


Abstract
An analog of epibatidine (EB) was synthesized with an iodine atom in the 2 position of the pyridyl ring. This analog, (±)-exo-2-(2-iodo-5-pyridyl)-7-azabicyclo[2.2.1]heptane (IPH), as well as its two stereoisomers, displayed high affinity for neuronal nicotinic receptors; therefore, radioiodinated IPH, [125I]IPH, was synthesized with specific radioactivities consistently >1000 Ci/mmol, and its properties as a radioligand for neuronal nicotinic receptors were evaluated. The characteristics of [125I]IPH binding in tissue homogenates appeared to be virtually identical to those reported for [3H]epibatidine binding; but the high specific radioactivity of [125I]IPH greatly facilitated measurements of nicotinic receptors in tissues with relatively low receptor densities and/or where tissues are in limited supply. Autoradiography with [125I]IPH provided clear localization of nicotinic receptors in brain and adrenal gland after film exposure times of ≤2 days. We conclude that [125I]IPH will be a very useful radioligand for the study of neuronal nicotinic receptors in brain and in peripheral ganglia.
Neuronal nicotinic cholinergic receptors are composed of α and β subunits, and at least seven α and three β subunits have been cloned from mammalian neuronal tissues (for reviews, see Galzi and Changeux, 1995;Lindstrom, 1995; Sargent, 1993). These subunits have discrete but overlapping distributions, and it is believed that different α and β subunits can combine to form distinct nicotinic receptor subtypes. Although these subtypes have certain properties in common, including the fundamental function of gating cations through their channels, they appear to differ with regard to their channel conductances (Papkeet al., 1989; Mulle and Changeux, 1990) pharmacology (Duvoisin et al., 1989; Luetje and Patrick, 1991), rates of desensitization (Gross et al., 1991) and anatomical distribution (Mulle et al., 1991).
Neuronal nicotinic receptor binding sites in brain can be labeled and studied with several different radioactive agonists, including [3H]ACh (Schwartz et al., 1982), [3H]nicotine (Romano and Goldstein, 1980; Marks and Collins, 1982), [3H]n-methylcarbamylcholine (Abood and Grassi, 1986; Boska and Quiron, 1987) and [3H]cytisine (Pabreza et al., 1991). Over their useful concentration ranges as radiolabeled ligands, all of these agonists appear to label predominantly a single subtype of neuronal nicotinic receptor, one which contains α-4 and β-2 subunits (Whiting et al., 1991; Flores et al., 1992). In fact, because of a lower affinity for most other nicotinic receptor subtypes, these radiolabeled agonists are not useful probes for the receptor subtype(s) in the adrenal gland, autonomic ganglia or related cell lines, most of which contain receptors composed predominantly of α-3 and/or α-5 subunits in association with β-2 and/or β-4 subunits. Similarly, these nicotinic agonists do not have high affinity for nicotinic receptors composed of α-7 subunits and therefore do not label them in binding studies.
In contrast to most other nicotinic agonists, epibatidine (EB), a chlorine-containing alkaloid initially isolated from the skin of the Ecuadoran frog Epipedobates tricolor by Daly and colleagues (Spande et al., 1992), has high potency and efficacy at several different neuronal nicotinic receptor subtypes (Badio and Daly, 1994; Gerzanich et al., 1995). As a radiolabeled ligand, [3H]EB has proved to be useful as a very high-affinity probe for several different subtypes of neuronal nicotinic receptors. Thus, in addition to rat and human brain (Houghtling et al., 1994, 1995), this broad-spectrum nicotinic agonist binds with high affinity to neuronal nicotinic receptors in rat adrenal gland (Houghtling et al., 1994, 1995), chick retina (McKayet al., 1994), IMR-32 human neuroblastoma cells (Davilá-Garcı́a et al., 1995) and rat adrenal pheochromocytoma PC12 cells (Xiao et al., 1995). In addition, [3H]EB binds to several different defined subtypes of recombinant neuronal nicotinic receptors expressed in frog oocytes (Gerzanich et al., 1995) and in stably transfected mammalian cell lines (Xiao et al., 1996).
EB, like all other radioactive nicotinic agonist probes that have been available, has been radiolabeled with tritium, [3H], which limits the specific radioactivity that can be achieved in the molecule to 29 Ci/mmol/[3H] atom incorporated. This limitation, in turn, imposes certain difficulties or restrictions in measuring nicotinic receptors in very small tissues (e.g.,sympathetic ganglia or retina), or where cultured cells are in limited supply or in cells or tissues with a relatively low density of receptors. In addition, because of the relatively low specific radioactivity of [3H]-labeled compounds, autoradiography of nicotinic receptors with these ligands usually requires film exposure times of ≥2 months.
A ligand that retained most of the binding properties of epibatidine but incorporated radioactive iodine, [125I], with its much higher specific radioactivity (2175 Ci/mmol/[125I] atom incorporated) and higher energy could circumvent these limitations. Therefore, IPH, an analog of EB in which iodine replaces chlorine, was synthesized in unlabeled and [125I]-labeled forms (fig. 1). Here we assess the binding properties of IPH and the utility of [125I]IPH as a radiolabeled probe of nicotinic receptors in brain and other neural tissues.
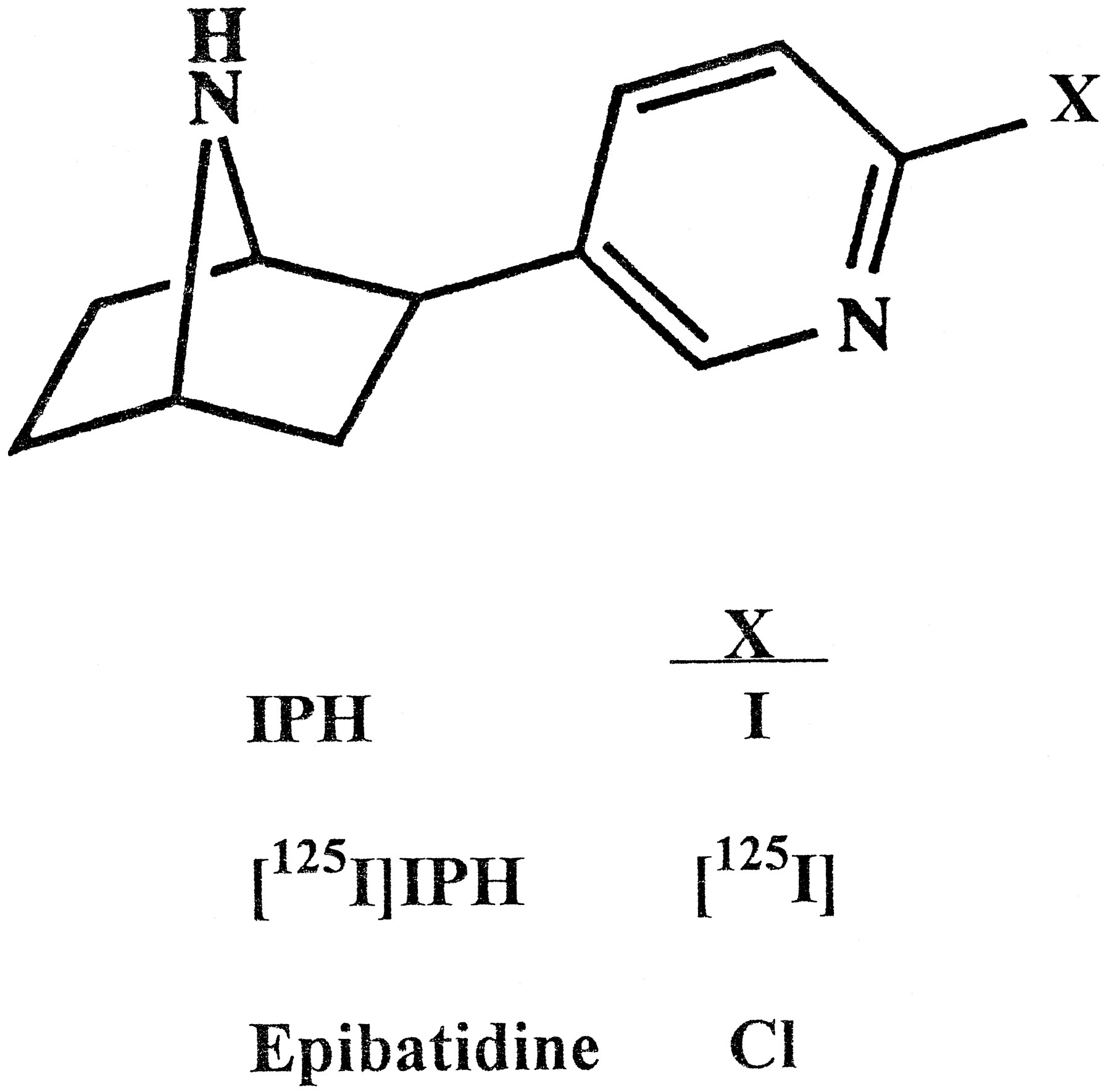
Structures of IPH, [125I]IPH and EB.
Materials and Methods
Synthesis of IPH and [125I]IPH.
The bromopyridyl precursor of IPH, (±)-exo-2-(2-bromo-5-pyridyl)-7-azabicyclo[2.2.1]heptane, was synthesized as described by Horti et al. (1996). Unlabeled and [125I]-labeled IPH were then synthesized as described by Musachio et al. (in press). The products were purified by reverse-phase HPLC, and radiochemical purity was found to be >95%. The [125I]IPH was kept in aqueous ammonium formate (0.4–1.6 M) at −20° and could be used for >2.5 months without noticeable differences in its binding properties (other than its specific activity). The specific radioactivities of the several different batches of [125I]IPH used here were consistently >1000 Ci/mmol.
The enantiomers of nonradioactive IPH were separated by chiral HPLC using a 1 × 25 cm Chiracel o.d. column (Daicel Chemical Industries, Fort Lee, NJ). The mobile phase consisted of 92:8:0.1 hexane/2-propanol/triethylamine at a flow rate of 3.5 ml/min (UV detection at 254 nm). Several 1-mg injections of racemic IPH were made on the chiral column. The resolved enantiomers, designated IPH-A and IPH-B (t R = 18.3 and 20.3 min, respectively), were collected, and the solvent was removed by rotary evaporation. Reinjection of the separated enantiomers indicated that they were >99% optically pure. Because the neuronal nicotinic receptor binding sites did not display stereospecificity for the enantiomers of IPH (see Results), studies to measure the specific rotation of each enantiomer, which would have required synthesis and resolution of much greater amounts of IPH, were not carried out.
Drugs and reagents.
[3H]EB (56.5 Ci/mmol) was obtained from New England Nuclear Research Products (Boston, MA). Drugs and chemicals were purchased from Sigma Chemical (St. Louis, MO), except for (±)-EB and DHβE, which were purchased from Research Biochemicals (Natick, MA).
Tissue preparation.
Rat tissues were obtained from adult male Sprague-Dawley rats. The forebrain was obtained by a single cut just behind the colliculi and excluded the cerebellum and medulla. Specific rat brain regions were dissected with the aid of an atlas (Paxinos and Watson, 1982). Normal human cerebral cortex was obtained from the Brain Resource Center at the Neuropathology Laboratory at Johns Hopkins University School of Medicine. The tissues were suspended in 50 mM Tris·HCl buffer (pH 7.4 at room temperature) and homogenized with a Brinkmann Polytron. The homogenates were centrifuged at 35,000 × g for 10 min and the pellets were resuspended in fresh buffer.
Binding assays.
Aliquots of tissue homogenates equivalent to 0.5 to 10 mg of tissue (30 to 600 μg of protein) were added to tubes containing 50 mM Tris·HCl buffer (pH 7.4) and [125I]IPH or [3H]EB at the indicated concentrations and incubated for 4 hr at 24°C. The assay volumes were maintained at 2.5 ml for saturation studies and 1 ml for competition and tissue distribution studies. Reactions were started by the addition of tissue. Nonspecific binding was determined in tissues incubated with [125I]IPH or [3H]EB in the presence of 300 μM (−)-nicotine hydrogen tartrate, and specific binding was defined as the difference between total binding and nonspecific binding. In drug competition studies, drugs were dissolved in buffer and added at the indicated concentrations. When ACh was used in competition studies, tissues were preincubated for 30 min in buffer containing 1 mM diisopropyl fluorophosphate to inhibit cholinesterases. Incubations were terminated by vacuum filtration through Whatman GF/C filters, which were mounted on a Brandel cell harvester and pre-wet with 0.5% polyethylenimine to reduce binding to the filter (Schwartz et al., 1982). The filters were washed three times with 4-ml aliquots of buffer and then counted in a Gamma counter for [125I]IPH binding or in a scintillation counter for [3H]EB binding. To conserve material where high concentrations of ligand were required, such as in binding saturation studies, the specific radioactivity of [125I]IPH was sometimes reduced by adding unlabeled IPH. This did not appear to affect the results.
Autoradiography.
Autoradiography of [125I]IPH binding sites was carried out in 16-μm cryostat-cut sections of rat brain and adrenal gland. The sections were incubated with ∼500 pM of [125I]IPH (1200 Ci/mmol) and then rinsed, dried and apposed to autoradiographic film (Hyperfilm, Amersham, Arlington Heights, IL) as described previously for studies with [3H]EB (Perry and Kellar, 1995), except that the films were exposed to [125I]IPH-labeled sections for 1 to 2 days instead of 4 to 6 months.
Data analysis.
Saturation and competition binding data were analyzed by nonlinear regression analyses (Accufit Saturation Two-Site and Accufit Competition Programs; Beckman Instruments, Fullerton, CA). The data were fit to a one-site and a two-site model. The simpler model was accepted unless the two-site model gave a statistically better fit of the data (P < .05 by F test). Hill coefficients were calculated from binding saturation and drug competition studies and were analyzed statistically using Student’s t test to assess differences from unity. Concentrations of radioligands used in calculations were corrected for the amount of ligand bound to tissue, which was consistently <20% of the total ligand present.
Results
Binding of the stereoisomers of nonradioactive IPH to nicotinic receptors.
Nicotinic receptor binding sites in brain discriminate between the stereoisomers of nicotine (Romano and Goldstein, 1980;Schwartz et al., 1982; Marks and Collins, 1982;) but not between those of EB (Badio and Daly, 1994; Damaj et al., 1994; Dukat et al., 1993; Houghtling et al., 1995). The (+)- and (−)-isomers of EB also appear to be equipotent in activating nicotinic receptors (Badio and Daly, 1994; Damaj et al., 1994; Li et al., 1993). To determine whether this lack of stereospecificity for EB extends to IPH, in which the chlorine atom is replaced by the larger iodine atom, the stereoisomers of (±)-IPH were separated on a chiral HPLC column and their affinities for nicotinic receptors were examined in competition assays against [3H]EB. As shown in figure 2, the two stereoisomers of IPH (referred to here as IPH-A and IPH-B) and racemic IPH are virtually equipotent in competing for [3H]EB binding in rat forebrain homogenates, and they are only slightly less potent than racemic EB.

Competition by the two stereoisomers of IPH (IPH-A and IPH-B), racemic IPH and (±)EB for nicotinic cholinergic binding sites in rat forebrain membrane homogenates labeled by 1 nM [3H]EB. The mean ± S.D. of two independent measurements of the IC50 values were 3 ± 0.7, 2.7 ± 0.2, 1.3 ± 0.2 and 0.4 ± 0.2 nM, respectively. The n H values were all between 1 and 1.1.
[125I]IPH Binding constants.
Racemic [125I]IPH was synthesized, and its use for measuring neuronal nicotinic receptor binding sites was evaluated. As shown in figure 3A, in rat forebrain homogenates, specific binding of [125I]IPH represented >95% of total binding at concentrations of ≤500 pM and ≥80% of total binding even at concentrations of 4 nM. Binding of [125I]IPH over this concentration range had a Hill coefficient (nH ) close to 1 and fit a model for a single class of binding sites with a dissociation constant (Kd ) of about 90 pM and a density (B max) of approximately 98 fmol/mg of protein (fig. 3B).

Saturation binding of [125I]IPH in rat forebrain membrane homogenates. A, Semilog plot of specific binding of [125I]IPH over a concentration range of 5 pM to 8.7 nM. Inset, saturation plots of specific binding (•) and nonspecific binding (▵) of [125I]IPH. B, Scatchard plot of specific binding data shown in A. Inset, Hill plot of the specific binding data shown in A. Data shown are representative of 6 analyses. The mean ± S.E.M. for the K d andB max values were 92.4 ± 10 pM and 97.6 ± 4 fmol/mg of protein, respectively. The mean ± S.E.M. for the Hill coefficient (n H) was 0.98 ± 0.1.
The pharmacology of [125I]IPH binding sites.
The pharmacological characteristics of the [125I]IPH binding site in rat forebrain were determined in drug competition assays (fig. 4 and table1). Among nicotinic agonists examined, (±)-EB was the most potent competitor, followed by cytisine, nicotine, ACh and carbachol. DHβE was the most potent antagonist examined, being >70 times more potent than curare in competing for binding sites in brain. The ganglionic antagonist mecamylamine did not compete effectively for [125I]IPH binding sites, which is consistent with its lack of potency in competing for brain binding sites labeled by other nicotinic agonist ligands. Similarly, α-bungarotoxin, which has high affinity for receptors containing α-7 subunits, did not compete effectively for [125I]IPH binding sites in brain. All of the agonists examined competed for [125I]IPH binding sites with Hill coefficients close to 1; in contrast, the two effective antagonists, DHβE and curare, competed with shallower slopes, yielding Hill coefficients significantly <1 (table 1). The competition curves for these two antagonists are fit best by a model for two populations of binding sites, and in each case the higher affinity site represents ∼85% of the total population of nicotinic sites labeled by [125I]IPH. Because of the relatively small fraction of the total (15%) represented by the lower affinity site(s) and the limited number of antagonist concentrations used in these studies, we consider these two-site analyses to be preliminary.

Competition by nicotinic drugs for binding sites labeled by [125I]IPH in rat forebrain membrane homogenates. The tissues were incubated with drugs at the indicated concentrations in the presence of ∼200 pM of [125I]IPH. When competition by ACh was examined, tissues were preincubated with diisopropyl fluorophosphate to inhibit cholinesterases. Curves are representative of 3 to 5 independent measurements. The mean ± S.E.M. of the IC50 andn H values are shown in table 1.
Competition by nicotinic drugs for binding sites in rat forebrain homogenates labeled with ∼200 pM of [125I]IPH.
Distribution of [125I]IPH binding sites in brain and other tissues.
The relative distribution of [125I]IPH binding sites in homogenates from brain and other tissues was determined using a ligand concentration of ∼500 pM. This concentration will occupy >80% of the nicotinic receptors with aKd value of ≤100 pM and ∼60% of the nicotinic receptors with a Kd value of ≤350 pM; thus, binding at this concentration should label most neuronal nicotinic receptor binding sites (one exception is the receptor composed of α-7 subunits, which has low affinity for EB and IPH). As shown in figure 5, among the rat brain regions dissected, [125I]IPH binding is highest in the thalamus, intermediate in the amygdala, caudate/putamen and cerebral cortex, somewhat lower in the cerebellum and lowest in the hippocampus and hypothalamus. In human cerebral cortex, [125I]IPH binding is approximately one fourth that in rat cerebral cortex (fig. 5).

Relative distribution of nicotinic binding sites labeled by ∼500 pM [125I]IPH in membrane homogenates from several areas of rat brain and human cerebral cortex. Values are the mean ± S.E.M. from 4 to 6 independent measurements. Rat forebrain (FB) is shown for comparison. Thal, thalamus; Amy, amygdala; CuP, caudate putamen; Ctx, cerebral cortex; Cb, cerebellum; Hip, hippocampus; Hypo, hypothalamus.
Most brain nicotinic receptors with high affinity for agonists are thought to be the α-4/β-2 subtype (Whiting et al., 1991;Flores et al., 1992). To determine whether [125I]IPH also labels other neuronal nicotinic receptor subtypes, measurements were made in the superior cervical ganglion and adrenal gland, both of which are thought to be devoid of α-4 receptor subtypes (Rogers et al., 1992; Henderson et al., 1994; Mandelzys et al., 1994) and in the retina, which probably contains more than one subtype of nicotinic receptor, including both an α-3 and an α-4 subtype (McKay et al., 1994). As shown in figure 6, both the superior cervical ganglion and the retina have high densities of nicotinic sites labeled by [125I]IPH, whereas the adrenal gland has a relatively low density. However, measurements in the isolated adrenal cortex and adrenal medulla indicate that the nicotinic receptors in the adrenal gland are concentrated in the medullary portion, which contains chromaffin cells, whereas fewer receptors are found in the adrenal cortex (fig. 6, inset). A low but measurable amount of specific binding could be detected in the liver (possibly related to ganglia) and in the pituitary gland but not in the kidney or in fat cells (data not shown).

Relative distribution of nicotinic binding sites labeled by ∼500 pM [125I]IPH in homogenates from rat superior cervical ganglia (SCG), retina and adrenal gland. Inset, comparison of [125I]IPH binding in separated adrenal medulla and adrenal cortex. All values are the mean ± S.E.M. from 4 to 6 independent measurements.
Autoradiography with [125I]IPH.
To further assess the value of [125I]IPH as a ligand for neuronal nicotinic receptors, we carried out receptor autoradiography in cryostat-cut sections of rat brain and adrenal gland. After incubation with [125I]IPH, the sections were apposed to autoradiographic film in cassettes for exposure times of 1 to 2 days. The autoradiographs of labeled binding sites in rat brain (fig.7 A and B) indicate that in general, the distribution of [125I]IPH binding sites in brain is similar to that found with other radiolabeled nicotinic agonists, including [3H]ACh, [3H]nicotine and [3H]cytisine (Clarke et al., 1985; Markset al., 1992; Happe et al., 1994; Anderson and Arneric, 1994), and it is virtually identical to the distribution found with [3H]EB (Perry and Kellar, 1995). Thus, in the sections examined, [125I]IPH prominently labeled sites within the cerebral cortex, caudate/putamen, olfactory tubercle, subiculum, dentate gyrus and septal nuclei, as do other radiolabeled nicotinic agonists, but in addition, [125I]IPH, like [3H]EB, labeled a dense concentration of nicotinic sites in the optic tract and optic chiasm, which is not seen with other nicotinic agonist radioligands. This labeling of the optic tract and chiasm probably reflects binding to a non-α-4 receptor subtype, possibly a receptor containing an α-3 subunit (McKay et al., 1994), for which most other radiolabeled nicotinic agonists have low affinity. [125I]IPH also labeled a dense concentration of nicotinic sites in the septofimbrial/fornix nuclei.

Autoradiographs of nicotinic binding sites labeled by [125I]IPH in rat brain and adrenal gland. Slide-mounted tissue sections (16 μm) were incubated with 500 pM of [125I]IPH, washed, dried and then apposed to autoradiographic X-ray film in cassettes for 2 days. A, Sagittal section of the rat brain. B, Coronal section of rat brain at the level of the optic chiasm. C, Cross section of a rat adrenal gland. D, Autoradiographic blank obtained by incubating a rat brain section with 500 pM [125I]IPH in the presence of 300 μM nicotine and then exposing the section to X-ray film as above. Cb, cerebellum; Cpu, caudate putamen; Ctx, adrenal gland cortex; DEn, dorsal endopyriform nucleus; DG, dentate gyrus; DLG, lateral geniculate nucleus; Fr, frontal cortex, GP, globus pallidus; LD, laterodorsal nucleus of thalamus; Med, adrenal medulla; ox, optic chiasm; Par, parietal cortex; Pn, pontine nucleus; PrS, presubiculum; Sfi/f, septofimbrial nucleus/fornix; Tu, olfactory tuberculum.
Autoradiographic measurements in the adrenal gland revealed a dense concentration of [125I]IPH binding sites within the medullary portion of the gland and much sparser or nearly no binding sites in the cortical region of the gland (fig. 7C). This distribution is consistent with the results of binding in membrane homogenates from the dissected adrenal medulla and cortex shown in figure 6. Finally, as shown in figure 7D, nonspecific binding of [125I]IPH to tissue sections is virtually absent.
Discussion
[125I]IPH binds with very high affinity to sites that have the characteristics of neuronal nicotinic receptors. Both the pharmacological properties and distribution of the binding sites in rat brain are nearly identical to those reported for [3H]EB (Houghtling et al., 1995). However, the affinity of [125I]IPH for the majority of these nicotinic sites in rat brain appears to be slightly lower than that of [3H]EB. Moreover, [125I]IPH binding in rat brain fits a model for a single class of sites, whereas in previous studies, [3H]EB binding was fit best to a model for two classes of sites (Houghtling et al., 1995). In rat forebrain, [125I]IPH binding appears to represent primarily the α-4/β-2 subtype of nicotinic receptor, which is the predominant subtype bound with high affinity by most nicotinic agonists in forebrain (Whiting et al., 1991; Flores et al., 1992). However, [125I]IPH can also bind other subtypes of nicotinic receptors with high affinity. Thus, in rat adrenal gland and superior cervical ganglia, which do not express mRNA coding for either α-2 or α-4 subunits of nicotinic receptors (Rogers et al., 1992; Henderson et al., 1994;Mandelzys et al., 1994), most of the [125I]IPH binding probably represents a receptor subtype containing an α-3 subunit; and in the retina [125I]IPH may bind to two or more subtypes one containing an α-3 subunit and another an α-4 subunit (McKay et al., 1994). Furthermore, although [125I]IPH binding in rat forebrain is fit best by a model for a single class of receptors, binding competition data with DHβE and curare are fit best by a model for two sites. This suggests that [125I]IPH actually labels at least two nicotinic receptors in brain that can be distinguished by these two antagonists. Preliminary studies indicate that in each case, the second binding site in brain constitutes only ∼15% of the total receptor population bound by [125I]IPH.
Thus, the fit of [125I]IPH binding to one class of sites probably reflects the fact that the rat forebrain contains predominantly α-4/β-2 receptors, with non-α-4/β-2 nicotinic receptors concentrated in specific nuclei and tracts, such as the medial habenula, interpeduncular nucleus, fasciculus retroflexus, optic nerve and olfactory bulb (Perry and Kellar, 1995; Marks et al., 1996). It is likely that this, along with the somewhat lower affinity of [125I]IPH for α-4/β-2 receptors (which, by diminishing the difference in affinities between α-4/β-2 and non-α-4/β-2 receptors, would make it harder to distinguish two classes of sites), results in [125I]IPH binding fitting a model for a single site.
The high affinity and high specific activity of [125I]IPH make it particularly useful for measuring nicotinic receptor binding sites in peripheral ganglia, which have evaded labeling with most other ligands. Thus, these studies with [125I]IPH revealed a high density of nicotinic receptors in superior cervical ganglia, where they are critical for neurotransmission in the sympathetic nervous system. Similarly, [125I]IPH binding allowed comparison of the distribution of nicotinic receptors in the adrenal medulla and adrenal cortex. Both homogenate binding and autoradiographic studies demonstrate that the nicotinic receptors of the adrenal gland are concentrated in the medullary region of the gland, where they are believed to be located on chromaffin cells. The nicotinic receptors on these cells mediate the release of epinephrine and norepinephrine in response to stimulation by ACh released from the splanchnic nerve or by exogenous nicotine.
The high affinity of IPH for a broad spectrum of neuronal nicotinic receptors, along with its very low nonspecific binding and its ready penetration into the brain after parenteral injection, makes it potentially very useful as a radioiodinated ligand for in vivo measurements of nicotinic receptors. In fact, initial studies with [125I]IPH in mouse and [123I]IPH in baboon (Musachio et al., 1997) suggest that it should prove useful as a ligand for in vivo imaging of brain nicotinic receptors by single-photon emission computed tomography.
In summary, [125I]IPH should be a very useful new tool for the study of neuronal nicotinic receptors. Like its analog EB, [125I]IPH binds with high affinity to several different neuronal nicotinic receptor subtypes. Furthermore, its high specific radioactivity, coupled with its high affinity and low nonspecific binding, permits direct measurements of nicotinic receptors in autonomic nervous system ganglia, adrenal gland and retina. Nicotinic receptors in these tissues are crucial for neurotransmission, but they have been difficult or impossible to measure with other ligands. Finally, the high specific radioactivity of [125I]IPH allows autoradiographic images of neuronal nicotinic receptors to be obtained with film exposure times of 1 or 2 days instead of the 2 months or longer usually required with [3H] ligands.
Footnotes
-
Send reprint requests to: Kenneth J. Kellar, Ph.D., Department of Pharmacology, Georgetown University School of Medicine, Washington DC 20007.
-
↵1 This work was supported in part by National Institutes of Health Grant DA06486. The Brain Resource Center at Johns Hopkins University is supported by National Institutes of Health Grant AG05146.
- Abbreviations:
- ACh
- acetylcholine
- DHβE
- dihydro-β-erythroidine
- EB
- epibatidine
- IPH
- (±)-exo-2-(2-iodo-5-pyridyl)-7-azabicyclo[2.2.1]heptane
- HPLC
- high performance liquid chromatography
- Received January 8, 1997.
- Accepted March 10, 1997.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}