


Abstract
Peripherally administered cytokines profoundly affect the central nervous system (CNS). One mechanism by which they could affect the CNS is by crossing the blood-brain barrier (BBB) to interact directly with brain receptors. Human and murine IL-1α (hIL-1α; mIL-1α) are transported across the murine BBB with a high rate of transport into the posterior division of the septum (PDS), but it is unknown whether BBB transport is relevant to their actions. Here, we injected species-specific blocking antibodies into the PDS to determine whether transport across the BBB is required for blood-borne hIL-1α to affect memory. Retention was impaired in a dose-dependent manner when hIL-1α was injected either by tail vein (i.v.) or into the PDS, with the PDS route being 1000 times more potent. About 70% of the memory impairment induced by i.v. hIL-1α was reversed by injecting a blocking antibody (Ab) specific for hIL-1α into the PDS. This shows that much of the memory impairment induced by hIL-1α depends on its ability to cross the BBB. Ab specific for mIL-1α was also effective in reversing memory impairment, showing that hIL-1α releases mIL-1α from endogenous stores. Whether the mIL-1α was released from peripheral stores, which would require it to cross the BBB, or from brain stores is unknown. In conclusion, these results show that exogenous, blood-borne hIL-1α affects memory by releasing mIL-1α from endogenous stores and by crossing the BBB to act at sites within the PDS.
Cytokines administered peripherally can have profound effects on the central nervous system (CNS) (Plata-Salaman, 1991). For example, peripherally administered interleukin-1 (IL-1) can alter feeding, sleep, glucose levels, thermogenesis, exploratory behavior, drinking, reproductive behavior, and cognitive functions (Spadaro and Dunn, 1990; Kent et al., 1992; Bianchi and Panerai, 1993; Kent et al., 1996; Langhans and Hrupka, 1999). In many cases, the effect of the peripherally administered cytokine can be blocked when antagonists to that same cytokine are given directly into the brain (Kent et al., 1992, 1996;Cremona et al., 1998; del Rey et al., 1998). This shows that the peripheral and central levels are somehow linked, with the peripheral cytokine able to increase its levels in the CNS by some mechanism.
Several mechanisms have been proposed by which a peripheral cytokine could influence its levels in the CNS (Blatteis, 1992; Dantzer, 1994;Faggioni et al., 1995; Kluger et al., 1995; Watkins et al., 1995). Stimulation of the afferent vagus, release of soluble factors at the circumventricular organs, and stimulation of immune cell transport across the blood-brain barrier (BBB) all depend on direct or indirect self-stimulated release of the cytokine into the brain from endogenous stores. Transport of cytokines directly across the BBB has also been demonstrated (Banks et al., 1989) and is the only mechanism in which the exogenous, peripherally administered cytokine directly interacts with brain receptors.
The BBB of the mouse transports human IL-1α (hIL-1α), murine IL-1α (mIL-1α), and murine IL-1β (mIL-1β) by a saturable mechanism in the blood-to-brain direction (Banks et al., 1991). The rate of transport is modest, with about 0.05 to 0.08% of an i.v. dose of hIL-1α taken up per gram of brain. This is similar to or exceeds the level of uptake of many blood-borne substances that affect brain function by crossing the BBB (Banks and Kastin, 1996). However, it remains controversial whether transport across the BBB is relevant to the actions of cytokines.
The transport of hIL-1α is particularly high into the posterior division of the septum (PDS) (Maness et al., 1995). The PDS receives input fibers from the hippocampus and subiculum (Ramon y Cajal, 1901;Raisman, 1966; Herkenham and Nauta, 1977; Swanson and Cowan, 1979;Morley, 1986). Fibers from the PDS project to the habenular nuclei and, either directly or by way of the habenular nuclei, to the interpeduncular nuclei in the midbrain. The PDS, therefore, is positioned to contribute limbic input for a pathway connecting the hippocampus to the midbrain. Functions of the midbrain nucleus are similar to those known to be modified by cytokines, such as feeding, drinking, exploratory and avoidance behaviors, sleep cycles, and reproductive behavior. The hippocampus is important in learning and memory and so the PDS is well positioned to mediate the limbic effects on cognition.
The rodent PDS does not take up human IL-1β (hIL-1β), however (Maness et al., 1995). Unpublished evidence suggests that hIL-1β, unlike hIL-1α, is not transported by a saturable system across the murine BBB. This is consistent with recent evidence of species specificity in rodents, with rat IL-α, but not hIL-1α, being transported by a saturable system across the BBB of the rat (Plotkin et al., 2000).
Here, we determined whether transport across the BBB is relevant to the CNS actions of blood-borne IL-1α. The above-mentioned findings suggest that IL-1α injected directly into the PDS should mimic some of the CNS effects of i.v. IL-1α. We, therefore, compared the effects of injection of hIL-1α either intravenously or directly into the PDS on memory. We also determined the ability of injections into the PDS of antibody (Ab) directed specifically at human or murine IL-1α to block the effects on memory of i.v. hIL-1α. Because the mouse cannot produce hIL-1α, any blockade by the human specific antibody would be a direct demonstration that blood-borne IL-1α had crossed the BBB to influence CNS function. We compared these results to those for i.v.-injected hIL-1β, a cytokine that is not transported across the BBB by a saturable system, when Ab specific to human or murine IL-1β was injected into the PDS.
Materials and Methods
Measurement of Memory in Mice.
All animal studies were carried out with protocols approved according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Experimentally naive 2-month-old ICR mice from our colony were trained in a T-maze footshock avoidance apparatus as previously described (Flood and Morley, 1993; Farr et al., 1999). The maze consisted of a black plastic alley with a start box at one end and two goal boxes at the other. A stainless steel rod floor ran throughout the maze. The start box was separated from the alley by a plastic guillotine door that prevented the mouse from entering the alley until the training trial began. After placing a mouse in the start box, a training trial was begun by simultaneously raising the guillotine door and sounding a buzzer. A footshock was applied 5 s later. The goal box chosen by the mouse on the first trial was designated as “incorrect”. Footshock was continued until the mouse entered the other goal box, which on all subsequent trials was designated as the “correct” choice for that particular mouse. At the end of each trial, the mouse was removed from the goal box and returned to its home cage. A new trial began after placing the mouse back in the start box by sounding the buzzer and raising the guillotine door. Footshock was applied 5 s later unless the mouse had entered its correct goal box. The acquisition training conditions used were an intertrial interval of 45 s, a doorbell type buzzer of 65 dB as the conditioned stimulus warning, and a footshock at 0.4 mA (scrambled grid floor shocker model E13-08; Coulbourn Instruments, Allentown, PA). These parameters were selected based on previous studies that allow the control group to show good retention (mean trials to criterion scores of 6–7) but permitting the detection of impaired retention by memory-impairing drugs. h-IL1α, IL-1β, Ab, goat serum (GS), or vehicle were administered within 3 min after training was complete. Retention was tested 1 week after training and five avoidances in six consecutive trials were used as criterion.
Administration of Cytokines and Antibodies.
Forty-eight hours before training, mice were anesthetized with methoxyflurane, placed in a stereotaxic instrument, and the scalp deflected. A hole was drilled 0.1 mm anterior and 0.5 mm lateral to either side of the bregma. Immediately after training, mice were again placed in the stereotaxic instrument under enflurane anesthesia and 2.0 μl of solution injected over a 60-s period through 30-gauge blunt tubing made of stainless steel (Small Parts, Inc., Miami, FL) attached to a 10-μl syringe with polyethylene-10 tubing and driven by a Sage syringe pump (model 341A) into each PDS within 3 min of training. The depth of the needle was 3.4 mm from the surface of the skull at an angle of 6°. The site of the injection was confirmed histologically based on a mouse brain stereotaxic atlas (Slotnick and Leonard, 1975).
For tail vein injections, mice were anesthetized with enflurane immediately after training. In the studies in which the mice received both a tail vein and a septal injection, mice received the septal injection first and received the tail vein injection while still in the stereotaxic instrument and under anesthesia.
Cytokines and species-specific antibodies were purchased from R & D Systems (Minneapolis, MN). All antibodies directed at cytokines were IgG antibodies raised in goat against Escherichia coli-derived recombinant cytokines, selected for their ability to neutralize bioactivity, and with endotoxin levels of less than 10 ng/mg antibody. Ab-h1α (catalog number AB-200-NA, lot number AL06) has a neutralization dose (ND50, concentration of antibody required to yield one-half-maximal inhibition of the [3H]thymidine incorporation by murine T-helper D10.G4.1 cells when the cytokine is present at a concentration just high enough to elicit a maximum response) of 14 ng/ml in the presence of 50 pg/ml hIL-1α (Ab/cytokine ratio of 280:1) and does not neutralize the biological activity of recombinant mIL-1α, hIL-1β, or mIL-1β. Ab-m1α (catalog number AB-400-NA, lot number BM03) has an ND50 of 30 ng/ml in the presence of 50 pg/ml mIL-1α (Ab/cytokine ratio of 600:1) and does not neutralize the biological activity of recombinant hIL-1α, mIL-1β, or hIL-1β. Ab-h1β (catalog number MAB201, lot numberAWE05) has an ND50 of 2 ng/ml in the presence of 50 pg/ml hIL-1β (Ab/cytokine ratio of 40:1) and does not neutralize the biological activity of recombinant mIL-1β, hIL-1α, or mIL-1α. Ab-m1β (catalog number AF-401-NA, lot number NP07) has an ND50 of 10 ng/ml in the presence of 50 pg/ml mIL-1β (Ab/cytokine ratio of 200:1) and does not neutralize hIL-1β. The cytokines injected into mice were recombinant (E. coli-derived) with endotoxin levels of less than 0.1 ng/μg cytokine, greater than 97% pure, and carrier-free. Goat serum (Vector Laboratories, Burlingame, CA) was used as a control for the blocking antibodies.
Permeability of Human IL-1β across Murine BBB.
The hIL-1β was labeled with 131I by the iodobead method (Pierce Chemical, Rockford, IL) and purified on a column of G10 Sephadex. Male ICR mice were anesthetized with urethane and the left jugular vein and right carotid artery exposed. The I-hIL-1β (105 cpm) was injected into the jugular vein in 0.2 ml of lactated Ringer's solution with 1% bovine serum albumin. In one-half of the mice, 1 μg/mouse was included in the i.v. injection. Blood was taken from the carotid artery 1 to 10 min after i.v. injection and the mouse decapitated immediately after taking the blood. Serum was obtained by centrifuging the whole blood at 5000gfor 10 min at 4°C. The level of radioactivity in whole brain and serum was measured in a gamma counter. The brain/serum ratio was plotted against exposure time and the unidirectional influx rate (Ki) measured from the following equation:
Statistics.
Means are reported with their standard errors and the n per group. Student's t test was used to compare two means. Analysis of variance (ANOVA) was used to compare more than two means and this was followed by Dunnett's ttest comparing the indicated group to the other groups. Ap < 0.05 was taken as indicating a statistically significant difference. The regression lines for brain/serum ratios versus exposure times were calculated with the Prism 3.0 statistical software package and are reported with their n, the correlation coefficient (r), and p value. Regression lines were compared for statistical differences with the Prism 3.0 package as well. Slopes were first compared and if no statistical differences were found, the intercepts were then compared.
Results
Preliminary studies indicated that 1 ng/PDS of hIL-1α injected after training produced a high degree of impairment in retention. A dose-response curve was constructed by injecting doses of hIL-1α ranging from 0.03 to 1.0 ng/PDS (Fig. 1) and a control dose of normal saline (NS), with an n of 7 to 10 mice/group. The ANOVA showed there were statistically significant differences among the groups [F(4,42) = 6.25,p < 0.001] and Dunnett's t test showed that the control was significantly different from the 0.1 (p < 0.05), 0.3 (p < 0.05), and 1.0 (p < 0.01) ng/PDS doses of hIL-1α.

Human IL-1α effects on retention: dose-response curves. The dose of hIL-1α given by i.v. injection is in units of μg/mouse (open circles, solid line) and the dose injected into each PDS was in units of ng/PDS (open squares, broken line). *p < 0.05 compared with 0 dose; **p < 0.01 compared with 0 dose.
Preliminary studies indicated that 1 μg/mouse of hIL-1α injected by tail vein after training produced a high degree of impairment in retention. A dose-response curve was constructed by injecting doses of hIL-1α ranging from 0.03 to 1.0 μg/mouse (Fig. 1) and a control dose of NS, with an n of 9 to 10/group. The ANOVA showed there were statistically significant differences among the groups [F(4,43) = 7.25, p < 0.001] and Dunnett's t test showed that the control was significantly different from the 0.3 (p < 0.05) and 1.0 (p < 0.01) μg/mouse doses of hIL-1α. Figure 1shows 0.3 ng/PDS produced about the same degree of impairment in retention as did the i.v. dose of 0.3 μg/mouse.
We determined whether species-specific antibodies injected into the PDS could reverse the impairment in retention induced by i.v. hIL-1α (Fig. 2). This and the other antibody studies are outlined in Table 1. We chose the i.v. dose of 0.3 μg/mouse of hIL-1α because it produced both a statistically significant and a submaximal response. We assumed, based on Fig. 1, if the effect on memory was due to the ability of hIL-1α to cross the BBB, there should be about 0.3 ng/PDS of hIL-1α to neutralize. We calculated a need to inject 1000-fold more antibody on a weight basis (100:1 antibody-to-cytokine ratio and 10-fold more because the molecular weight of the antibody is about 10 times greater than the cytokine), which is 0.3 μg/PDS. Therefore, we injected 0.3 μg/mouse of hIL-1α i.v. and 0.3 μg/PDS of antibody. The ANOVA showed statistical differences among the groups: F(4,51) = 7.43, p < 0.001. The post test showed the group receiving hIL-1α/GS was different from all other groups atp < 0.05. Ab-h1α and Ab-m1α each produced about a 70% reversal of the hIL-1α-induced impairment in retention and together they produced about a 97% reversal.
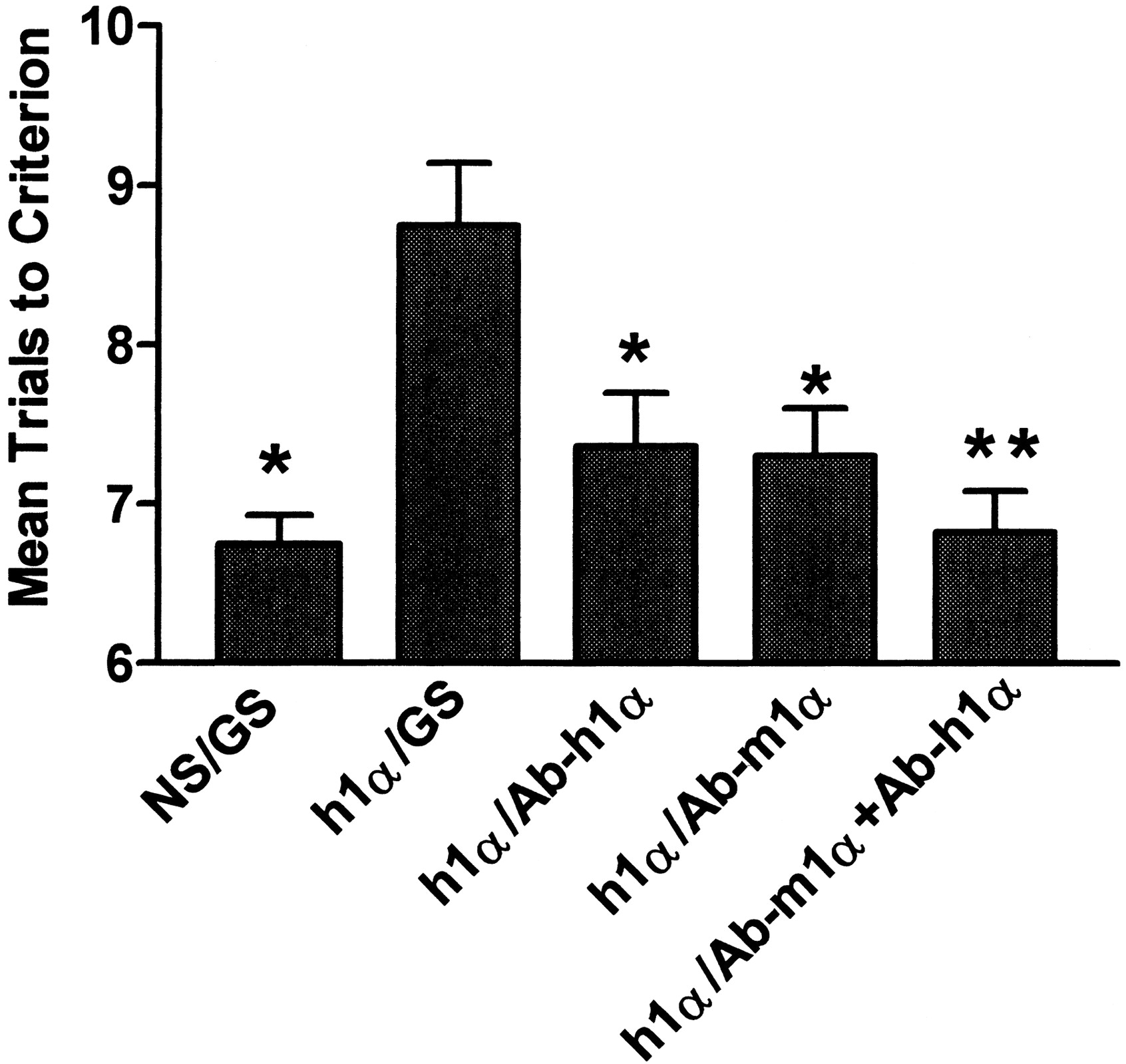
Effects of PDS injection of species-specific antibodies on impairment of retention induced by i.v. hIL-1α. The substance injected intravenously (NS; h1α is hIL-1α) is shown to the left of the slash and the substance injected into the PDS (GS; Ab-h1α is antibody specific for hIL-1α; Ab-m1α is antibody specific for mIL-1α) is shown to the right. Ab-h1α and Ab-m1α were equally effective at inhibiting the memory-impairing effect of i.v. hIL-1α. *p < 0.05 and **p < 0.01 compared with h1α/GS.
Summary of main experimental paradigms and results
We used hIL-1β to determine whether similar results could be produced for a cytokine not crossing the BBB by a saturable transporter. We confirmed previous, unpublished results here by showing that I-hIL-1β uptake by brain was not saturated by inclusion of 1 μg/mouse of unlabeled hIL-1β in the i.v. injection:Ki = 0.59 ± 0.13 μl/g-min (I-hIL-1β only) versus 0.61 ± 0.09 μl/g-min. Figure3 shows that, just as for hIL-1α, hIL-1β was able to impair retention after either i.v. (ttest = 2.4, p < 0.05) or PDS injection (t test = 7.1, p < 0.01). We then determined whether 0.3 μg/PDS of Ab-m1β or Ab-h1β could reverse the effect of i.v. hIL-1β (0.3 μg). The ANOVA showed a significant effect [F(2,24) = 7.9, p < 0.01] and the post test showed that Ab-m1β (p < 0.01) but not Ab-h1β blocked the effects of hIL-1β on retention (Fig.4).

Human IL-1β impairs retention after i.v. or PDS injections. The dose was 1 μg/mouse i.v. or 1 ng/PDS. *p < 0.05 and **p < 0.01 in comparison with respective NS group.

Effects of PDS injection of species-specific antibodies on impairment of retention induced by i.v. hIL-1β. The substance injected intravenously (h1β is hIL-1β) is shown to the left of the slash and the substance injected into the PDS (Ab-h1β is antibody specific for hIL-1-β; Ab-m1β is antibody specific for mIL-1β) is shown to the right. Only Ab-m1β inhibited the memory-impairing effects of i.v. hIL-1β.
Discussion
Here, we showed that a CNS effect of an intravenously administered cytokine was dependent in large part on the ability of that cytokine to cross the BBB. Specifically, we showed that memory impairment induced by i.v. human IL-1α can be largely blocked by injection into the PDS of an antibody specific for human IL-1α.
We first determined whether hIL-1α could impair memory. Cytokines, including the interleukins, have numerous effects on the CNS. Many of these effects are related to the induction of sickness behavior, including impairment of cognitive processes (Dantzer and Kelley, 1989). Therefore, it was reasonable that learning and memory might be impaired by IL-1α. Additionally, IL-1β, which uses the same receptor as IL-1α, also impairs learning and memory and has elevated levels in the hippocampus in conditions associated with sickness behavior (Pugh et al., 2001).
We chose not to test learning because the acute effects of IL-1α could be a confounder (McGaugh, 1973). To assess effects on learning, a substance is given immediately before testing. But the sickness behavior induced by IL-1α could lead to a decline in motivation, rather than an impairment in the ability to learn. To assess effects on memory, a substance is given after learning has occurred and memory is tested after the acute effects have passed, a week after IL-1α administration in this case. Therefore, the mice were not exposed to the acute effects of IL-1α during training or testing when sickness behavior could have interfered with performance.
Any effect on memory resulting from the transport of IL-1α across the BBB is likely to occur at an i.v. dose of 1 μg/mouse or less because the BBB transporter is saturated at that dose (Banks et al., 1991). Any higher dosing could further increase brain levels, but only through the much less efficient, nonsaturable extracellular pathways and transmembrane diffusion (Balin et al., 1986; Broadwell and Sofroniew, 1993). We found that 1 μg/mouse of IL-1α given i.v. produced a high level and statistically significant impairment in retention. There was a dose-dependent relationship between the log dose of i.v. IL-1α and the number of trials needed to reach criterion, with the dose of 0.3 μg/mouse producing a statistically significant impairment.
Given that a little less than 0.1% of an i.v. dose of IL-1α enters the brain (Banks et al., 1991), we used 1 ng/mouse as the high dose administered into the PDS. We chose the PDS because of its high rate of blood-to-brain transport for hIL-1α. Transport rates of closely related cytokines, including hIL-1β and IL receptor antagonist, are not especially high (Maness et al., 1995). The PDS is positioned to act as a limbic modulatory component on a pathway connecting the hippocampal formation to midbrain nuclei, areas that affect learning and memory. Other studies indicate that IL-1 receptor ligands may be particularly effective at impairing learning and memory tasks mediated through the hippocampus (Pugh et al., 2001); the memory paradigm we used here is considered to be a hippocampal task. We found that injection of hIL-1α into the PDS produced a dose-response curve very similar to i.v. injection of hIL-1α, except the doses were 1000-fold lower (Fig. 1).
Injection of a substance in a small volume directly into brain tissue produces a pattern of distribution entirely different from an i.c.v. injection. After i.c.v. injection, a substance diffuses throughout the CSF space of the brain (Davson and Segal, 1996; Maness et al., 1998). Penetration into brain tissue occurs from all brain/CSF contacting surfaces, but relevant concentrations in brain tissue seldom extend even to 0.1 cm from the contacting surface (Maness et al., 1996,1998). No saturable brain-to-blood transport system has been described for any cytokine to date, but cytokines injected into the lateral ventricle do enter the blood as they are reabsorbed with the CSF at the arachnoid villi. This slow brain-to-blood transfer combines with a relatively long half-life in blood and a relatively small volume of distribution to produce significant elevations in the blood. In fact, the level of exogenous IL-6, tumor necrosis factor-α, and IL-1α in the blood and peripheral tissues about 20 min after injection is higher after i.c.v. than after i.v. injection (Chen et al., 1997; Chen and Reichlin, 1998; Di Santo et al., 1999). A real potential exists that the effects seen with i.c.v. injection are mediated through receptors outside the CNS (Clark et al., 1983; Bodnar et al., 1989; Yao et al., 1993). In contrast, a substance injected directly into brain tissue in a small volume diffuses very slowly from its initial site of injection (Cserr and Berman, 1978; Cserr, 1984; de Lange et al., 1993; Banks et al., 1994). Brownian motion is the primary force behind diffusion with local metabolism, physical obstruction by cells or fiber tracts, and reabsorption across capillaries acting to further impede diffusion within brain tissue. Therefore, hIL-1α injected into the PDS, in comparison with hIL-1α injected i.c.v., should be available to receptors only in the area of the PDS.
The above-mentioned results show that mechanisms exist by which hIL-1α administered i.v. or into the PDS can impair memory. They do not demonstrate that hIL-1α given i.v. is acting at receptors behind the BBB, although the relative magnitude of the doses is consistent with that mechanism. To determine the extent to which hIL-1α given i.v. exerts its effects after crossing the BBB, we gave antibodies specific to human (Ab-h1α) or murine (Ab-m1α) IL-1α directly into the PDS. Because the only source of hIL-1α in the mouse was the exogenous, peripherally administered cytokine, Ab-h1α could reverse memory impairment only to the extent that hIL-1α crossed the BBB to enter the PDS. We chose 0.3 μg/mouse as the i.v. dose to give, which produced a statistically significant, submaximal effect on the linear portion of the dose-response relation. Based on the similarity in magnitude of effect as shown in Fig. 1, this dose would be expected to produce a level of 0.3 ng/PDS if the effect were totally caused by IL-1α released or transported into the PDS. We chose an Ab dose of 3 μg/PDS, which would give an Ab/cytokine molar ratio of 100:1, because IgG Ab has a molecular weight about 10 times that of IL-1α. The Ab/cytokine mass ratio of 1000:1 is somewhat larger than the Ab/cytokine ratios based on the ND50 values, which ranged from 40:1 to 600:1 and so should have inhibited more than 50% of any effect caused by the specific cytokine. We found that about 70% of the memory impairment induced by i.v. hIL-1α was reversed by Ab-h1α (Fig. 2). This shows that much of the impairment in memory was attributable to the transport of hIL-1α across the BBB. Goat serum, which contains nonspecific antibodies, was used as control and did not reverse the effect of hIL-1α on memory.
Memory impairment was also reversed by Ab-m1α (Fig. 2). This is consistent with hIL-1α impairing memory through one of the mechanisms whereby IL-1α mediates its own release from endogenous stores. These results do not indicate whether the mIL-1α blocked by Ab-m1α was released from stores within the brain or from peripheral stores. Release from peripheral stores would have required the mIL-1α to cross the BBB to reach the PDS. When Ab-m1α and Ab-h1α were given together, the effects of i.v. hIL-1α were almost totally blocked, strongly suggesting that the memory-impairing effects of IL-1α are primarily mediated through the PDS.
To test the specificity of these findings, we determined whether hIL-1β could impair memory and whether any such effects could be blocked with Ab-h1β or Ab-m1β. Previous studies have shown that hIL-1β does not enter the PDS (Maness et al., 1995), although whether mIL-1β does has not been determined. We also confirm previous, unpublished findings that hIL-1β, unlike mIL-1β, does not cross the BBB of the mouse in a saturable manner. Here, I-hIL-1β did have a measurable uptake, but in the absence of self-inhibition this most likely represents the transport of degradation products across the BBB. Therefore, any effect of i.v. hIL-1β would not be expected to be caused by transport of exogenous cytokine across the BBB. We found that hIL-1β impaired retention after either i.v. injection or injection into the PDS (Fig. 3). The effects were at the same doses as for hIL-1α and were of the same magnitude. However, 0.3 μg/PDS of Ab-h1β was unable to block the impairment in retention induced by i.v. hIL-1β, even though this antibody was the most potent based on the ND50 (Fig. 4). This clearly demonstrates that hIL-1β is not impairing memory by crossing the BBB to act within the PDS. Ab-m1β injected into the PDS blocked nearly all of the memory-impairing effect of i.v. hIL-1β. This demonstrates both that the PDS is an important site for the mediation of the central effects of blood-borne IL-1β and also that the effects on memory result from the self-release of IL-1β from endogenous stores. It is not clear whether the source of the mIL-1β acting at the PDS was from peripheral stores, which would require transport across the BBB, or from brain stores released directly into the PDS.
In conclusion, we showed that hIL-1α and hIL-1β impair memory after either i.v. injection or injection into the PDS. For hIL-1α, which crosses the murine BBB by a saturable transport system, the memory-impairing effect after i.v. administration is partially blocked by injection into the PDS of antibodies specific for either hIL-1α or mIL-1α. For hIL-1β, which unlike hIL-1α and mIL-1β does not cross the murine BBB by a saturable system, the memory-impairing effect after i.v. administration is blocked by antibody specific for mIL-1β but not by antibody specific for hIL-1β. These results show the PDS to be an important site for mediating the effects of IL-1α and IL-1β. The results also show the importance of self-release from endogenous stores in mediating the effects of IL-1α and IL-1β, although it is not clear whether the endogenous cytokines must cross the BBB to induce their effects at the PDS. The results clearly show that the memory-impairing effect of hIL-1α given i.v. into the mouse is partially mediated by its ability to cross the BBB at the PDS.
Acknowledgments
We thank Sandra M. Robinson for technical assistance.
Footnotes
-
This study was supported by Veterans Affairs Merit review, R01 AA12743, and R01 NS41863.
- Abbreviations:
- CNS
- central nervous system
- IL-1
- interleukin-1
- BBB
- blood-brain barrier
- hIL-1α
- human interleukin-1α
- mIL-1α
- murine interleukin-1α
- mIL-1β
- murine interleukin-1β
- PDS
- posterior division of the septum
- hIL-1β
- human interleukin-1β
- Ab
- antibody
- GS
- goat serum
- Ab-h1α
- blocking antibody specific for human IL-1α
- Ab-h1β
- blocking antibody specific for human IL-1β
- Ab-m1α
- blocking antibody specific for murine IL-1α
- Ab-m1β
- blocking antibody specific for murine IL-1β
- ND50
- concentration of antibody required to yield one-half-maximal inhibition of the [3H]thymidine incorporation by murine T-helper D10.G4.1 cells when the cytokine is present at a concentration just high enough to elicit a maximum response
- I-hIL-1β
- hIL-1β radioactively labeled with131I
- Ki
- unidirectional influx rate
- ANOVA
- analysis of variance
- NS
- normal saline
- CSF
- cerebrospinal fluid
- Received June 22, 2001.
- Accepted August 3, 2001.
- U.S. Government





{kind=link}
{kind=link}
{kind=link}
{kind=link}