


Abstract
We previously reported that the intraplantar (i.pl.) application of nociceptin/orphanin FQ (N/OFQ) at extremely low doses elicited a nociception through a substance P (SP) release from nociceptor endings. In the present study, the nociception induced by SP (and N/OFQ) was abolished by intrathecal (i.t.) injection of neurokinin1(SP receptor) antagonist, suggesting the involvement of the stimulation of nociceptive primary SP neuron and SP release into spinal synapses. On the other hand, similar low doses of N/OFQ (i.t.) exerted nociceptive responses, characterized by scratching, biting, and licking, and these responses were blocked by an neurokinin1antagonist (i.t.) or capsaicin pretreatment or in tachykinin 1 gene knockout mice (tac1−/− mice), suggesting that N/OFQ receptor (NOR) also exists on the spinal terminals of SP neurons. When wide ranges of N/OFQ doses were used, a typical bell-shaped dose-response relationship was observed in both peripheral and central nociception tests. Furthermore, N/OFQ (1 nmol) administered i.pl. blocked SP (i.pl.)-induced flexor responses, which were abolished by pertussis toxin pretreatment or in NOR gene knockout (NOR−/− ) mice. On the other hand, N/OFQ administered i.t. blocked SP (i.t.)-induced scratching, biting, and licking in capsaicin-pretreated and tac1−/− mice, and this antinociception was abolished in NOR−/− mice. All these findings suggest that N/OFQ has biphasic actions depending on doses in the nociceptors and spinal synapses and has postsynaptic antinociceptive actions in spinal cord by modulating SP signaling.
The family of the G protein-coupled opioid receptors consisting of the μ, δ, and κ receptors (Evans et al., 1992; Kieffer et al., 1992;Fukuda et al., 1993; Thompson et al., 1993; Yasuda et al., 1993) was recently extended by a novel member (Fukuda et al., 1994; Mollereau et al., 1994; Nishi et al., 1994) that did not bind any of the typical opioid receptor ligands (Meunier, 1997). Identification of the orphan receptor in this way led to the advent of “reverse pharmacology” to identify the corresponding physiological ligands. Two independent groups discovered the identical natural ligand: heptadecapeptide, named nociceptin (Meunier et al., 1995) or orphanin FQ (Reinscheid et al., 1995). Despite the fact that nociceptin/orphanin FQ (N/OFQ) and its receptor (NOR) are structurally similar to dynorphin A and opioid receptors, accumulated reports reveal that N/OFQ produces hyperalgesic (or antiopioid) effects when injected into the brain and spinal cord (Meunier, 1997). Along this line, repeated i.c.v. injection of antisense oligonucleotide for NOR resulted in significant analgesia in mice (Meunier et al., 1995). However, there are reports that N/OFQ also has antinociceptive (analgesic) effects on the tail-flick test when injected into the brain and spinal cord (Meunier, 1997). In addition, mice with a genetic lesion in the NOR gene (NOR−/− mice) displayed normal baseline nociceptive responses in several analgesic paradigms. Thus, the role of N/OFQ in pain signaling in vivo remains unclear.
Most recently, we reported that the peripheral application of extremely low doses of N/OFQ produced nociceptive responses in the simple and very sensitive peripheral nociception test (Inoue et al., 1998a,b). Because the nociceptors are distant from the cell body in the dorsal root ganglion, the site of actions of various pharmacological reagents affecting such behavioral responses could be confined to nerve endings (or nociceptors). In addition, taking into account that primary afferent neurons are bipolar cells, actions on the nociceptors of such neurons could be expected similarly on the other, spinal synapses. For this purpose, we analyzed the nociceptive responses characterized by scratching, biting, and licking (SBL responses) induced by N/OFQ administered intrathecally (i.t.), on the analogy to the peripheral paradigm. Here, we found opposite N/OFQ-induced actions on peripheral and central nociceptive responses depending on the doses used, and we discuss their possible mechanisms.
Materials and Methods
Animals.
Male ddY mice weighing 20 to 22 g were used in most of experiments. In some experiments, mice with a targeted disruption of tachykinin 1 gene (tac1−/−) and the wild type (tac1+/+; Zimmer et al., 1998) were used, whereas in some other experiments, homozygous mice lacking the NOR gene (NOR−/−) and the wild type (NOR+/+; Nishi et al., 1997) were used. The experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by The Declaration of Helsinki.
Drugs.
N/OFQ was obtained from Sawady Technology (Tokyo, Japan). Substance P (SP) was from Peptide Institute (Osaka, Japan). MEN-10376 was obtained from Research Biochemicals Inc. (Natick, MA). Capsaicin was obtained from Nacalai Tesque (Kyoto, Japan). Pertussis toxin (PTX) was obtained from Funakoshi (Tokyo, Japan). CP-96345 and CP-96344 were generously provided by Pfizer Central Research (Sandwich, Kent, UK). N/OFQ, SP, PTX, CP-96345, and CP-96344 were dissolved in physiological saline, MEN-10376 was dissolved in 25% dimethyl sulfoxide, and capsaicin was dissolved in 10% ethanol and 10% Tween 80 in physiological saline.
Evaluation of Nociceptive Flexor Responses
All experiments were performed in compliance with the relevant laws and institutional guidelines. Nociceptive flexor responses induced by intraplantar (i.pl.) injection (2 μl) of nociceptive substances were evaluated in mice as previously described (Inoue et al., 1997, 1998a,b;Tokuyama et al., 1998). Briefly, mice were lightly anesthetized with ether and held in a cloth sling with their four limbs hanging free through holes. The sling was suspended on a metal bar. All limbs were tied with strings; three were affixed to the floor, whereas the fourth was connected to an isotonic transducer and recorder. Mice were lightly anesthetized with ether, and a small incision was made in the surface of right hindlimb planta. Two polyethylene cannulas (outer diameter, 0.61 mm) filled with drug solution were connected to a microsyringe. Because we used light and soft polyethylene cannulas, they did not fall off the paw during the experiments. All experiments were started after complete recovery (20–30 min) from the light ether anesthesia, and i.pl. injection of second saline did not show any significant flexor responses. Because the intensity of flexor responses differs from mice to mice, we used the greatest response among spontaneous and nonspecific flexor responses occurring immediately after cannulation as the maximal reflex. Nociceptive activity was represented by the percentage of maximal reflex observed before drug challenges in the beginning of each experiment. In this case, N/OFQ (or SP) filled in a tandem manner separated by an air space in cannulas. N/OFQ (or SP) challenge was performed at 5-min intervals. In most experiments using wide ranges of N/OFQ doses, we evaluated the average of flexor responses to two consecutive challenges per each dose, and at the most, four different doses were tested in each mouse, to obtain stable responses throughout experiments. In some experiments, the antinociceptive effect of N/OFQ was expressed as the ratio of the response observed for the average of two repeated control SP-induced responses in the beginning of the experiments. In this case, SP was administered i.pl. at 10 and 5 min before and 5, 10, 20, and 30 min after N/OFQ injection.
Evaluation of Nociceptive SBL Responses.
When nociceptive substances are i.t. injected in mice, nociceptive responses characterized by scratching of the limbs and biting and licking of the paws or tail (SBL responses) are observed. The nociceptive activity was evaluated by the total response time (given in seconds) over 30 min after the i.t. injection (Hylden and Wilcox, 1980) of nociceptive substance in 5 μl, according to the previous report (Sakurada et al., 1991).
Capsaicin Treatment.
Capsaicin was injected s.c. into the back of newborn (P4) ddY mice at a dose of 50 mg/kg. This treatment is known to cause a degeneration of small-diameter afferent sensory neurons (Hiura and Ishizuka, 1989). As a control, the vehicle (10% ethanol and 10% Tween 80 in physiological saline) used for dissolving capsaicin was injected into mice. For nociceptive tests, capsaicin- or vehicle-pretreated mice weighing 20 to 22 g were used. Gross behavioral changes in treated mice were not observed.
Immunohistochemistry.
Immunohistochemical experiments using capsaicin-treated mice were performed by using primary anti-SP antibody (Chemicon International, Temecula, CA) at a dilution of 1:40. The secondary antibody was fluorescein isothiocyanate-labeled goat anti-rabbit IgG (Vector Laboratories, Peterborough, UK) at a dilution of 1:300.
Statistical Analysis.
The data were analyzed using Student’s t -test after multiple comparisons ANOVA. The criterion of significance was set at p < .05. All results are expressed as mean ± S.E.
Results
N/OFQ-Induced Nociceptive and Antinociceptive Actions through Nociceptors.
When N/OFQ was injected into the plantar surface (i.pl.) of mouse hindlimb, it induced very short-acting but constant nociceptive flexor responses on repeated challenges, as previously reported (Inoue et al., 1998a). N/OFQ-induced nociceptive responses were dose dependent in a wide range of doses from 0.01 to 100 fmol (i.pl.), whereas they started declining from 1 pmol to 1 nmol (i.pl.; Fig. 1A). The nociceptive response to an application of N/OFQ at high doses (10 pmol to 1 nmol) administered in the beginning of experiments was equal to that on repeated challenges of this peptide (data not shown). However, approximately 20% of nociceptive responses remained even when 1 nmol of N/OFQ i.pl. was used. On the other hand, SP induced similar dose-dependent nociceptive responses in ranges between 10 fmol and 100 pmol i.pl., as previously reported (Inoue et al., 1998a).

Nociceptive and antinociceptive effects of N/OFQ in the peripheral nociceptive flexor test. A, dose-response curves of N/OFQ- and SP-induced flexor responses in ddY mice. Each peptide (in 2 μl) was administered i.pl. to mice. Results represent the percentage of each N/OFQ- (or SP-) induced response to the maximal one, as described in the text. B, time course of inhibitory effects of N/OFQ (1 nmol) on SP (10 pmol)-induced flexor responses and their blockade by PTX (10 ng i.pl.) treatment in ddY mice. SP challenges for control responses were performed twice at 5-min interval, followed immediately by N/OFQ injection through another cannula. The results represent the percentage ratio of SP response at the indicated time after N/OFQ (or vehicle) injection to the average of control SP responses. In experiments using PTX, this toxin injection was performed 5 min before N/OFQ injection. *p < .05, compared with vehicle-treated mice. #p < .05, compared with N/OFQ alone. C, loss of inhibition of SP (10 pmol)-induced nociception by N/OFQ (1 nmol) at 30 min after N/OFQ (or vehicle) injection in NOR−/− mice. All data represent the mean ± S.E. from five separate experiments.
To characterize the attenuated effects of N/OFQ at higher doses, we examined how N/OFQ at a high dose affects SP responses. When N/OFQ (1 nmol i.pl.) was administered 5 min after the second control challenge of SP (10 pmol i.pl.), it showed an immediate but transient weak nociceptive response. However, pretreatment with N/OFQ inhibited SP responses (Fig. 1B). SP responses were completely abolished 30 min after the N/OFQ injection. Such N/OFQ (i.pl)-induced antinociceptive actions were completely reversed by PTX (10 ng i.pl.) pretreatment (Fig. 1B) and were abolished in NOR−/− mice (Fig. 1C).
N/OFQ and SP Stimulate Primary SP Neurons in Exerting Peripheral Nociceptive Responses.
Because we found that N/OFQ administered i.pl. exerts nociceptive responses through an SP release from nociceptor endings (Inoue et al., 1998a), it is evident that N/OFQ stimulates SP-containing primary afferent neurons, such as C-fibers. As shown in Fig. 2, N/OFQ (1 fmol i.pl.)-induced nociceptive flexor responses were indeed blocked by 10 and 100 pmol of CP-96345, a neurokinin1 (NK1) antagonist, administered i.t. 30 min before N/OFQ (i.pl.) challenge but not by 100 pmol of CP-96344, an inactive isomer (Snider et al., 1991). Similarly SP (i.pl.)-induced responses were also blocked by CP-96345 but not by CP-96344. These findings suggest that both N/OFQ and SP stimulate primary afferent nociceptive SP neurons.

Blockade of N/OFQ- (or SP)-induced nociceptive flexor responses by i.t. injected NK1 antagonist. The results represent the flexor responses to N/OFQ (1 fmol i.pl.) or SP (10 pmol i.pl.) as the percentage of maximal reflex. CP-96345 (an NK1 antagonist) or CP-96344 (an inactive derivative) at doses indicated in the figure was administered i.t. 30 min before the test. Data are the mean ± S.E. from five separate experiments. *p < .05, compared with vehicle-treated mice (Veh).
N/OFQ (i.t.)-Induced Nociceptive SBL Responses through an SP Release from Spinal Synapses.
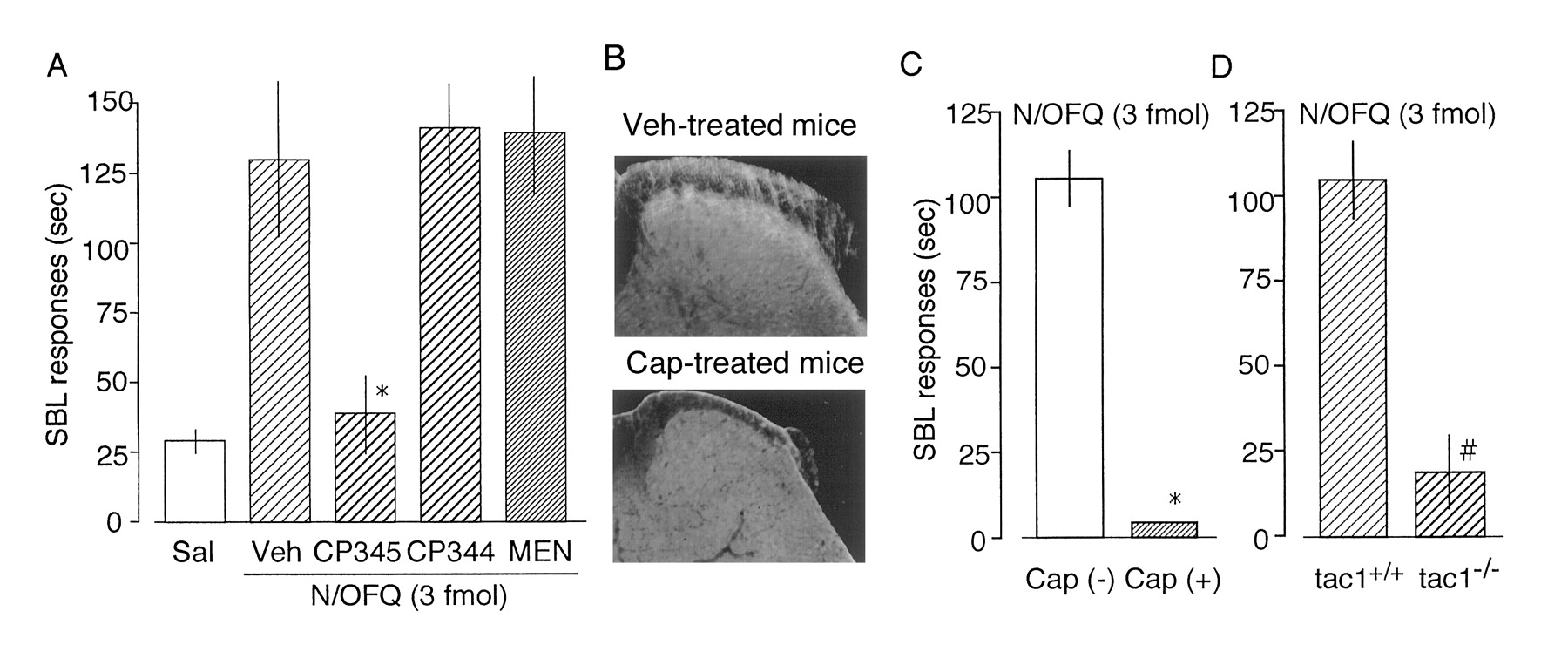
Because there are reports that SP administered i.t. produces nociceptive SBL behaviors (Hylden and Wilcox, 1983; Takahashi et al., 1987), it is expected that N/OFQ may release SP from spinal synapses. According to the above-mentioned results, N/OFQ was administered i.t. to determine such SBL responses. As shown in Fig. 3A, N/OFQ showed a dose-dependent SBL response in a wide range of doses from 3 amol to 3 fmol. However, there also was a decline when the dose increased from 30 fmol to 300 pmol. Unlike the case with peripheral responses, SBL responses were decreased to the control level when 1 nmol i.t. of N/OFQ was administered. Such a bell-shaped curve with N/OFQ contrasts with the typical dose-dependent curve achieved with SP. N/OFQ (3 fmol i.t.)-induced SBL response was abolished in NOR−/− mice (Fig. 3B), indicating that this response is mediated through its receptor. Just like with peripheral responses, N/OFQ (3 fmol i.t.)-induced SBL responses were blocked by CP-96345 (10 nmol i.t.) but not by CP-96344 (10 nmol i.t.; Fig. 4A). In addition, these responses were not affected by MEN-10376 (2 nmol i.t.), a specific NK2 receptor antagonist (Maggi et al., 1991), which blocked NK A-induced but not SP-induced SBL responses (data not shown).

Nociceptive effects of N/OFQ evaluated by nociceptive SBL responses. A, dose-response curves of N/OFQ- or SP-induced SBL responses in ddY mice. Each peptide was administered i.t. in a volume of 5 μl into mice. B, loss of N/OFQ (3 fmol i.t.)-induced SBL responses in NOR−/− mice. Data are the mean ± S.E. from seven separate experiments. *p < .05, compared with NOR+/+ mice.

N/OFQ-induced SBL responses through an SP release. A, blockade of N/OFQ (3 fmol i.t.)-induced SBL responses to NK1 antagonist in ddY mice. Saline (in 5 μl) was administered i.t. to show the basal SBL response (Sal). Saline (Veh, 5 μl), CP-96345 (CP345, 10 nmol), CP-96344 (CP344, 10 nmol), or MEN-10376 (MEN, 2 nmol, NK2 antagonist) was administered i.t. 5 min before N/OFQ injection (i.t.). Data are the mean ± S.E. from six separate experiments. *p < .05, compared with Veh. B, confirmation of capsaicin-induced loss of SP-immunoreactivity in the dorsal horn of the spinal cord in mice used for the test. C and D, loss of N/OFQ (3 fmol i.t.)-induced SBL responses in capsaicin (Cap)-pretreated ddY mice or in tac1−/− mice. Data are the mean ± S.E. from six separate experiments. *p < .05, compared with Cap(−) mice. #p < .05, compared with tac1+/+ mice.
Peripheral pain is transduced to the dorsal horn of the spinal cord through small-diameter primary afferent neurons, such as Aδ- and C-fibers. Capsaicin treatment of neonatal animals leads to the degeneration of C-fibers (Hiura and Ishizuka, 1989), resulting in a reduction in SP-like immunoreactivity in the substantia gelatinosa of spinal cord (Fig. 4B). In such mice, N/OFQ (3 fmol i.t.)-induced SBL response was completely blocked (Fig. 4C). This finding was further supported by the experiments using tac1−/− mice but not in wild-type littermates (Fig. 4D).
N/OFQ-Induced Antinociceptive Actions in Spinal Cord.
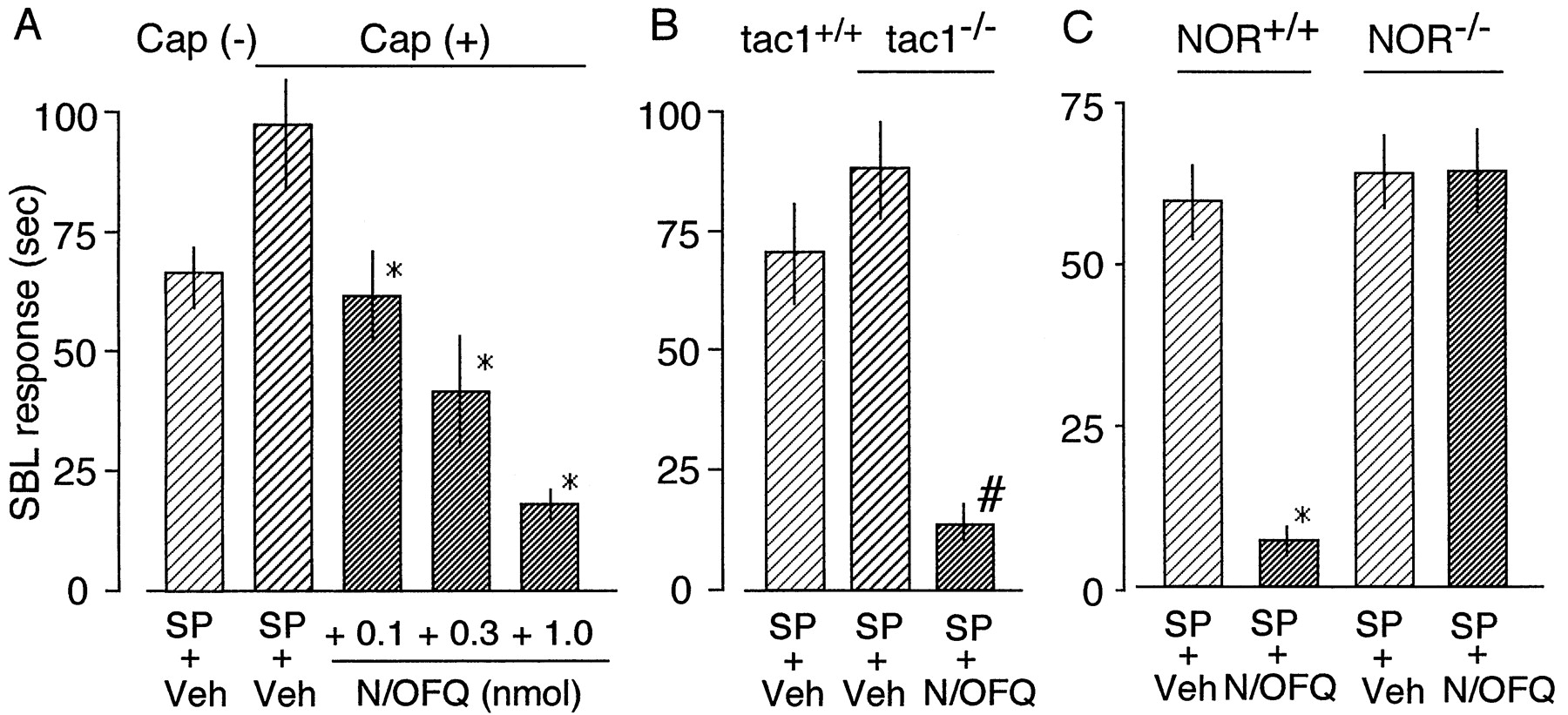
To determine why high doses of N/OFQ were less efficacious than low doses of this peptide (Fig. 3A), we assessed the possibility that N/OFQ may also exert antinociceptive effects at high doses. For this purpose, we used capsaicin-pretreated mice to remove the contribution of the presynaptic actions of N/OFQ. As shown in Fig.5A, SP (100 pmol i.t.)-induced SBL responses (mean ± S.E.) in such mice significantly increased from 71.6 ± 4.3 s (n = 8) to 97.0 ± 11.2 s (n = 8), being consistent with the previous report that capsaicin pretreatment causes a supersensitization to SP (Mantyh and Hunt, 1985). These changes are generally accepted as a denervation supersensitization. In such mice, the SP-induced SBL responses were inhibited by N/OFQ in a dose-dependent manner (Fig. 5A), with 50% of antinociceptive dose (AD50, mean ± S.E.) of 0.17 ± 0.02 nmol i.t. Similar findings were observed when tac1−/− mice were used (Fig. 5B). SP responses in tac1−/− mice (87.6 ± 10.1 s,n = 5) were slightly higher than those in tac1+/+ mice (70.3 ± 10.4 s,n = 5), but there was no significant difference. The supersensitization and/or up-regulation of NK1 receptor have been previously reported elsewhere (Mantyh and Hunt, 1985; Zimmer et al., 1998). To confirm that N/OFQ antinociceptive action mediates through NOR, we used NOR−/− mice. SP (100 pmol)-induced SBL responses in NOR−/− mice were slightly higher than those in NOR+/+ mice, but there was no significant difference (Fig. 5C). However, N/OFQ also inhibited SP-induced SBL responses from 59.8 ± 5.7 s (n = 5) to 6.9 ± 2.2 s (n = 5) in NOR+/+ mice but not in NOR−/− mice (Fig. 5C). These findings strongly suggest that the antinociceptive activity of N/OFQ involves the inhibition of postsynaptic SP responses through NOR.

Inhibitory effects of N/OFQ on SP-induced nociceptive responses in the spinal cord. A, dose-dependent inhibition of SP (100 pmol i.t.)-induced SBL responses to various doses of N/OFQ (i.t.) in capsaicin-treated mice. *p < .05, compared with SP + Veh in Cap(+) mice. B, inhibition of SP (100 pmol i.t.)-induced SBL responses to N/OFQ (1 nmol i.t.) in tac1−/− mice. *p < .05, compared with SP + Veh in tac1−/− mice. C, loss of N/OFQ-induced antinociception in NOR−/− mice. There was a significant (*p < .05, compared with SP + Veh) inhibition of SP (100 pmol i.t.)-induced SBL responses to N/OFQ (1 nmol i.t.) in NOR+/+ mice, whereas there was no significant inhibition in NOR−/− mice. All data are the mean ± S.E. from six separate experiments.
Discussion
The C-fiber polymodal nociceptors could be good targets for studying the molecular and cellular bases of inflammatory pain because they respond to inflammatory mediators, such as bradykinin from plasma, histamine from mast cells, serotonin from platelets, SP from C-fiber nociceptors, and prostaglandins from various cells. In addition, these neurons possess SP as a neurotransmitter and have properties that are selectively degenerated by capsaicin treatment. The peripheral nociception test used in this study has been developed for the purpose of analyzing in vivo signaling mechanisms at the level of C-fiber nociceptor endings (Inoue et al., 1998a). This test has several advantages over many other assays of analgesia: 1) it is sufficiently sensitive to assess very weak and short-acting nociceptive responses induced by the local application of extremely small amounts of pain-producing substance, 2) the nociceptive responses in this test appear to involve relatively simple molecular and neuronal mechanisms because they are attributed to the stimulation of identified receptors, and 3) because the peripheral nerve endings are distant from the cell body in the dorsal root ganglion, the site of actions of various pharmacological reagents affecting such behavioral responses could be confined to nerve endings.
In this test, N/OFQ administered i.pl. elicited biphasic nociceptive and antinociceptive actions, depending on the doses administered. As previously reported (Inoue et al., 1998a), N/OFQ at extremely low doses (10 amol to 100 fmol) was found to produce nociception in mice through an SP release from nociceptor endings via activation of Gαi and phospholipase C (PLC). However, N/OFQ at higher doses than 100 fmol showed declining responses. We speculate that the biphasic dose-response relationship of N/OFQ is not due to desensitization because the nociceptive response to an application of N/OFQ at high doses (10 pmol to 1 nmol) administered in the beginning of experiments was equal to that on repeated challenges of this peptide (data not shown). In addition, the stable flexor responses are shown on repeated challenges of N/OFQ at 100 fmol, a dose at which the peptide shows a maximal response (data not shown). Such a biphasic action might be explained by the fact that 1 nmol of N/OFQ blocked SP-induced nociceptive actions. In other words, N/OFQ at a high dose inhibits the nociception by SP that is released by N/OFQ itself through its specific receptor and PTX-sensitive G proteins. We speculate that the antinociceptive signaling on high doses of N/OFQ is due to the release of βγ subunits from Gαi/othrough NOR, inhibiting the production of free Gαq/11 through the NK1 receptor as postreceptor cross-talk. This possibility is based on the fact that the stoichiometry of receptor/Gαq/11 coupling is quite low compared with that of receptor/Gαi/o (Pang and Sternweis, 1990). Alternatively, Gαipossesses a weaker intrinsic activity to stimulate PLC and may be a competitive antagonist (or partial agonist) against Gαq, which possesses a stronger intrinsic activity for PLC, as reported elsewhere (Ueda et al., 1995a,b, 1999).
Taking into account that primary afferent neurons (or nociceptors) are bipolar cells, the in vivo signaling at the peripheral side could be also expected on the other, central side. Here, we obtained important findings that N/OFQ- or SP-induced peripheral responses were blocked by i.t. injected NK1 antagonist. This suggests that these peptides stimulate nociceptive primary SP neurons. In the present study measuring nociceptive responses characterized by SBL responses, N/OFQ administered centrally (or i.t.) also produced potent nociceptive actions at extremely low doses ranging from 3 amol to 3 fmol (i.t.). As reported in the case with peripheral mechanisms, N/OFQ-induced SBL responses were found to be mediated through an SP release from the spinal terminals because they were abolished by NK1 but not NK2 antagonist (Fig. 4A), in capsaicin-pretreated mice (Fig. 4C), in which primary afferent nociceptive SP neurons of a small diameter had been selectively degenerated (Hiura and Ishizuka, 1989) and in tac1−/− mice (Fig. 4D). The potency of N/OFQ-induced nociceptive response was extremely high compared with that of SP response in nerve endings and spinal cord (Figs. 1A and 3A). We are speculating on the working hypothesis that one molecule of N/OFQ may release many molecules of SP and that this mechanism works as an amplification in pain signaling. Alternatively, there is another possibility that endogenous SP released by N/OFQ may have the advantages of efficient accessibility to the receptor compared with exogenously administered SP. On the other hand, it was speculated that N/OFQ might bind to the NK1 receptor because the N/OFQ-induced peripheral (Inoue et al., 1998a) and central (Fig. 4A) nociceptive responses were completely blocked by the NK1 receptor antagonist. Regarding this issue, we propose a working hypothesis. Because we observed that N/OFQ-induced actions were completely abolished in NOR−/−mice and by treatment with PTX or Gi antisense oligodeoxynucleotides (Figs. 1, 3, and 5; Inoue et al., 1998a), it is unlikely that these actions were mediated through NK1 receptor. This view was also supported by the finding that N/OFQ at high doses antagonized the SP-induced SBL responses (Fig. 5A). N/OFQ showed a bell-shaped dose-response relationship in the spinal cord as well as peripheral paradigm. Unlike the case with the peripheral paradigm, we must separately consider the contribution of postsynaptic mechanisms to the antinociceptive actions by N/OFQ administered centrally, although presynaptic mechanisms could be discussed analogously. For this purpose, we used capsaicin-pretreated mice devoid of presynaptic mechanisms. In such mice, SP (i.t.) responses were enhanced compared with vehicle-treated mice (Fig. 5). N/OFQ at higher doses (0.1–1 nmol i.t.) inhibited the SP-induced SBL responses in a dose-dependent manner. Similar results were obtained in tac1−/− mice, in which slight enhancement of SP responses and their marked inhibition by N/OFQ (1 nmol i.t.) were observed. Because all of these central effects of N/OFQ were completely abolished in NOR−/− mice, it is evident that there are specific actions through NOR. We speculate that the mechanism of this antinociceptive signaling by N/OFQ in the central level is also due to the postreceptor cross-talk, as in the case of peripheral mechanisms.
Thus, it is very likely that N/OFQ has dose-related opposite modulator actions on SP-mediated nociception in nociceptor endings and spinal cord. In a previous report (Mamiya et al., 1998; Nishi et al., 1997), NOR−/− mice showed no significant changes in pain threshold in various nociception tests. Because different modalities of pain may involve different primary afferent systems (Kuraishi et al., 1985; Zimmer et al., 1998), the presynaptic and postsynaptic contributions of NOR system may differ quantitatively and qualitatively with the mixed modalities of pain and may contribute to the sensory discrimination of different nociceptive stimuli. Important issues to discuss involving presynaptic and postsynaptic mechanisms is the source of N/OFQ in the spinal cord and the physiological and pathophysiological roles of N/OFQ. In the limited literature, it was reported that N/OFQ-like immunoreactivity seems to originate from central, rather than primary, afferent neurons (Riedl et al., 1996) and that N/OFQ gene expression was markedly increased by adjuvant-induced inflammation (Andoh et al., 1997). The most interesting issues of the present biphasic actions of N/OFQ would be the physiological and pathophysiological roles. In the present experiments, SP (100 pmol)-induced SBL responses in NOR−/− mice were slightly higher but not significantly different from those in NOR+/+ mice (Fig. 5C). However, SP responses were higher in NOR−/− mice than in NOR+/+ mice when the SBL nociception test was evaluated with different doses of SP and with different evaluation times (H.U., M.I., A.Y., H.T., Y. Nakata, A. Inoue, manuscript in preparation). This suggests that N/OFQ may have some pain inhibitory actions in vivo at the downstream neuronal mechanism of the SP neurons. The present finding that N/OFQ postsynaptically inhibits the actions of SP may be related to this issue. The increased N/OFQ release, possibly mediated through enhanced N/OFQ gene expression during inflammation (Andoh et al., 1997), may stimulate SP release through the facilitative autoreceptor and increase the pain sensation under such a pathophysiological condition. The N/OFQ-induced SP release seen in the present study may be related to the pain facilitative role of this peptide. However, further detailed studies are important for a better understanding of the physiological and pathophysiological roles of N/OFQ.
Footnotes
-
Send reprint requests to: Dr. Hiroshi Ueda, Department of Molecular Pharmacology and Neuroscience, Nagasaki University School of Pharmaceutical Sciences, 1-14 Bunkyo-machi, Nagasaki 852-8521, Japan. E-mail: ueda{at}net.nagasaki-u.ac.jp
-
↵1 JSPS research fellow.
- Abbreviations:
- N/OFQ
- nociceptin/orphanin FQ
- SP
- substance P
- i.pl.
- intraplantar
- i.t.
- intrathecally
- SBL responses
- scratching, biting, and licking
- tac1−/− mice
- tachykinin 1 gene knockout mice
- PLC
- phospholipase C
- PTX
- pertussis toxin
- Received March 15, 1999.
- Accepted June 21, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}