


Abstract
This study was aimed at clarifying the gender differences in the urinary excretion of organic anions and the gene expression of organic anion transporters in rats. The renal clearance with regard to the plasma concentration (CLurine,p) of taurocholate, dibromosulfophthalein (DBSP), and zenarestat, all substrates and/or inhibitors of organic anion transporting polypeptide 1 (Oatp1), was much higher in female than in male rats. The following results imply that the transport system(s) for the reabsorption of zenarestat across the luminal side exhibits a gender difference: 1) the renal uptake clearance assessed by an in vivo integration plot analysis of zenarestat from the blood side does not show any clear gender differences; 2) the renal clearance with regard to the kidney concentration (CLurine,k) of zenarestat in female rats was approximately 30 times higher than in male rats; and 3) both CLurine,p and CLurine,k were increased in male rats by the coinfusion of DBSP, which is an inhibitor of organic anion transporters. Northern and Western blot analyses confirmed a previous finding that the gene expression of Oatp1, which is localized at the apical plasma membrane of the kidney, was much higher in the kidneys of male rats. Overall, a gender difference in urinary excretion is commonly observed for several organic anions, including Oatp1 substrates and inhibitors, and Oatp1 and/or transporters that have a similar substrate specificity to Oatp1 could be involved in such a phenomenon involving its substrates.
The kidney plays an important role in the elimination of therapeutic agents from the body, and the renal excretion of xenobiotics involves at least three processes, including glomerular filtration, secretion, and reabsorption via the renal tubules. Several types of xenobiotic transporters have been recently identified and are suggested to be primarily involved both in the renal secretion and reabsorption of organic anions and cations. These include the organic anion transporter (OAT) family, the organic anion transporting polypeptide (OATP), and the organic cation transporter (Oct) family (Sekine et al., 2000; van Aubel et al., 2000; Dresser et al., 2001). As far as the OATP family in rats is concerned, Oatp1 (gene symbol, Slc21a1), Oatp3 (Slc21a7), Oat-K1 (Slc21a4), and Oat-K2 are expressed in the kidney (Bergwerk et al., 1996; Masuda et al., 1997, 1999; Abe et al., 1998), whereas in the case of the OAT family, both Oat1 (Slc22a6) and Oat3 (Slc22a8) have been reported to be expressed in rat kidney (Sekine et al., 1997; Sweet et al., 1997; Kusuhara et al., 1999). In humans, a strong mRNA band for OAT4 (Slc22a11) has also been detected in the kidney (Cha et al., 2000). Oat1 and Oat3 are localized at the basolateral membrane in the kidney, whereas the gene products of Oatp1, Oat-K1, and Oat-K2 have been identified at the brush-border membrane (Bergwerk et al., 1996;Masuda et al., 1997, 1999; Tojo et al., 1999; Hasegawa et al., 2002). As ATP-dependent primary active transporters, P-glycoprotein (Abcb1), multidrug resistance-associated protein 1 (Mrp1) (Abcc1), and Mrp2 (Abcc2) are expressed in the kidney (Thiebaut et al., 1987; Schaub et al., 1997; Peng et al., 1999). Oligopeptide transporters (PEPT1 and PEPT2), which accept β-lactam antibiotics and angiotensin-converting enzyme inhibitors as substrates, are also expressed in the kidney (Fei et al., 1994; Saito et al., 1996).
Some xenobiotic transporters have been reported to exhibit gender differences in their expression. For example, the gene product of Oct2 (Slc22a2) in male rat kidney is higher than that in female rat kidney, whereas the mRNA expression of Oct1 (Slc22a1), Oct3 (Slc22a3), and Oat1 does not show any gender difference (Urakami et al., 1999, 2000). The expression of Oct2 in the kidney is possibly under hormonal regulation and is increased by testosterone treatment. Gender differences and hormonal regulation have recently been reported in the expression of Oat2 and Oat3 (Kobayashi et al., 2001). The expression of Oatp1 mRNA is also higher in male than in female rat kidney and regulated by testosterone (Lu et al., 1996), although the relevance of such gender differences in terms of the renal excretion of organic anions is still unclear.
Oatp1 is composed of 670 amino acids and has 12 membrane-spanning domains (Jacquemin et al., 1994). It has been shown that Oatp1 mRNA is localized in the liver, kidney, brain, lung, skeleton, muscle, and proximal colon (Jacquemin et al., 1994). Immunomorphological investigations have revealed that the gene product detected by Oatp1 antibody is localized at the apical plasma membrane of the kidney in the S3 segment of the proximal tubule of the outer medulla (Bergwerk et al., 1996). Oatp1 is thought to be the multispecific anion transporter that accepts many types of organic anions, including taurocholate (TCA) and other bile acids, as substrates (Eckhardt et al., 1999; Burckhardt and Wolff, 2000). The driving force is considered to be reduced glutathione (Li et al., 1998). Bergwerk et al. (1996) have reported that Oatp1 in the liver is present in the form of a 83-kDa protein, whereas that in the kidney is only 37 kDa under reducing conditions. Thus, the structure of the Oatp1 gene product might differ from organ to organ, although the difference between the liver and kidney in terms of the function of Oatp1 remains to be clarified.
In addition, some organic anions are reported to exhibit a gender difference in their urinary excretion in rats. These include zenarestat, torsemide, eugurane sodium, clentiazem, and nilvadipine metabolites (Kling et al., 1991; Tanaka et al., 1991a; Nakamura et al., 1993; Terashita et al., 1994; Sato et al., 2000). The degree of urinary excretion of these compounds is much smaller in male than in female rats. A series of earlier studies has revealed that the renal clearance of zenarestat, an aldose reductase inhibitor for the treatment of diabetic neuropathy, in female rats is approximately 50 times higher than that in male rats (Tanaka et al., 1991a). Since its unbound fraction in plasma and glomerular filtration rate (GFR) are relatively similar in males and females, the reason for this gender difference cannot be due to a difference in glomerular filtration (Tanaka et al., 1993). Furthermore, the renal clearance of zenarestat in female rats is inhibited by probenecid, which is a typical inhibitor of the renal secretion of organic anions, whereas the renal clearance in male rats is unaffected (Tanaka et al., 1991a). Hypophysectomized male rats and normal female rats exhibit similar urinary excretion of zenarestat, and treatment of male and female hypophysectomized rats with testosterone resulted in the urinary excretion of zenarestat comparable with that by intact adult male rats (Tanaka et al., 1992), suggesting hormonal regulation of its excretion system(s).
In the light of the above findings, we formed the hypothesis that the gender difference in the urinary excretion of some organic anions may be due to a difference in the expression of Oatp1. Since Oatp1 can take up a variety of types of organic anions and is localized at the apical membrane to a higher degree in male rats, the higher urinary excretion of anionic compounds in female rats may be explained by a gender difference in the reabsorption process mediated by Oatp1. To obtain evidence for such a hypothesis, in the present study, we have investigated the existence of gender differences in the urinary excretion of the substrate and inhibitor of Oatp1, TCA, and dibromosulfophthalein (DBSP), respectively. We also report here that zenarestat has an affinity for Oatp1 and that the gender difference in its urinary excretion results from its transport across the apical membranes. Gender differences in the gene expression of other organic anion transporters were also examined in the present study to identify potential candidates that may be involved in the mechanism governing such gender differences in urinary excretion.
Experimental Procedures
Materials.
Zenarestat (FK-366) was donated by Fujisawa Yakuhin Kogyo (Osaka, Japan). Inulin- and HPLC-grade acetonitrile were purchased from Wako Pure Chemical Industries (Osaka, Japan). DBSP was purchased from Société d'Etudes et de Researches Biologiques (Paris, France). Isopropyl ether was purchased from Hayashi Pure Chemical Industries (Osaka, Japan). [3H]Estradiol 17β-d-glucuronide (E217βG; 55 μCi/nmol) and [3H]TCA (3 μCi/nmol) were purchased from PerkinElmer Life Sciences (Boston, MA). All other chemicals and reagents were commercial products of analytical grade. Seven-week-old male and female Sprague-Dawley rats were purchased from Charles River Japan (Tokyo, Japan) and had free access to water and food. The studies reported on in this article have been carried out in accordance with the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by the U.S. National Institutes of Health.
Infusion Study.
The bladder was catheterized using polyethylene tubing (no. 8; o.d. 2.33 mm; Hibiki, Tokyo, Japan). The body temperature of the rats was maintained by keeping them under suitable lighting. Zenarestat dissolved in saline was administered via the femoral vein at a priming dose of 1 mg/kg and continued as a sustained infusion of 3 mg/kg/h in the volume of 6 ml/h. Inulin was administered at a priming dose of 10 mg/kg and continued as a sustained infusion of 20 mg/kg/h. Blood was collected at 30, 60, 90, 120, and 150 min. Urine specimens were collected at 0 to 30, 30 to 60, 60 to 90, 90 to 120, and 120 to 150 min. In the inhibition study, DBSP was injected after mixing it with zenarestat injection solution. The highest dose of DBSP exhibits a transport maximum in its biliary excretion (Goto et al., 2002). The urinary clearance with regard to the plasma concentration (CLurine,p) was calculated as the ratio of the cumulative excreted amount over 150 min to the area under the curve over 150 min (AUC0–150min). The AUC was calculated based on the trapezoidal rule where the initial plasma concentration (C0) was calculated by dividing the loading dose by V0. The urinary clearance with regard to the kidney concentration (CLurine,k) was calculated as the ratio of the urinary excretion rate from 120 to 150 min to the kidney concentration at 150 min. The GFR was assumed to be equal to the total clearance (CLtot) of inulin.
Intravenous Bolus Injection.
Rats underwent bile duct cannulation using polyethylene tubing (PE10; i.d. 0.61 mm; BD Biosciences, Bedford, MA), and the bladder was catheterized as described for the infusion study. DBSP (2 mg/kg) or [3H]TCA (5 μCi/kg) was administered via the femoral vein. Zenarestat (1 mg/kg) was also administered via the femoral vein to rats without bile or bladder cannulation and, then, 1 and 3 min after administration, the animals were killed, and the liver and kidneys were removed immediately (integration plot analysis). The plasma concentration-time profile was fitted to a biexponential equation, and the AUC was estimated by integration. The CLtot was calculated as Dose/AUC0–∞. The CLurine,p was calculated as the ratio of the cumulative excreted amount over 180 min to the AUC0–180min. The V0was calculated by dividing the dose by theC0 calculated from the biexponential equation. The initial slope in a plot (designated as the integration plot) of the amount of zenarestat in tissue/plasma concentration of zenarestat versus the AUC/plasma concentration of zenarestat gives the tissue uptake clearance.
Determination of the Amount of Each Compound.
For the determination of zenarestat, 0.1 M citrate buffer (pH 4.7; 100 μl) was added to 50 μl of rat plasma, followed by 0.5 N HCl (100 μl) and isopropyl ether (400 μl). This sample was shaken for 5 min, then centrifuged for 2 min at 3000 rpm. The organic layer (300 μl) was added to 0.01 N NaOH (50 μl), shaken for 5 min, and then centrifuged for 2 min at 3000 rpm. The organic layer was removed and the aqueous layer (20 μl) was added to 1 M phosphate buffer (pH 5.5) (30 μl), and 20 μl were subjected to HPLC. Kidney was homogenated in 2.5 volumes of 0.1 M citrate buffer (pH 4.7). Then, 100 μl of urine or 300 μl of kidney homogenate were used instead of 50 μl of plasma. HPLC conditions were as follows: column, Nucleosil 5C18 (5 μm; GL Sciences, Tokyo, Japan), 4.0 mm i.d. × 15 cm; guard column, Nucleosil 5C18 (5 μm; GL Sciences), 4.0 mm i.d. × 1 cm; mobile phase, 20 mM phosphate buffer (pH 6.0)/acetonitrile (70/30); flow rate, 1 ml/min; column temperature, 40°C. Inulin in plasma and urine was analyzed by spectrophotometric assay using a modification of the method ofHeyrovsky (1956). Briefly, a 10-μl sample was added to 2 μl of 0.5% β-indolylacetic acid in ethanol and 80 μl of concentrated hydrochloric acid. After 24 h at ambient temperature, the absorption of the samples was measured at 530 nm. [3H]TCA was determined by counting the radioactivity in a Tri-Carb liquid scintillation spectrometer (Packard BioScience, Meriden, CT). For the determination of DBSP, plasma and urine specimens were diluted with 50 mM Tris/HCl buffer (pH 7.4) and then made alkaline by the addition of NaOH, the concentration of DBSP being determined in a dual wavelength spectrophotometer (Hitachi, Tokyo, Japan) at 575 and 640 nm.
Determination of the Plasma Unbound Fraction (fu) and Blood to Plasma Concentration Ratio (Rb).
[3H]TCA or DBSP was added to blank whole blood and incubated for 15 min, and the drug concentration in the whole blood (50 μl) was determined. The residual blood was then centrifuged for 5 min at 10,000 rpm (TOMY RL-100; Tomy Seiko, Tokyo, Japan), and the drug concentration in 50 μl of supernatant (plasma) was determined. The residual plasma was centrifuged through an Amicon YMT membrane (MPS-3 system; Millipore Corporation, Bedford, MA) at 3000 rpm for 5 min at 37°C. All the plasma protein binding data were normalized with respect to the filter blank.
Uptake Studies in Oatp1-Transfected LLC-PK1Cells.
Uptake of [3H]E217βG was determined as described previously (Sugiyama et al., 2001). Expression of Oatp1 was induced by incubating cells for 24 h in the presence of sodium butyrate (5 mM) before starting the transport experiments (Eckhardt et al., 1999). All transport assays were performed in Krebs-Henseleit buffer (120 mM NaCl, 23.8 mM NaHCO3, 4.83 mM KCl, 0.96 mM KH2PO4, 1.20 mM MgSO4, 12.5 mM HEPES, 5 mM glucose, and 1.53 mM CaCl2 adjusted to pH 7.4). The uptake was calculated as the ratio of the intracellular amount to the concentration in the medium. To obtainKi, the uptake of [3H]E217βG within the linear range (2 min) was measured and fitted to the following equation:
Northern Blot Analysis.
Northern blot analysis was performed as previously described (Ogawa et al., 2000). A 2-μg sample of mRNA, extracted from liver and kidney using ISOGEN (Nippon Gene, Tokyo, Japan) and Oligotex dT30 super (Takara, Tokyo, Japan) according to the manufacturer's protocol, was electrophoresed on 1% agarose/formaldehyde gel and transferred to a nitrocellulose filter. The filter was hybridized at 42°C with a full-length cDNA of Oat2, Oat3, and Mrp2, and 2190 to 2700 bp of cDNA of Oatp1 and 2700 to 3610 bp of cDNA of Oatp2 randomly labeled with [32P]dCTP. The filter was then washed with 0.1% standard saline citrate/0.1% SDS at 55°C. No cross-reaction was observed for this Oatp1 probe with cRNA of Oatp2 and Oatp3 or for this Oatp2 probe with cRNA of Oatp1 and Oatp3.
Western Blot Analysis.
Anti-rat Oatp1 serum was previously raised against a fusion protein with the C-terminal 40 amino acids and maltose-binding protein as described in Eckhardt et al. (1999). Membrane fractions were prepared as previously described (Ogawa et al., 2000), diluted with the sample buffer, with or without reducing agent, and denatured at 95°C for 2 min before separation on 3.75% stacking and 10% resolving SDS-polyacrylamide gels. Proteins were transferred electrophoretically to polyvinylidene difluoride membranes using a blotter (Trans-Blot; Bio-Rad, Richmond, CA) at 15 V for 1 h. The membrane was blocked with Tris-buffered saline containing 0.05% Tween 20 (TBST) and 5% bovine serum albumin for 1 h at room temperature. After washing with TBST (three times for 10 min), the membrane was incubated with anti-rat Oatp1 serum (dilution 1:500), allowed to bind 125I-labeled sheep anti-rabbit IgG antibody diluted 1:200 in TBST containing 5% bovine serum albumin for 1 h at room temperature, and washed with TBST (three times for 5 min). Then, the membranes were exposed to Fuji imaging plates (FujiFilm, Kanagawa, Japan) for 3 h at room temperature and analyzed with an imaging analyzer (BAS 2000; FujiFilm).
Results
Gender Difference in the Urinary Excretion of Organic Anions.
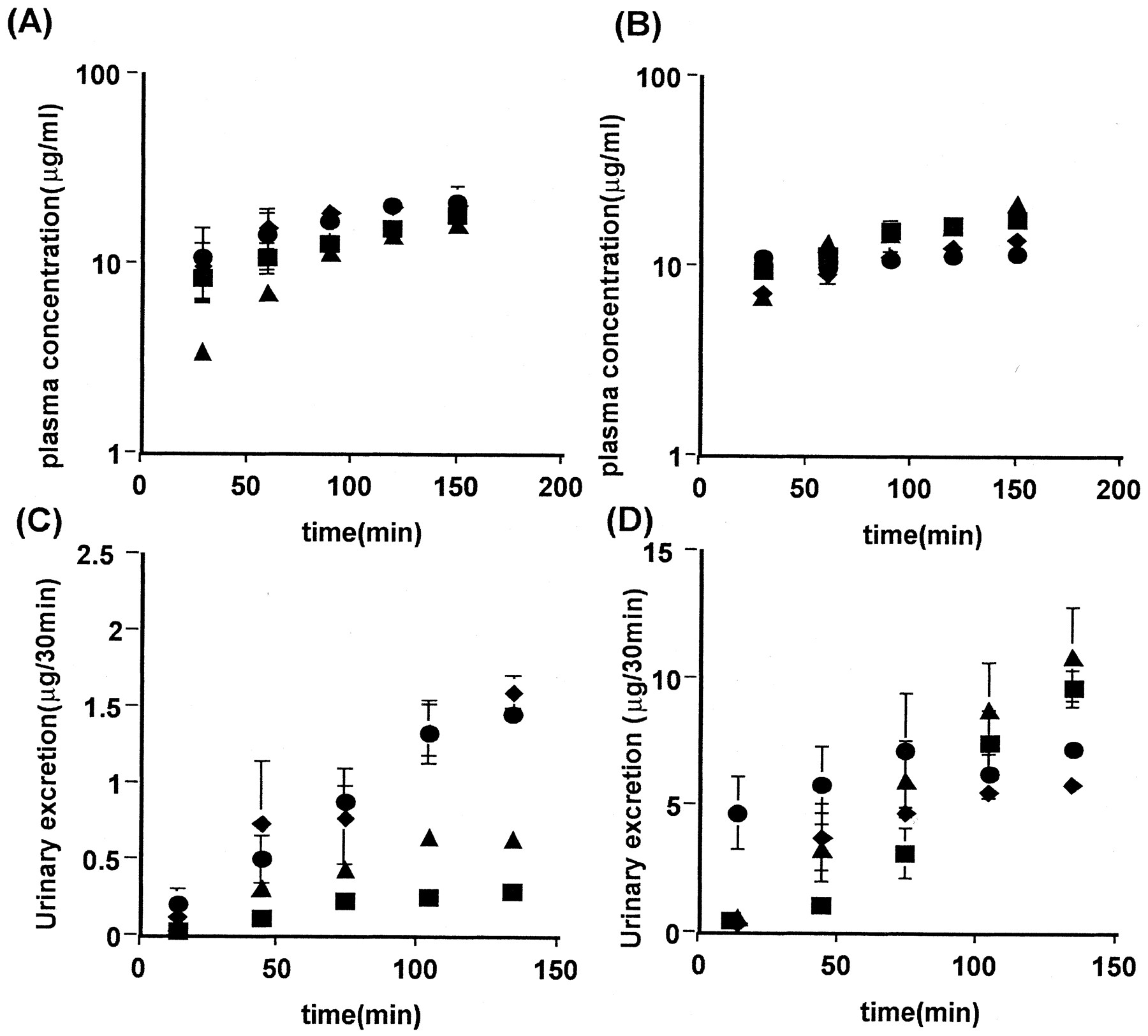
The plasma concentration profile of TCA and DBSP showed a minimal gender difference, whereas their urinary excretion over 180 min in female rats was 3.8 and 7.7 times higher than in males, respectively (Fig. 1). Neither the CLtot, the fu, nor the Rb of TCA and DBSP showed any evidence of a gender difference (Table 1). The CLurine,p of TCA and DBSP in female rats was 4.4 and 7.7 times higher than in male rats, respectively (Table1). During the constant infusion of zenarestat alone, its urinary excretion rate was also higher in female than in male rats (Fig. 2, C and D), the CLurine,p in female rats being 18 times higher than that in male rats (Table 2), whereas the plasma concentration profile showed only a minimal gender difference (Fig. 2, A and B).

Time profiles of the plasma concentration (A and C) and cumulative urinary excretion (B and D) of [3H] TCA (A and B) and DBSP (C and D) in male and female rats after intravenous bolus injection. Male (▪) and female (♦) rats received an intravenous bolus injection of [3H]TCA (5 μmol/kg) or DBSP (2 mg/kg). Each point and vertical bar represent the mean ± S.E. of three or four rats.
Pharmacokinetic parameters of [3H]TCA and DBSP

Effect of DBSP on the plasma concentration (A and B) and urinary excretion (C and D) of zenarestat in male (A and C) and female (B and D) rats. Rats received an intravenous priming dose of zenarestat (1 mg/kg) and a subsequent infusion of both 3 mg/h/kg zenarestat and DBSP at 0 (▪), 30 (▴), 75 (♦), or 150 (●) μmol/h/kg. Each point and vertical bar represent the mean ± S.E. of four or five rats.
Pharmacokinetic parameters of zenarestat
Gender Difference in the Transport of Zenarestat across the Apical Side.
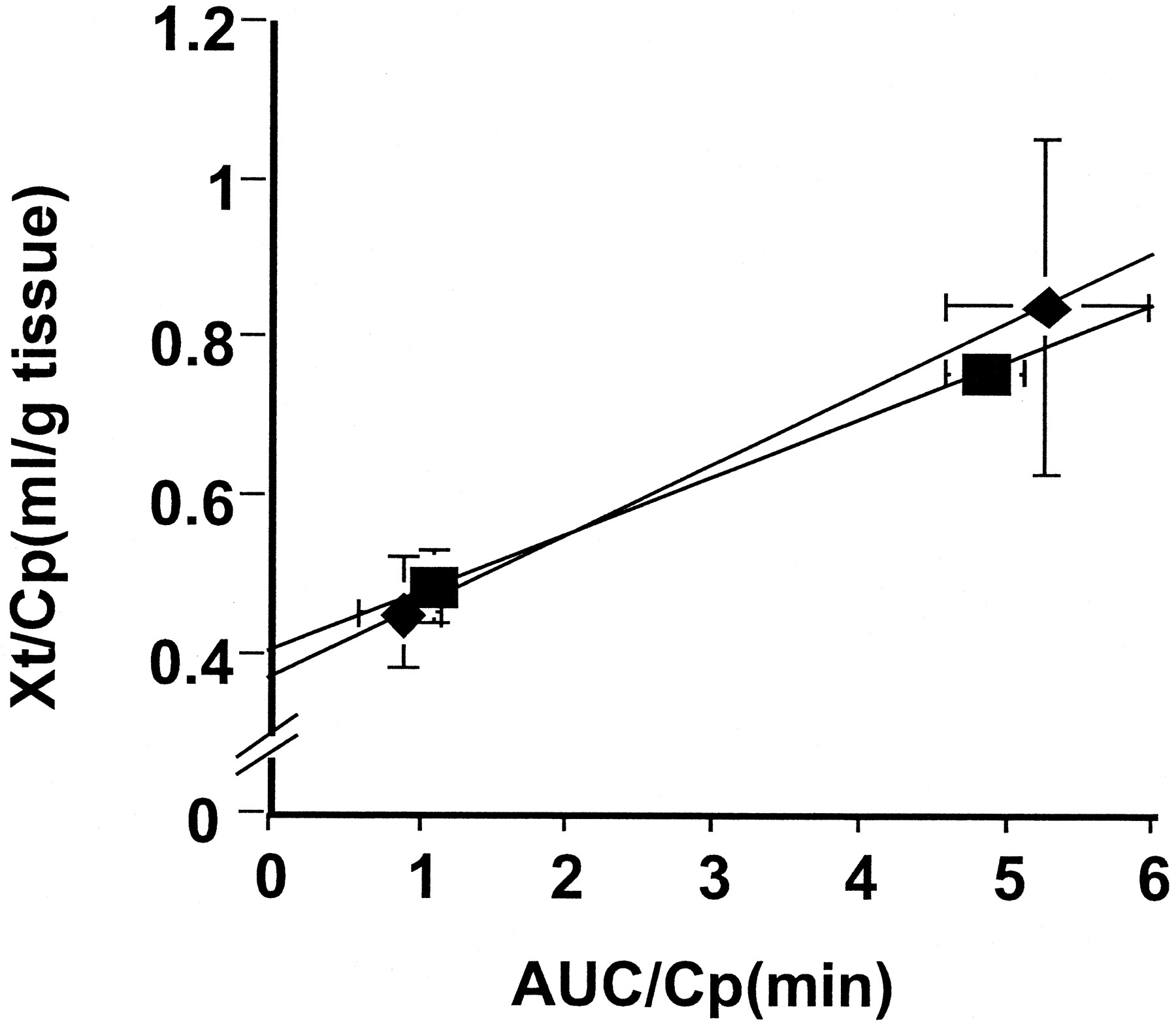
To examine the gender difference in the transport of zenarestat across the antiluminal membranes, its renal uptake after 1 and 3 min following a bolus injection was examined (Fig.3). The tissue-to-plasma concentration ratio was similar for the two genders (Fig. 3). The slope of the integration plot in male and female rats (0.0731 and 0.0892 ml/min/g tissue, respectively) was similar (Fig. 3).

Integration plot of the renal uptake of zenarestat. The time profiles of the plasma and tissue concentrations of zenarestat were measured after its intravenous bolus injection (1 mg/kg) in male (▪) and female (♦) rats. Data are expressed as integration plots. The initial slope represents the CLuptake. Each point and the vertical and horizontal bars represent the mean ± S.E. of three independent experiments.
Next, to analyze the transport clearance across the luminal side, the CLurine,k was examined when it was found that the clearance in female rats was 31 times higher than that in male rats (Table 2). The effect of DBSP on plasma zenarestat concentrations was not as marked (Fig. 2, A and B), whereas DBSP increased the urinary excretion of zenarestat in male rats (Fig. 2C). Both CLurine,p and CLurine,kwere increased by DBSP in male rats, the CLurine,p and CLurine,k at 150 μmol/h/kg DBSP being 2.6 and 5.7 times higher than in the control (Table 2). The CLurine,p was increased, whereas the CLurine,k was decreased by DBSP in female rats, although the change compared with the control was less than 50% (Table 2). Such a phenomenon might be explained by inhibition of efflux transport from tubular cells to the luminal and antiluminal spaces. Further studies are required to examine such a hypothesis. In the presence of 150 μmol/h/kg DBSP, the CLurine,k in female rats was 3.7 times higher than that in male rats (Table 2). Thus, the gender difference in CLurine,k in the presence of DBSP was not as marked compared with the control.
Zenarestat Has an Affinity for Oatp1.
The uptake of [3H]E217βG, a typical substrate of Oatp1, by Oatp1-transfected LLC-PK1 cells was inhibited by zenarestat with a Ki value of 8.97 μM (Fig. 4).
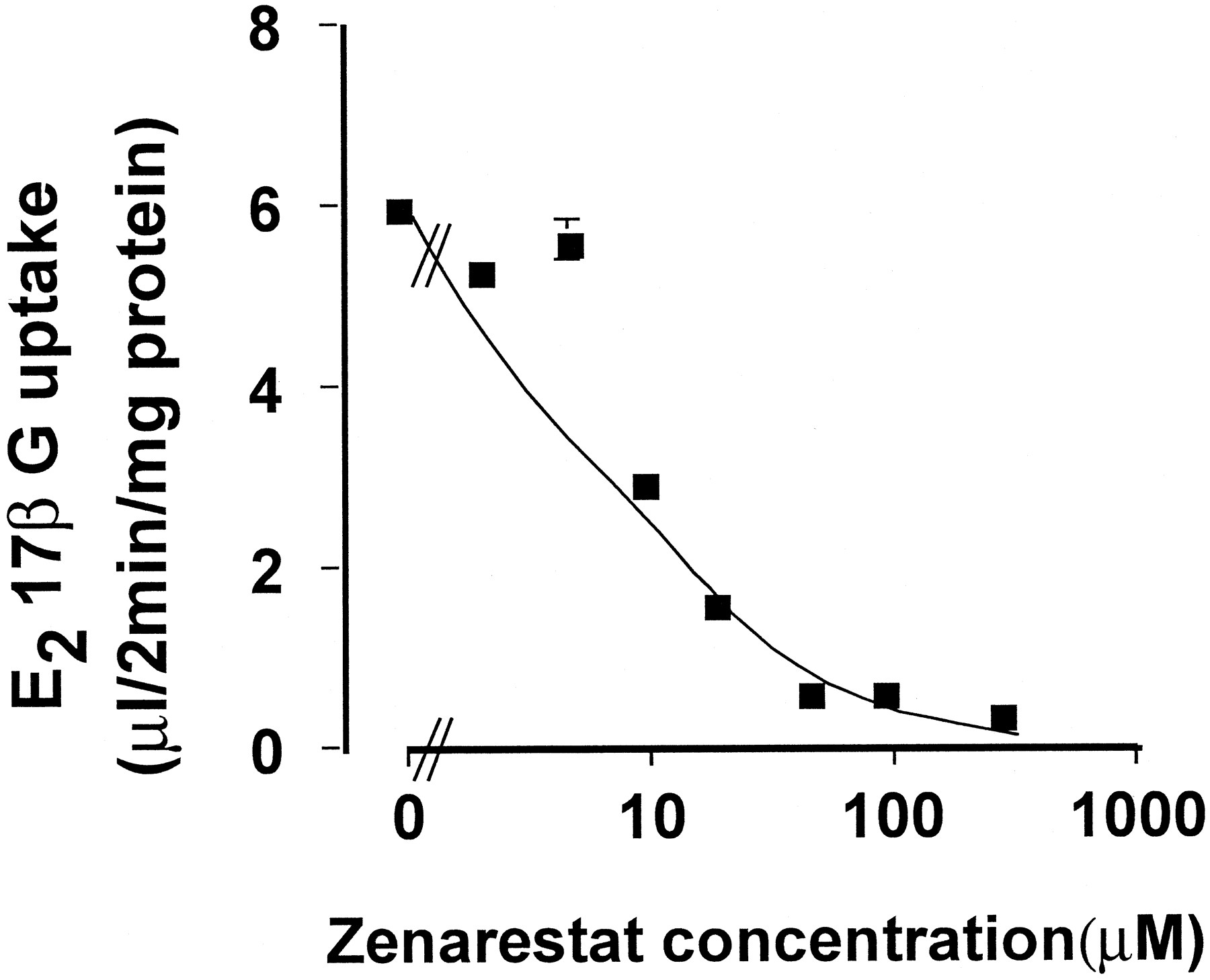
Inhibition of the uptake of [3H]E217βG by zenarestat in Oatp1-transfected LLC-PK1 cells. LLC-PK1 cells stably transfected with rat Oatp1 were incubated at 37oC for 2 min with [3H]E217βG (0.3 μCi/ml). The uptake in vector-transfected cells was subtracted from that in Oatp1-transfected cells and normalized by the substrate concentration in the medium. The solid line indicates the fitted line. Each point and vertical bar represent the mean ± S.E. of three experiments.
Gender Difference in the Expression of Organic Anion Transporters.
The mRNA expression of Oatp1 in the kidney was much higher in male than in female rats, whereas this gender difference was minimal in the liver (Fig. 5A). Such higher Oatp1 expression was also confirmed by Western blotting (Fig. 5C). These findings were compatible with previous findings (Lu et al., 1996; Goto et al., 2002). Bergwerk et al. (1996) have reported that renal Oatp1 migrates as 33- and 37-kDa peptides under reduced conditions, whereas liver Oatp1 migrates as 83-kDa protein. In the present study, the size of renal Oatp1 was also smaller than that of hepatic Oatp1, although multiple bands cannot be clearly seen in the kidney (Fig. 5B). Oudar et al. (1991) reported a gender difference in the relative volume of both the cortex and medulla with regard to the total kidney volume. The difference in the relative volume of the medulla was at most 2-fold greater in female than in male rats and could have a minor effect on the observation of higher Oatp1 expression in male than in female rats. The mRNA of Oatp2 was not detected in either male or female rat kidney (Fig. 5A). The mRNA of Mrp2 in the liver did not show any clear gender difference, whereas that in the kidney could not be detected under these conditions (Fig. 5A). The mRNA of Oat2 in the kidney was much higher in female than in male rats, whereas the mRNA of Oat3 in the liver was higher in male than in female rats (Fig. 5A), and these results were compatible with recent findings (Kobayashi et al., 2001).

Northern blot analysis of Oatp1, Oatp2, Mrp2, Oat2, and Oat3 (A and B) and Western blot analysis of Oatp1 (C) in kidney and liver. Panels A and B, 2 mg of poly (A)+ RNAs from male and female rat tissues were used to detect the mRNA of these transporters (A) and β-actin (B). A typical result is shown, and reproducible results were obtained on three occasions. Panel C, 30 μg of crude membrane from kidney and liver of three male and female rats was separated by SDS-polyacrylamide gel electrophoresis. Data represent the results obtained from three rats.
Discussion
Several reports have shown gender differences in the urinary excretion of certain types of organic anions in rats (Tanaka et al., 1991a; Nakamura et al., 1993; Terashita et al., 1994; Sato et al., 2000), although the mechanism for such gender differences is not yet fully understood. Considering that Oatp1 takes up a variety of types of organic anions into cells (Eckhardt et al., 1999) and that the gene expression of Oatp1 in kidney is higher in male than in female rats (Lu et al., 1996), the urinary excretion of its substrates may also exhibit a gender difference if Oatp1 is mainly involved in the renal transport of the substrates. Actually, we have previously reported that the urinary excretion of E217βG, a typical substrate of Oatp1, is more than 250 times higher in female than in male rats (Goto et al., 2002). Therefore, here, we attempted to examine such a gender difference in urinary excretion for other substrates and inhibitors, including therapeutic agents. We found that the CLurine,p of TCA and DBSP, a substrate and inhibitor of Oatp1, respectively, is higher in female than in male rats (Fig. 1, Table 1). In addition, zenarestat, which exhibits a similar gender difference in its urinary excretion (Fig. 2; Tanaka et al., 1991a), has been shown to have an affinity for Oatp1 in the present study (Fig. 4). Therefore, at least some types of Oatp1 substrates or inhibitors commonly exhibit a gender difference in their urinary excretion in rats.
To examine the reason for such gender differences, further kinetic analysis was performed in vivo to investigate the excretion of zenarestat. Since the uptake of zenarestat by the kidney is not very different between males and females (Fig. 3), there may be only a minimal gender difference in the renal uptake of zenarestat. In addition, a gender difference may be present in zenarestat transport across the apical membrane since the CLurine,k of zenarestat was higher in female than in male rats (Table 2). This CLurine,k was increased by DBSP (Table 2), suggesting that such transport can be inhibited by DBSP. Considering both the GFR (Table 2) and fu of zenarestat (0.0033 and 0.0056 in male and female rats, respectively; Tanaka et al., 1993), the estimated glomerular filtration clearance for this compound (fuGFR) is 1.2 to 2.2 ml/h/kg, which is much higher than the CLurine,p of zenarestat in control male rats (Table 2), suggesting extensive reabsorption of this compound in males.
Considering that substrates or inhibitors of Oatp1 commonly exhibit higher urinary excretion in female rats (Figs. 1 and 2) and that Oatp1 is expressed at a higher level in male rats (Fig. 5; Lu et al., 1996) on the apical membranes of tubular epithelial cells (Bergwerk et al., 1996), such a gender difference can be explained if we hypothesize that Oatp1 is involved in the reabsorption of its substrates. The urinary excretion in castrated male rats was increased to a level comparable with that found in female rats (Tanaka et al., 1991b), and treatment of male and female gonadectomized or hypophysectomized rats with testosterone resulted in urinary excretion of zenarestat that was characteristic of that in male rats. (Tanaka et al., 1991b, 1992). On the other hand, the mRNA for Oatp1 is reduced in the kidney of castrated male rats, and treatment of castrated male and female rats with testosterone increased its expression (Lu et al., 1996). The Oatp1 gene product is also reduced in the kidney of castrated male rats, whereas it is increased in female rats treated with testosterone (Goto et al., 2002). Urinary clearance of both zenarestat (Table 2) and E217βG (Goto et al., 2002) is increased by the Oatp1 inhibitor DBSP. These findings are compatible with the hypothesis that the reabsorption of these compounds, which is possibly mediated by Oatp1 and/or transporters that have a similar substrate specificity to Oatp1, can be inhibited by DBSP. However, in the presence of the highest dose of DBSP (150 μmol/h/kg), the CLurine,k of zenarestat in female rats was still 4 times higher than in males (Table 2), whereas the CLurine,p of E217βG showed only a minimal gender difference at the same DBSP dose (Goto et al., 2002). Since the CLurine,p of zenarestat at 150 μmol/h/kg of DBSP was still lower than the fuGFR as calculated above (Table 2), reabsorption may not be completely inhibited by DBSP and/or other mechanism(s) may be involved in the gender difference in renal excretion. Interestingly, the urinary excretion of zenarestat is reduced by probenecid in female rats, whereas such inhibition is minimal in males (Tanaka et al., 1991a), suggesting that some probenecid-sensitive transporter, involved in the secretion of zenarestat, is expressed in the kidneys of female rats. Since probenecid is an inhibitor of Oatp1, the inhibition of Oatp1 by probenecid cannot be ruled out in the studies by Tanaka et al. (1991a). Nevertheless, considering their results together with the present findings, multiple transporters, including a probenecid-sensitive transporter and a male-specific reabsorption transporter, might be involved in the gender difference seen in zenarestat excretion. Further studies are needed to clarify this hypothesis.
Northern blot analysis revealed that Oatp1 gene expression is much higher in male rats than in females, whereas no clear gender difference can be detected for Oatp2 or Mrp2, although one was observed both in Oat2 in kidney and Oat3 in liver (Fig. 5). Lu et al. (1996) have also reported a similar gender difference in mRNA for Oatp1 where the probe for such Oatp1 detection corresponded to 254 to 1436 bp, which was located in the open reading frame of Oatp1. Considering that the OATP family shows a high degree of sequence homology (85 and 87% homology between Oatp1 and Oatp2, and between Oatp1 and Oatp3, respectively), in this study, we used a probe consisting of the noncoding region of Oatp1, which showed only a minimal cross-reaction with the cRNA of Oatp2 and Oatp3. Cattori et al. (2000) have reported another Oatp member, Oatp4, which exhibits liver-specific expression in male rats. Considering these results, the gene expression of Oatp1, apart from the other three clones in kidney, is higher in male rats than in females, although there is still a need to exclude the possibility that other, perhaps unknown, members of the OATP family may affect the Northern blot analysis.
In addition to Oatp1, the mRNA of Oat2 showed a gender difference in the kidney (Fig. 5) as reported by Kobayashi et al. (2001). Oat2 takes up a variety of types of substrates, including salicylate, prostaglandin E2, indomethacin, and 3′-azido-3′-deoxythymidine (Sekine et al., 1998; Morita et al., 2001), because its substrate specificity is different from that of Oatp1 to some extent. However, the present findings alone are not enough to rule out the possibility that Oat2 may be involved in the gender difference in the urinary excretion of certain types of organic anions. In fact, bromosulfophthalein, a structural analog of DBSP and TCA, has been reported to inhibit Oat2-mediated transport (Sekine et al., 1998;Morita et al., 2001). Therefore, further studies are needed to clarify whether this transporter is involved in the gender difference in the secretion and/or reabsorption of organic anions.
In conclusion, the urinary excretion of several Oatp1 substrates and inhibitors exhibits a gender difference in rats, and a DBSP-sensitive reabsorption process at the apical side is involved to some extent in the gender difference seen in the urinary excretion of zenarestat.
Footnotes
-
DOI: 10.1124/jpet.102.033878
- Abbreviations:
- Oat
- organic anion transporter
- Oatp
- organic anion transporting polypeptide
- Oct
- organic cation transporter
- TCA
- taurocholate
- GFR
- glomerular filtration rate
- DBSP
- dibromosulfophthalein
- HPLC
- high-pressure liquid chromatography
- E217βG
- [3H]estradiol 17β-d-glucuronide
- AUC
- area under the curve
- CLtot
- total clearance
- CLurine,p
- urinary clearance with respect to plasma concentration
- CLurine,k
- urinary clearance with respect to the kidney concentration
- bp
- base pair(s)
- TBST
- Tris-buffered saline/Tween 20
- Mrp
- multidrug resistance-associated protein
- Received February 6, 2002.
- Accepted April 8, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}