


Abstract
This study investigated the role of a multispecific organic anion transporter, Oatp1a4/Slco1a4, in drug transport across the blood-brain barrier. In vitro transport studies using human embryonic kidney 293 cells expressing mouse Oatp1a4 identified the following compounds as Oatp1a4 substrates: pitavastatin (Km = 8.3 μM), rosuvastatin (Km = 12 μM), pravastatin, taurocholate (Km = 40 μM), digoxin, ochratoxin A, and [d-penicillamine2,5]-enkephalin. Double immunohistochemical staining of Oatp1a4 with P-glycoprotein (P-gp) or glial fibrillary acidic protein demonstrated that Oatp1a4 signals colocalized with P-gp signals partly but not with glial fibrillary acidic protein, suggesting that Oatp1a4 is expressed in both the luminal and the abluminal membranes of mouse brain capillary endothelial cells. The brain-to-blood transport of pitavastatin, rosuvastatin, pravastatin, and taurocholate after microinjection into the cerebral cortex was significantly decreased in Oatp1a4(−/−) mice compared with that in wild-type mice. The blood-to-brain transport of pitavastatin, rosuvastatin, taurocholate, and ochratoxin A, determined by in situ brain perfusion, was significantly lower in Oatp1a4(−/−) mice than in wild-type mice, whereas transport of pravastatin and [d-penicillamine2,5]-enkephalin was unchanged. The blood-to-brain transport of digoxin was significantly lower in Oatp1a4(−/−) mice than in wild-type mice only when P-gp was inhibited by N-(4-[2-(1,2,3,4-tetrahydro-6,7-dimethoxy-2-isoquinolinyl)ethyl]-phenyl)-9,10-dihydro-5-methoxy-9-oxo-4-acridine carboxamide (GF120918). Taken together, these results show that Oatp1a4 can mediate the brain-to-blood and blood-to-brain transport of its substrate drugs across the blood-brain barrier. The brain-to-plasma ratio of taurocholate, pitavastatin, and rosuvastatin was close to the capillary volume in wild-type mice, and it was not affected by Oatp1a4 dysfunction. Whether Oatp1a4 can deliver drugs from the blood to the brain remains controversial.
Drugs have to cross the blood-brain barrier (BBB) before reaching their target in the central nervous system. The BBB consists of a tight monolayer of brain capillary endothelial cells that are characterized by highly developed tight junctions between adjacent endothelial cells and a paucity of fenestra and pinocytotic vesicles, thereby limiting paracellular transport across the endothelial cells. In addition, xenobiotic transporters act as a barrier by actively effluxing drugs into the blood circulation, thus limiting the transcellular pathway of drugs and other xenobiotic compounds (Kusuhara and Sugiyama, 2005; Hermann and Bassetti, 2007; Ohtsuki and Terasaki, 2007). Indeed, dysfunction of ATP-binding cassette transporters (such as P-gp, BCRP, and multidrug resistance-associated protein 4) results in a significant increase in brain penetration by their substrate drugs (Fromm, 2004; Enokizono et al., 2007, 2008; Ose et al., 2009).
In vivo studies using microinjection into the cerebral cortex have shown that the elimination of diverse amphipathic and hydrophilic organic anions and nucleotide analogs from the cerebral cortex involves saturable mechanisms (Kusuhara and Sugiyama, 2005). Oatp1a4/Slco1a4, a homolog of the hepatic multispecific organic anion transporter Oatp1a1, was cloned from rat brain using homology screening (Noé et al., 1997). Oatp1a4 exhibits broad substrate specificities for amphipathic compounds and has a high affinity for the cardiac glycoside digoxin (Noé et al., 1997; Cattori et al., 2001). Oatp1a4 is abundantly expressed in the brain and liver. Immunohistochemical staining of rat brain has shown that the Oatp1a4 protein is present in both the luminal and the abluminal membranes of the BBB (Gao et al., 1999). Because Oatp1a4 mediates cellular uptake, this membrane localization suggests that it may mediate uptake from both the brain and the blood sides. Kinetic analyses of efflux across the BBB in rats after microinjection of Oatp1a4 inhibitors, such as taurocholate and digoxin, into the cerebral cortex have indicated that Oatp1a4 plays a major role in the uptake of amphipathic organic anions (including 17β-estradiol glucuronide, pitavastatin, and dehydroepiandrosterone sulfate) across the abluminal membrane and partly accounts for the uptake of pravastatin in the net efflux from the brain to the blood (Asaba et al., 2000; Sugiyama et al., 2001; Kikuchi et al., 2004). Mutual inhibition studies suggest that the uptake of BQ-123 and estrone sulfate is mediated by transporters distinct from Oatp1a4, although they are Oatp1a4 substrates (Kitazawa et al., 1998; Hosoya et al., 2000). Furthermore, it has been suggested that Oatp1a4 prevents accumulation of 24S-hydroxycholesterol, an endogenous liver X receptor agonist produced de novo from cholesterol in the brain, by facilitating its elimination across the BBB (Ohtsuki et al., 2007). In addition, in situ brain perfusion studies have suggested that Oatp1a4 also mediates the uptake of prostaglandin E1, morphine-6-glucuronide, and [d-penicillamine2,5]-enkephalin (DPDPE) from the blood (Dagenais et al., 2001; Bourasset et al., 2003; Taogoshi et al., 2005). Brain uptake of DPDPE in particular is limited by P-gp, and thus Oatp1a4-mediated uptake is more commonly observed in P-gp knockout mice and is characterized by inhibition by digoxin, verapamil, and fexofenadine (Dagenais et al., 2001).
The role of Oatp1a4 in drug transport at the BBB has only been investigated using Oatp1a4 inhibitors. Thus, the present study was undertaken to investigate the role of Oatp1a4 in drug transport across the BBB using Oatp1a4(−/−) mice. Mouse Oatp1a4 (mOatp1a4) has been cloned (van Montfoort et al., 2002) and found to be abundantly expressed in the liver, and to a lesser degree in the brain and kidney. Functional expression of mOatp1a4 in Xenopus laevis oocytes has demonstrated that mOatp1a4 efficiently transports digoxin and BQ-123, although the Km value of digoxin for mOatp1a4 is markedly greater than that for rat Oatp1a4 (rOatp1a4) (Noé et al., 1997; van Montfoort et al., 2002).
Materials and Methods
Chemicals.
[3H]Pitavastatin and unlabeled pitavastatin, [3H]rosuvastatin and unlabeled rosuvastatin, and [3H]pravastatin were kindly donated by the Kowa Company Ltd. (Tokyo, Japan), AstraZeneca (Cheshire, UK), and Sankyo (Tokyo, Japan), respectively. [14C]Sucrose and [3H]ochratoxin A were obtained from Moravek Biochemicals (Brea, CA). [14C]Carboxyl-inulin, [3H]taurocholate, [3H]digoxin, and [3H]DPDPE were obtained from PerkinElmer Life and Analytical Sciences (Waltham, MA). GF120918 was a gift from GlaxoSmithKline (Ware, UK). All other chemicals used in the experiments were of analytical grade.
Construction of a Stable Transfectant of mOatp1a4 in HEK293 Cells.
Based on the nucleotide sequence (GenBank accession no. AB031814), the cDNA encoding a full open reading frame of mOatp1a4 was cloned from mouse brain cDNA using polymerase chain reaction (PCR). The mOatp1a4 cDNA was subcloned into pcDNA3.1(+) (Invitrogen, Carlsbad, CA) and transfected into HEK293 cells using lipofection 2000 reagent (Invitrogen), according to the manufacturer's protocol. The transfectants were selected after culture in the presence of Zeocin (800 μg/ml; Invitrogen), and maintained at 37°C in 5% CO2 and 95% humidity in Dulbecco's modified Eagle's medium (high glucose; Invitrogen) supplemented with 10% fetal bovine serum, 1% antibiotic-antimycotic (Invitrogen), and Zeocin (200 μg/ml).
Determination of Cellular Uptake Using a Stable Transfectant of mOatp1a4.
Uptake was initiated by addition of the radiolabeled ligands to the incubation buffer after cells had been washed twice and preincubated at 37°C for 15 min in Krebs-Henseleit buffer. The Krebs-Henseleit buffer consisted of 23.8 mM NaHCO3, 118 mM NaCl, 4.83 mM KCl, 1.2 mM MgSO4, 0.96 mM KH2PO4, 1.53 mM CaCl2, 5 mM d-glucose, and 12.5 mM HEPES adjusted to pH 7.4. The uptake was terminated at specified times by the addition of buffer at 4°C, and cells were washed three times. The radioactivity associated with the cells and medium was determined in a liquid scintillation counter. The protein concentrations in aliquots of cell lysate were determined using the method of Lowry et al. (1951). Ligand uptake was assessed from the cell-to-medium ligand concentration ratio, calculated as the concentration of ligand associated with the cells divided by that in the medium.
Kinetic Analysis for the in Vitro Transport Study.
Mock- and mOatp1a4-HEK293 cells were incubated for 0.5 and 2 min to determine the cellular accumulation of pitavastatin and rosuvastatin and taurocholate, respectively, at 37°C. Specific uptake was determined by subtracting the uptake by mock-HEK293 cells from that by mOatp1a4-HEK293 cells. Kinetic parameters for the specific uptake by mOatp1a4 were obtained from eq. 1 (Michaelis-Menten equation):
 where v is the uptake rate of the substrate (picomoles per minute per milligram of protein), S is the substrate concentration in the medium (micromoles), Km is the Michaelis constant (micromoles), and Vmax is the maximum uptake rate (picomoles per minute per milligram of protein). The experimental data were fitted to the equation using nonlinear regression analysis with weighting as the reciprocal of the observed values, using the MULTI program; the damping Gauss-Newton method algorithm was used for curve fitting (Yamaoka et al., 1981).
where v is the uptake rate of the substrate (picomoles per minute per milligram of protein), S is the substrate concentration in the medium (micromoles), Km is the Michaelis constant (micromoles), and Vmax is the maximum uptake rate (picomoles per minute per milligram of protein). The experimental data were fitted to the equation using nonlinear regression analysis with weighting as the reciprocal of the observed values, using the MULTI program; the damping Gauss-Newton method algorithm was used for curve fitting (Yamaoka et al., 1981).
Experimental Animals.
Oatp1a4(−/−) mice were obtained from Deltagen (San Carlos, CA). Oatp1a4(−/−) mice were generated by homologous recombination using a targeting vector in which a 385-bp fragment including exons 1, 2, and 3, corresponding to position of nucleotides 240 to 624 of the mouse Oatp1a4 cDNA clone (GenBank accession no. AB031814), was replaced by a lacZ/Neo insert. Genotyping was performed by PCR analysis using gene-specific (GS) primer [endogenous (E)] (5′-CACAGAGATGAGAAGAGGGTGTTGG-3′), GS primer [targeted (T), E] (5′-GCAAGCATTTGTGGTAAACCTGAGC-3′), and neomycin cassette-specific (Neo) primer (T) (5′-GGGCCAGCTCATTCCTCCCACTCAT-3′). Wild-type (C57BL/6J) mice were supplied by Oriental Yeast Co., Ltd. (Tokyo, Japan). All mice (10–18 weeks of age) were maintained under standard conditions with a reverse dark/light cycle. Food and water were available ad libitum. All experiments involving animals in this study were performed according to the guidelines provided by the Institutional Animal Care Committee (Graduate School of Pharmaceutical Sciences, The University of Tokyo).
Quantification of mRNA Expression of Various Transporters in the Mouse Cerebral Cortex.
The mRNA levels of Oatp1a4, Oatp1c1, Oat3, Mdr1a, Bcrp, Mrp4, and glyceraldehyde-3-phosphate dehydrogenase were quantified using the real-time PCR method. Total RNA was isolated from the cerebral cortex of each of three mice using ISOGEN (Wako Pure Chemicals, Tokyo, Japan). Real-time PCR was performed using a QuantiTect SYBR Green PCR kit (QIAGEN, Valencia, CA) and a LightCycler system (Roche Diagnostics, Mannheim, Germany). An external standard curve was generated by dilution of the target PCR product, which was purified by agarose gel electrophoresis. The absolute concentration of the external standard was measured using PicoGreen dsDNA Quantification Reagent (Invitrogen).
Isolation of Mouse Brain Capillary-Enriched Fraction.
A modification of a method described previously was used for capillary isolation (Ball et al., 2002). In brief, the cortex was homogenized in 0.32 M sucrose (1 g of brain/20 ml of sucrose) using a Polytron homogenizer (Kinematica, Littau-Lucerne, Switzerland). The homogenate was centrifuged at 4°C for 10 min at 2200g, and the resulting pellet was suspended in 25% BSA and centrifuged at 4°C for 10 min at 2200g. The supernatant was decanted, and the pellet was washed three times with buffer (10 mM Tris-HCl and 0.5 mM dithiothreitol, pH 7.6). The purity of the brain capillary-enriched fraction was confirmed by γ-GTP activity (Goldstein et al., 1975).
Western Blot Analysis.
Crude membranes from liver homogenates and brain capillaries were prepared as follows. Liver homogenates and brain capillaries were centrifuged at 4°C for 15 min at 2000g, the supernatant was collected and centrifuged at 4°C for 15 min at 100,000g, and the resulting pellet was resuspended in PBS containing 0.1% protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO) and stored at −80°C until used. The protein concentration was measured using the method of Lowry et al. (1951). The specimens were loaded onto an 8.5% SDS-polyacrylamide gel electrophoresis with a 3.75% stacking gel. Proteins were electroblotted onto a polyvinylidene difluoride membrane (Pall, Port Washington, NY), and the membrane was then blocked for 1 h at room temperature with Tris-buffered saline containing 0.05% Tween 20 (TTBS) and 2.5% ECL Advance Blocking Agent (GE Healthcare, Little Chalfont, Buckinghamshire, UK). The membrane was washed with TTBS and incubated overnight at 4°C with rabbit anti-mOatp1a4 antiserum, which was raised in rabbits against a synthetic peptide consisting of the 20 carboxyl-terminal amino acids of mOatp1a4 (CTNVHRSPTMQNDGERKTKL)-keyhole limpet hemocyanin conjugate (1:1000 in TTBS containing 2.5% ECL Advance Blocking Agent). Detection was performed by binding to horseradish peroxidase-labeled anti-rabbit IgG antibody (1:5000 in TTBS containing 2.5% ECL Advance Blocking Agent; GE Healthcare). Immunoreactivity was detected with an ECL Advance Western Blotting Detection Kit (GE Healthcare). Antigen adsorption was performed by incubating rabbit anti-mOatp1a4 antiserum with mOatp1a4 antigen (50 μg/ml) at 4°C overnight. After the membrane was stripped with Restore Western Blot Stripping Buffer (Pierce Chemical, Rockford, IL), it was incubated with the antigen-adsorbed anti-mOatp1a4 antiserum.
Identification of Membrane Localization of Oatp1a4 in the BBB by Immunohistochemical Staining.
Frozen sections of the brain were prepared from C57BL/6J wild-type and Oatp1a4(−/−) mice by GenoStaff (Tokyo, Japan). Mouse brain specimens were embedded in OTC compound and frozen by dry ice, and 8-μm-thick sections were sliced by cryostat. Before staining, the sections were pretreated by heat-induced epitope retrieval and fixed to glass slides in methanol (−20°C). The sections were incubated in 1% Triton X-100 for 30 min at room temperature, washed three times with PBS, and then incubated in PBS containing 5% bovine serum albumin (BSA-PBS) to block nonspecific protein binding. The sections were then washed three times with PBS and incubated overnight at 4°C in a 1:50 dilution of rabbit anti-mOatp1a4 antiserum and a 1:40 dilution of monoclonal anti-P-gp antibody (C219; Signet Laboratories, Dedham, MA), or a 1:20 dilution of rabbit anti-mOatp1a4 antiserum and a 1:200 dilution of mouse monoclonal anti-GFAP antibody (Millipore Bioscience Research Reagents, Temecula, CA) in BSA-PBS. After incubation, the sections were washed three times with PBS and then were incubated for 1 h at room temperature in a 1:250 dilution of secondary antibodies [Alexa 594 anti-rabbit IgG and Alexa 488 anti-mouse IgG (Invitrogen); or Alexa 488 anti-rabbit IgG and Alexa 594 anti-mouse IgG] and TO-PRO-3 iodide (Invitrogen) in BSA-PBS. The sections were mounted in VECTASHIELD mounting medium (Vector Laboratories, Burlingame, CA) and were viewed under a confocal laser microscope [Zeiss LSM 510 apparatus (Carl Zeiss, Thornwood, NY) or Leica TCS-SP5 (Leica Microsystems CMS GmbH, Mannheim, Germany)].
Determination of Brain-to-Blood Transport of Oatp1a4 Substrates Across the BBB Using Microinjection Technique to the Cerebral Cortex.
The efflux of test compounds from the brain after microinjection into the cerebral cortex was investigated using the brain efflux index (BEI) method as described previously (Sugiyama et al., 2003). Tritium-labeled compounds (70 nCi/mouse) with the nonpermeable reference compound [14C]carboxyl-inulin (2 nCi/mouse) in 0.5 μl of ECF buffer (122 mM NaCl, 25 mM NaHCO3, 10 mM d-glucose, 3 mM KCl, 1.4 mM CaCl2, 1.2 mM MgSO4, 0.4 mM K2HPO4, and 10 mM HEPES, pH 7.4) were injected into the S2 region (4.5 mm lateral to the bregma, 2.5 mm in depth). After intracerebral microinjection, mice were decapitated, and the radioactivity that remained in the ipsilateral cerebrum was determined. The 100 − BEI (percent), which represents the percentage of the test compounds remaining in the cerebrum, is described by eq. 2:
 The elimination rate constant of the compounds eliminated from the brain (kel) was obtained by fitting the 100 − BEI (percent) versus time data. A least-squares regression program (MULTI) (Yamaoka et al., 1981) was used for the calculation.
The elimination rate constant of the compounds eliminated from the brain (kel) was obtained by fitting the 100 − BEI (percent) versus time data. A least-squares regression program (MULTI) (Yamaoka et al., 1981) was used for the calculation.
Determination of Blood-to-Brain Transport of Oatp1a4 Substrates Across the BBB Using in Situ Brain Perfusion.
In situ brain perfusion was performed as described previously (Dagenais et al., 2000). In brief, mice were anesthetized by intraperitoneal injection of pentobarbital sodium (50 mg/kg) and then injected intravenously with GF120918 (10 mg/kg and injection volume of 3.3 ml/kg) dissolved in a 3:2 mixture of propylene glycol-water) 15 min before the perfusion of test compounds. The right common carotid artery was catheterized with a polyethylene-10 tube mounted on a 30-gauge needle. Before insertion of the catheter, the common carotid artery was ligated caudally. During surgery, the body temperature was maintained by placing the mice on a heated plate. The syringe containing the perfusion fluid was placed in an infusion pump (PerkinElmer Life and Analytical Sciences, Waltham, MA) and connected to the catheter. Before perfusion, the thorax of the animal was opened and the heart was cut. Perfusion was started immediately at a flow rate of 1 ml/min. The perfusion fluid consisted of Krebs-Henseleit bicarbonate buffer (25 mM NaHCO3, 118 mM NaCl, 4.7 mM KCl, 1.2 mM MgSO4, 1.2 mM NaH2PO4, 1.2 mM CaCl2, and 10 mM d-glucose). The perfusion fluid was bubbled with 95% O2 and 5% CO2 for pH control (7.4) and was warmed to 37°C in a water bath. Tritium-labeled compounds were added to the perfusate at a concentration of 1 μCi/ml, with carbon-labeled sucrose as a vascular volume marker. The 1-min perfusion was terminated by decapitation. The brain was then removed, and the cortex of the right cerebral hemisphere was placed in a preweighed vial and weighed. The radioactivity associated with the brain and perfusion fluid specimens was determined in a liquid scintillation counter.
The apparent volume of brain distribution (Vbrain) was calculated from the amount of radioactivity in the right cortex using eq. 3:
 where Xtot (disintegrations per minute per gram of brain) is the amount of tracer measured in the right cortex, Cperf (disintegrations per minute per milliliter) is the concentration of tritium-labeled compound in the perfusate, and Vvasc is the brain vascular volume. Vvasc was estimated from the tissue distribution of [14C]sucrose, which is known to diffuse very slowly across the BBB. The brain uptake clearance was calculated from eq. 4:
where Xtot (disintegrations per minute per gram of brain) is the amount of tracer measured in the right cortex, Cperf (disintegrations per minute per milliliter) is the concentration of tritium-labeled compound in the perfusate, and Vvasc is the brain vascular volume. Vvasc was estimated from the tissue distribution of [14C]sucrose, which is known to diffuse very slowly across the BBB. The brain uptake clearance was calculated from eq. 4:
 where T is the perfusion time (minutes).
where T is the perfusion time (minutes).
Determination of the Brain-to-Plasma Concentration Ratios of [3H]Taurocholate, Pitavastatin, and Rosuvastatin after Intravenous Infusion in Wild-Type and Oatp1a4(−/−) Mice.
Male C57BL/6J mice and Oatp1a4(−/−) mice (10–18 weeks) weighing approximately 25 to 35 g were used for these experiments. Under pentobarbital anesthesia (30 mg/kg), the jugular vein was cannulated with a polyethylene-10 catheter for the administration of drugs. The mice then received a constant intravenous infusion of [3H]taurocholate (400 μCi/h/kg), pitavastatin (20 μmol/h/kg), or rosuvastatin (20 μmol/h/kg). Blood samples were collected from the jugular vein at 10 min ([3H]taurocholate) or 120 min (pitavastatin and rosuvastatin) after treatment, and the brain was excised immediately. Plasma specimens were obtained by centrifugation of the blood samples (10,000g). The plasma and brain concentrations of [3H]taurocholate were determined in a liquid scintillation counter, and those of pitavastatin and rosuvastatin were determined with LC-MS analysis.
Quantification of Pitavastatin and Rosuvastatin in Plasma and Brain Specimens.
The brain was homogenized with a 4-fold volume of phosphate-buffered saline to obtain a 20% brain homogenate. Plasma specimens (10 μl) were mixed with 40 μl of ethanol, and brain homogenates (100 μl) were mixed with 400 μl of ethanol. All of these mixed solutions were centrifuged at 15,000g for 10 min. The supernatants of the plasma specimens were mixed with an equal volume of water and subjected to LC-MS analysis. The supernatants of the brain specimens (350 μl) were evaporated, and the pellets were reconstituted with 50 μl of 20% ethanol. The reconstituted specimens were centrifuged at 15,000g for 10 min, and an aliquot of the supernatant was subjected to LC-MS analysis.
An LCMS-2010EV equipped with a Prominence LC system (Shimadzu, Kyoto, Japan) was used for the analysis. Samples for pitavastatin were separated on an L-column ODS (5 μm, 2.1 × 150 mm; Chemicals Evaluation and Research Institute, Tokyo, Japan) in isocratic mode at a flow rate of 0.3 ml/min, and those for rosuvastatin were separated on a CAPCELL PAK C18 MGII column (3 μm, 2 mm × 50 mm; Shiseido, Tokyo, Japan) in binary gradient mode at a flow rate of 0.4 ml/min. The composition of the mobile phase for pitavastatin was acetonitrile-0.05% formic acid (40:60), and the total run time was 5 min. As for rosuvastatin, acetonitrile and ammonium acetate (10 mM) were used for the mobile phase, and the acetonitrile concentration was initially 20%, increased linearly to 50% over 2 min, kept at 50% over 1 min, and finally reequilibrated at 20% for 3 min. Pitavastatin and rosuvastatin were eluted at 3.7 and 3.2 min, respectively. In the mass analysis, pitavastatin and rosuvastatin were detected at mass/charge ratios of 422.15 and 482.15, respectively, under positive electrospray ionization conditions. The interface voltage was −3.5 kV, and the nebulizer gas (N2) flow was 1.5 l/min. The heat block and curved desolvation line temperatures were 200 and 150°C, respectively.
Statistical Analysis.
Data are presented as the mean ± S.E. for three to seven animals, unless otherwise specified. A Student's two-tailed unpaired t test and one-way analysis of variance followed by Tukey's multiple comparison test were used to identify significant differences among groups, where appropriate. Statistical significance was set at P < 0.05. The mean differences of the elimination rate constant for elimination of the compounds from the brain (kel) between wild-type mice and Oatp1a4(−/−) mice with 95% confidence intervals were used for detection of significant difference. When the 95% confidence interval did not cross 0, the result was regarded as significant.
Results
Time Profiles and Concentration Dependence of the Uptake of Various Compounds by mOatp1a4-Transfected HEK293 Cells.
The time profiles of the uptake of pitavastatin, rosuvastatin, pravastatin, taurocholate, digoxin, ochratoxin A, and DPDPE by mock- and mOatp1a4-HEK293 cells are shown in Fig. 1. For each of the compounds the mOatp1a4-HEK293 cells accumulated more than the mock-transfected cells. The specific uptake of pitavastatin and rosuvastatin and taurocholate by mOatp1a4 was determined at 0.5 and 2 min, respectively, because the initial velocity was linear during these periods. The concentration dependencies were examined for pitavastatin, rosuvastatin, and taurocholate by mOatp1a4 are shown as Eadie-Hofstee plots (Fig. 2). Nonlinear least-squares regression analysis showed that the Km and Vmax values for pitavastatin, rosuvastatin, and taurocholate for mOatp1a4 were 8.27 ± 1.06 μM and 1.65 ± 0.14 nmol/min/mg protein, 11.7 ± 2.5 μM and 1.74 ± 0.27 nmol/min/mg protein, and 40.3 ± 17.0 μM and 0.14 ± 0.03 nmol/min/mg protein, respectively.

Time profiles of the uptake of various compounds by mOatp1a4-transfected HEK293 cells. The uptake of [3H]pitavastatin (0.1 μM) (A), [3H]rosuvastatin (0.1 μM) (B), [3H]pravastatin (0.1 μM) (C), [3H]taurocholate (1 μM) (D), [3H]digoxin (0.05 μM) (E), [3H]ochratoxin A (0.02 μM) (F), and [3H]DPDPE (0.005 μM) (G) by mOatp1a4-transfected HEK293 cells at 37°C was examined. ● and ○, uptake by mOatp1a4-transfected HEK293 cells and vector-control (mock-transfected) cells, respectively. Each point represents the mean ± S.E. n = 3–4.

Concentration dependence of the uptake of [3H]pitavastatin (A), [3H]rosuvastatin (B), and [3H]taurocholate (C) by mOatp1a4-transfected HEK293 cells. The uptake of [3H]pitavastatin (A), [3H]rosuvastatin (B), and [3H]taurocholate (C) was determined at various substrate concentrations: pitavastatin, 0.5 to 50 μM; rosuvastatin, 0.5 to 50 μM; and taurocholate, 5 to 500 μM. Mock- and mOatp1a4-HEK293 cells were incubated for 0.5 min (A and B) and 2 min (C) at 37°C to determine the cellular accumulation. Specific uptake was determined by subtracting the uptake by mock-HEK293 cells from that by mOatp1a4-HEK293 cells. ——, fitted line obtained by nonlinear regression analysis. Each point represents the mean ± S.E. n = 3.
mRNA Expression of Xenobiotic Transporters in the Cerebral Cortex of Wild-Type and Oatp1a4(−/−) Mice.
The mRNA expression levels of transporters in the cerebral cortex of wild-type and Oatp1a4(−/−) mice were determined using real-time PCR to check the adaptive regulation of transporters of Oatp1a4(−/−) mice at the BBB. In Oatp1a4(−/−) mice, the mRNA expression level of Oatp1a4 was reduced compared with that of wild-type mice (Table 1). Quantification of mRNA by real-time PCR showed that there were no significant differences between wild-type and Oatp1a4(−/−) mice in the mRNA expression levels of Oatp1c1, Oat3, Mdr1a, Bcrp, and Mrp4 in the cerebral cortex (Table 1).
mRNA expression of transporters in the cerebral cortex
All mRNA levels were determined by real-time PCR and normalized by the value of Gapdh. n = 3, mean ± S.E.
Genotyping of Oatp1a4(−/−) Mice.
Genotyping was performed by PCR analysis using GS and Neo primers. PCR with GS (E) and GS (T, E) primers produces a fragment of 232 bp in wild-type and heterozygous mice, whereas PCR with GS (T, E) + Neo (T) primers produces a fragment of 476 bp in heterozygous and Oatp1a4(−/−) mice (Fig. 3A). The Oatp1a4(−/−) mice were fertile and exhibited no obvious abnormalities.

Expression of Oatp1a4 at the BBB. A, representative Oatp1a4 genotype analysis of Oatp1a4 wild-type, heterozygous, and nullizygous mice. The endogenous and targeted alleles were detected by PCR analysis. PCR products for the GS (E) + GS (T, E) primer pair and the GS (T, E) + Neo (T) primer pair are 232 and 476 bp, respectively. Identifications of wild-type (+/+), heterozygous (+/−), and nullizygous (−/−) offspring are shown. The design of the primer sets is described under Materials and Methods. B, protein expression of Oatp1a4 in the crude membrane fractions of liver, brain capillaries, and the choroid plexus. Crude membrane fractions were prepared from mouse liver homogenate and brain capillaries. Each sample (liver, 2 μg of protein/lane; brain capillaries, 80 μg of protein/lane; and choroid plexus, 20 μg of protein/lane) was subjected to SDS-polyacrylamide gel electrophoresis (8.5% separating gel). Oatp1a4 protein was detected by rabbit anti-mOatp1a4 antiserum preincubated with or without mOatp1a4 antigen. L, liver (crude membrane); BBB, brain capillary-enriched fraction (crude membrane); CPx, choroid plexus. C, double immunostaining of Oatp1a4 with P-gp in brain sections from wild-type (a–c) and Oatp1a4(−/−) mice (d). P-gp was used as a marker of luminal expression: red, Oatp1a4; green, P-gp; blue, nuclei stained with TO-PRO-3. Scale bar, 10 μm (a–c) and 20 μm (d). D, double immunostaining of Oatp1a4 with GFAP in brain sections from wild-type mice. Top panels show the en face image and the bottom panes show the vertical image. The white rigid line in the upper panel indicates the site where the vertical image was obtained. Arrow and arrowhead, luminal and abluminal localization of Oatp1a4, respectively. GFAP was used as a marker of astrocyte: green, Oatp1a4; red, GFAP; blue, nuclei stained with TO-PRO-3. Scale bar, 5 μm.
Protein Expression and Membrane Localization of Oatp1a4 in Mouse Brain Capillaries.
The protein expression of Oatp1a4 in the mouse brain capillaries, liver, and choroid plexus was determined by Western blot analysis. An antiserum against mOatp1a4 detected a protein of approximately 83 kDa in the brain capillary-enriched fraction and a protein of approximately 76 kDa in the liver and choroid plexus (Fig. 3B). The difference in the molecular mass was probably accounted for by differences in the degree of glycosylation. These signals were abolished after preabsorption of the mOatp1a4 antiserum with the antigen, indicating that they were specific for the antigen peptide (Fig. 3B). The membrane localization of Oatp1a4 in mouse brain capillary endothelial cells was examined by immunohistochemical staining using frozen sections of mouse brain, with P-gp used as a marker of the luminal side of the brain capillaries and GFAP used as an astrocyte marker. The Oatp1a4 signal (Fig. 3C, red) overlapped with the P-gp signal (Fig. 3C, green) on one side of the nucleus (Fig. 3C, blue). Furthermore, the Oatp1a4 signals (Fig. 3D, green) were detected between the nucleus (Fig. 3D, blue) and GFAP signals (Fig. 3D, red) in the cross section. These results indicate that Oatp1a4 is expressed in both the luminal and the abluminal membranes in mouse brain capillary endothelial cells. However, no signals were detected by mOatp1a4 antiserum in Oatp1a4(−/−) mice brains, showing the specificity of the signals detected by mOatp1a4 antiserum (Fig. 3C).
Elimination Rates of Pitavastatin, Rosuvastatin, Pravastatin, and Taurocholate from the Brain Across the BBB in Wild-Type and Oatp1a4(−/−) Mice.
The time profiles of the amount of pitavastatin, rosuvastatin, pravastatin, and taurocholate remaining in the brain of wild-type and Oatp1a4(−/−) mice after microinjection into the cerebral cortex are shown in Fig. 4, and the elimination rate constants (kel) are summarized in Table 2. The 95% confidence interval of the mean differences of kel between wild-type mice and Oatp1a4(−/−) mice did not cross 0 (Table 2); the elimination of these compounds from the cerebral cortex after microinjection was therefore significantly delayed in Oatp1a4(−/−) mice compared with wild-type mice with statistical significance.
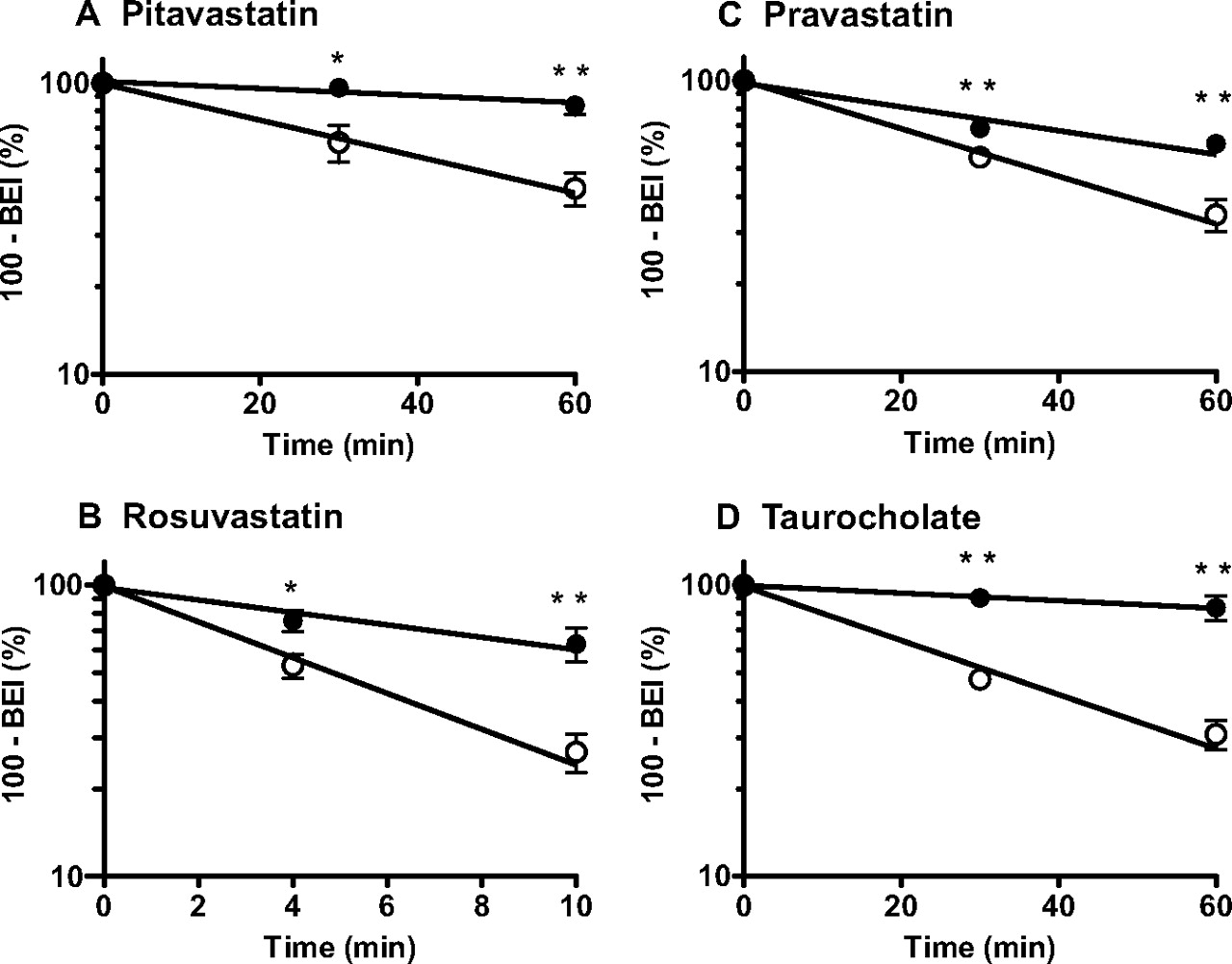
Time profiles of [3H]pitavastatin (A), [3H]rosuvastatin (B), [3H]pravastatin (C), and [3H]taurocholate (D) remaining in the ipsilateral cerebrum after intracerebral microinjection. Remaining radioactive compounds in the cerebrum of wild-type (○) and Oatp1a4(−/−) mice (●) were determined using the BEI method. ——, fitted line obtained by nonlinear regression analysis. Each point represents the mean ± S.E. n = 3–4. Statistically significant differences between wild-type and Oatp1a4(−/−) mice: ∗, P < 0.05; ∗∗, P < 0.01.
Elimination rate constants of compounds from the brain
The elimination rate constant for elimination of the compounds from the brain (kel) was obtained by fitting the 100 − BEI (percent) versus time data. The data used for the calculation are shown in Fig. 4. Data represent mean ± computer-calculated S.D.
BBB Permeability to Oatp1a4 Substrates Determined by in Situ Brain Perfusion in Wild-Type and Oatp1a4(−/−) Mice.
The brain uptake of pitavastatin (0.1–100 μM), rosuvastatin (0.1 μM), pravastatin (0.1 μM), taurocholate (1 μM), ochratoxin A (0.1 μM), and DPDPE (0.1 μM) was evaluated using an in situ brain perfusion technique in wild-type and Oatp1a4(−/−) mice. The uptake of pitavastatin by the brain was saturable, and this component accounted for the major part of the total uptake in wild-type mice (Fig. 5A). The brain uptake clearances of pitavastatin, rosuvastatin, pravastatin, taurocholate, and ochratoxin A were 57 ± 9, 24 ± 4, 11 ± 2, 11 ± 0, and 24 ± 3 μl/min/g brain, respectively, in wild-type mice (Fig. 5, A–E), whereas clearance of DPDPE was negligible (0.091 ± 0.091 μl/min/g brain). The uptake clearance of pitavastatin, rosuvastatin, taurocholate, and ochratoxin A was significantly reduced in Oatp1a4(−/−) mice (15 ± 4, 7.8 ± 1.6, 0.57 ± 0.37, and 10 ± 2 μl/min/g brain, respectively), whereas that of pravastatin was unchanged (Fig. 5). A similar brain vascular space was observed in wild-type mice (10 ± 2 μl/g brain) and Oatp1a4(−/−) mice (8.9 ± 1.6 μl/g brain) in this section.

Comparison of brain uptake of [3H]pitavastatin (A), [3H]rosuvastatin (B), [3H]taurocholate (C), [3H]ochratoxin A (D), and [3H]pravastatin (E) by the cerebral cortex of wild-type and Oatp1a4(−/−) mice after perfusion for 1 min. The uptake clearance was determined by an in situ brain perfusion technique in wild-type (WT) mice (□) and Oatp1a4(−/−) [knockout (KO)] mice (■). The concentrations in the perfusate were 0.1 μM for [3H]rosuvastatin (B), 1 μM for [3H]taurocholate (C), 0.1 μM for [3H]ochratoxin A (D), and 0.1 μM for [3H]pravastatin (E). Each bar represents the mean ± S.E. n = 3–6. Statistically significant differences between wild-type and Oatp1a4(−/−) mice: ∗, P < 0.05; ∗∗, P < 0.01; ∗∗∗, P < 0.001.
Effect of GF120918 on Brain Uptake of Digoxin in Wild-Type and Oatp1a4(−/−) Mice.
The brain uptake of digoxin (0.1 μM) was determined using an in situ brain perfusion technique in wild-type and Oatp1a4(−/−) mice treated with or without GF120918. The brain uptake clearance of digoxin was very low and similar in wild-type and Oatp1a4(−/−) mice. Wild-type and Oatp1a4(−/−) mice predosed with the P-gp inhibitor GF120918 before brain perfusion showed significantly increased brain uptake of digoxin (Fig. 6). The brain uptake of digoxin was significantly reduced in Oatp1a4(−/−) mice only when the brain efflux function of P-gp was prohibited by GF120918 (Fig. 6). The brain vascular spaces in wild-type mice and Oatp1a4(−/−) mice were 9.7 ± 2.5 and 7.8 ± 1.9 μl/g brain, respectively, and were unchanged by pretreatment with GF120918 (7.8 ± 1.3 and 7.0 ± 2.0 μl/g brain, respectively).

Comparison of brain uptake of [3H]digoxin by the cerebral cortex in wild-type and Oatp1a4(−/−) mice with or without predosing with GF120918. GF120918 (10 mg/kg and injection volume of 3.3 ml/kg; dissolved in a 3:2 mixture of propylene glycol-water) was injected intravenously into mice 15 min before the perfusion. The uptake clearance was determined by an in situ brain perfusion technique in wild-type mice (□) and Oatp1a4(−/−) mice (■). The concentration of [3H]digoxin in the perfusate was 0.1 μM. Each bar represents the mean ± S.E. n = 3. Statistical significance was assessed by one-way analysis of variance followed by Tukey's multiple comparison test. ∗∗, P < 0.01, significantly different uptake between wild-type and Oatp1a4(−/−) mice predosed with GF120918. †††, P < 0.001, significantly different uptake with or without predosing with GF120918.
Brain/Plasma Ratio of [3H]Taurocholate, Pitavastatin, and Rosuvastatin Between Wild-Type and Oatp1a4(−/−) Mice.
[3H]Taurocholate was selected to examine the effect of Oatp1a4 dysfunction on the in vivo uptake by the brain. The brain/plasma ratio of [3H]taurocholate determined at 10 min after the start of the infusion was 7.6 ± 0.6 μl/g brain (n = 7) in wild-type mice and was unchanged in Oatp1a4(−/−) mice (6.2 ± 0. 9 μl/g brain, n = 5). Because of its rapid elimination from the systemic circulation, [3H]taurocholate is not a suitable compound to examine the effect of Oatp1a4 dysfunction on the brain concentration after longer exposure. Instead, pitavastatin and rosuvastatin were used as probes. There was no difference in the brain/plasma ratio of pitavastatin and rosuvastatin (microliters per gram of brain) determined at 120 min after the start of the infusion between wild-type and Oatp1a4(−/−) mice: 22 (19 and 26, n = 2) versus 16 (20 and 12, n = 2) and 8.3 (5.6 and 11, n = 2) versus 9.0 (7.9 and 10, n = 2), respectively.
Discussion
A number of studies have suggested that the Oatp transporter family mediates the cellular uptake from the brain side and the blood side of the BBB (Dagenais et al., 2001; Sugiyama et al., 2001; Kikuchi et al., 2004; Taogoshi et al., 2005; Ohtsuki et al., 2007). In the present study, the role of Oatp1a4 in the brain-to-blood and blood-to-brain transport of drugs across the BBB was investigated using Oatp1a4(−/−) mice.
In vitro transport studies using HEK293 cells expressing mOatp1a4 were performed to identify Oatp1a4 substrates. Anionic drugs (including pitavastatin, rosuvastatin, pravastatin, and ochratoxin A), a bile acid (taurocholate), a peptide drug (DPDPE), and a cardiac glycoside (digoxin) were all Oatp1a4 substrates (Fig. 1). Among the drugs tested, pitavastatin exhibited the greatest transport activity. The Km values of pitavastatin, taurocholate, and rosuvastatin were determined (Fig. 2), and those for the former two were found to be similar to those determined in rOatp1a4 (Abe et al., 1998; Kikuchi et al., 2004).
Western blot analysis elucidated the fact that the molecular mass of Oatp1a4 was greater in the brain capillary-enriched fraction than that in the liver and choroid plexus (Fig. 3B). To identify membrane localization of mOatp1a4 in brain capillaries, double immunostaining of Oatp1a4 together with P-gp or GFAP, markers for luminal membrane of the brain capillaries and astrocytes, respectively, was performed. Overlap of the Oatp1a4 signals with P-gp signals indicates luminal expression of mOatp1a4. This finding is also supported by cross section of brain capillaries showing that the intense signals were present between the nuclei (Fig. 3D). Furthermore, we could detect the Oatp1a4 signals, which were not superimposed on P-gp signals (Fig. 3C). In the cross section, the signals were located between the nucleus and GFAP, suggesting that Oatp1a4 is also expressed on the abluminal membrane. Considering the difference in the intensities, we speculate that Oatp1a4 is more abundantly expressed in the luminal membrane than in the abluminal membrane in mouse brain. This pattern is consistent with human brain in which OATP1A2, corresponding to mOatp1a4 in humans (72% homology at the amino acid level), was detected only in the luminal membrane by immunohistochemical staining (Bronger et al., 2005) but not with the recent data in which rOatp1a4 protein was exclusively recovered in the abluminal membrane-enriched fraction (Roberts et al., 2008).
There was no marked change in the expression of other xenobiotic transporters in the cerebral cortex, at least at the mRNA level (Table 1). Therefore, it is most likely that the alteration in drug transport across the BBB observed in Oatp1a4(−/−) mice was due to a lack of Oatp1a4 in the BBB. The role of Oatp1a4 in drug transport across the BBB in the brain-to-blood and blood-to-brain directions was investigated using microinjection into the cerebral cortex and in situ brain perfusion, respectively, in wild-type and Oatp1a4(−/−) mice. Considering the high plasma protein binding of the compounds tested and short transit time of the blood circulation through the brain, reuptake of these organic anions effluxed into the blood circulation would be negligible (so-called sink condition). The brain-to-blood transport of the four Oatp1a4 substrates tested across the BBB was significantly decreased in Oatp1a4(−/−) mice compared with that in wild-type mice (Fig. 4). The impact of Oatp1a4 dysfunction was profound for pitavastatin and taurocholate, whereas that for rosuvastatin and pravastatin was moderate (Table 2). Because the transport activity of rosuvastatin by mOatp1a4 is considerably higher than that for taurocholate and pravastatin (Fig. 1), the impact of Oatp1a4 dysfunction on the brain-to-blood transport does not depend solely on Oatp1a4 activity. In a previous study, rOat3 was suggested to mediate the brain-to-blood transport of pravastatin in rats (Kikuchi et al., 2004). mOat3 is also expressed on the abluminal membrane of mouse brain capillaries (Ohtsuki et al., 2004), which may account for the discrepancy.
The impact of Oatp1a4 dysfunction on the blood-to-brain transport was also investigated using Oatp1a4(−/−) mice (Fig. 5). In wild-type mice, saturation was observed in the blood-to-brain transport of pitavastatin (Fig. 5A). The saturable transport of pitavastatin was probably diminished in Oatp1a4(−/−) mice because the blood-to-brain transport of pitavastatin determined at the highest concentration in wild-type mice was similar to that in Oatp1a4(−/−) mice (Fig. 5A). Namely, the saturable mechanisms is ascribed to Oatp1a4-mediated transport. In addition, the blood-to-brain transport of rosuvastatin, taurocholate, and ochratoxin A was also significantly reduced in Oatp1a4(−/−) mice, whereas that of pravastatin was not (Fig. 5, B–E). The absence of an effect of Oatp1a4 dysfunction on the brain uptake of pravastatin seems reasonable, considering that pravastatin has the lowest transport activity among the Oatp1a4 substrates tested (Fig. 1). It is worth mentioning that the uptake of pitavastatin, rosuvastatin, ochratoxin A, and pravastatin remaining in Oatp1a4(−/−) mice is greater than that of taurocholate, suggesting involvement of other transporter(s) to the blood-to-brain transport of these compounds. In contrast, the brain uptake of DPDPE was almost negligible in wild-type mice, whereas DPDPE exhibits moderate transport activity in Oatp1a4-HEK393 cells (Fig. 1G). This discrepancy will be accounted for by P-gp-mediated efflux (Dagenais et al., 2001). Such cooperation of Oatp1a4 and P-gp was observed for digoxin in this study. The blood-to-brain transport of digoxin was significantly lower in Oatp1a4(−/−) mice compared with that in wild-type mice only with GF120918 pretreatment (Fig. 6). GF120918 is a dual inhibitor of P-gp and Bcrp (Jonker et al., 2000; Lentz et al., 2000). Considering the fact that digoxin is not a BCRP substrate (Pavek et al., 2005), the effect of GF120918 is ascribed to the inhibition of P-gp.
Taken together, these results indicate that Oatp1a4 mediates both the blood-to-brain and brain-to-blood transport of amphipathic organic anions across the BBB. Oatp1a4 is an active uptake transporter for organic anions, but it is also known to mediate the efflux down the electrochemical potential (Li et al., 2000). The fact that Oatp1a4 is expressed both in the luminal and abluminal membranes suggests the possibility that Oatp1a4 mediates the uptake and/or subsequent efflux process in the transcellular transport across the BBB. Because the present study determined the overall transport across the BBB, the critical step mediated by Oatp1a4 could be identified.
It was previously shown that administration of lovastatin and pravastatin reduced total β-amyloid peptides in the brain of TgCRND8 mice expressing mutant human amyloid precursor protein (Chauhan et al., 2004) and that administration of valsartan reduced Alzheimer's disease-type neuropathology and also attenuated the development of β-amyloid-mediated cognitive deterioration in Tg2576 mice, a mouse model of Alzheimer's disease (Wang et al., 2007). It is of great concern that Oatp1a4 could deliver anionic drugs to the brain. The brain/plasma ratio of [3H]taurocholate, pitavastatin, and rosuvastatin after intravenous infusions, the blood-to-brain transport of which was significantly decreased in Oatp1a4(−/−) mice (Fig. 5), were very low despite Oatp1a4-mediated transport. Their distribution volumes in the brain were very close to those in the capillary space in wild-type mice, and the impact of Oatp1a4 dysfunction was below the limit of detection. Because these anionic compounds are highly bound to the plasma proteins, the duration time might not be sufficient to detect Oatp1a4-mediated transport. In addition, ATP-binding cassette transporters may limit their brain penetration at the BBB as observed for DPDPE (Dagenais et al., 2001) and digoxin (Fig. 6). We also reported that the low distribution volume of an active form of oseltamivir (Ro 64-0802) in the brain, which is close to the capillary space in wild-type mice, was ascribed to the Mrp4-mediated efflux at the BBB (Ose et al., 2009). BCRP and MRP4 as well as P-gp may restrict the brain penetration of the selected Oatp1a4 substrates from the blood. Whether Oatp1a4 can deliver drugs to the brain remains controversial. Therefore, whether Oatp1a4 can deliver drugs and other endogenous substrates from the blood to the brain also remains controversial.
In conclusion, Oatp1a4 is expressed in both the luminal and the abluminal membranes of mouse brain capillary endothelial cells and can mediate the transcellular transport of amphipathic organic anions across the BBB in both the blood-to-brain and brain-to-blood directions. Whether Oatp1a4 can deliver drugs and other endogenous substrates from the blood to the brain remains controversial.
Acknowledgments.
We thank Kowa Company Ltd. (Tokyo, Japan) for providing radiolabeled and unlabeled pitavastatin, AstraZeneca (Cheshire, UK) for providing radiolabeled and unlabeled rosuvastatin, and Sankyo (Tokyo, Japan) for providing radiolabeled pravastatin. We also thank Dr. Glynis Nicholls (GlaxoSmithKline Research and Development) for the kind gift of GF120918.
Footnotes
This work was supported in part by Grant-in-Aid for Scientific Research (A) from the Ministry of Education, Culture, Sports, Science and Technology [Grant 20249008] (to Y.S.); and Grant-in-Aid for Scientific Research (B) from the Ministry of Education, Culture, Sports, Science and Technology [Grant 20390046] (to H.K.).
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.109.029454
-
- BBB
- blood-brain barrier
- P-gp
- P-glycoprotein
- Bcrp/BCRP
- breast cancer resistance protein
- Oatp
- organic anion transporter peptide
- BQ-123
- cyclo-d-Trp-d-Asp-Pro-d-Val-Leu
- DPDPE
- [d-penicillamine2,5]-enkephalin
- m
- mouse
- r
- rat
- GF120918
- N-(4-[2-(1,2,3,4-tetrahydro-6,7-dimethoxy-2-isoquinolinyl)ethyl]-phenyl)-9,10-dihydro-5-methoxy-9-oxo-4-acridine carboxamide, Elacridar
- HEK
- human embryonic kidney
- PCR
- polymerase chain reaction
- bp
- base pairs
- GS
- gene specific
- E
- endogenous
- T
- targeted
- Neo
- neomycin cassette-specific
- Oat
- organic anion transporter
- Mdr
- multidrug resistance
- Mrp/MRP
- multidrug resistance-associated protein
- BSA
- bovine serum albumin
- PBS
- phosphate-buffered saline
- TTBS
- Tris-buffered saline containing 0.05% Tween 20
- GFAP
- glial fibrillary acidic protein
- BEI
- brain efflux index
- LC
- liquid chromatography
- MS
- mass spectrometry
- Ro 64-0802
- [3R,4R,5S]-4-acetamido-5-amino-3-(1-ethylpropoxy)-1-cyclohexene-1-carboxylate phosphate.
- Received July 11, 2009.
- Accepted October 13, 2009.
- Copyright © 2010 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}