


Abstract
Organic anion transporter 1 (OAT1/SLC22A6) is predominantly expressed in the proximal tubules of the kidney. Cumulative studies have shown its critical role in the tubular secretion of a variety of organic anions, including several clinically important drugs. In addition, OAT1 is also involved in the pharmacological effect of diuretics and the nephrotoxicity of antiviral drugs. In contrast to these functional characterizations, the regulatory mechanism of OAT1 expression is poorly understood. It was recently demonstrated that the expression of Oat1 was markedly reduced in the kidneys of hepatocyte nuclear factor 1α (Hnf1α)-null mice. However, in vitro evidence for the involvement of HNF1α and further analyses are required to illustrate the transcriptional regulation of OAT1 genes in more detail. Computational analysis of the potential transcription factor binding sites revealed that the HNF1-motif was conserved in the proximal-promoter region of human and mouse OAT1 genes. The mRNA expression of mouse organic anion transporter 1 was drastically reduced in Hnf1α-null mice compared with that in wild-type mice, which was consistent with a previous report (Maher et al., 2006). Forced expression of HNF1α alone or both HNF1α and HNF1β enhanced the activity of human and mouse OAT1 promoters in the transactivation assays, whereas HNF1β alone was not active. Mutations in the HNF1-motif significantly reduced this transactivation. Direct binding of HNF1α/HNF1α homodimer and HNF1α/HNF1β heterodimer to the HNF1-motif found in the human OAT1 promoter was demonstrated by electrophoretic mobility shift assays. These results provide convincing evidence for the involvement of HNF1α/β in the constitutive expression of human and mouse OAT1 in the kidney.
Secretary transport from blood to urine across the renal proximal tubules is an important pathway in the renal elimination of many compounds, including both endobiotics and xenobiotics. Cumulative studies have shown that organic anion transporter 1 (OAT1/SLC22A6) and organic anion transporter 3 (OAT3/SLC22A8) account for the uptake of anionic compounds from the systemic circulation into the renal proximal-tubular epithelial cells at the basolateral membrane (Enomoto and Endou, 2005; Sekine et al., 2006). So far, many groups, including us, have characterized the transport properties of OAT1 and OAT3 and demonstrated that several drugs, such as β-lactam antibiotics, nonsteroidal anti-inflammatory drugs, antiviral drugs, and loop and thiazide diuretics are substrates of these transporters. Generation of Oat1- and Oat3-null mice confirmed the essential role of these transporters in the renal secretion of drugs (Sweet et al., 2002; Eraly et al., 2006). The impairment in Oat1 function in the kidney affects not only the tubular secretion of drugs but also their pharmacodynamics. For instance, loss of Oat1 in the kidney results in the reduced secretion of furosemide, leading to attenuation of the diuretic effect of this drug. It was also suggested that the accumulation of antiviral drugs, adefovir and cidofovir, in the kidney via OAT1 is associated with their nephrotoxicity (Ho et al., 2000; Cihlar et al., 2001). In contrast to these functional characterizations, information about the mechanism underlying the kidney-specific expression of OAT1 remains limited.
We have recently characterized the transcriptional regulation of OAT3 and urate transporter 1 (URAT1/SLC22A12) and shown that the coordinated action of hepatocyte nuclear factor 1 α/β (HNF1α/β) and DNA methylation determines the kidney-specific expression of these transporters (Kikuchi et al., 2006, 2007). HNF1 consists of two isoforms, HNF1α and HNF1β, and activates the transcription of target genes via direct binding to their promoters after forming homodimers or heterodimers between the two isoforms (Mendel and Crabtree, 1991; Tronche and Yaniv, 1992). These transcription factors were originally identified to be involved in the maintenance of hepatic gene expression, such as albumin, α1-antitrypsin, and α- and β-fibrinogen, as well as some of the organic anion transporters in the liver (Shih et al., 2001). As a result, cumulative reports including the analyses of gene-disrupted animals suggest the importance of HNF1α/β in extrahepatic organs, such as the pancreas and kidney (Pontoglio et al., 1996; Lee et al., 1998; Gresh et al., 2004).
In the kidney, HNF1α exhibits restricted distribution within the proximal tubules, whereas HNF1β is expressed along the entire nephrons (Lazzaro et al., 1992; Pontoglio et al., 1996). HNF1 normally exists as the HNF1α/HNF1β heterodimer or HNF1β/HNF1β homodimer in the proximal tubules, whereas HNF1β/HNF1β homodimer is predominantly expressed in the other segments. As for renal organic anion transporters, mRNA expression of Oat1, as well as Oat2 and Oat3 in the kidney, is reduced in Hnf1α-null mice, suggesting the role of HNF1α in the transcription of organic anion transporters not only in the liver but also in the kidney (Maher et al., 2006). Computational analysis of the human and mouse OAT1 5′-flanking sequences revealed that the HNF1-motif is conserved in the proximal-promoter region of OAT1 genes (Fig. 1). On the other hand, Ogasawara et al. (2007) recently reported that HNF4α, an orphan member of the nuclear receptor superfamily, enhances the promoter activity of human OAT1, whereas the effect of HNF1α or HNF1β was minimal. HNF4α forms homodimers to bind to a DNA sequence corresponding to a direct repeat of AGGTCA-like hexamers separated by one or two nucleotides (DR1 or DR2, respectively) or an inverted repeat of the hexamers separated by eight nucleotides (IR-8) (Sladek et al., 1990; Fraser et al., 1998; Prieur et al., 2005) and regulates the hepatic expression of human organic anion transporter 2 (SLC22A7) and organic cation transporter 1 (SLC22A1) under both physiological and pathological conditions (Popowski et al., 2005; Saborowski et al., 2006). Although HNF4α is found in the proximal tubules in the kidney (Jiang et al., 2003), the physiological significance of HNF4α in the kidney is poorly recognized due to the embryonic death of Hnf4α-null mice (Chen et al., 1994). Furthermore, it has yet to be investigated whether HNF1α/β is actually involved in the transcription of the human OAT1 gene, as is the case with OAT3 and URAT1.
In accordance, the purpose of the present study was to examine whether HNF1α and/or HNF1β is involved in the transcriptional regulation of human and mouse OAT1 genes and to further confirm the importance of HNF1 in the regulation of organic anion transporters in the kidney.
Materials and Methods
Materials. All reagents were purchased from Wako Pure Chemicals (Osaka, Japan) unless stated otherwise.
Preparation of Total RNA and Quantitative Polymerase Chain Reaction. Total RNA was isolated from the kidney of 7 to 14-week-old male (n = 3) and female (n = 4) wild-type or Hnf1α-null mice (Lee et al., 1998), and it was treated with DNase I to eliminate the contaminated genomic DNA. The total RNA was reverse-transcribed using a random-nonamer primer (Takara, Shiga, Japan), and real-time quantitative polymerase chain reaction (PCR) was performed as previously described (Kikuchi et al., 2006) using the primers shown in Table 1 to quantify the mRNA expression of mouse organic anion transporter 1 (mOat1). The mRNA expression of mOat1 was normalized by the mRNA expression of GAPDH and statistically analyzed by the Student's t test.
Oligonucleotides used for the production of promoter constructs, site-directed mutagenesis, EMSA, and quantitative PCR With regard to the oligonucleotides used for EMSA and site-directed mutagenesis, the HNF1-motif in the hOAT1 promoter is underlined. Boldface type indicates the difference in the sequence of the per and mut compared with the wild-type sequence found in the hOAT1 promoter.
Isolation of the 5′-Flanking Region of the hOAT1 and mOat1 Genes. The transcriptional start site of the hOAT1 and mOat1 gene was identified based on the information in the public database, Database of Transcriptional Start Sites (http://dbtss.hgc.jp/), with the reference sequence identification for hOAT1 and mOat1 (NM_004790 and NM_008766, respectively). The position of the potential transcription factor binding sites in human and mouse OAT1 promoter regions was determined using MatInspector (http://www.genomatix.de/) or NUBIScan (http://pages.unibas.ch/wtt/Products/Nubiscan/nubiscan.html) (Podvinec et al., 2002). The 919- and 110-base pair (bp) 5′-flanking regions of the human and mouse OAT1 gene were amplified by PCR using human and mouse genomic DNA as a template, respectively. An artificial KpnI or HindIII restriction site was added to the primer sequences, which are shown in Table 1. The PCR products were digested with KpnI and HindIII after subcloning into pGEM-T Easy vector (Promega, Madison, WI) and ligated into pGL3-Basic vector (Promega) predigested with KpnI and HindIII, yielding the hOAT1_-919/+11_HNF1wt and mOat1_-110/+10_HNF1wt promoter-reporter construct. A series of hOAT1 5′-truncated promoter fragments (-623/+11, -318/+11, and -111/+11) were PCR-amplified using hOAT1_-919/+11_HNF1wt plasmid as a template (the primers are shown in Table 1) and then inserted into the pGL3-Basic vector as described above, yielding the following promoter constructs: hOAT1_-623/+11_HNF1wt, -318/+11_HNF1wt, and -111/+11_HNF1wt. The sequence identity of all the constructs with the respective genomic sequences was verified by DNA sequencing. Plasmid DNA was prepared using the GenElute Plasmid Midiprep kit (Sigma-Aldrich, St. Louis, MO).

Alignment of the proximal promoters of human and mouse OAT1 genes. Nucleotide sequences of human and mouse OAT1 promoter regions were aligned using Genetyx-win version 8 software to illustrate the high homology of the 5′-flanking sequences between species. Nucleotide numberings are relative to the transcriptional start sites indicated by an arrow (+1), and homologous sequences between species are boxed. The putative HNF1-motif and the IR-8 element are shaded, and CpG dinucleotides in each sequence are reverse-colored.
Site-Directed Mutagenesis. All of the mutated promoter fragments (HNF1mut) having a 4-bp-disrupted HNF1-motif were generated with a QuikChange XL Site-Directed Mutagenesis Kit (Stratagene, La Jolla, CA) using internally mutated oligonucleotides with sense sequence (as shown in Table 1) according to the manufacturer's instructions. The introduction of mutations was verified by DNA sequencing. The positions and bases that will replace the original sequences were decided based on the information in the database of transcription factors TRANSFAC (http://www.gene-regulation.com/); highly conserved bases in the consensus HNF1-motif were mutated into bases with the lowest frequency at the corresponding position.
Cell Culture, Transfections, and Luciferase Assays. Cell culture and transfections were performed as described previously. In transactivation assays, 0.5 μg of empty pcDNA3.1+ control vector, 0.5 μg of HNF1α expression vector, 0.25 μg of HNF1α and HNF1β expression vectors, or 0.5 μg of HNF1β expression vector was cotransfected with 0.5 μg of the corresponding promoter construct and 0.05 μg of internal standard pRL-SV40 into HEK293 cells. The promoter activity was measured as relative light units of firefly luciferase per unit of Renilla luciferase. The difference in the promoter activity between wild-type and HNF1-mutated reporter constructs was statistically analyzed by the Student's t test.
In Vitro Translation. In vitro translation was performed using TnT Quick Coupled Transcription/Translation kits (Promega) according to the manufacturer's instructions. One microgram of empty pcDNA3.1+ control vector, 1 μg of HNF1α expression vector, 0.5 μgof HNF1α and HNF1β expression vectors, or 1 μg of HNF1β expression vector was added to the TnT Quick master mix. The mixture was then incubated at 30°C for 75 min and used for additional analyses. The HNF1α and HNF1β proteins were prepared at least twice, and the reproducible binding to the labeled OAT1 probe was confirmed.
Electrophoretic Mobility Shift Assay. Double-stranded oligonucleotide probes were generated by hybridizing single-stranded complementary oligonucleotides with sense sequences (shown in Table 1). The sequence “wt” corresponds to the wild-type HNF1-motif found in the hOAT1 promoter, and “per” corresponds to the perfect consensus sequence for the HNF1-motif, whereas “mut” denotes the wild-type sequence mutated in the motif. Electrophoretic mobility shift assay (EMSA) was performed as previously described with Dig Gel Shift Kit, 2nd Generation (Roche Diagnostics, Indianapolis, IN) (Kikuchi et al., 2006). In brief, 2.5 μl of in vitro-translated HNF1α and/or HNF1β was incubated on ice for 30 min in the binding solution containing 0.92 pmol of digoxigenin-labeled probe. A 100-fold excess of unlabeled oligonucleotides and 1 μg of antibody against HNF1α or HNF1β (Santa Cruz Biotechnology, Inc., Santa Cruz, CA) were added to the reaction for competition and supershift assays, respectively. The experiments were independently performed three times, and the representative images are shown.
Results
Computational Analysis of the Potential Transcription Factor Binding Sites in Human and Mouse OAT1 Promoters. The promoter sequences of the human and mouse OAT1 genes were obtained from the National Center for Biotechnology Information genome database and aligned using Genetyx-win version 8 software (Fig. 1). The promoter region of OAT1 was highly conserved between human and mouse, especially up to approximately 100 bp upstream of the transcriptional start site. Computational analysis using MatInspector revealed that the HNF1-motif is conserved at -56 to -44 relative to the transcriptional start site in both promoters, indicating the functional importance of this motif. Analysis using NUBIScan demonstrated that the IR-8 element was located at -123 to -104 in the hOAT1 promoter, but not in the mOat1 promoter, suggesting the possible existence of a species difference in the regulation of OAT1 expression.
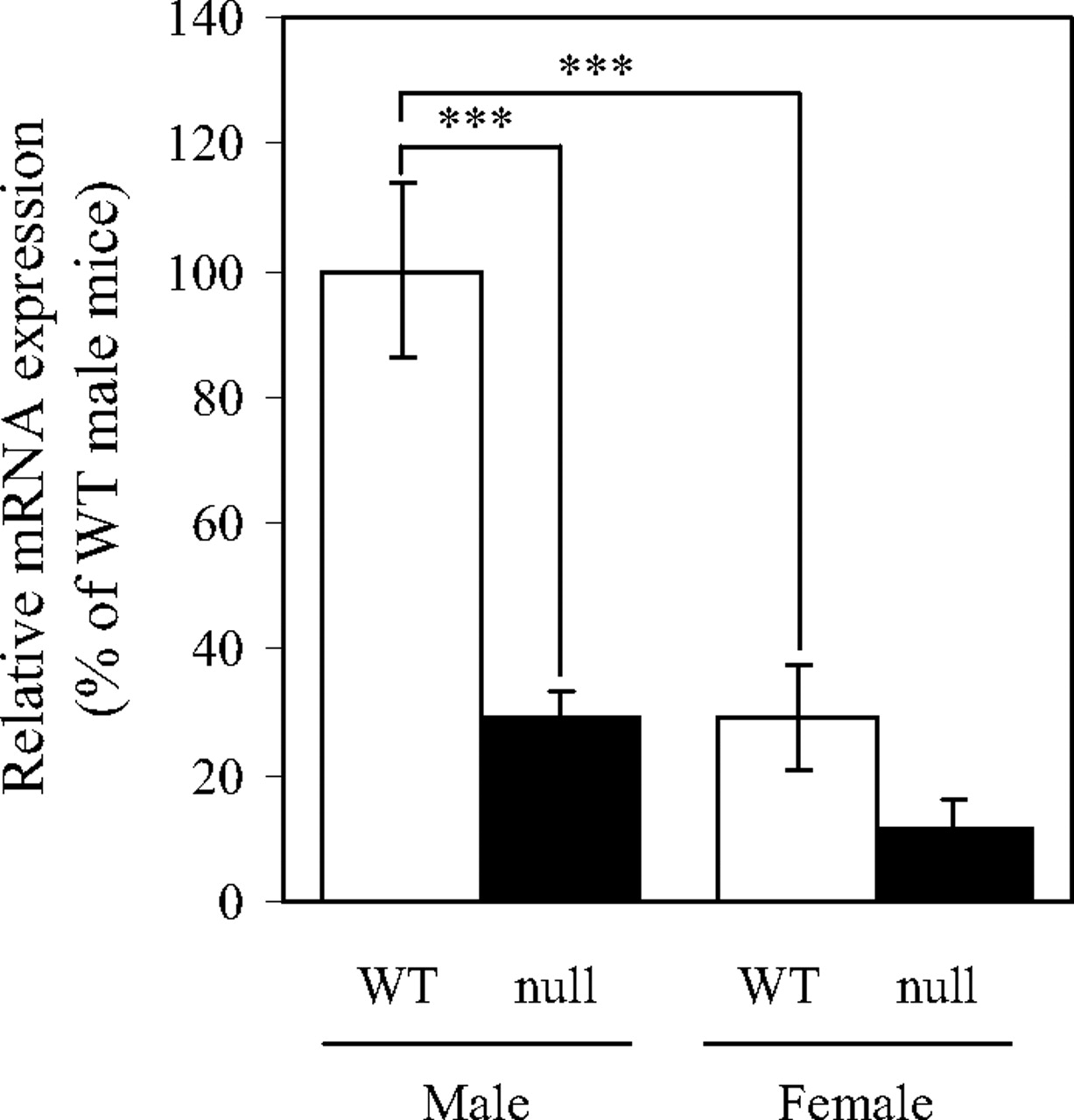
Impaired Expression of mOat1 in the Kidney of Hnf1α-Null Mice. The mRNA expression of mOat1 in male and female Hnf1α-null mice was measured by real-time quantitative PCR (Fig. 2). The expression of mOat1 mRNA was approximately 3-fold higher in male kidney than in female in wild-type controls, which is consistent with the previous findings (Buist and Klaassen, 2004). Inactivation of HNF1α led to a significant reduction in the expression of mOat1 in male mice. In females, the expression of mOat1 mRNA was reduced by two-thirds in Hnf1α-null mice compared with wild-type mice. These results clearly document the importance of HNF1α in the transcriptional regulation of the mOat1 gene in the kidney.
Transactivation of the Human and Mouse OAT1 Promoter by HNF1α/β. We have previously demonstrated that endogenous expression of HNF1α and HNF1β is negligible in HEK293 cells (Kikuchi et al., 2006). Thus, this cell line was used in additional studies to investigate the effect of exogenously expressed HNF1α and/or HNF1β on OAT1 gene promoter activity. The protein expression of exogenously transfected HNF1α or HNF1β in HEK293 cells was confirmed by Western blot analysis after preparation of nuclear extracts from those cells (data not shown). The promoter activity of mOat1 was stimulated by cotransfection of HNF1α alone or both HNF1α and HNF1β, whereas HNF1β alone did not show any enhancement (Fig. 3A). These results provide mechanistic evidence for the involvement of HNF1α in the transcription of mOat1 in the kidney.
To investigate whether HNF1α and/or HNF1β regulate hOAT1-promoter activity, a series of 5′-truncated-promoter constructs of hOAT1, with or without the mutation in the HNF1-motif, was cotransfected with HNF1α and/or HNF1β into HEK293 cells, and the luciferase activity was measured (Fig. 3B). All of the wild-type promoter constructs showed a marked increase in luciferase activity by forced-expression of HNF1α alone or both HNF1α and HNF1β compared with the pcDNA3.1+-transfected control. The effect of HNF1β on the promoter activity of hOAT1 was not marked, regardless of the length of the promoter construct. No additional enhancement was observed by extending the hOAT1 promoter from -111 to -919, suggesting that HNF1α/β binds to the hOAT1-promoter region within 111 bp upstream of the transcriptional start site, where the HNF1-motif was found. The functional relevance of this HNF1-motif was investigated by introducing mutations into the motif. The increase in the luciferase activity caused by the forced expression of HNF1α alone or both HNF1α and HNF1β was attenuated by approximately 50 to 70% in the HNF1-motif-mutated reporter compared with the wild-type reporter. These results strongly suggest that the HNF1-motif found in the hOAT1-proximal promoter is essential for transactivation of the promoter activity by HNF1α alone or both HNF1α and HNF1β.

Relative mRNA expression of mOat1 in wild-type and Hnf1α-null mice. mRNA expression of mOat1 in the kidneys of male or female wild-type (WT, □) and Hnf1α-null mice (null, ▪) was quantified as described under Materials and Methods. The data were normalized by the mRNA expression of GAPDH. The relative mRNA expression was given as a ratio with respect to the mRNA expression of mOat1 in male wild-type mice that was taken as 100%. Results are presented as the mean ± S.E. of three (male) or four (female) mice. ***, P < 0.001, significantly different between the indicated data points.
Direct Binding of HNF1α/β to the hOAT1 Promoter. To confirm the direct binding of HNF1α/β to the hOAT1 promoter, EMSA was performed with the oligonucleotide probe corresponding to the HNF1-motif found in the hOAT1 promoter and in vitro-translated HNF1α and/or HNF1β (Fig. 4A). The expression of in vitro-translated HNF1α or HNF1β was confirmed by Western blot analysis with specific antibodies against HNF1α or HNF1β, respectively (data not shown). When the probe was incubated with HNF1α alone or both HNF1α and HNF1β, one- (Fig. 4A, band a, lane 3) or two-shifted bands (Fig. 4A, bands a and b, lane 7) were observed, respectively, whereas there was no shifted band when the probe was incubated with HNF1β alone (lane 11). In all samples using the in vitro-translated products, a broad signal was detected below the bands (Fig. 4A, bands a and b). It is likely that this signal represents nonspecific binding to the labeled probe because the signal was also detected when in vitro-translated empty pcDNA3.1+ was incubated with the probe (Fig. 4A, lane 2). Formation of both bands a and b (Fig. 4A) was completely abolished by adding a 100-fold excess of the unlabeled competitor corresponding to the consensus sequence for the HNF1-motif (Fig. 4A, lanes 4 and 8) and partly inhibited by the competitor corresponding to the hOAT1 wild-type promoter sequence (Fig. 4A, lanes 5 and 9). On the other hand, these bands were not affected by the addition of the mutated competitor (Fig. 4A, lanes 6 and 10). These results suggest that bands a and b (Fig. 4A) can be ascribed to the binding of HNF1α or HNF1β to the HNF1-motif in the hOAT1 promoter.
To demonstrate the specific binding of HNF1 to the hOAT1 promoter, supershift analysis was performed using antibodies against HNF1α or HNF1β (Fig. 4B). The addition of an anti-HNF1α antibody resulted in the supershift of both bands a and b (Fig. 4B, lanes 2 and 5), whereas the addition of anti-HNF1β antibody abolished band b but not band a (Fig. 4B, lanes 3 and 6). The supershifted bands were barely detectable when both HNF1α and HNF1β were incubated with the labeled probe (Fig. 4B, lanes 5 and 6), probably due to the low intensity of the shifted bands. The mobility of bands a and b (Fig. 4B) in the present study nearly coincides with that of the shifted bands that reflect the interaction of HNF1α/HNF1α homodimer and HNF1α/HNF1β heterodimer with the HNF1-motif in hOAT3 and human URAT1 promoters, respectively (Kikuchi et al., 2006, 2007). These results strongly suggest that bands a and b (Fig. 4B) reflect the binding of HNF1α/HNF1α homodimer and HNF1α/HNF1β heterodimer, respectively.

Transactivation of human and mouse OAT1 promoters by HNF1α and/or HNF1β. A, HEK293 cells were transfected with mOat1 wild-type promoter (mOat1_-110/+10_HNF1wt), or a promoterless pGL3-Basic plasmid, together with empty pcDNA3.1+ vector (white bars), HNF1α expression vector (black bars), HNF1α, and HNF1β expression vectors (gray bars), or HNF1β expression vector (hatched bars). The luciferase activity was measured as described under Materials and Methods and was shown as the factor of induction over background activity measured in cells transfected with pGL3-Basic together with pcDNA3.1+. B, the activity of a series of hOAT1 wild-type or HNF1-mutated promoters in the presence or absence of exogenously expressed HNF1α and/or HNF1β was measured as described in A. Results are presented as the mean ± S.E. of triplicate samples. *, P < 0.05, **, P < 0.01, and ***, P < 0.001, significantly different between wild-type and the corresponding HNF1-mutated promoters.
Discussion
Cumulative evidence suggests that HNF1α and/or HNF1β play a critical role in the expression of drug transporters in the liver and kidney. Several groups, including us, have shown that the HNF1-motif located within approximately 100 bp of the 5′-flanking region is involved in the transcriptional regulation of these transporters, i.e., -65 to -53 in the OAT3 promoters, -70 to -58 in the URAT1 promoters, -51 to -39 in the organic anion-transporting polypeptide 1B1 promoter, and -60 to -48 in the organic anion-transporting polypeptide 1B3 promoter (Jung et al., 2001; Kikuchi et al., 2006, 2007). Consistent with this finding, a database search for the potential transcription factor binding sites revealed that an HNF1-motif is conserved from -56 to -44 relative to the transcriptional start sites of the human and mouse OAT1 promoters (Fig. 1). These observations as well as the reduced expression of mOat1 in the kidney of Hnf1α-null male mice (Maher et al., 2006) prompted us to investigate the role of HNF1α/β in the transcriptional regulation of human and mouse OAT1 genes.
The involvement of HNF1α and/or HNF1β in the transcriptional regulation of human and mouse OAT1 genes was revealed by in vivo (Fig. 2) and in vitro experiments (Figs. 3 and 4). It is generally accepted that the transactivation potency of HNF1β is lower than that of HNF1α (Rey-Campos et al., 1991). Indeed, exogenous expression of HNF1β alone enhanced the promoter activity of OAT3 and URAT1 by at most 50% compared with that of HNF1α alone (Kikuchi et al., 2006, 2007). In the present study, HNF1β alone hardly stimulated the promoter activity of the human and mouse OAT1 genes, and no direct binding of HNF1β/HNF1β homodimer to the hOAT1 promoter could be detected. HNF1β exhibited less activity against the OAT1 promoters than OAT3 and URAT1 promoters. Therefore, it is likely that the contribution of HNF1β/HNF1β homodimer to the expression of OAT1 is much smaller than that of OAT3 and URAT1.
Ogasawara et al. (2007) demonstrated that HNF4α markedly transactivated the hOAT1 promoter through direct binding to the IR-8 element, whereas neither HNF1α nor HNF1β affected the promoter activity. On the other hand, we have provided clear evidence for an essential role of HNF1α in human and mouse OAT1 expression. We also confirmed that HNF4α transactivates the hOAT1 promoter. Cotransfection of HNF4α into HEK293 cells enhanced the promoter activity of the hOAT1_-919/+11_HNF1wt construct 3-fold compared with the pcDNA3.1+-transfected control (data not shown). Therefore, the experimental conditions may not be appropriate for examining the effect of HNF1α/β in the previous report. The two studies used different cell lines as host. We used HEK293 cells that lack the endogenous expression of HNF1α/β, whereas Ogasawara et al. (2007) used OK cells in which the endogenous expression of HNF1α/β has not been investigated. The following difference in the reporter construct used could be another reason: hOAT1_-2747/+88 (Ogasawara et al., 2007) versus hOAT1_-919, -623, -318, or -111/+11_HNF1wt (present study).

Direct binding of HNF1α and HNF1β to the hOAT1 promoter. A, competition assays are shown. A digoxigenin-labeled probe corresponding to the hOAT1 wild-type promoter containing the HNF1-motif was incubated with in vitro-translated HNF1α and/or HNF1β, or pcDNA3.1+ as a negative control, in the presence or absence of a 100-fold excess of unlabeled competitor (per, wt, or mut) as indicated. B, supershift analysis is shown. The probe was incubated with HNF1α alone or both HNF1α and HNF1β in the presence or absence of a specific antibody against HNF1α (α) or HNF1β (β) as indicated. The DNA-protein complex was detected as described under Materials and Methods. The representative images of three independent experiments are shown.
In the kidney, the expression of both HNF1α and HNF4α is restricted to the proximal tubules (Lazzaro et al., 1992; Pontoglio et al., 1996; Jiang et al., 2003), consistent with the regional distribution of hOAT1 (Hosoyamada et al., 1999). Thus, the proximal tubule-specific expression of hOAT1 in the kidney is, at least partly, explained by the concerted effect of HNF1α/HNF1β heterodimer and HNF4α/HNF4α homodimer. However, the tissue distribution of HNF1αβ and HNF4α is much wider than that of OAT1 (Sladek et al., 1990; Blumenfeld et al., 1991; Rey-Campos et al., 1991; Miquerol et al., 1994; Drewes et al., 1996). Although OAT1 is exclusively expressed in the kidney, HNF1α, HNF1β, and HNF4α are expressed in extrarenal tissues, such as the liver, intestine, and pancreas. We have recently shown that the kidney-specific expression of OAT3 and URAT1 genes is regulated by the synergistic effect of transcriptional activation by HNF1α/β and repression by DNA methylation in the promoter region. However, unlike the minimal promoter regions of hOAT3 and human/mouse URAT1, there are few CpG dinucleotides in the human and mouse OAT1 promoter regions up to -400 bp (Fig. 1). It is noteworthy that OAT1 and OAT3 genes occur as a tightly linked pair on the same chromosome in the human and mouse genome with intergenic distances of 8.3 kilobase pairs in humans and 7.5 kilobase pairs in mice (Eraly et al., 2003). Both genes are transcribed to the same direction with OAT3 upstream of OAT1. The clustering of OAT1 and OAT3 in the genome raises the possibility that the methylation status of the OAT3 promoter region may affect the widespread chromatin configuration including the OAT1 gene, enabling DNA methylation-dependent gene silencing of OAT1 in extrarenal tissues. Future studies are required to show the in vivo relevance of the epigenetic regulation for OAT1 expression.
In conclusion, the present study clearly demonstrates that HNF1α/HNF1β heterodimer plays a key role in the constitutive expression of human and mouse OAT1 genes. The transcriptional activation by HNF1 is a common feature among renal organic anion transporters, further emphasizing the role of this transcription factor in the regulation of endobiotics/xenobiotics transport in the kidney.
Footnotes
-
T.S. and R.K. contributed equally to this work.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.107.128249.
-
ABBREVIATIONS: OAT, organic anion transporter; URAT, urate transporter; HNF, hepatocyte nuclear factor; hOAT, human OAT; PCR, polymerase chain reaction; mOat, mouse organic anion transporter; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; bp, base pair; HEK, human embryonic kidney; wt, wild type; mut, mutated wild-type sequence; EMSA, electrophoretic mobility shift assay.
- Received July 6, 2007.
- Accepted November 19, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}