

Abstract
Because multiple molecular signal transduction pathways regulate cerebellar long-term depression (LTD), which is thought to be a possible molecular and cellular basis of cerebellar learning, the systematic relationship between cerebellar LTD and the currently known signal transduction pathways remains obscure. To address this issue, we built a new diagram of signal transduction pathways and developed a computational model of kinetic simulation for the phosphorylation of AMPA receptors, known as a key step for expressing cerebellar LTD. The phosphorylation of AMPA receptors in this model consists of an initial phase and an intermediate phase. We show that the initial phase is mediated by the activation of linear cascades of protein kinase C (PKC), whereas the intermediate phase is mediated by a mitogen-activated protein (MAP) kinase-dependent positive feedback loop pathway that is responsible for the transition from the transient phosphorylation of the AMPA receptors to the stable phosphorylation of the AMPA receptors. These phases are dually regulated by the PKC and protein phosphatase pathways. Both phases also require nitric oxide (NO), although NO per se does not show any ability to induce LTD; this is consistent with a permissive role as reported experimentally (Lev-Ram et al., 1997). Therefore, the kinetic simulation is a powerful tool for understanding and exploring the behaviors of complex signal transduction pathways involved in cerebellar LTD.
- cerebellar long-term depression
- kinetics
- simulation
- signal transduction
- positive feedback loop
- phosphorylation
Cerebellar long-term depression (LTD), thought to be a molecular and cellular basis for cerebellar learning (Ito, 1989; Lisberger, 1998; Kawato, 1999), is a process involving a decrease in the synaptic strength between parallel fiber (PF) and Purkinje cells (PCs) induced by the conjunctive activation of PFs and climbing fiber (CF) (Ito, 1989; Linden and Connor, 1995). Multiple signal transduction pathways have been shown to be involved in this process (Linden and Connor, 1993; Fiala et al., 1996; Daniel et al., 1998). PF is thought to transmit its signal through glutamate (Kano and Kato, 1987; Linden et al., 1991) and nitric oxide (NO) (Shibuki and Kimura, 1997; Daniel et al., 1998), and CF is thought to transmit its signal through corticotropin-releasing factor (CRF) (Miyata et al., 1999) and Ca2+ influx via voltage-gated Ca2+ channels (Ekerot and Kano, 1985; Sakurai, 1990; Crepel and Jaillard, 1991; Linden and Connor, 1993; Daniel et al., 1998). Both PF- and CF-mediated signals are transmitted into PCs through multiple signaling pathways, including mitogen-activated protein (MAP) kinase and Ca2+, and finally lead to the phosphorylation of AMPA receptors by protein kinase C (PKC) (Fiala et al., 1996; Daniel et al., 1998; Matsuda et al., 2000). In addition, NO, by diffusing into PCs, inhibits protein phosphatase through the cGMP and soluble guanylnyl cyclase (sGC) pathways, resulting in inhibition of the dephosphorylation of AMPA receptors (Daniel et al., 1998). Therefore, the signals from PF and CF dually regulate the phosphorylation of AMPA receptors through the PKC and protein phosphatase pathways. The phosphorylation of AMPA receptors has been shown to be a key step for cerebellar LTD expression through internalization (Matsuda et al., 2000; Wang and Linden, 2000). However, because of the complex nature of the signaling pathways underlying the phosphorylation of AMPA receptors in cerebellar LTD, the systematic relationship between the synaptic inputs and the output responses remains obscure.
To understand and explore the behaviors of the complex signal transduction pathways, it is important to use the computational framework of kinetic simulation. Accordingly, by taking advantage of the recently developed program GENESIS/kinetikit (Bhalla and Iyengar, 1999), we built a computational simulation model for the phosphorylation of AMPA receptors in cerebellar LTD on the basis of kinetic parameters. It was reasonable to incorporate only biochemical parameters of post-translational biochemical reactions, but not those for gene expression and protein synthesis, into the simulation, simply because no kinetic parameters of gene expression and protein synthesis have thus far been available in cerebellar LTD. The phosphorylation of AMPA receptors in the kinetic model consisted of the initial and intermediate phases; the former was mediated by Ca2+, diacylglycerol (DAG), and arachidonic acid (AA)-mediated PKC pathway and the latter by the MAP kinase-mediated positive feedback loop pathway. In addition, NO was required for both the initial and intermediate phases, consistent with a permissive role as shown experimentally (Lev-Ram et al., 1997). Therefore, the kinetic simulation of cerebellar LTD provided us with a novel method for understanding and exploring the complex nature of the signal transduction pathways involved in cerebellar LTD.
MATERIALS AND METHODS
Block diagram showing the phosphorylation of AMPA receptors in cerebellar LTD. The conjunctive stimulation of PF and CF has been shown to elicit cerebellar LTD (Ito, 1989). In addition, multiple signal transduction pathways have been shown to regulate cerebellar LTD (Ito, 1989; Linden and Connor, 1993; Fiala et al., 1996;Daniel et al., 1998). According to the literature described below, a new block diagram of cerebellar LTD was reconstructed (see Fig.1A). Cerebellar LTD has been shown to be mediated by the modulation of AMPA receptors (Linden and Connor, 1993; Daniel et al., 1998), and the decrease in the EPSP has been thought to be mediated by the internalization of phosphorylated AMPA receptors (Matsuda et al., 2000; Wang and Linden, 2000). PF mediates its signals to PCs by releasing glutamate, resulting in the activation of AMPA receptors and type-1 G-protein-coupled metabotropic receptors (mGluR1s) (Aiba et al., 1994; Conquet et al., 1994; Daniel et al., 1998; Ichise et al., 2000). The activation of AMPA receptors leads to an influx of Ca2+ through the activation of a Na+/Ca2+ pump (Brorson et al., 1994; Daniel et al., 1998) and to the activation of MAP kinase cascades through the activation of Lyn tyrosine kinase (Hayashi et al., 1999). The activation of mGluR1 leads to the activation of GTP-binding proteins, Gq, and the subsequent activation of phospholipase C (PLC), resulting in the production of DAG and inositol-1,4,5-phosphate (IP3) (Linden and Connor, 1993; Fiala et al., 1996; Daniel et al., 1998). IP3 mobilizes Ca2+from the internal Ca2+ store through IP3 receptors (Finch and Augustine, 1998; Takechi et al., 1998). The elevation of Ca2+ leads to the activation of cytosolic phospholipase A2(PLA2), resulting in the production of AA and the subsequent activation of PKC (Bhalla and Iyengar, 1999). PKC has been shown to be involved in the induction of cerebellar LTD (Crepel and Krupa, 1988; Linden and Connor, 1991). DAG, Ca2+, and AA activate protein kinase C (Nishizuka, 1992).
In addition, PF has recently been shown to produce NO (see Fig.1B) (Shibuki and Kimura, 1997). NO, diffusing into PCs, activates sGC, and the activated sGC catalyzes GTP into cGMP (Bredt and Snyder, 1992; Stone and Marletta, 1996; Lev-Ram et al., 1997; Daniel et al., 1998). Then, cGMP activates cGMP-dependent protein kinase (PKG) (Wang and Robinson, 1997). PKG phosphorylates its substrate, G-substrate (Nairn et al., 1985; Wang and Robinson, 1997). Recently, cDNA of G-substrate was cloned (Endo et al., 1999; Hall et al., 1999), and it was shown that phosphorylated G-substrate preferentially inhibits protein phosphatase 2A (PP2A) rather than protein phosphatase 1 (Endo et al., 1999). In this simulation, therefore, we assumed that the dephosphorylation of AMPA receptors is mediated by PP2A. CF has been known to depolarize PCs, resulting in the influx of Ca2+ into PCs through voltage-gated Ca2+ channels (Ito, 1989;Linden and Connor, 1993; Daniel et al., 1998). In this study, therefore, Ca2+ elevation induced by the stimulation of PF and CF, or PF or CF alone, was assumed on the basis of the experimental results using Ca2+indicators (Miyakawa et al., 1992; Midtgaard et al., 1993; Kohda et al., 1995; Wang et al., 2000), not by the kinetic simulation. Recently, CRF found in CF was shown to play a permissive role in cerebellar LTD (Miyata et al., 1999) and to activate MAP kinase without Ca2+ elevation (Rossant et al., 1999). In this simulation, therefore, MAP kinase was activated by PKC, Lyn, and CRF through the activation of Raf and MAP/extracellular signal-regulated kinase (MEK). Activated MAP kinase phosphorylates and activates PLA2, resulting in the production of AA and the subsequent activation of PKC (Bhalla and Iyengar, 1999). Therefore, the PKC–MAP kinase pathway interacts at this point. This connection should result in a positive feedback loop (Bhalla and Iyengar, 1999). In this study, therefore, we attempted to set the parameters to fit the time course of the phosphorylation of the AMPA receptors to that of EPSP as reported previously (Chen and Thompson, 1995). We built the NO/cGMP pathway according to literature (see Fig. 1B, Table 1). All of the kinetic parameters in Figure 1B are shown in Table 1. All of the other kinetic parameters used in this study were based on previous reports (Bhalla and Iyengar, 1999) and are shown on our web site (http://www.erato.atr.co.jp/∼kuroda/supplementary_info.html). All of the inputs used in this study are also shown in Appendix (see Fig. 6).
Kinetic simulations. The phosphorylation of AMPA receptors in cerebellar LTD was simulated based on the following two biochemical reactions: protein–protein (molecule–molecule) interactions and enzymatic reactions. The protein–protein interactions included interactions such as NO–GS and cGMP–PKG. These reactions can be given by the following formulation:
 Experimentally, in most cases, Kf andKb are not available in literature. However,Kd, the dissociation constant, has generally been reported. Therefore, based on the reportedKd values,Kf andKb were calculated by the following definition:
Experimentally, in most cases, Kf andKb are not available in literature. However,Kd, the dissociation constant, has generally been reported. Therefore, based on the reportedKd values,Kf andKb were calculated by the following definition:
 If A is greatly in excess, the total amount of product, [AB], that can be formed is limited by the amount of B. Therefore, the reactions should follow the pseudo first-order reactions. When the half time (t½) of the pseudo first-order reaction is experimentally obtained, Kf andKb can be calculated by the following equation or by plotting [A] versus apparent rate constant, which can be estimated from t½, giving a line with a y-intercept of Kb and slope of Kf:
If A is greatly in excess, the total amount of product, [AB], that can be formed is limited by the amount of B. Therefore, the reactions should follow the pseudo first-order reactions. When the half time (t½) of the pseudo first-order reaction is experimentally obtained, Kf andKb can be calculated by the following equation or by plotting [A] versus apparent rate constant, which can be estimated from t½, giving a line with a y-intercept of Kb and slope of Kf:
 The enzymatic reactions include phosphorylation and dephosphorylation. These reactions can be given by the following formulation of Michaelis-Menten:
The enzymatic reactions include phosphorylation and dephosphorylation. These reactions can be given by the following formulation of Michaelis-Menten:
 where E, S, and P denote enzyme, substrate, and product, respectively.
where E, S, and P denote enzyme, substrate, and product, respectively.
As in the case of protein–protein interactions, the values ofK1 andK2 are not generally given in literature. However, K3 can be calculated by the experimentally shownKcat value, given byVmax divided by the concentration of the enzyme. The values of Km are also generally reported. Therefore, on the basis of theKm and K3 values, the values of K1 and K2 were calculated by the following definition:
 Unless apparent rate constants are available, we assume thatK2 is ∼2–20× greater thanK3 because K2 is generally greater than K3 in many enzymes.
Unless apparent rate constants are available, we assume thatK2 is ∼2–20× greater thanK3 because K2 is generally greater than K3 in many enzymes.
Once the above parameters determined on the basis of the kinetic values were found to be robust enough to reproduce cerebellar LTD elicited under normal conditions (Chen and Thompson, 1995), we simulated the following experiments. All of the numerical computations were performed with the kinetics library, which is an extension to GENESIS (Bhalla and Iyengar, 1999).
RESULTS
Building a kinetic simulation model for the phosphorylation of AMPA receptors in cerebellar LTD
To develop a computational kinetics simulation for the phosphorylation of AMPA receptors in cerebellar LTD, we first developed a block diagram for the phosphorylation of AMPA receptors in cerebellar LTD on the basis of reported data (Fig.1A). Because recent evidence has suggested that the phosphorylation of AMPA receptors is a key step for the expression of cerebellar LTD through the internalization of the phosphorylated AMPA receptors (Matsuda et al., 2000; Wang and Linden, 2000; Xia et al., 2000), we measured concentrations of phosphorylated and nonphosphorylated AMPA receptors. According to literature, cerebellar LTD involves the phosphorylation of AMPA receptors of PCs by both the activation of PKC (Crepel and Krupa, 1988; Linden and Connor, 1993; Fiala et al., 1996; Daniel et al., 1998;Matsuda et al., 2000) and the inhibition of protein phosphatase by the NO/cGMP pathway (Nairn et al., 1985; Ito and Karachot, 1992; Ajima and Ito, 1995; Daniel et al., 1998). Raf, MAP kinase, and PKC have been shown to form a potential positive feedback loop (Bhalla and Iyengar, 1999) (Fig. 1A, pink andyellow lines), and PKC has been shown to be regulated by the Ca2+-, DAG-, and AA-mediated linear pathway (Fig. 1A, blue and yellow lines) and by the positive feedback loop pathway. Because PP2A has been shown to be phosphorylated by PKG (Endo et al., 1999), we assume that the action of the NO/cGMP pathway is finally mediated by PP2A (Fig. 1A, green line, B, Table1). Therefore, the phosphorylation of AMPA receptors is dually regulated by PKC and PP2A. In the block diagram (Fig. 1A), the number of initial concentrations of molecules was 28. Of the 28, 22 were determined by literature (Table 1) or by the kinetic simulation (Bhalla and Iyengar, 1999), and 6 were assumed. The numbers of protein–protein interactions and enzymatic reactions were 30 and 25, respectively. Regarding the Kd values, 24 of a total of 30 reactions were determined by literature or by the kinetic simulation (Bhalla and Iyengar, 1999), and 6 were assumed. Regarding the values of Km andKcat, 20 of a total of 25 reactions were determined by literature or by the kinetics simulation (Bhalla and Iyengar, 1999), and 5 were assumed. The bistable behavior of the phosphorylation of AMPA receptors in cerebellar LTD (see below) was robust over a wide range of most of the assumed parameters, but it was very sensitive to parameters for PKC–Raf and PKC–AMPA receptors because the former includes parameters for the intersection point of the positive feedback loop (see below), and the latter includes direct parameters for the phosphorylation of AMPA receptors. Taking advantage of the kinetic parameters available in literature with some assumptions, we built the computational kinetic simulation model of the phosphorylation of AMPA receptors in cerebellar LTD.

A, Block diagram of the phosphorylation of AMPA receptors in cerebellar LTD. The stimulation of PF and CF results in the elevation of the phosphorylation of AMPA receptors through the activation of PKC via the activation of both linear cascades, including DAG, Ca2+, and AA (blue and yellow lines), and a positive feedback loop including PLA2(pink and yellow lines), and through the inhibition of protein phosphatase 2A (green line). The dashed-line boxindicates the generation of Ca2+, which was reconstituted by the previous observation (Miyakawa et al., 1992;Midtgaard et al., 1993; Kohda et al., 1995; Wang et al., 2000) but not by the kinetics simulation. The arrows andbars denote the stimulatory and inhibitory pathways, respectively. B, Detailed block diagram of the NO/cGMP pathway. The bidirectional arrows denote reversible reactions, and the unidirectional arrows denote irreversible reactions. Enzymes are located in the middle of the segment. The numbers indicate the reactions that have kinetic parameters shown in Table 1.PDE, Phosphodiesterase.
Kinetic parameters in the NO/cGMP pathway
The conjunctive stimulation of PF and CF induces the stable phosphorylation of AMPA receptors in cerebellar LTD
On the basis of the kinetic parameters, we tried to make the kinetic simulation reproduce the experimental results, i.e., that the conjunctive stimulation of PF and CF induces cerebellar LTD (Chen and Thompson, 1995). Cerebellar LTD consists of an initial peak followed by a stable phase (Chen and Thompson, 1995). It has been shown that gene expression and protein synthesis are required for the late phase of cerebellar LTD (Linden, 1996); however, these experimental conditions are different from those of the kinetic simulation because, under the experimental conditions, only PCs were used, and the noninvolvement of NO was assumed (Linden, 1996). By considering the fact that only post-translational biochemical reactions were incorporated into the kinetic simulation, it is reasonable to assume that the stable phase of LTD can be divided into two phases: an intermediate phase maintained by the post-translational biochemical reactions and a late phase maintained by the protein synthesis and gene expression. Moreover, because the time course of the phosphorylation of AMPA receptors in cerebellar LTD has not been determined and the phosphorylation of AMPA receptors is important for the expression of cerebellar LTD, we assumed here that the phosphorylation of AMPA receptors also consists of three phases (initial peak, intermediate phase, and late phase) and built the simulation to reproduce the time course of EPSP in cerebellar LTD. In the kinetic simulation, the conjunctive stimulation of PF and CF was found to induce the phosphorylation of AMPA receptors and to reduce the concentration of nonphosphorylated AMPA receptors (Fig.2A). The stimulation of either PF or CF alone failed to induce the stable phosphorylation of AMPA receptors in cerebellar LTD (Fig.3). The time course of the phosphorylation of AMPA receptors consists of three phases: an initial phase including the initial peak of the phosphorylated AMPA receptors (0–10 min), an intermediate phase including the sustained phosphorylation of AMPA receptors (10–40 min), and a late phase including the disappearance of the AMPA receptor phosphorylation (>40 min). The time course of the nonphosphorylated AMPA receptors in the initial and intermediate phases is similar to that of the amplitude of EPSP evoked by the conjunctive stimulation of PF and CF (Chen and Thompson, 1995) (Fig. 2A). In the late phase, however, the concentration of the nonphosphorylated AMPA receptors begins to increase and reaches the basal level at ∼90 min because of the dephosphorylation of the AMPA receptors.
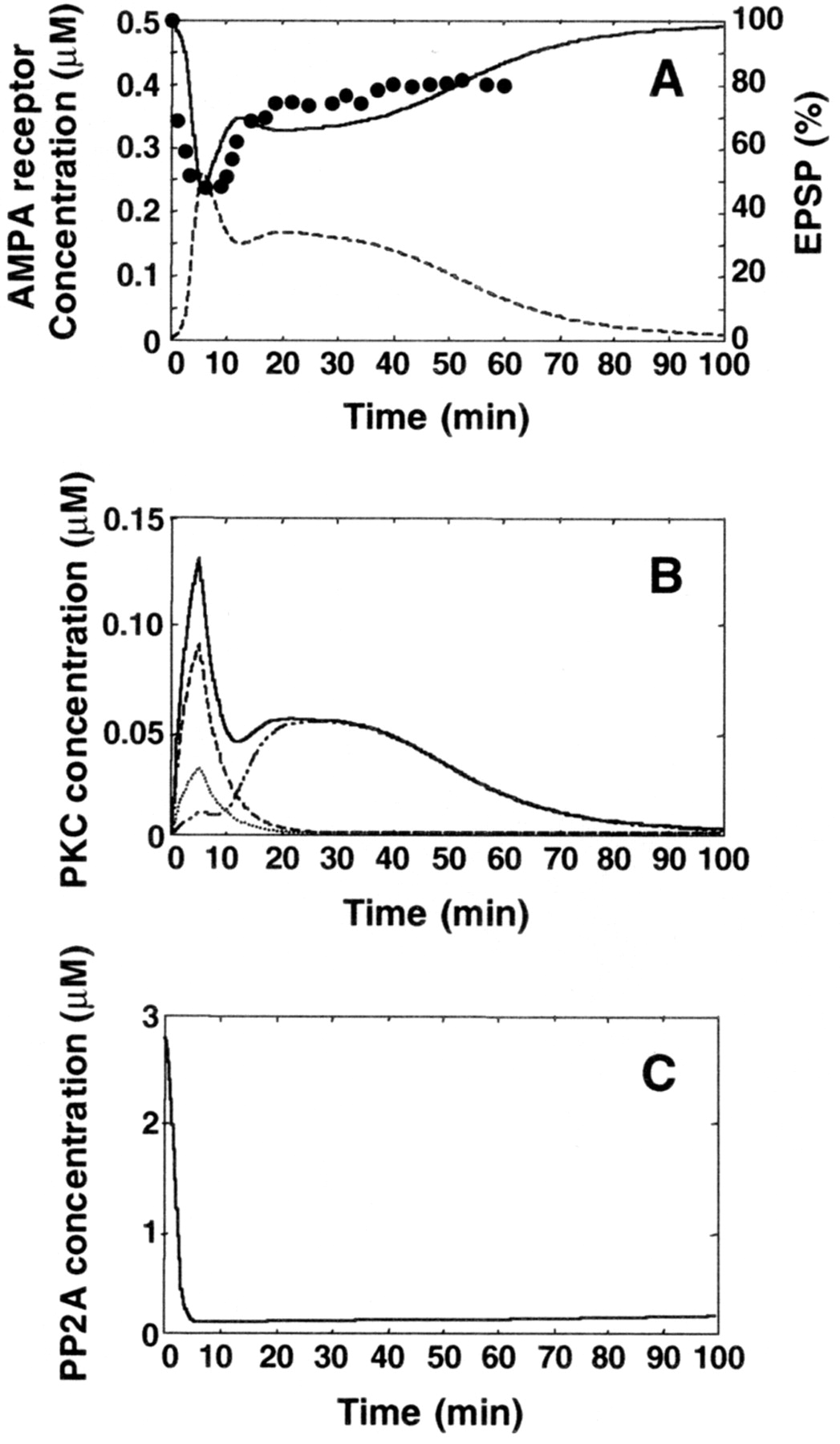
Conjunctive stimulation of PF and CF to induce the stable phosphorylation of AMPA receptors. The simulation was run at 1 Hz for a 5 min conjunctive stimulation of PF and CF. The time course of the indicated molecule concentration is plotted. A, The phosphorylation of AMPA receptors induced by the conjunctive stimulation of PF and CF. The dashed line indicates the concentration for the phosphorylated AMPA receptors. The dark line indicates the concentration of the nonphosphorylated AMPA receptors. The changes of EPSP induced by the 10 min conjunctive stimulation of PF and CF is replotted (●) from the earlier observation (Chen and Thompson, 1995). Note that in the kinetic simulation, the 5 min stimulation of PF and CF is used because 5 min stimulation is optimal according to a previous report (Karachot et al., 1994). B, The activation of PKC by the stimulation. Thedark line indicates the total PKC activity; thedashed line indicates Ca2+-activated PKC activity; the dotted line indicates DAG-activated PKC activity; the dashed–dotted line indicates AA-activated PKC activity. C, The inhibition of PP2A activity.

Requirement for the conjunctive stimulation of PF and CF to induce the stable phosphorylation of AMPA receptors. The simulation was run under conditions similar to those in Figure2A except that the following simulations were used: both PF and CF, PF alone, or CF alone. The time courses of the indicated molecules by the conjunctive stimulation of PF and CF (straight line), by the stimulation of PF alone (dotted line), and by the stimulation of CF alone (dashed line) are plotted. A, The phosphorylated AMPA receptors; B, active PKC;C, active PP2A; D, active MAP kinase;E, active PKG.
Consistently, the activation of PKC also consists of three phases (Fig.2B): an initial phase mediated by the direct activation by Ca2+, DAG, and AA, an intermediate phase mediated by the activation of the MAP kinase-mediated positive feedback loop (Fig. 3D), and a late phase mediated by the inactivation of PKC activity (see also Fig. 5). This positive feedback loop is essential for the transition from the transient phosphorylation of AMPA receptors to the stable phosphorylation of the AMPA receptors and is responsible for the bistability of the AMPA receptor phosphorylation (Figs.4, 5).

Optimal duration of the stimulation to induce the stable phosphorylation of AMPA receptors in cerebellar LTD. The duration of the conjunctive stimulation of PF and CF at 1 Hz was varied between 0 and 15 min with 1 min intervals, and the simulation was run. The concentrations of the phosphorylated AMPA receptors (A), active PKC (B), active PP2A (C), and active MAP kinase (D) at 30 min after the onset of the stimulation are shown.

The role of each signaling molecule and pathway in inducing the phosphorylation of AMPA receptors in cerebellar LTD. To explore the roles of the signaling molecules and pathways in inducing the phosphorylation of AMPA receptors in cerebellar LTD, the simulation was run under the same conditions as in Figure2A except that the concentration of the indicated molecules shown below was held at the basal level or the indicated pathway was deleted. A, The phosphorylated AMPA receptors; B, active PKC; C, active PP2A;D, active MAP kinase; E, active PKG.Blue line, None; yellow line, PKC; green line, NO; pink line, Ca2+; cyan line, PLA2; black line, DAG; brown line, PLA2 activated by Ca2+elevation; purple line, PLA2 activated by MAP kinase. Note that the time courses of the active PP2A (C) or active PKG (E) were exactly the same, except without NO (green), and expressed as a single blue line.
The PP2A activity markedly decreases by the stimulation and becomes lowest at 5 min and then gradually reactivates (Fig. 2C). The inactivation of PKC and the reactivation of PP2A lead to the dephosphorylation of the AMPA receptors, resulting in the disappearance of the phosphorylated AMPA receptors in the late phase. The reactivation of PP2A is mediated by the degradation of NO. Although NO is degraded very fast and consequently the PKG activity returns to the basal level after the stimulation (Fig. 3E), the reactivation of PP2A is very slow because the dephosphorylation step of G-substrate, an inhibitor of PP2A, is assumed to be quite slow. Because PP2A activity is inhibited by G-substrate phosphorylated by PKG, it is interesting to test whether this prolonged phosphorylation of G-substrate really occurs in vivo if antibody that specifically recognizes the phosphorylated G-substrate will be available in the future. The inactivation of PKC activity is mediated by the degradation of PKC itself and by the reactivation of PP2A activity. Although the reactivation of PP2A is very small, this small reactivation of PP2A blocks the activation of the positive feedback loop, and the activity of the loop becomes lower than the threshold, which is a critical point for bistability, resulting in the transition from the activation state to the inactivation state of the loop. These results indicate that the current signal transduction pathways as shown in Figure 1 are able to produce the initial and intermediate phases for the phosphorylation of AMPA receptors in cerebellar LTD elicited by the conjunctive stimulation of PF and CF.
Requirement for the conjunctive stimulation of PF and CF to induce the stable phosphorylation of AMPA receptors
Cerebellar LTD has been shown to be induced by the conjunctive stimulation of PF and CF, but not by the stimulation of either PF or CF alone (Ito, 1989; Daniel et al., 1992; Hemart et al., 1994; Chen and Thompson, 1995). We therefore asked whether the stable phosphorylation of AMPA receptors also requires the conjunctive stimulation of both PF and CF. In the kinetic simulation, the stimulation of PF alone did not induce the intermediate phase of the phosphorylation of AMPA receptors, although the initial phase was induced to a lesser extent (Fig. 3,dotted line). The stimulation of CF did not induce either the initial or intermediate phase (Fig. 3, broken line). The difference between the initial phase by the stimulation of PF and that by the stimulation of CF was caused by the absence of NO production by the stimulation of CF, but not by that of PF. However, an experimental stimulation similar to the CF alone stimulation in the kinetic simulation has been shown to induce an initial sharp decrease in the EPSC (Daniel et al., 1992; Hemart et al., 1994). The failure of the kinetic simulation to reproduce this observation raises the possibility that other mechanisms in addition to the phosphorylation of AMPA receptors underlie the initial phase of cerebellar LTD (see Discussion).
Optimal duration of the stimulation to induce the stable phosphorylation of AMPA receptors in cerebellar LTD
We next examined what length of time of the conjunctive stimulation by PF and CF is required to induce the stable phosphorylation of AMPA receptors. In the simulation, the stable phosphorylation of AMPA receptors was not induced by a time <3 min (Fig. 4A). Time >5 min induced the stable phosphorylation of AMPA receptors, and the optimal duration of the stimulation was found to be 6 min. The optimal duration in the simulation (6 min) of the phosphorylation of AMPA receptors was almost the same as that of the cerebellar LTD observed in an experiment (5 min) (Karachot et al., 1994). This result indicates that the phosphorylation of AMPA receptors shows bistability within a middle time scale of up to 40 min. The activation of PKC also showed a similar bistability (Fig. 4B). The PP2A activity first decreased linearly and then reached a plateau (Fig. 4C), indicating that the inhibition of PP2A does not exhibit bistability. Therefore, the bistability in the phosphorylation of AMPA receptors is mediated by the prolonged activation of PKC. The activation of the MAP kinase-mediated positive feedback loop has been reported to be responsible for the bistability (Fig. 4D) (Bhalla and Iyengar, 1999). In addition, we found that the inhibition of PP2A is required for this bistability (Fig. 5). Because bistability in the phosphorylation of AMPA receptors has not been shown experimentally, this result provides an interesting testable prediction of bistability in the phosphorylation of AMPA receptors in cerebellar LTD. A recently established antiphosphorylated AMPA receptors antibody (Matsuda et al., 1999) should allow us to test this prediction in the future.
The role of each signaling molecule and pathway in the induction of the phosphorylation of AMPA receptor in the cerebellar LTD
We next explored the roles of signaling molecules and pathways in the phosphorylation of AMPA receptors (Fig. 5). Without PKC activation, the phosphorylation of AMPA receptors did not increase in the kinetic simulation (Fig. 5A, yellow line) because of the lack of activation of PKC (Fig. 5B). This finding is consistent with an earlier observation that the inhibition of PKC activity results in the complete disruption of LTD in mice expressing inhibitory peptide of PKC in PCs (De Zeeuw et al., 1998). In contrast, cerebellar LTD has been shown to be unimpaired in mice lacking the PKCγ gene, one of the isoforms of PKC (Chen et al., 1995). This discrepancy may be attributable to the possibility that PKC isoforms function redundantly or that there is a PKC-independent pathway. Various experiments suggest the involvement of PKC in the induction of LTD (Linden and Connor, 1991; Ito and Karachot, 1992). Taken together, the former possibility is more likely. Without NO production, no stable phosphorylation of AMPA receptors was observed (Fig. 5A,green line) because of the lack of activation of PKG (Fig. 5E) and that of inhibition of PP2A (Fig.5C), despite the fact that PKC was fully activated (Fig.5B). This finding is consistent with the observation that NO is essential for the induction of all phases of LTD, although NO itself shows no ability to elicit LTD (Lev-Ram et al., 1997). In addition, these results support the idea that LTD is dually regulated by the PKC and NO pathways. Furthermore, without Ca2+elevation, the stable phosphorylation of AMPA receptors did not occur with a lesser initial phase (Fig. 5A, pink line) because of the partial activation of PKC (Fig.5B). This finding is also consistent with the fact that Ca2+ is essential for the induction of all phases of LTD (Sakurai, 1990; Konnerth et al., 1992). Without DAG elevation, both the initial and intermediate phases of the AMPA receptor phosphorylation were observed, but the phosphorylation was less stable compared with that under the normal condition (Fig.5A, black line) because of the partial activation of MAP kinase (Fig. 5D). This finding indicates that DAG contributes to the stable phosphorylation of AMPA receptors in both the initial phase and the intermediate phase. Experimentally, it is difficult to analyze the role of DAG because no specific inhibitors of DAG are available. Moreover, the inactivation of PLC, which produces both DAG and IP3, results in the depletion of both products. Consequently, the simulation is useful for analyzing the role of signal transduction pathways, which is experimentally difficult to analyze. Without PLA2 activation, the intermediate phase of the AMPA receptor phosphorylation disappeared, with a slight reduction in the initial phase (Fig. 5A,cyan line). The positive feedback loop was responsible for the intermediate phase because without MAP kinase-mediated PLA2 activation, the intermediate phase disappeared without affecting the initial phase (Fig.5A, purple line) because of the lack of the stable activation of MAP kinase (Fig. 5D). Lack of the activation of either Raf or MEK resulted in the same results as that without MAP kinase-mediated PLA2 activation. This finding indicates that the positive feedback loop including PLA2 is responsible for the intermediate phase of the AMPA receptor phosphorylation. This result is consistent with an earlier observation that PLA2 regulates the intermediate phase of LTD (Linden, 1995). Without the activation of PLA2 by Ca2+elevation, the initial phase came to be slightly reduced, and the intermediate phase came to be less stable (Fig. 5A,brown line).
DISCUSSION
The initial phase of the AMPA receptor phosphorylation in cerebellar LTD was dependent on the direct activation of PKC by linear cascades including DAG, Ca2+, and AA, whereas the intermediate phase was mediated by the activation of the positive feedback loop. The inactivation of PLA2resulted in the disappearance of the intermediate phase without a remarkable change in the initial phase, indicating that the positive feedback loop was responsible for the intermediate phase. Consistent with this result, the selective inhibition of PLA2 has been reported to convert cerebellar LTD to short-term depression (STD), and the application of free unsaturated fatty acids has been shown to result in an apparent conversion from STD to LTD in cultured PCs by the stimulation, where STD, not LTD, comes to be normally induced (Linden, 1995). Taken together, it is likely that PLA2 and AA serve as regulators of the positive feedback loop and are responsible for the intermediate phase of LTD.
The lack of the activation of Raf, MEK, or MAP kinase led to the same result as that without the MAP kinase-mediated PLA2 activation. MAP kinase and MEK have also been shown to be required for the induction of both the initial and intermediate phases of cerebellar LTD (Kawasaki et al., 1999). The reason that MAP kinase and MEK are not involved in the induction of the initial phase in the simulation may be attributable to the possibility that an unknown part of the MAP kinase cascade is missing in the simulation. MAP kinase has been shown to be required for the mGlu receptor activity (Kawasaki et al., 1999). Therefore, the molecular linkage between MAP kinase and mGlu receptors needs to be clarified and incorporated into the kinetic simulation.
In all phases, the inhibition of PP2A by NO is essential for the phosphorylation of AMPA receptors. This is consistent with an earlier observation that NO is essential for cerebellar LTD, but NO itself is insufficient to induce LTD (Lev-Ram et al., 1997), suggesting that PP2A acts as a gate signal of cerebellar LTD. This result indicates that a significant amount of PF stimulation is required to induce cerebellar LTD via inhibition of PP2A, and that spontaneous PF activity may be insufficient to inhibit PP2A activity enough to be required for the stable phosphorylation of AMPA receptors. If only a small amount of PP2A is easily inhibited by spontaneous PF activity, the PF activity together with spontaneous CF activity may cause unexpected and unwarranted cerebellar LTD. Therefore, the reason such a large amount of PP2A is needed may be caused partially by the role of the NO–PP2A pathway as a gate signal for the induction of cerebellar LTD. In this study, spontaneous PF activity was not included. However, if spontaneous PF activity is included, the PP2A may reach to the some basal level, which may not be small enough to induce LTD, and should be inhibited further by NO signal, leading to induced LTD. Therefore, with or without spontaneous PF activity, the action of PP2A would be similar. In addition, PP2A has another role in the kinetic simulation; the dephosphorylation of Raf and MEK. Raf and MEK regulate the positive feedback loop, and the inhibition of the dephosphorylation of these molecules was required for the activation of the positive feedback loop. Therefore, PP2A also has a permissive role in the phosphorylation of AMPA receptors as well as in the activation of the positive feedback loop.
We compared the phosphorylation of AMPA receptors in the kinetic simulation and cerebellar LTD shown experimentally. In the initial phase, some results of the AMPA receptor phosphorylation in the kinetic simulation did not correlate with the experimental observations of cerebellar LTD. The stimulation of CF alone did not induce an apparent initial phase in the kinetic simulation (Fig. 3), whereas a similar protocol can induce an initial sharp peak of EPSC decrease (Daniel et al., 1992; Hemart et al., 1994). Additionally, without NO or Ca2+ elevation, a small peak in the phosphorylation of the AMPA receptors could still be observed in the kinetic simulation (Fig. 5A), whereas this initial peak was not observed by the addition of blockers of Ca2+ (Sakurai, 1990; Eilers et al., 1997) or NO (Lev-Ram et al., 1997). These discrepancies raise the possibility that other mechanisms underlie the expression of the initial phase of cerebellar LTD. It has been shown that increasing Ca2+ in dendrite activates Ca2+-dependent K+ channels, resulting in shunting PF-induced EPSP (Gruol et al., 1991; Khodakhah and Ogden, 1993; Muller et al., 1998). Additionally, it has been shown recently that the activation of postsynaptic mGluR1 in PC dendrites transiently depresses synaptic transmissions at PF–PC synapses by presynaptic mechanisms involving Ca2+ increase in PC dendrites and retrograde signaling (Levenes et al., 2000) and that retrograde inhibition of Ca2+ influx occurs through endogenous cannabinoids at excitatory synapses on PCs (Kreitzer and Regehr, 2001). These mechanisms are likely to be the main mechanisms for the expression of the initial phase of cerebellar LTD. The kinetic parameters in these two mechanisms, although unknown at present, should allow us to test whether these mechanisms can explain the expression of the initial phase of cerebellar LTD by use of the kinetic simulation. Accordingly, the kinetics simulation is a powerful tool for testing whether the phosphorylation of AMPA receptors can explain the expression of cerebellar LTD. Lists of the kinetic simulation results, the corresponding experimental results, and model predictions are summarized in Appendix .
The phosphorylation of AMPA receptors has been shown to trigger the internalization of phosphorylated AMPA receptors (Matsuda et al., 2000;Wang and Linden, 2000). A recent study has demonstrated that, in cortical neurons, the activation of AMPA receptors without the activation of NMDA receptors leads to the rapid and almost complete internalization of AMPA receptors within 5 min after the stimulation and the slow reinsertion of the AMPA receptors to the cell surface, resulting in a reduction in the number of AMPA receptors at the cell surface (Ehlers, 2000). It is possible that a similar mechanism is involved in cerebellar LTD. Taken together with our results, it is likely that the phosphorylation of AMPA receptors is a key step for cerebellar LTD expression through the internalization of the AMPA receptors at least in the intermediate phase. However, the internalization of phosphorylated AMPA receptors was not incorporated into our model because of the absence of kinetic parameters. The absence of phosphorylated AMPA receptor internalization in the kinetic simulation may affect the results by affecting the balance between the phosphorylated and nonphosphorylated AMPA receptor concentrations; however, it is likely that the results would be similar if the internalization step was incorporated into kinetic simulation because the internalization process itself does not affect the signaling pathways. If the kinetic parameters of the phosphorylated AMPA receptor internalization is determined, we can address the question of whether the phosphorylation of AMPA receptors and the subsequent internalization of the AMPA receptors can explain the intermediate phase as well as the initial phase of cerebellar LTD.
There are some discrepancies of the signaling pathways between the slice and the culture conditions. However, cerebellar LTD can be elicited robustly under both conditions. For example, under some experimental conditions (Linden et al., 1991, 1995), NO is not thought to be essential for the induction of cerebellar LTD. It is possible that the concentration or the activity of PP2A decreases, or that an unidentified molecule except NO regulates the PP2A pathways to compensate and induce LTD under such conditions. Additionally, no evidence of the roles of PLA2 and AA has been demonstrated in the slice condition. If this positive feedback loop does not work in the slice, then an alternative feedback loop such as the Rap1–MAP kinase-mediated positive feedback loop found in PC12 cells (York et al., 1998) is responsible for the intermediate phase. The fact that cerebellar LTD robustly occurs under both conditions strongly suggests that LTD is elicited by similar mechanisms in both conditions even if detailed mechanisms are different. If one pathway operates in only one of the conditions, changing the concentration of molecules of a certain type might compensate and induce the cerebellar LTD, or another redundant pathway having a similar characteristics might come to be a replacement. Therefore, despite the discrepancies between these conditions, the kinetic simulation model is still useful for understanding the mechanism of cerebellar LTD.
It has been shown that glial fibrillary acidic proteins (Shibuki et al., 1996), expressed in astrocytes but not in neurons, and the δ2 subunit of the glutamate receptor channels (Hirano et al., 1995;Kashiwabuchi et al., 1995) are essential for the induction of LTD. The phosphorylation of AMPA receptors has been shown to lead to the internalization of the AMPA receptors (Matsuda et al., 2000; Xia et al., 2000). Although the detailed mechanisms are still unknown, the molecules and mechanisms involved should be incorporated into the kinetics model in the future. Local Ca2+release within the dendritic spines of PCs has been shown recently to be required for LTD induction (Miyata et al., 2000). Therefore, the specific localization of molecules and accessibility should also be quantitatively determined and incorporated into the kinetics model. In addition to phosphorylation, Ca2+-dependent K+ channels (Gruol et al., 1991; Khodakhah and Ogden, 1993; Muller et al., 1998), presynaptic retrograde signaling (Levenes et al., 2000), and retrograde inhibition of presynaptic Ca2+ influx by endogenous cannabinoids (Kreitzer and Regehr, 2001) have been shown to underlie the expression of cerebellar LTD. Electrical processes, such as the channel activity of AMPA receptors and Ca2+-dependent K+ channels, coupled with biochemical processes should also be incorporated into the kinetic simulation, provided the apparent kinetic parameters come to be determined. Moreover, the role of Raf in the induction of LTD has yet to be shown experimentally. Therefore, the present kinetic simulation should not be regarded as a definitive model, but as a complementary method for exploring cerebellar LTD in addition to experimental methods. It should also be emphasized that, even if the simulation can reproduce some experimental results, this does not exclude the possibility that unknown pathways or molecules are additionally needed for the induction of cerebellar LTD. In any case, experiments together with the approach using kinetic simulation should greatly improve our understanding of the behaviors of the complex biochemical reactions underlying cerebellar LTD.
The inputs used in this study
The temporal waveforms of inputs such as Ca2+, NO and glutamate used in this study are shown (Fig. 6). The concentration of CRF is assumed to be constantly 0.1 μm during the stimulation because no data were available. In the case of the stimulation of PF or CF alone, Ca2+ peak concentrations are reduced to 2.3 and 2.6%, respectively, according to an earlier report (Wang et al., 2000).

The inputs used in this study. The temporal waveforms of inputs, such as Ca2+(A), NO (B), and glutamate (C), which are induced by a single stimulation of PF and CF, are shown.
Summary of the kinetic simulation results and the corresponding experimental results together with the predictions
The phosphorylation of AMPA receptors and the changes of EPSC (Chen and Thompson, 1995) induced by the conjunctive stimulation of PF and CF (Fig. 2)
The initial and the intermediate phases are reproduced by the kinetic simulation. The late phase is not reproduced because the late phase of cerebellar LTD has been shown to require gene expression and protein synthesis (Linden, 1996), and our kinetic simulation does not include both of them. The time courses of the activation of PKC and PP2A have yet to be determined. However, antibodies against the phosphorylated AMPA receptors (Matsuda et al., 2000) and the phosphorylated G-substrate, an inhibitor of PP2A, should allow us to measure these time courses in the future.
The conjunctive stimulation of PF and CF induces the stable phosphorylation of AMPA receptors in cerebellar LTD (Fig. 3)
In the kinetic simulation, neither the initial nor intermediate phase is significantly induced by the stimulation of either PF or CF alone. However, an experimental stimulation similar to the CF alone stimulation in the kinetic simulation has been shown to induce an initial sharp decrease in the EPSC (Daniel et al., 1992; Hemart et al., 1994). The failure of the kinetic simulation to reproduce this observation raises the possibility that other mechanisms in addition to the phosphorylation of AMPA receptors underlie the initial phase of cerebellar LTD. It has been shown that increasing Ca2+ in dendrite activates Ca2+-dependent K+ channels, resulting in shunting PF-induced EPSP (Gruol et al., 1991; Khodakhah and Ogden, 1993; Muller et al., 1998). Additionally, it has been shown recently that the activation of postsynaptic mGluR1 in PC dendrites transiently depresses synaptic transmissions at PF–PC synapses by a presynaptic mechanism involving Ca2+ increases in PC dendrites and retrograde signaling (Levenes et al., 2000) and that retrograde inhibition of Ca2+ influx occurs through endogenous cannabinoids at excitatory synapse on PCs (Kreitzer and Regehr, 2001). These mechanisms are likely to be the main mechanisms for the expression of the initial phase of cerebellar LTD.
Bistability of the phosphorylation of AMPA receptors in cerebellar LTD (Fig. 4)
In the kinetic simulation, the phosphorylation of AMPA receptors by PKC shows bistability. Because bistability in the phosphorylation of AMPA receptors has not been shown experimentally, this result provides an interesting testable prediction of bistability in the phosphorylation of AMPA receptors in cerebellar LTD. A recently established antiphosphorylated AMPA receptors antibody (Matsuda et al., 1999) should allow us to test this prediction in the future.
The role of each signaling molecule and pathway in the induction of the phosphorylation of AMPA receptor in the cerebellar LTD (Fig. 5)
In the kinetic simulation, the roles of PKC, NO, Ca2+, and PLA2 are basically consistent with the experimental observations (Sakurai, 1990;Konnerth et al., 1992; Lev-Ram et al., 1997; De Zeeuw et al., 1998). However, without NO or Ca2+ elevation, a small peak in the phosphorylation of the AMPA receptors can still be observed in the kinetic simulation, whereas this initial peak is not observed by the addition of blockers of Ca2+ (Sakurai, 1990; Eilers et al., 1997) or NO (Lev-Ram et al., 1997). As indicated above, these discrepancies raise the possibility that other mechanisms underlie the expression of the initial phase of cerebellar LTD.
In addition, DAG contributes to the stable phosphorylation of AMPA receptors in both the initial and intermediate phases in the kinetic simulation. Experimentally, it is difficult to analyze the role of DAG because no specific inhibitors of DAG are available. Moreover, the inactivation of PLC, which produces both DAG and IP3, results in the depletion of both products. Consequently, the simulation is useful for analyzing the role of signal transduction pathways, which is experimentally difficult to analyze.
Footnotes
S.K. was supported by the Inheritance and Variation Group, PRESTO (Japan Science and Technology Corporation, Japan). We thank G. Ferriol for his help in the kinetic simulation, M. Ito, T. Hirano, H. Qadota, K. Kaibuchi, and K. Doya for critically reading this manuscript, and K. Kurihara and N. Matsumoto for their technical assistance.
Correspondence should be addressed to Shinya Kuroda, Kawato Dynamic Brain Project, ERATO, Japan Science and Technology, 2-2-2 Hikaridai, Seika-cho, Souraku-gun, Kyoto 619-0288. E-mail:kshinya{at}erato.atr.co.jp.
N. Schweighofer's present address: Learning Curve, 2F Fuji Bldg 40, 15-14 Sakuragaoka-cho, Shibuya-ku, Tokyo 150-0031.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}