


Abstract
Repeated intracerebroventricular injections of antisense oligodeoxynucleotides (ODNs) were used to selectively restrict the expression of cloned mu and delta opioid receptors (OR) in the mouse brain. Reduction of mu anddelta OR-like immunoreactivity was observed in brain structures of experimental mice. A random-sequence ODN used as a control showed no effect. ODNs to OR decreased radiolabeling of neural structures after intracerebroventricular injection of125I-immunoglobulins G directed to muor delta OR. The potencies of opioids binding themu OR, [d-Ala2,N-MePhe4,Gly-ol5]enkephalin and morphine were significantly attenuated in mice injected with ODNs to this receptor, an effect not seen for the deltaOR-binding agonists, [d-Pen2,5]enkephalin and [d-Ala2]deltorphin II. In morphine-dependent mice, ODNs to mu OR reduced the incidence of naloxone-precipitated withdrawal jumping, body weight loss and diarrhea. The ODN directed to nucleotides 7–26 of thedelta OR mRNA selectively impaired antinociception induced by [d-Ala2]deltorphin II (delta-2), but not that of [d-Pen2,5]enkephalin (delta-1) or morphine. It also diminished the incidence of withdrawal signs precipitated by naloxone in morphine-dependent mice. Thus, the clonedmu OR mediates morphine-evoked antinociception as well as physical dependence. The involvement of delta-2 OR in the development and/or expression of morphine dependence is suggested.
Opioids exert their effects after binding to specific receptors located throughout the central and peripheral nervous systems. Pharmacological experiments have suggested diversity among opioid receptors. The main types are designatedmu, delta and kappa (Martin et al., 1976; Lord et al., 1977). Mu opioid receptors are generally assumed to mediate antinociceptive effects of opioids at the supraspinal level (Höllt et al., 1985). Further, the interaction of morphine at the mu receptor is known to be the initial step in the development of tolerance to and physical dependence on this alkaloid (DeLander et al., 1984;Aceto et al., 1986). The existence of two populations of themu opioid receptor, named mu-1 andmu-2, has been proposed (Pasternak and Snyder, 1975; Wolozin and Pasternak, 1981). The mu-1 subtype appears to be linked to the modulation of opioid-induced supraspinal analgesia, whereasmu-2 is mostly associated with respiratory depression, inhibition of gastrointestinal transit and physical dependence (Linget al., 1984). The existence of mu, delta andkappa opioid receptors was confirmed by molecular cloning (see for review Uhl et al., 1994). Notwithstanding the ample pharmacological evidence for subtypes of these opioid receptors, up till now only three species have been cloned, the mu-1, delta-2 and kappa-1 subtypes.
The role of delta OR in supraspinal antinociception has been well documented (Porreca et al., 1987;Sánchez-Blázquez and Garzón, 1989). The dissimilar antinociceptive profiles exhibited by the delta-selective agonists, DPDPE and [d-Ala2]deltorphin II, led to the proposal of the existence of subtypes delta-1 anddelta-2 (Mattia et al., 1991; Sofuoglu et al., 1991). Interestingly, delta receptors have been implicated in the neural adaptive response to chronic morphine administration (Abdelhamid et al., 1991). The nonequilibrium and long-lasting antagonist at delta-2 OR, naltrindole 5′-isothiocyanate, has been shown to substantially reduce the development of physical dependence on morphine (Miyamoto et al., 1993).
The use of antisense ODNs in the inhibition of mRNA translation into functional proteins is a validated approach in in vitrosystems (Caceres and Kosik, 1990; Holopainen and Wojcik, 1993). This strategy is also used to impair receptor-mediated functions in in vivo studies (Wahlestedt et al., 1993; Zhang and Creese, 1993). ODNs to opioid receptors were reported to selectively block the antinociceptive effects of mu OR agonists in rats (Rossi et al., 1994; Chen et al., 1995), the spinal antinociception induced by delta-preferential compounds (Standifer et al., 1994; Tseng et al., 1994), the supraspinal antinociception induced by thedelta-2 agonist [d-Ala2]deltorphin II (Lai et al., 1994; Bilsky et al., 1996) and the antinociceptive actions exerted by the kappa opioid agonist U-50,488H in mice (Chien et al., 1994). In the present study, ODNs to mRNAs coding for mu anddelta OR were used to further investigate the role of these receptors in the development of tolerance to and physical dependence on chronic morphine administration. Thus, increasing amounts of the ODNs were i.c.v. injected to mice for 5 consecutive days. The efficacy and selectivity achieved by such treatment was established through changes in opioid-evoked antinociception and immunodetection studies. The results indicate that the ODNs produced changes in the expression ofmu and delta OR. Participation of the clonedmu OR in opioid-mediated analgesia as well as in morphine dependence was indicated. Additional evidence is provided for the involvement of the delta-2 OR in the development and/or expression of morphine dependence.
Materials and Methods
Animals and evaluation of analgesia.
Albino male mice CD-1 (Charles River, Barcelona, Spain) weighing 22 to 25 g were used throughout. Animals were kept at 22°C and a 12 h light/dark cycle (8 a.m./8 p.m.) was established. Food and water were provided ad libitum. Mice were housed and used strictly in accordance with the guidelines of the European Community about Care and Use of Laboratory Animals. To reduce the possibility of interference from spinal events, all substances were i.c.v. administered in a final volume of 4 μl to ether-anesthetized mice. The warm water (52°C) tail-flick test was used to measure the antinociceptive effect. Latencies in seconds were determined before treatment (basal latency) and also after the administration of the substance under study. Base-line latencies ranged from 1.3 to 2.2 s and were not affected by ODN administration. A cut-off time of 10 s was allotted to minimize the risk of tissue damage. Antinociception was expressed as a percentage of the MPE according to the following formula: %MPE = 100 × (test latency − base-line latency)/(cut-off time − base-line latency). A single i.c.v. dose of the opioids was given, and antinociception was assessed 30 min after morphine, 15 min after DAMGO or DPDPE and 10 min after [d-Ala2]deltorphin II. All compounds were dissolved in distilled water except for [d-Ala2]deltorphin II which was dissolved in 1% dimethyl sulfoxide. Agonist solutions were made up immediately before use. Statistical analysis of the results was accomplished by the Fisher’s Exact Probability test or analysis of variance (ANOVA) followed by the Student-Newman-Keuls test. The level of significance was set at P < .05.
Induction of physical dependence on chronic morphine treatment.
Animals were subcutaneously implanted with 10 ml/kg b.wt. of a suspension containing: 50% saline (0.9% NaCl in distilled water), 42.5% mineral oil (Sigma 400–5), 7.5% Arlacel (Sigma A-8009) and 0.1 g/ml morphine base. Development of tolerance was monitored by measuring the analgesic response to the chronic opioid by the tail-flick test. After 72 h of sustained morphine release, dependence was evaluated through precipitating withdrawal syndrome by s.c. administration of 1 mg/kg naloxone. Groups of three mice were placed in acryl-glass boxes (28 × 28 × 40 cm) and the number of jumps recorded over the next 10 min (Sánchez-Blázquez and Garzón, 1994).
Synthesis of ODNs.
End-capped phosphorothioate ODNs were synthesized on a CODER 300 DNA synthesizer by phosphoramidite chemistry (Matteucci and Caruthers, 1981). The introduction of phosphorothioate linkages was achieved by tetraethylthiuram disulfide sulfurization (Vu and Hirschbein, 1991). Crude ODNs were purified by reverse-phase chromatography with COP cartridges (Cruachem, Glasgow, U.K.). The eluted ODNs in 50% acetonitrile-water were then lyophilized (Rouan RC 1009/RCT 90, France). Sequences were as follows: ODN-μ16–32 5′-C*T*GATGTTCCCTGGG*C*C-3′ a 17-base oligo that corresponds to nucleotides 16 to 32 of the identical strand of the mouse mu OR (Min et al., 1994), this sequence bears no homology to any other cloned receptor (GeneBank Database); ODN-μun 5′-C*G*CCCCAGCCTCTTCCT*C*T-3′ (Rossi et al., 1994), which corresponds to a specific 5′-untranslated region of rat MOR-1 clone (Chen et al., 1993; Wang et al., 1993; Thompson et al., 1993) and mouse MOR cDNA (Min et al., 1994). ODN-δ7–265′-G*C*ACGGGCAGAGGGCACC*A*G-3′ corresponding to nucleotides 7 to 26 of the murine delta OR gene sequence (Evans et al., 1992; Kieffer et al., 1992) and identical with that used by Lai et al. (1994). ODN-δ29–465′-A*G*AGGGCACCAGCTCC*A*T-3′ (Standiffer et al., 1994) that corresponds to nucleotides 29 to 46 of the murine deltaOR gene sequence. A random oligo (ODN-RD) with the sequence 5′-C*C*CTTATTTACTACTTTC*G*C-3′ served as a control (Gillardonet al., 1994; Sánchez-Blázquez et al., 1995).
Administration of ODNs.
ODN solutions were made up in the appropriate volume of sterile water immediately before use. To assess the specificity of treatment with ODNs three separate control groups of mice were included: noninjected animals (naive), those which received the vehicle i.c.v. (saline), and animals injected with a random sequence ODN i.c.v. (ODN-RD). Each ODN treatment was performed i.c.v. on a distinct group of 16 to 24 mice by the following schedule: on days 1 and 2 with 1 nmol, days 3 and 4 with 2 nmol, on day 5 with 3 nmol. On day 6 the opioid agonists were i.c.v. injected and their antinociceptive activity evaluated by the warm water tail-flick test. On day 6, parallel groups of animals received 3 nmol of the corresponding ODN and, also by s.c. route, an oily suspension containing morphine. The administration of the ODNs (3 nmol) was then continued for 2 consecutive days. Injections were made into the right lateral ventricle of the mouse as described previously (Sánchez-Blázquez et al., 1995; Bilsky et al., 1996). Animals were lightly anesthetized with ether and injections were performed with a 10-μl Hamilton syringe at a depth of 3 mm at a point 2 mm lateral and 2 mm caudal from the bregma. The 4 μl were infused at a rate of 1 μl every 5 s. After this the needle was maintained for an additional period of 10 s. Successive injections were then passed through the same orifice. To minimize the possibility that repetitive i.c.v. injections would originate neurotoxicity, an interval of 24 h was allotted between administrations of the ODNs (Chiasson et al., 1994).
For immunodetection assays, ODNs to mRNA coding for mu anddelta OR were chronically infused into the lateral ventricles of rats. Under anesthesia a 25-gauge stainless steel cannula was implanted stereotaxically into albino male Wistar rats (240–270 g). A vinyl tubing connected the cannula to an osmotic minipump (Alzet 2 ML4, Alzet Corp., Palo Alto, CA), which was placed under the skin in the lumbar region. The ODNs were delivered at 2.5 μl/h (0.1 nmol/h) for 21 days. The cannula was permanently fixed to the skull by dental acrylic (Yang et al., 1993). The following oligos were used: ODN-μun, 5′-C*G*CCCCAGCCTCTTCCT*C*T-3′ (Rossi et al., 1994), which correspond to a specific 5-untranslated region of MOR-1 clone (Chen et al., 1993; Wang et al., 1993; Thompson et al., 1993), and 5′-A*C*TGCAGCTCCGCA*G*G-3′ corresponding to nucleotides 22 to 37 of the rat delta OR gene (Abood et al., 1994).
Antibodies.
The antibodies used in the study were: antiserum MAS/2 (Garzón et al., 1995) generated in rabbits against the amino-terminal portion (MDSSTGPGNTSDCSDP) of the clonedmu OR, antiserum MU/2EL (Garzón and Sánchez-Blázquez, 1995) raised against the peptide sequence 208–216 (TKYRQGSID) of this receptor and antiserum Δ/1 (Garzónet al., 1994) generated against the N-terminal peptide sequence (1–16: MELVPSARAELQSSPL) of the murine deltareceptor. Anti-OR IgGs were purified as described previously (Garzón et al., 1995).
Iodination of IgGs.
IgGs to mu anddelta opioid receptors were purified by affinity chromatography to the antigenic peptide and subsequent iodination according to Greenwood et al. (1963), (with chloramine-T and Na125I) with minor modifications (Sánchez-Blázquez et al., 1996). The reaction was started by mixing 10 μl of a freshly made 0.1 mg/ml solution of chloramine-T in 50 mM sodium phosphate buffer (pH 7.4), with 65 μl of a solution containing 80 μg purified IgGs and 500 μCi Na125I (NEZ 033A; specific activity, 17 Ci/mg) in 70 mM sodium phosphate buffer (pH 7.4). The reaction was stopped after 60 s with 50 μl of chloramine-T stop buffer [2.4 mg/ml sodium metabisulfite, 10 mg/ml tyrosine (saturated), 10% glycerol, 0.1% xylene cylanol in 10 mM sodium phosphate (pH 7.4), 0.9% NaCl ]. Labeled IgGs were separated from free iodine on a Sephadex G-25 column (PD-10 Pharmacia) first equilibrated with 30 ml of 10 mM sodium phosphate (pH 7.4), 0.9% NaCl, 1% bovine serum albumin and then with 100 ml of 10 mM sodium phosphate (pH 7.4), 0.9% NaCl. The reaction material was eluted with 6 ml of 10 mM sodium phosphate (pH 7.4), 0.9% NaCl and 0.5 ml fractions were collected. The IgGs were obtained in two fractions.
Electrophoresis and immunoblotting.
Membranes from mouse and rat striatum were SDS-solubilized and about 80 μg protein/lane were subsequently resolved by SDS/PAGE in 8 cm × 11 cm × 0.15 cm gel slabs (gradient, 7–18%T; 2.6%C). The resolved proteins were transferred to 0.2-μ polyvinylidene difluoride membranes (Bio-Rad, Richmond, CA). Free sites were blocked with nonfat dry milk (Blocker, Bio-Rad) in Tris-buffered saline (TBS) for 1 h at 37°C. Antimu and delta OR antibodies at 1:1000 dilution in TBS-0.05% Tween 20 (TTBS) were incubated at 6°C for 24 h. Secondary antiserum [goat antirabbit IgG (H+L) horseradish peroxidase conjugate (Bio-Rad, no. 170-6515)] in TTBS was added at 1:3000 dilution and left for 3 h. The substrate solution contained 1 mg/ml 3,3′-diaminobenzidine, 0.02% hydrogen peroxide, 0.04% nickel chloride in 0.1 M Tris buffer (pH 7.2). Immunoblots were analyzed by densitometry by an instrument with reflectance capabilities (Bio-Rad, GS-700 Imaging Densitometer).
Autoradiographic experiments.
Mice that had received either saline, ODN-RD or ODNs to opioid receptors were i.c.v. injected with 4 μl of the 125I-IgGs (about 4,000,000 cpm/mouse). The radiolabeled IgGs were administered bilaterally into the cerebral ventricles. After 24 h brains were removed and frozen on dry ice. Coronal cryostat sections (20 μm) were cut at various levels of the neuraxis, mounted onto gelatin-subbed slides and dried. Brain sections were exposed to a tritium-sensitive film (Hyperfilm3H, Amersham, Arlington Heights, IL) for 20 days at −80°C. Kodak LX-24 developer (3 min) and Kodak AL-4 fixer (5 min) were used to develop the films.
Drugs.
DAMGO, DPDPE and [d-Ala2]deltorphin II were purchased from Peninsula Laboratories (San Carlos, CA), morphine sulfate from Merck (Darmstadt, Germany) and naloxone hydrochloride from Sigma-Aldrich Quı́mica (Madrid, Spain).
Results
Effect of in vivo i.c.v. administration of ODNs tomu and delta OR on supraspinal analgesia induced by various opioid agonists.
In mice receiving the ODNs to the mu OR, the preferential agonists of this receptor, DAMGO and morphine, showed diminished antinociceptive potency. This effect was not observed for agonists binding delta OR (fig.1). Chronic injection of the ODN directed to nucleotides 7 to 26 of the mRNA coding for the cloned delta OR significantly impaired antinociception promoted by the selective agonist of the delta-2 OR [d-Ala2]deltorphin II. Interestingly, the activity of DPDPE, agonist at delta-1 OR, was fully expressed. The ODN29–46 to delta OR impaired the analgesia promoted by these two delta OR-binding agonists. The analgesic potency of the mu OR-preferential agonists was not altered by these ODNs to delta OR (fig. 1). Base-line latencies of naive mouse and mice treated with saline, ODN-RD or the ODNs to opioid receptors were not significantly different. When compared with nonpreinjected animals, the similar potency of the analgesic substances in mice injected with saline or a random oligo showed that the responsiveness of the mouse to the antinociceptive test was not altered by repeated i.c.v. administration.
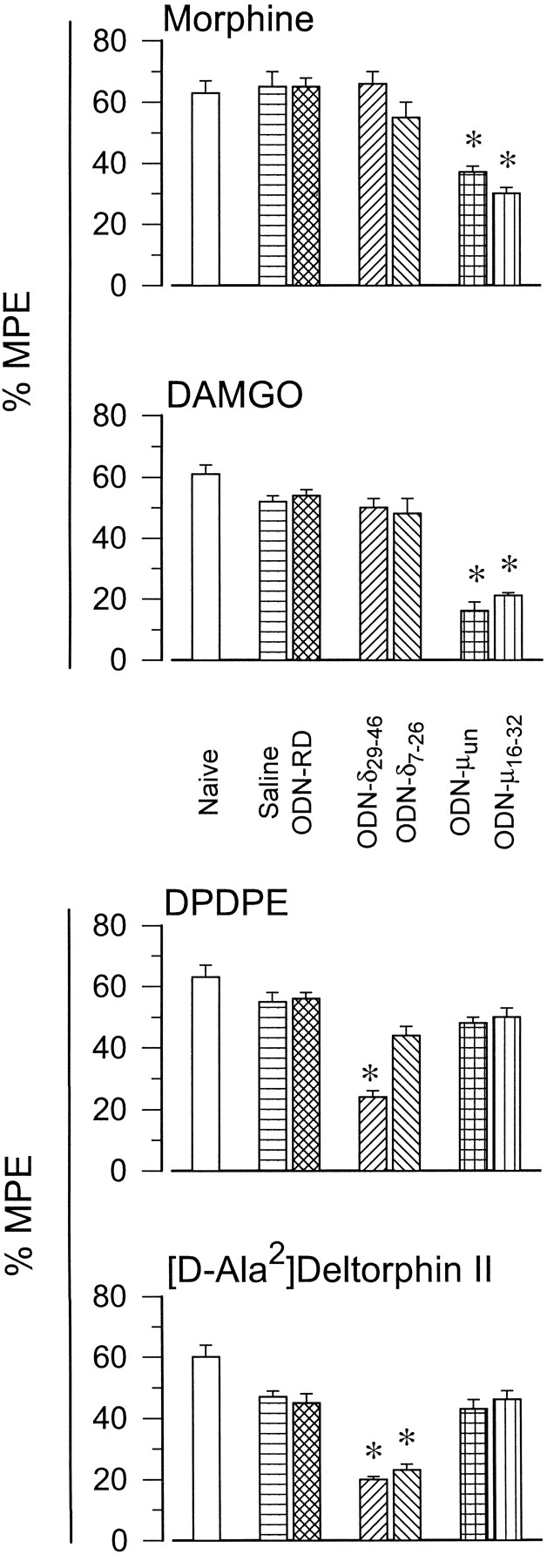
Effect of chronic i.c.v. administration of ODNs tomu and delta OR on the supraspinal analgesia induced by opioid agonists. Animals were injected with increasing amounts of the ODNs on a once-daily schedule for 5 consecutive days (see “Materials and Methods”). On day 6 the antinociceptive activity of opioids at supraspinal level was evaluated by the tail-flick test. Latencies were measured 30 min after morphine, 15 min after DAMGO or DPDPE and 10 min after [d-Ala2]deltorphin II. For each opioid and treatment (saline, ODN-RD, ODN-μ and ODN-δ) a different group of mice was used. Antinociception is expressed as %MPE. Values are the mean ± S.E.M. from groups of 10 to 15 mice each. * Significantly different from the control group receiving saline or the ODN-RD instead of the ODN to the corresponding opioid receptor type. Analysis of variance, Student-Newman-Keuls test, P < .05.
Chronic administration of ODNs to mu anddelta OR on the development of morphine tolerance.
A time-dependent decrease of the antinociceptive activity elicited by chronic morphine was observed (fig. 2). During the first hour after administration of the oily morphine suspension, analgesia reached the predetermined cut-off time of 10 s. The mice then rapidly developed tolerance to this effect, which was almost absent 24 h later. The ODNs to delta OR did not significantly alter the antinociceptive effect of morphine nor the rate of evolution of tolerance. Mice receiving the ODNs to mu OR showed inferior analgesia in response to the chronic opioid. The decrease that might take place beyond the cut-off time of 10 s could not be evaluated. ODNs to mu OR did not modify the rate at which tolerance developed to the chronic morphine administration.

Effect of ODNs to mu anddelta OR on the development of tolerance to morphine-induced antinociception. Mice were implanted at hour zero with the oily suspension containing morphine. Analgesia was evaluated by the warm water tail-flick test and expressed as a %MPE. Values are the mean ± S.E.M. from groups of 15 to 20 mice. Both ODNs tomu OR produced a similar reduction of chronic morphine-evoked antinociception. The effect of ODN-μ16–32 is shown. ODNs to delta OR lacked an effect on this paradigm. The data corresponding to ODN-δ7–26 are depicted. Control mice receiving saline or ODN-RD gave identical results. * Significantly different from the control group, ANOVA-Student-Newman-Keuls test; P < .05.
In vivo administration of ODNs to mu anddelta OR: Effect on morphine withdrawal syndrome precipitated by naloxone.
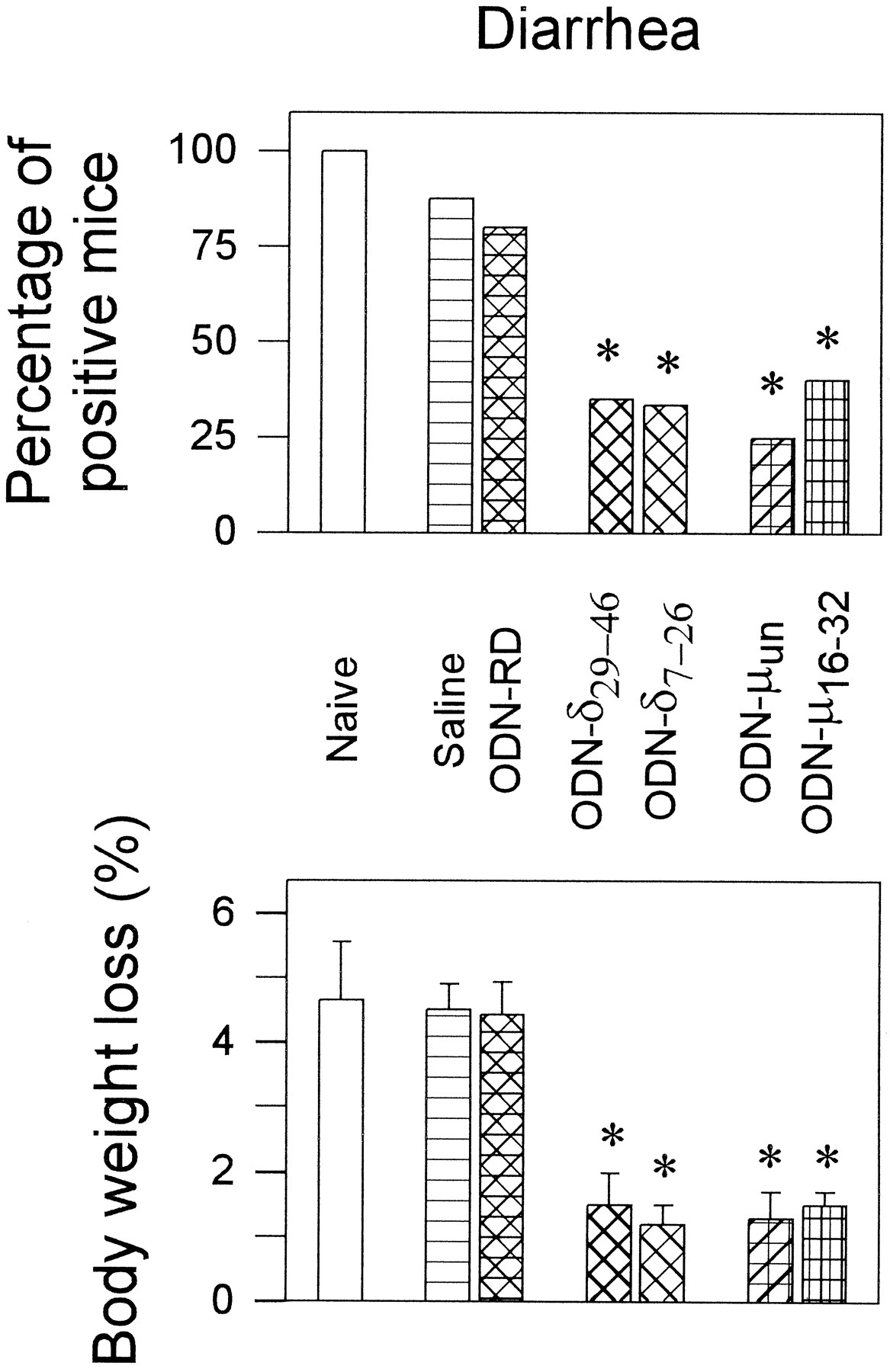
Naloxone (1 mg/kg s.c.) precipitated the withdrawal syndrome in animals that had received chronic morphine for 72 h. A daily i.c.v. injection for 5 consecutive days of saline or ODN-RD did not significantly reduce withdrawal jumping or diarrhea when compared with noninjected mice (naive) chronically exposed to morphine (figs. 3 and 4). Mice treated with antisense ODNs to mu OR before chronic morphine implantation exhibited low incidence of naloxone-elicited jumping behavior. The average number of jumps per mouse was from 24 ± 6.5 (ODN-RD-treated mice) to 4.5 ± 1,9 (ODN-μ16–32-treated) or to 1.6 ± .8 (ODN-μun-treated) (fig. 3, lower panel). The number of animals that displayed the symptom was also reduced substantially (fig.3, upper panel). This treatment also diminished the incidence of other withdrawal signs, such as loss of body weight and diarrhea (fig. 4). Thirty minutes after the administration of naloxone, the body weight of saline- or ODN-RD-treated mice was reduced by about 4.5 ± .6% (n = 12), a value significantly greater than that observed for mice injected with the ODNs to the mu OR (about 1.5%, n = 12) (fig. 4).

Treatment with ODNs to mu anddelta OR on the jumping behavior elicited by naloxone challenge in morphine-dependent mice. Mice were implanted with morphine 72 h before injecting 1 mg/kg s.c. of the opioid antagonist naloxone. Jumps were recorded during the 10 min that followed the challenge. The upper panel shows the percentage of animals presenting the symptom versus the total number of mice in the group. The average number of jumps was calculated, either as mice displaying the sign (middle panel) or with respect to all the mice in the group, including the zeroes of the animals not showing the symptom (lower panel). Groups of 16 to 24 mice were used. * Significantly different from the control group (ODN-RD), Fisher′s Exact Probability test (upper panel), or ANOVA-Student-Newman-Keuls test (middle and lower panels; values are the mean ± S.E.M.); P < .05.

Effect of ODNs to mu anddelta OR on naloxone-precipitated diarrhea and loss of body weight in morphine-dependent mice. Details as in figure 3.
Administration of the ODN-δ7–26 also alleviated certain withdrawal symptoms. Repeated i.c.v. injections of this ODN to thedelta OR reduced the percentage of animals showing withdrawal jumping (fig. 3, upper panel) and the average number of jumps of the group of mice (fig. 3, lower panel). However, the average number of jumps elicited by the jumping mice was similar to that elicited by saline or ODN-RD-treated animals (fig. 3, middle panel). The ODN-δ7–26 was also able to reduce naloxone-precipitated diarrhea and loss of body weight (fig. 4). In contrast, the ODN-δ29–46 to delta OR did not significantly reduce naloxone-precipitated jumping behavior, although it alleviated the body weight loss and diarrhea (figs. 3 and 4).
Immunoblots of mouse and rat striatum after in vivoi.c.v. administration of ODNs to mu and deltaOR.
The neural structures were obtained 6 days after commencing repeated administration of ODNs. Rats implanted with osmotic minipumps guided into the lateral ventricle were sacrificed after 3 weeks of continuous delivery of ODNs. Immunoblots of SDS-solubilized membranes from mouse striatum showed immunoreactive proteins at molecular masses of about 60 and 80 kdaltons for mu OR (fig.5, A, B and D) and 50 kdaltons for delta OR (fig. 5, C, E and F). These are glycosylated proteins because the immunosignals shifted to lower masses in the range of 40 kdaltons after enzymatic (Liu-Chen et al., 1993; Garzón et al., 1995) or chemical removal (Garzón et al., 1994) of the oligosaccharides. Glycoproteins exhibit anomalous mobility in SDS-PAGE chromatography that greatly depends on acrylamide concentration and buffer systems. These considerations might apply for the diverse masses described for these glycosylated opioid receptors (see for example Garzón et al., 1995). Both antisera to mu OR, MAS/2 and MU/2El, labeled identical bands (fig.5A). The immunoreactivity observed in control animals receiving the random sequence ODN was similar to that of naive mice. In mice undergoing repeated injections with the ODN-μun (fig. 5B), a slight but significant reduction of the mu OR-like immunoreactivity could be observed (19 ± 5%*, mean ± S.E.M. from four independent determinations; * indicates a significant difference from the group receiving the random ODN, ANOVA, Student-Newman-Keuls test, P < .05). A greater decrease, about 62 ± 7%*, was achieved infusing continuously the ODN-μun into the rat brain (fig. 5D). In this neural tissue the immunosignals associated with delta OR were not altered by treatment with ODNs to mu OR (fig. 5F). The subchronic administration of ODNs to delta OR produced small decreases of delta OR-like immunoreactivity in mouse striatum: ODN-δ7–26, 17 ± 3%*; ODN-δ29—46, 25 ± 6%* (fig. 5C). Thedelta OR-related immunosignals appeared notably diminished (54 ± 7%*) in rats receiving chronically the ODN-δ22–37 (fig. 5 E). The labeling of an antisera (S/1) directed to alpha subunits of Gi2-transducer proteins (Sánchez-Blázquez et al., 1995) was not influenced by the ODNs to delta OR (fig. 5G).

Immunoblots of SDS extracts from mouse and rat striatum with antipeptide antibodies to mu anddelta OR. Mice received repeated i.c.v. injections of the ODNs to mu and delta OR on a 5-day schedule. Rats were implanted s.c. with osmotic minipumps guided into the lateral ventricle and the ODNs were then infused continuously for 3 weeks. Equal amounts (80 μg/lane) of SDS-solubilized protein (RDvs. ODN) were loaded in each gel lane. SDS-PAGE and immunoblotting were performed as described under “Materials and Methods.” (A, B, D) Immunoreactive bands at molecular masses of about 60 and 80 kdaltons for mu OR were observed. (C, E, F) Bands of about 50 kdaltons for delta OR. (G) The S/1 antisera to Gi2α subunits labeled a 38 kdaltons band. Immunoblots were analyzed by optical densitometry, the percent of decrease in immunoreactivity (ODN to OR versus RD) was computed and expressed as mean ± S.E.M. from four independent experiments (see “Results”). The molecular weight standards used were: α2-macroglobulin (180,000), β-galactosidase (116,000), fructose-6-phosphate kinase (84,000), pyruvate kinase (63,000), fumarase (52,000), lactic dehydrogenase (41,000), triosephosphate isomerase (36,000). Biotinylated SDS molecular weight markers: phosphorylase b (subunit, 97,000), catalase (subunit, 58,100), alcohol dehydrogenase (subunit, 39,000), carbonic anhydrase (29,000), trypsin inhibitor (20,100) and lysozyme (14,300). MAS/2 and MU/2El, antibodies to the N-terminal and second extracellular loop of the cloned mu OR, respectively; Δ/1, antibody to the N-terminal of the cloned delta OR; RD, random oligodeoxynucleotide; ODN, antisense oligodeoxynucleotide to the corresponding opioid receptor.
Autoradiographic studies.
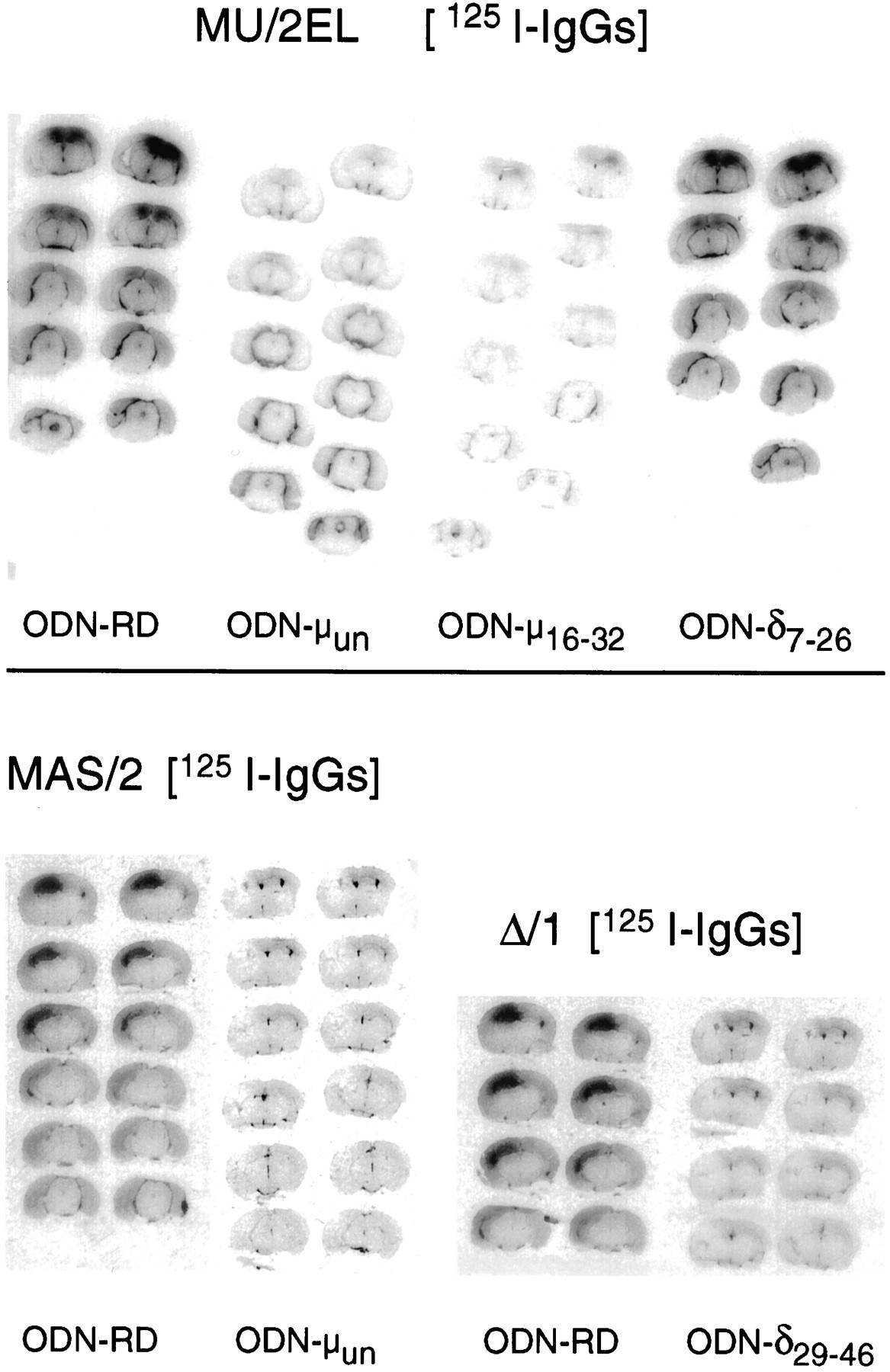
Radiolabeling of mouse brain neural structures could be observed 24 h after injecting i.c.v. affinity-purified 125I-IgGs to mu anddelta OR. The greatest amount of labeling was localized in periventricular areas. Strong radiostaining was also found over the cortical, septal and hippocampal regions. Differences in the staining achieved with MU/2EL and MAS/2 antibodies are probably caused by variations in diffusion into the brain tissue or access to some subpopulations of MU-OR. The glycosylation of the N-terminal peptide sequence of the MU-OR might account for a smaller population of receptors being tagged by MAS/2 because this antibody is directed to this region. In brain sections obtained from animals receiving denatured 125I-IgGs or preabsorbed with the corresponding antigenic peptides, the immunosignals were practically absent (not shown). The specificity of this labeling is suggested by previous results showing that these anti OR antibodies diminished the specific binding of opioid agonists to mouse brain membranes (Garzón and Sánchez-Blázquez 1995; Garzón et al., 1994, 1995). Mice chronically treated with ODNs to mu OR (ODN-μun and ODN-μ16–32) displayed a substantial reduction of the immunolabeling promoted by MU/2EL125I-IgGs. Similarly, the signals originated by MAS/2125I-IgGs were reduced in mice receiving the ODN-μun (fig. 6). These immunosignals were preserved when using the ODN7–26 directed todelta OR. The Δ/1 125I-IgGs showed a weak binding to brain sections obtained from mice injected with the ODN-δ29–46 (fig. 6). The ODN-δ7–26 also reduced the delta OR-related immunosignals without affecting these originated by the antibodies to mu OR (not shown).

In vivo i.c.v. injection of125I-IgGs to mu and delta OR: radiolabeling of mouse brain sections. Twenty-four hours after i.c.v. administration of the 125I-IgGs to mu OR (MAS/2 and MU/2EL) and delta OR (Δ/1), cryostat sections (20 μm) were cut at various levels of the neuraxis, mounted on gelatin-subbed slides, dried and placed against a tritium-sensitive film (Hyperfilm-3H; Amersham). Each ODN was studied in at least three mice. Images from representative animals are depicted. ODN-RD, random oligodeoxynucleotide; ODN, antisense oligodeoxynucleotide to the corresponding opioid receptor.
Discussion
Synthetic ODNs have been successfully used as pharmacological tools to investigate the physiology of various neural receptor systems (Wahlestedt et al., 1993; Zhang and Creese, 1993; Laiet al., 1994; Standifer et al., 1994; Tsenget al., 1994; Sánchez-Blázquez et al., 1995; Bilsky et al., 1996). Repeated in vivo injection into the lateral ventricle of an ODN complementary to bases 16 to 32 of the murine mu OR mRNA, significantly reduced the supraspinal antinociception elicited by muOR-binding agonists. Selectivity of the effect was indicated because the analgesia produced by selective agonists of delta OR was not altered. A similar profile for opioid-evoked antinociception could be observed for the ODN-MORun directed to bases −87 to −69 upstream from the initiation codon ATG (Rossi et al., 1994). The ODN to nucleotides 7 to 26 of the cloned delta OR diminished the analgesic effect promoted by the agonist atdelta-2 OR, [d-Ala2]deltorphin II, without affecting the pattern exhibited by DPDPE, an agonist atdelta-1 OR. These data confirm and extend previous reports describing the possibility of selectively inhibiting antinociception produced by delta-2 OR-binding agonists (Bilsky et al., 1994, 1996; Lai et al., 1994). The repeated administration of an ODN complementary to nucleotides 29 to 46 of the murine delta OR (Standifer et al., 1994) inhibited the antinociception elicited by DPDPE (delta-1) and [d-Ala2]deltorphin II (delta-2) in the mouse. No changes were observed for themu OR-binding agonists DAMGO and morphine.
It has been reported that cloned mu OR mediates morphine-evoked antinociception in rats. Microinjection into the periaqueductal grey matter of an ODN with a sequence complementary to the 5′-untranslated region of the rat MOR-1 receptor clone completely blocked the analgesic profile exhibited by morphine (Rossi et al., 1994). Moreover, i.c.v. administration of an oligo targeted at the first 18 nucleotides of the coding region of the ratmu OR diminished the antinociceptive actions of morphine as determined by the cold water tail-flick test (Cheng et al., 1995). Probably by reducing the number of functional receptors, the ODNs to cloned mu OR greatly decreased the antinociceptive potency of mu OR-binding agonists. These ODNs also diminished the antinociception promoted by s.c. implantation of morphine. Tolerance to this impaired analgesia was observed. In these mice a weak dependence and/or expression of morphine withdrawal syndrome was also seen. This substantiates the idea that tolerance to and dependence on opioids are parts of a common mechanism (Way et al., 1969). Repeated administration for 5 days of either ODN16–32 or ODNun to mu OR, before starting chronic morphine, promoted a significant reduction of naloxone-precipitated withdrawal symptoms. Morphine-dependent mice injected with the ODNs to mu OR, although exhibiting a reduced morphine withdrawal syndrome, showed no better appearance than dependent mice that had received saline or the ODN-RD. This observation is in contrast to that described for compounds such as clonidine or the N-acetyl derivative of the endogenous opioid peptide β-endorphin. These substances not only alleviated morphine withdrawal syndrome but also improved the general appearance of tolerant-dependent mice (Garzón and Sánchez-Blázquez, 1992). It is rather likely that clonidine and N-acetyl β-endorphin, acting at neural substrates other than mu OR (G proteins, cAMP production, etc.), trigger a series of responses that counteract the effects of chronic morphine. The ODNs to mu OR reduced the number of receptors available to morphine, thus impairing the first step leading to the development of tolerance and dependence. However, they apparently did not induce responses to neutralize the effects of the opioid.
It is generally accepted that mu OR drives the initial step in both the relief of pain as well as the addictive side effects of morphine. In agreement with this idea, high levels of mu OR mRNAs are detected in the thalamic structures and the periaqueductal grey matter, where mu OR agonists modulate pain transmission. The mRNA is also found in regions associated with the expression of morphine withdrawal syndrome, e.g., the locus ceruleus (Mansour et al., 1995). The withdrawal syndrome might be considered as the summation of several separate actions originating at the pharmacologically defined subtypes of muOR, mu-1 and mu-2 (Ling et al., 1984). The i.c.v. injection into mice of antibodies generated against the second extracellular loop of the mu OR (MU/2EL), or the first 16 amino acids of its N terminus (MAS/2), has also been shown to reduce analgesia and the expression of multiple symptoms of withdrawal syndrome (mu-1 and mu-2 mediated) (Garzón and Sánchez-Blázquez, 1995; Garzón et al., 1995; Sánchez-Blázquez et al., 1996). Accordingly, treatment with ODNs or antibodies to mu OR did not separate the effects assigned to each of the pharmacologically defined subtypes of the mu OR (Ling et al., 1984). It is therefore conceivable that both subtypes, mu-1 and mu-2 OR, are highly homologous proteins. The alternative splicing of precursor mRNA (Zimprich et al., 1995) or the existence of various glycosylated forms (Liu-Chen et al., 1993; Garzón and Sánchez-Blázquez, 1995; Garzónet al., 1995) of this mu OR might account for the pharmacological subtypes.
Notably, a series of studies performed in the mouse led to the proposal of delta OR being involved in the development of morphine dependence (Abdelhamid et al., 1991). Miyamoto et al. (1993) demonstrated that continuous blockade ofdelta-2 opioid receptors by the long-lasting antagonist 5′NTII inhibited the development of physical dependence on chronic morphine. In agreement with these findings, ODN7—26, which selectively blocks delta-2 OR-evoked analgesia (Lai et al., 1994; Bilsky et al., 1996; present work), markedly reduced the number of animals exhibiting the naloxone-precipitated jumping behavior. Both ODN7–26 and ODN29–46to delta OR promoted reductions in the incidence ofmu-2 OR-related withdrawal symptoms, i.e., weight loss and diarrhea. ODN29–46, which reduced bothdelta-1 OR- and delta-2 OR-evoked antinociception, lacked effect on the jumping behavior promoted by naloxone in morphine-tolerant dependent mice. These differences in the activity of the ODNs to delta OR might be interpreted in terms of nonspecific effects such as alterations of nonopioid systems. However, the possibility exists that, in delta-2 OR-deficient mice, treated with the ODN7–26 or receiving 5′NTII, the endogenous ligands reduce the incidence of morphine withdrawal jumping behavior by acting at delta-1 OR. This idea implies that activation of molecular subtypes of thedelta OR may lead to distinct/opposite roles in the chronic effects of morphine. In addition, the data obtained with different ODNs to this cloned delta receptor indicate that both subtypes might have closely related amino acid sequences.
Effective treatment with an ODN is expected to reduce the levels of the protein encoded by the targeted mRNA. In this work, the muand delta OR were labeled by previously characterized specific antibodies (Garzón et al., 1994, 1995). After five repeated injections of the ODNs to mu anddelta OR, Western blotting analysis of synaptosomal fractions from mouse brain areas revealed small reductions in these proteins. This agrees with previous studies with ODNs which show dramatic alterations in behavior accompanied by modest changes in protein abundance (Wahlestedt et al., 1993). To promote consistent decreases of these mu and delta OR in the present work, ODNs had to be continuously infused into the lateral ventricle of the brain for several weeks. Therefore, the turnover of these opioid receptors appears to be slower than that of other cell membrane proteins, i.e., the alpha subunits of G transducer proteins (Sánchez-Blázquez et al., 1995). An important reduction of mu and deltaOR-like immunoreactivity could be attained when labeling was performedin vivo to mice treated for 5 days with the ODNs to these receptors. It seems that ODNs preferentially affect a small pool of receptors of recent synthesis or with rapid turnover. These receptors appear to be correctly orientated in the cellular membrane because the antibodies after i.c.v. injections were able to bind antigenic peptide sequences in the extracellular domains. The major pool of receptors, probably old and lacking functionality, might be located extrasynapticaly. This pool would be much less affected by ODN treatment (Wagner, 1994; Zhou et al., 1994) and not recognized by the in vivo injected antibodies. Thus, the possibility of a wider populations of receptors being tagged in Western blotting than in in vivo-initiated autoradiographic studies cannot be disregarded.
In summary, this study offers experimental evidence of the efficacy of the antisense strategy in the in vivo blocking of the synthesis of specific receptor proteins. It also shows the participation by cloned mu OR in antinociception and the addictive effects of morphine. It also confirms the involvement ofdelta-2 OR in the development and/or expression of morphine dependence without compromising antinociception produced bymu OR-binding agonists.
Footnotes
-
Send reprint requests to: Pilar Sánchez-Blázquez, Neurofarmacologı́a, Instituto Cajal, C.S.I.C., C/Doctor Arce, 37, 28002 Madrid, España (Spain).
-
↵1 Supported by CICYT SAF93–0058 and SAF95–0003.
- Abbreviations:
- ODN
- antisense oligodeoxynucleotide
- RD
- random oligodeoxynucleotide
- OR
- opioid receptor
- DAMGO
- [d-Ala2,N-MePhe4,Gly-ol5]enkephalin
- DPDPE
- [d-Pen2,5]enkephalin
- i.c.v.
- intracerebroventricular
- s.c.
- subcutaneous
- SDS
- sodium dodecyl sulfate
- PAGE
- polyacrylamide gel electrophoresis
- %MPE
- percentage of maximum possible effect
- ANOVA
- analysis of variance
- IgG
- immunoglobulin G
- Received August 5, 1996.
- Accepted November 18, 1996.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}