003).
Actions and affinities of apelin receptor antagonists
The role of the C-terminal phenylalanine of the apelin peptides has been of particular interest since the discovery that the recently identified homolog of ACE, ACE2, cleaves this residue from apelin peptides. Apelin-13 with the C-terminal phenylalanine mutated to alanine [Apelin-13(F13A); Gln-Arg-Pro-Arg-Leu-Ser-His-Lys-Gly-Pro-Met-Pro-Ala] has been shown to act as an apelin-specific functional antagonist in rats in vivo, blocking the hypotensive effects of apelin-13 (Lee et al., 2005). However, no in vitro confirmation of its antagonist activity has been made, and there are conflicting reports as to the effect of alanine substitution at this position on the activity of apelins at the apelin receptor expressed in cell lines. De Mota et al. (2000) found that substitution of the C-terminal phenylalanine of (Pyr1)apelin-13 for alanine abolished the agonist activity of this peptide at the rat apelin receptor expressed in CHO cells. However, two studies showed that substitution of the C-terminal phenylalanine for alanine in (Pyr1)apelin-13 or apelin-13 had little effect on their affinity or agonist activity at the human apelin receptor expressed in human embryonic kidney 293 cells (Fan et al., 2003; Medhurst et al., 2003). Apelin-13(F13A) has been demonstrated to have comparable affinity and agonist activity at the native apelin receptor in human cardiovascular tissues in vitro (Pitkin et al., 2009), indicating that this peptide is a full agonist in man. The disparity of these reports may reflect species differences in the structure activity relationships of the human and rat apelin receptor.
IX. Radiolabeled Ligands
A number of radiolabeled ligands for the apelin receptor have been synthesized based on the structure of the endogenous ligands (Table 4), but most are based on (Pyr1)apelin-13. [125I](Pyr1)apelin-13 binds to receptors in human left ventricle with a KD of 0.35 nM. It associates rapidly, with a half-time for association of 6 min, and dissociates with a half-time for dissociation of 53 min (Katugampola et al., 2001). It is noteworthy that analogs of this radioligand have been made by others with modifications at position 75. Hosoya et al. (2000) replaced the methionine at this position with norleucine to prevent possible oxidation during the radiolabeling process, because they found oxidized (Pyr1)apelin-13 to be inactive. The resulting radioligand, [125I](Pyr1)[Nle75,Tyr77]apelin-13, is commercially available. Medhurst et al. (2003) oxidized the methionine at this position, because the unoxidized form of the radioligand was very unstable.
Radiolabelled ligands of the apelin receptor
X. Physiological Roles
Since the discovery of apelin as the endogenous ligand for APJ in 1998, a number of physiological roles for the receptor have emerged, including regulation of cardiovascular function, fluid homeostasis, and the adipoinsular axis.
A. Cardiovascular Roles
An important role for the apelin system in cardiovascular regulation is supported by cardiovascular phenotypes in both the apelin receptor and apelin peptide knockout mouse (see section XIII). Apelin modulates vascular tone in vivo, causing a reduction in blood pressure when infused into rats (Lee et al., 2000; Reaux et al., 2001; Tatemoto et al., 2001; Cheng et al., 2003; Ishida et al., 2004; El Messari et al., 2004; Lee et al., 2005; Mitra et al., 2006; Iturrioz et al., 2007) and vasodilation of resistance vessels when infused into the human forearm (Japp et al., 2008), both responses mediated primarily by nitric oxide. In vitro, apelin causes vasodilation of human splanchnic artery, largely via a nitric oxide-dependent mechanism (Salcedo et al., 2007). Apelin also causes vasoconstriction of human saphenous vein (Katugampola et al., 2001) and mammary artery (Maguire et al., 2009) in vitro by a direct action on vascular smooth muscle. These data support a role for the apelin system in modulation of vascular tone, where apelin released from endothelial cells would act on apelin receptors on the endothelium to cause vasodilation or on underlying smooth muscle cells to cause vasoconstriction.
Apelin also modulates cardiac function. Apelin peptides have positive inotropic effects in rats (Berry et al., 2004; Jia et al., 2006; Atluri et al., 2007) and mice (Ashley et al., 2005) in vivo. In vitro studies have demonstrated that apelins are potent positive inotropic agents by a direct action on cardiac tissue in rat (Szokodi et al., 2002; Dai et al., 2006; Farkasfalvi et al., 2007) and human (Maguire et al., 2009). It is noteworthy that apelins are the most potent endogenous inotropic agents yet reported in isolated cardiac tissue, with EC50 values of 40 to 125 pM in human tissue (Maguire et al., 2009) and 33 pM in rat tissue (Szokodi et al., 2002).
In addition, apelin is a potent angiogenic factor (Kasai et al., 2004; Cox et al., 2006) and mitogen of endothelial cells (Kasai et al., 2004; Masri et al., 2004) and vascular smooth muscle cells (Li et al., 2008). Apelin and its receptor are required for normal cardiovascular development. In the frog, apelin and its receptor are required for normal heart morphology (Inui et al., 2006) and for angiogenesis of intersomitic vessels (Cox et al., 2006; Kälin et al., 2007). In the zebrafish, either an excess or deficit of apelin impairs the migration of anterior lateral plate mesoderm cells during gastrulation, such that the myocardial progenitos cells are not in the correct position to receive inductive signals required for differentiation, resulting in a deficiency of the heart (Scott et al., 2007; Zeng et al., 2007, Nornes et al., 2009). Mice lacking the apelin gene show retardation of retinal vascular development (Kasai et al., 2008) and narrow blood vessels in intersomitic vessels during embryogenesis (Kidoya et al., 2008), whereas APLNR(−/−) mice have cardiac developmental defects (Charo et al., 2009). It has recently been shown that apelin acts downstream of Cripto (official gene name tdgf-1) to specify murine embryonic stem cells toward the cardiac lineage (D'Aniello et al., 2009).
B. Fluid Homeostasis
The colocalization of apelin and its receptor with vasopressin in magnocellular neurons of the SON and PVN of the hypothalamus triggered investigation of their role in fluid homeostasis. Apelin, given to mice by intracerebroventricular injection, inhibits vasopressin neuron activity and vasopressin release, decreasing plasma vasopressin concentration and increasing diuresis (De Mota et al., 2004). Dehydration increases apelin (Reaux-Le Goazigo et al., 2004) and apelin receptor (O'Carroll and Lolait, 2003) expression and decreases vasopressin expression in rat magnocellular neurons (Reaux-Le Goazigo et al., 2004). In addition to its central effects, apelin has direct actions on the microvasculature of the kidney (Hus-Citharel et al., 2008). APLNR(−/−) mice have abnormal fluid homeostasis and altered responses to osmotic stress (Roberts et al., 2009; Roberts et al., 2010) (see section XIII). Osmotic stimuli have been show to exert opposing effects on plasma apelin and vasopressin in man; increased plasma osmolality increases plasma vasopressin and decreases plasma apelin, and vice versa (Azizi et al., 2008). However, the role of the apelin system in drinking behavior is unclear.
C. Adipoinsular Axis
Apelin is expressed and released by cultured adipocytes, identifying it as a novel adipokine (Boucher et al., 2005), and adipose tissue is a possible source of plasma apelin. Apelin expression in adipose tissue is regulated by factors such as fasting and refeeding (Boucher et al., 2005), insulin (Boucher et al., 2005; Wei et al., 2005), hypoxia (Glassford et al., 2007; Kunduzova et al., 2008), growth hormone (Kralisch et al., 2007), tumor necrosis factor α (Daviaud et al., 2006), and peroxisome proliferator-activated receptor γ coactivator 1α (Mazzucotelli et al., 2008). Whereas insulin stimulates adipose apelin expression (Boucher et al., 2005; Wei et al., 2005), apelin inhibits insulin secretion (Sörhede Winzell et al., 2005), presenting an interesting interaction between the two systems.
There is evidence for a role for apelin regulation of adiposity, peripherally administered apelin causing no change in food intake (Sunter et al., 2003; Higuchi et al., 2007) but decreasing adiposity, possibly by up-regulating uncoupling proteins and increasing energy expenditure (Higuchi et al., 2007). However, investigation of the role of central apelin on food intake and body weight has yielded disparate results (Taheri et al., 2002; O'Shea et al., 2003; Sunter et al., 2003; Valle et al., 2008). Apelin may also be involved in vascularization of adipose tissue (Kunduzova et al., 2008).
D. Other Roles
There is evidence for a role for the apelin system in gastrointestinal function, with expression of apelin and its receptor in several areas of the gastrointestinal tract during embryonic development and adulthood in rodents (Wang et al., 2004; Susaki et al., 2005; Wang et al., 2009). Proposed functions include stimulation of gastric cell proliferation and cholecystokinin secretion (Wang et al., 2004) and regulation of gastric acid secretion (Lambrecht et al., 2006). Apelin is expressed in osteoblasts and may have a role in stimulating proliferation and suppressing apoptosis in this cell type (Xie et al., 2006; Tang et al., 2007; Xie et al., 2007). Apelin has also been shown to inhibit apoptosis in neurons in vitro (Zeng et al., 2010) and to have antinociceptive effects (Xu et al., 2009). It has also been suggested that apelin may have an immunomodulatory role (Habata et al., 1999; Horiuchi et al., 2003; Leeper et al., 2009).
XI. Pathophysiological Roles
A. Cardiovascular Disease
A growing body of evidence links the apelin system with cardiovascular disease. A number of studies have noted changes in human plasma apelin levels in conditions of cardiac dysfunction, with sometimes conflicting results and reporting a very wide range of concentrations (20–4000 pg/ml) depending on the assay used and sample preparation. Overall, plasma apelin seems to rise in early heart failure (Chen et al., 2003) but to normalize or decrease in later stages (Chen et al., 2003; Chong et al., 2006; Miettinen et al., 2007). Treatments such as chronic resynchronization therapy and placement of a left ventricular assist device increase apelin peptide levels (Chen et al., 2003; Francia et al., 2007). Apelin receptor mRNA was found to be decreased in left ventricle from patients with idiopathic dilated cardiomyopathy (Földes et al., 2003). It is noteworthy that the apelin receptor is the most significantly up-regulated gene of >12,000 after mechanical offloading of failing myocardium (Chen et al., 2003). These data suggest that apelin may be up-regulated in early disease as a compensatory mechanism, increasing the force of contraction of the heart. Both apelin and its receptor may then be down-regulated later in disease when its positive inotropic effects would be detrimental, owing to the resultant increase in oxygen demand.
A beneficial role of apelin in heart failure is supported by data from animal models. Cardiac apelin is increased in response to hypoxia via hypoxia-inducible factor-1 (Ronkainen et al., 2007), and both apelin and its receptor are up-regulated in ischemic heart failure in rats (Atluri et al., 2007; Sheikh et al., 2008). Apelin protects against cardiac ischemia reperfusion injury (Simpkin et al., 2007; Zeng et al., 2009) and inhibits glucose deprivation induced cardiomyocyte apoptosis (Zhang et al., 2009c), suggesting that apelin may be up-regulated in response to cardiac hypoxia to protect the tissue from ischemia reperfusion injury.
The apelin system has recently been implicated in the pathogenesis of atherosclerosis by evidence from transgenic mouse models (see section XIII), although there are conflicting reports as to whether apelin signaling is beneficial or detrimental in these models. In man, apelin peptide is up-regulated in atherosclerotic coronary artery (Pitkin et al., 2009), and both apelin and its receptor are up-regulated in aortic valve stenosis, a process that displays some hallmarks of atherosclerosis (Peltonen et al., 2009), supporting an involvement of the apelin system in human atherosclerosis. Apelin also prevents aortic aneurism formation in mice (Leeper et al., 2009).
A role for the apelin system in hypertension has also been proposed. Spontaneously hypertensive rats have decreased cardiovascular apelin receptor and apelin mRNA and protein compared with control rats. Exercise training or long-term all-trans retinoic acid treatment improves hypertension in line with an increase in apelin receptor and apelin expression (Zhong et al., 2005; Zhang et al., 2006), consistent with the predominantly hypotensive effect seen in healthy individuals and animals. Spontaneously hypertensive rats have increased apelin expression in the rostral ventrolateral medulla, and overexpression of apelin in this area causes chronic hypertension in normotensive rats (Zhang et al., 2009b). Some single-nucleotide polymorphisms (SNPs) in the gene encoding the apelin receptor (APLNR) and apelin peptide (APLN) are associated with hypertension (see section XII). There is also evidence for a role in pulmonary hypertension (Andersen et al., 2009; Falcão-Pires et al., 2009) and portal hypertension (Tiani et al., 2009). It is noteworthy that apelin is able to cause vasodilation by a prostanoid-dependent mechanism in blood vessels from patients with atherosclerotic heart disease (Maguire et al., 2009). This suggests that apelin may have beneficial vasodilatory effects even in patient groups that display a degree of endothelial dysfunction.
The apelin system is proposed to have a role in pathophysiological angiogenesis, contributing to portosystemic collateralization and splanchnic neovascularization in portal hypertensive rats (Tiani et al., 2009), neoangiogenesis in liver cirrhosis (Principe et al., 2008), and tumor neoangiogenesis (Kälin et al., 2007; Sorli et al., 2007). However, angiogenic effects (Eyries et al., 2008) and induction of larger nonleaky vessels by apelin (Kidoya et al., 2010) appear to be beneficial in recovery from ischemia.
B. Obesity and Diabetes
Plasma apelin is positively correlated with body mass index in humans (Heinonen et al., 2005), and increased plasma and adipose tissue apelin expression in obese patients can be reversed by hypocaloric diet associated with weight loss (Castan-Laurell et al., 2008). However in mouse models of obesity, insulin level, rather than adiposity, is the major determinant of increased apelin expression (Boucher et al., 2005). This suggests that apelin expression may be increased in response to hyperinsulinemia associated with obesity rather than to obesity itself, although investigation of the effect of type 2 diabetes mellitus on plasma apelin has yielded disparate results (Li et al., 2006; Erdem et al., 2008).
The up-regulation of apelin in obesity may be beneficial in metabolic syndrome because peripheral apelin treatment improves glucose tolerance and enhances glucose utilization in obese insulin-resistant mice (Dray et al., 2008); conversely, apelin knockout mice have reduced insulin sensitivity (Yue et al., 2010). Apelin also decreases body adiposity, without altering food intake, in obese mice by increasing the expression of uncoupling proteins and energy expenditure (Higuchi et al., 2007). The up-regulation of apelin in obesity may also have beneficial cardiovascular effects; limiting hypertension and increasing cardiac contractility (see section XI.A).
C. Other Roles
Roles for the apelin system in cardiovascular disease and obesity, conditions associated with inflammation, have raised interest in a possible connection between apelin and inflammation. Apelin expression is induced by inflammatory mediators, such as tumor necrosis factor α (Daviaud et al., 2006), interleukin-6, and interferon-γ (Han et al., 2008) and plasma apelin levels correlate with markers of inflammation (Malyszko et al., 2008). It is possible that up-regulation of apelin in response to inflammation is a compensatory mechanism to limit the onset of metabolic disorders, such as insulin resistance and cardiovascular dysfunction.
The apelin system has also been suggested to be involved in HIV infection of CD4 positive cells (Choe et al., 1998; Edinger et al., 1998; Zhang et al., 1998), gastrointestinal disorders (Han et al., 2007), liver cirrhosis (Principe et al., 2008), and obstructive sleep apnea (Henley et al., 2009).
XII. Single-Nucleotide Polymorphisms
A number of SNPs within the gene encoding the apelin receptor (APLNR) are associated with cardiovascular disease risk. The G212A variant is associated with slower heart failure progression (Sarzani et al., 2007), whereas a SNP in the 5′ flanking region is associated with susceptibility to brain infarction (Hata et al., 2007). Three SNPs within APLNR, and two in the gene encoding apelin (APLN), have been found to be associated with hypertension (Li et al., 2009), and a number of SNPs in APLNR were associated with blood pressure responses to dietary sodium intervention (Zhao et al., 2010) in Han Chinese. In addition, one SNP in APLN is associated with fasting plasma glucose levels in Han Chinese male subjects (Zhang et al., 2009a).
XIII. Knockout Mouse Models
Mice lacking the gene encoding the apelin receptor [APLNR(−/−)] are not born in the expected Mendelian ratio (Charo et al., 2009). They have normal baseline blood pressure and heart rate but lack the hypotensive response to apelin seen in wild-type mice. In addition, angiotensin II produces a greater pressor effect in apelin receptor-deficient mice compared with wild-type mice, and double mutant mice, lacking both the apelin receptor and AT1a, have a higher baseline blood pressure than mice lacking AT1a only (Ishida et al., 2004). These data suggest a role for the apelin receptor in blood pressure regulation, particularly in opposing the effects of angiotensin II. APLNR(−/−) mice show cardiac developmental defects and modest decrements in contractile function under basal conditions, but under exercise stress, they have strikingly reduced exercise capacity (Charo et al., 2009), supporting a role for the apelin receptor in cardiac function.
It is noteworthy that apelin receptor-deficient mice on an ApoE(−/−) background have a reduced atherosclerotic burden compared with APLNR(+/+) ApoE(−/−) mice (Hashimoto et al., 2007). Likewise, apelin knockout mice show decreased neointima formation in a carotid ligation model (Kojima et al., 2010), implying that activation of the apelin receptor is detrimental in atherosclerosis. This contrasts with the findings of Chun et al. (2008), where apelin signaling opposed angiotensin II-induced atherosclerosis in ApoE(−/−)mice. APLNR(−/−) mice have been shown to have abnormal fluid homeostasis, drinking significantly less water, showing an inability to concentrate their urine to the same extent as wild-type mice (Roberts et al., 2009) and having altered responses to osmotic stress (Roberts et al., 2010), supporting a role for the apelin receptor in fluid homeostasis.
Apelin knockout mice are viable, fertile, appear healthy, and have normal body weight, normal water and food intake, and normal heart rates and heart morphology (Kuba et al., 2007; Charo et al., 2009). However, similar to APLNR(−/−) mice, these mice have slightly impaired cardiac function under basal conditions and markedly decreased exercise capacity (Charo et al., 2009). They also develop progressive impairment of cardiac contractility with age and present with more severely impaired heart function in response to pressure overload than wild-type mice (Kuba et al., 2007). This indicates that apelin is important in the maintenance of cardiac function in conditions of exercise stress, pressure overload and aging. Apelin-deficient mice also show impaired retinal vascularization and ocular development (Kasai et al., 2008) and narrow blood vessels in intersomitic vessels during embryogenesis (Kidoya et al., 2008). In addition, apelin knockout mice have diminished insulin sensitivity (Yue et al., 2010).
XIV. Conclusion
In conclusion, since the pairing of the apelin receptor with its endogenous ligand apelin in 1998, a number of physiological and pathophysiological roles have emerged for this receptor, including modulation of the cardiovascular system, fluid homeostasis, and the adipoinsular axis. Development of additional pharmacological tools, particularly a competitive antagonist, and further investigation of the roles of endogenous apelin will aid validation of this system as a drug target in a range of conditions.
Acknowledgments.
This work was supported by the British Heart Foundation [Grants PS/02/001, PG/09/050/27734, FS/06/017].
Footnotes
This article is available online at http://pharmrev.aspetjournals.org.
doi:10.1124/pr.110.002949.
-
↵1 Abbreviations:
- GPCR
- G-protein-coupled receptor
- IUPHAR
- International Union of Pharmacology
- AT
- angiotensin
- CHO
- Chinese hamster ovary
- ACE
- angiotensin-converting enzyme
- CNS
- central nervous system
- SON
- supraoptic nucleus
- PVN
- paraventricular nucleus
- SNP
- single-nucleotide polymorphism
- pKD
- the negative log 10 of the equilibrium dissociation constant of a ligand determined directly in a binding assay using a labeled form of the ligand.
- Copyright © 2010 by The American Society for Pharmacology and Experimental Therapeutics
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- I. Introduction
- II. The Apelin Receptor: Recommendations for Nomenclature
- III. Receptor Structure
- IV. Endogenous Agonists
- V. Receptor Distribution
- VI. Apelin Peptide Distribution
- VII. Synthetic Agonists
- VIII. Antagonists
- IX. Radiolabeled Ligands
- X. Physiological Roles
- XI. Pathophysiological Roles
- XII. Single-Nucleotide Polymorphisms
- XIII. Knockout Mouse Models
- XIV. Conclusion
- Acknowledgments.
- Footnotes
- References
- Figures & Data
- Info & Metrics
- eLetters