Article Text

Abstract
BACKGROUND Sufficient ATP concentrations maintain physiological processes and protect tissue from hypoxic damage. With decreasing oxygen concentration, ATP synthesis relies increasingly on the presence of phosphocreatine.
AIM The effect of exogenously applied creatine on phosphocreatine and ATP concentrations was studied under control and anoxic conditions.
METHODS Pregnant mice were fed orally with creatine monohydrate (2 g/kg body weight/day). Brainstem slices from these mice pups were compared with those from pups of non-creatine supplemented pregnant mice. Measurements were performed under normoxic and anoxic conditions. In addition, brainstem slices from non-creatine treated mice pups were incubated for 3 hours in control artificial cerebrospinal fluid (CSF) (n = 10) or in artificial CSF containing 200 μM creatine (n = 10). ATP and phosphocreatine contents were determined enzymatically in single brainstem slices.
RESULTS ATP concentrations were in the same range in all preparations. However, there was a significant increase of phosphocreatine in the brainstems from pups of creatine fed mice when compared with the brainstems of pups from non-creatine treated mice or in non-incubated brainstems of control animals. After 30 minutes anoxia, ATP as well as phosphocreatine concentrations remained significantly higher in creatine pretreated slices compared with controls.
CONCLUSION The data indicate that exogenous application of creatine is effective in neuroprotection.
- ATP
- creatine
- phosphocreatine
- neuroprotection
Statistics from Altmetric.com
The brain is one of the most metabolically active organs and depends on a continuous supply of energy to stabilise ionic homeostasis, energy consuming biochemical reactions, and physiological processes. The primary energy source is ATP generated via oxidative phosphorylation of NADH within mitochondria,1 ,2 which requires a sufficient oxygen supply. Under hypoxic conditions, ATP is primarily supplemented by the phosphocreatine pool, before anaerobic glycolysis is activated, leading to increased production of lactate and H+.1 ,3 ,4
The hypoxia induced fall of intracellular ATP results in multiple dysfunctions of neurones—for example, failure in Ca2+regulating mechanisms leading to an uncontrolled rise of intracellular Ca2+ and subsequent cell damage.4 This explains why survival time of immature rats under anoxic conditions is directly correlated with the preservation of brain energy stores for regulating ion homeostasis.5
Previous investigations demonstrated that the exogenous application of creatine increases the latency of anoxic depolarisation of neurones,6 thus prolonging cellular survival and stabilising synaptic transmission within the central nervous system.7 ,8 Previously, we described the application of prophylactic supplementation of creatine and the effects on the synaptic activity of the central respiratory network under anoxic conditions.9 Here, we present the changes in phosphocreatine and ATP concentrations within the brainstem in more detail using the identical experimental design.
The hypothesis that exogenous supplementation of creatine stabilises ATP concentrations during anoxic conditions through raised phosphocreatine pools was analysed in an in vitro brainstem preparation of neonatal mice.
Experimental setup
ATP and phosphocreatine contents in brainstem slices obtained from neonatal animals (P2) (n = 60) were determined enzymatically.
In a first experimental setup, pregnant mice were fed orally with creatine monohydrate (2 g/kg body weight/day) (Sigma, Deisenhofen, Germany) throughout pregnancy (20 ± 1 day). Brainstem slices (600 μM thick) from these mice pups (n = 10) were compared with those from pups born to non-creatine supplemented pregnant mice (n = 10). Thereafter, mice pups from controls (n = 10) as well as from creatine treated animals (n = 10) were exposed to anoxia for 30 minutes.
In a second series of experiments, brainstem slices from non-creatine treated mice pups were incubated for three hours in control artificial cerebrospinal fluid (CSF) (n = 10) or in artificial CSF containing 200 μM creatine (n = 10).
Preparation
Technical details of the transverse brainstem slice preparations have been described previously.10 The brainstem was isolated in ice cold artificial CSF and secured in a vibratome with its rostral end directed upwards. Slices were sectioned serially until the rostral boundary of the so called pre-Bötzinger complex became visible. This region is crucial for the generation of the respiratory rhythm.11 The transverse slice (600 μm thickness) containing the pre-Bötzinger complex is recognisable by cytoarchitectonic landmarks, such as the appearance of the inferior olive and the hypoglossal nucleus, and the disappearance of the facial nucleus. The next slice containing the pre-Bötzinger complex was transferred into a recording chamber for electrophysiological measurements or was immediately homogenised for biochemical analysis.
The preparation was superfused by artificial CSF (flow rate 10 ml/min) at 29°C and was continuously gassed with carbogen (95% O2 and 5% CO2). The artificial CSF contained 128 mM NaCl, 3 mM KCl, 1.5 mM CaCl2, 1 mM MgSO4, 24 mM NaHCO3, 0.5 mM NaH2PO4, and 30 mM D-glucose, and was equilibrated with carbogen at 27°C to pH 7.4. Anoxia was induced by gassing the artificial CSF with 95% N2 and 5% CO2 over a period of 30 minutes.
Biochemical analysis
The rationale of the biochemical analysis was to determine the ATP, ADP, phosphocreatine, and creatine contents of brainstem slices to obtain information about the creatine kinase regulated transfer of phosphate from phosphocreatine to ADP, which results in de novo production of ATP.
Immediately after withdrawal from the recording chamber all slices were homogenised by ultrasound in 250 μl 8% perchloric acid and centrifuged for 15 minutes at 15 500g. The supernatant fluid was neutralised by the addition of KHCO3and recentrifuged. ATP and phosphocreatine was measured enzymatically by adding 30 μl of a 100 mM triethanolamin hydrochloride stem solution (containing 7 mM MgCl2, 2 nmol (4 μl of 0.5 mM) NADP solution, 12 nmol ADP (4 μl of 3 mM) solution, 8 nmol glucose (4 μl of 2 mM), and 1 μl G6P-DH) to 100 μl of the homogenised brainstem slices.12 The enzymatic reaction was started with 0.5 μl hexokinase. ATP was measured using a photometer with light absorption at 366 nm. After the end of the biochemical reaction, 20 μl creatine kinase was added to measure phosphocreatine contents. All ATP and phosphocreatine values are given in μmol/g wet weight. Significant differences were determined by the Student's t test.
Results
Using enzymatic tests, ATP values did not differ significantly between normal brainstem slices and those from mice pups of animals supplemented with creatine during pregnancy. Table 1 gives the ATP concentrations. There were also no significant changes in ATP concentrations when slices were incubated for three hours in artificial CSF or artificial CSF containing 200 μM creatine: mean (SE) ATP concentrations were 7.2 (0.2) μmol/g in control slices and 7.5 (0.2) μmol/g in slices incubated in creatine supplemented artificial CSF. Both sets of values were significantly lower than in the group receiving oral supplementation via the mother animals.
ATP concentrations for controls and supplemented pups under normoxic and anoxic conditions
After 30 minutes anoxia, there was a 54% decrease in ATP values in neonatal control slices, whereas there was only an 8% decrease in ATP values in the creatine pretreated slices (p < 0.05) (fig 1).

Anoxic changes of ATP concentrations in brainstem slices of neonatal animals. Slices were taken either from untreated animals (“controls”) or were incubated for three hours in 200 μM creatine (“incubation”) or alternatively taken from pups that were pretreated by feeding the pregnant mother animals with creatine (“nutrition”) (n = 10 each group). Anoxia lasted for 30 minutes.
The mean (SE) phosphocreatine concentration was 0.8 (0.1) μmol/g in control slices and thus significantly different from the values found in slices obtained from pups of mother animals fed with creatine (mean, 1.9; SE, 0.1 μmol/g; p < 0.05). After three hours creatine incubation, the mean (SE) concentration of phosphocreatinine increased from 2.4 (0.3) μmol/g to 3.2 (0.4) μmol/g (p < 0.05; fig 2).
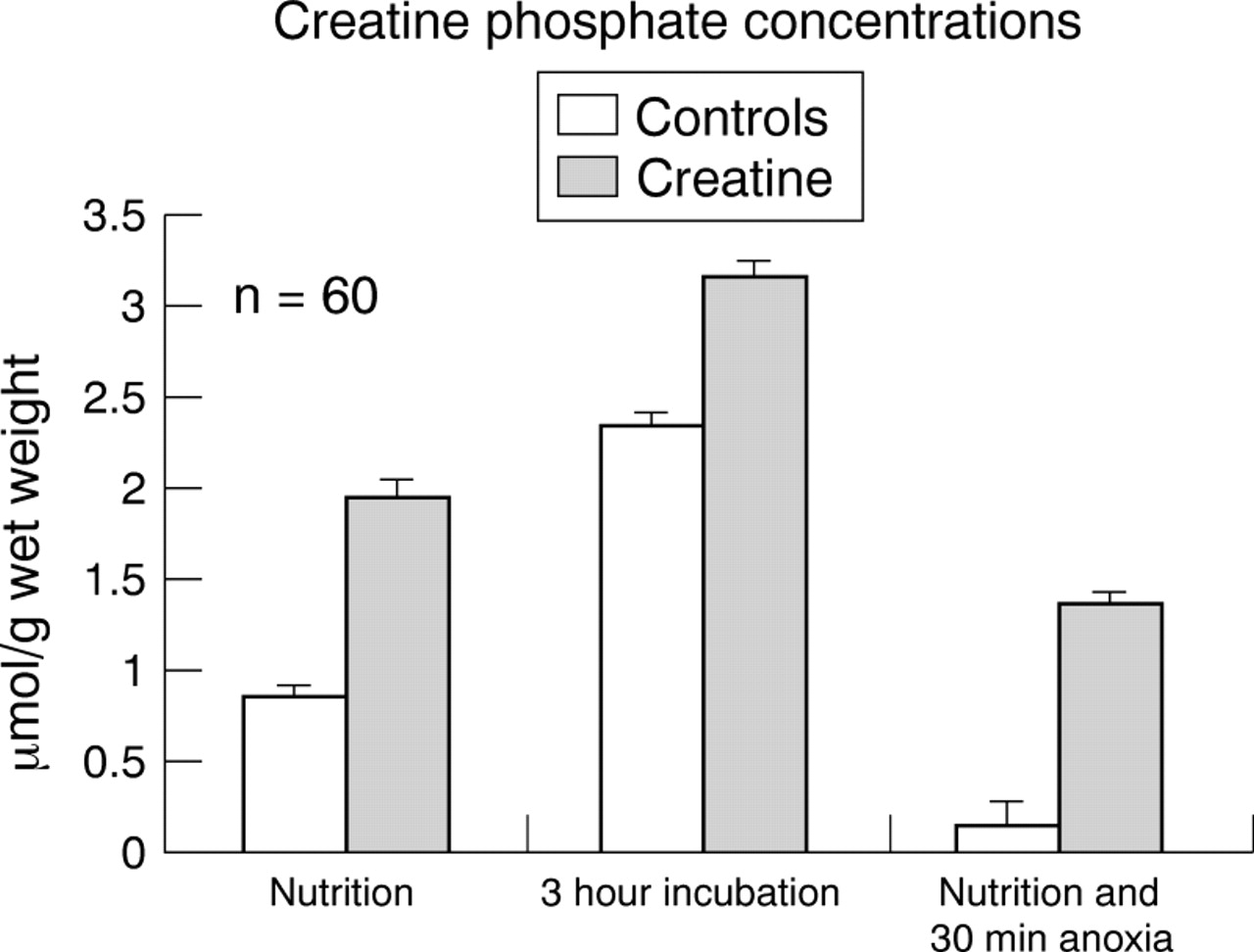
Anoxic changes of phosphocreatine concentrations in brainstem slices of neonatal animals. Slices were taken either from untreated animals (“controls”) or were incubated for three hours in 200 μM creatine (“incubation”), or alternatively taken from pups that were pretreated by feeding the pregnant mother animals with creatine (“nutrition”) (n = 10 each group). Anoxia lasted for 30 minutes.
The most pronounced preservation of phosphocreatine values was seen in incubated slices. After 30 minutes anoxia, the mean (SE) phosphocreatine concentration was 1.4 (0.1) μmol/g in creatine pretreated slices, whereas the phosphocreatine was almost completely depleted (mean, 0.1; SE, 0.1 μmol/g) in control slices (p < 0.05; fig 2).
Discussion
Using biochemical analysis, we found that oral treatment of pregnant mother animals with creatine or incubation of brainstem slices in creatine containing artificial CSF increased phosphocreatine concentrations and stabilised ATP values during anoxia in brain tissue of neonatal mice. ATP concentrations were lower, but not significantly different, in slices incubated for three hours either in normal or in creatine containing artificial CSF. However, phosphocreatine concentrations were significantly higher after slices were incubated in creatine containing artificial CSF or after they were incubated in artificial CSF without creatine. ATP concentrations were decreased in both groups after incubation for three hours. We interpret this finding to be the result of the low metabolic activity of slices held at a temperature of 27°C.
After 30 minutes anoxia, ATP was significantly reduced (54%) in control brainstem slices, while there was only a slight decrease of ATP values (8%) in slices that were pretreated with creatine. In addition, phosphocreatine was only slightly decreased in creatine pretreated slices, whereas it was almost completely depleted after 30 minutes anoxia in control tissue.
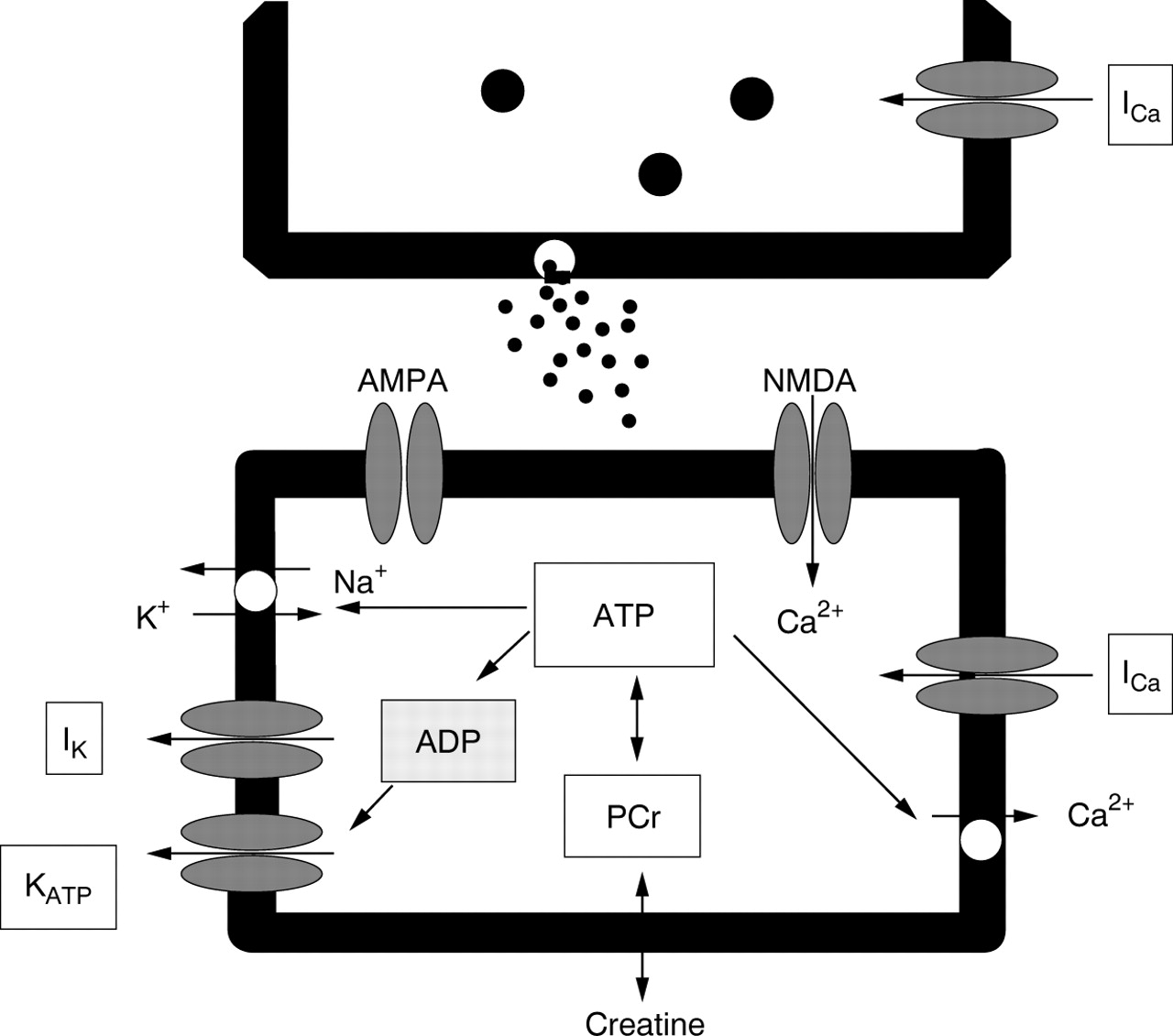
Physiological processes and life rescuing mechanisms—for example, the highly energy demanding hypoxic augmentation of respiratory activity,9 depend on a sufficient energy supply and hence a sufficient phosphocreatine pool. Under normal conditions, phosphocreatine decreases rapidly to 40–50% during hypoxic conditions, which is accompanied by a comparable fall in ATP values, indicating failure of ATP synthesis.13 ,14 Such energy depletion is correlated with the onset of hypoxic depression of the central respiratory network, as Pierard and co-workers15have shown by magnetic resonance spectroscopy. This neural response can easily be explained by the blockade of synaptic interaction within the network.15 Inhibitory synaptic transmission seems to be more sensitive to hypoxic stress,16 which leads to the danger of excitotoxicity of excitatory amino acids. Excessive excitation could lead to the massive influx of cations, which can lead to severe damage to neurones. This is particularly threatening to neonatal animals in which compensatory rescue mechanisms are underpowered.17 The immature isoform of the NMDA (N-methyl-D-aspartate) receptor reveals a higher open probability, and its deactivation is slower than that of adult NMDA receptors, resulting in an enhanced Ca2+ influx.17 This may be potentiated by a Ca2+ influx through depolarisation activated L-type Ca2+ channels.18 Immature brain tissue might not be able to cope with such a Ca2+overload. The reason is that creatine kinase is three to six times less active in immature than in mature animals, which results in a limited energy pool.19 ,20 The responsible cellular mechanisms for this observation are summarised in the scheme shown in fig3.
{kind=link}
{kind=link}
{kind=link}
Schematic illustration of the most essential consequences of ATP stabilisation in neurones. ATP turnover is enhanced in all cases of hypoxic augmentation of neuronal activity. Increased ADP production leads to activation of K+ outward currents (IK) preferentially flowing through ADP/ATP regulated KATP channels, and the membrane potential of presynaptic and postsynaptic elements remains negative. ATPases regulate electrogenic 3Na+/2K+ pumping, they control active Ca2+ pumping into intracellular organelles, and indirectly ensure effective Na+ coupled Ca2+transportation across the plasma membrane. Such ATP dependent stabilisation of the membrane potential and cytosolic Ca2+concentration also protects presynaptic Ca2+ inward currents that regulate release of the neurotransmitter glutamate. Unprotected hypoxic enhancement of specific and non-specific glutamate release would activate Ca2+ permeable NMDA-type (N-methyl-D-aspartate) glutamate receptors, apart from the normally activated Na+ permeable AMPA-type glutamate receptors. Such enhanced Ca2+ influx and depressed Ca2+sequestration would lead to further membrane depolarisation and an excitotoxic rise in intracellular Ca2+ concentration. Thus, stabilisation of the ATP pool protects synaptic transmission between neurones even during severe hypoxia. AMPA, α-amino-3-hydroxy-5-methylinoxazol- propion-acid.
In immature as well as in mature animals, cell death does not occur as long as the ATP content remains above 25% of control.21 This has important clinical implications, because stabilisation of a sufficient ATP content could be neuroprotective. ATP is needed for energy consuming processes such as 3Na+/2K+ and Ca2+pumping,22 which are the key processes for ionic homeostasis determining neuronal excitability and synaptic transmission. Inhibitory and excitatory synaptic transmission should also be protected by increasing ATP supply. This is consistent with the finding that preincubation of neocortical cells with creatine also had a pronounced protective effect against hypoxic depression of synaptic transmission.7 ,23 ,24
In conclusion, our data demonstrate that hypoxic energy failure in neonatal mice can be prevented by exogenous creatine applied before the hypoxic event. From this observation, the working hypothesis is derived that creatine might also be neuroprotective in humans.