

Abstract
The mechanisms establishing and maintaining dendritic spines in the mammalian CNS remain primarily unknown. We report a novel mechanism of neuronal spine plasticity in the developing preoptic area in which estradiol induces prostaglandin-E2(PGE2) synthesis that in turn increases the density of spine-like processes. Estradiol requires PGE2 synthesis,in vivo and in vitro, and upregulates the dendritic spine protein spinophilin, an effect attenuated by antagonism of the AMPA–kainate receptor. This signaling pathway may involve cross talk between neighboring neurons and astrocytes and appears specific to the preoptic area in that hippocampal neurons responded with an increase in spinophilin to estradiol but not PGE2. Regionally specific mechanisms of estradiol-mediated synaptic plasticity allow for epigenetic control of complex sex-typic behaviors.
A primary locus of excitatory synaptic transmission in the mammalian CNS is the dendritic spine (Colonnier, 1968; Larkman, 1991; Sorra and Harris, 2000). These protrusions from dendritic shafts exhibit dynamic changes in number, size, and shape in response to variation in hormonal status, developmental stage, and changes in afferent input (Fifkova, 1985; Munoz-Cueto et al., 1991; Woolley and McEwen, 1992; Moser et al., 1994; Murphy and Segal, 1996; McEwen and Alves, 1999). Understanding the mechanisms determining the formation and plasticity of dendritic spines remains a major goal of neuroscience.
Estradiol (E2) has proven a potent regulator of dendritic spines and synapses in several discrete loci within the developing and adult brain. Exposure to E2 during a critical developmental period orchestrates permanent sexually dimorphic synaptic patterning in regions of the hypothalamus (Matsumoto and Arai, 1980, 1986; Matsumoto et al., 1988; Perez et al., 1990; Pozzo Miller and Aoki, 1991), whereas postpubertal E2exposure controls dynamic changes in spine density in the hippocampus (Woolley et al., 1990; Woolley and McEwen, 1992; Woolley, 1998) and hypothalamus (Calizo and Flanagan-Cato, 2000). Elucidating the cellular and molecular mechanisms underlying E2 regulation of dendritic spine density provides a valuable model for establishing a relationship between modifications in synaptic patterning and complex behaviors.
Estradiol increases the level of prostaglandin-E2(PGE2) in such diverse tissues as kidney and brain (Katayama and Lee, 1985; Ma et al., 1997). Elevated PGE2 is positively correlated with inflammatory pain responses (Ito et al., 2001), induction of neuronal apoptosis after ischemic injury (Takadera et al., 2002), and increased tumorigenic and metastatic potential in estrogen-dependent breast cancers (Karmali et al., 1983; Kundu et al., 2001). The synthesis of prostanoids begins with the oxygenative cyclization of arachidonic acid by cyclooxygenase. The inducible isoform of cyclooxygenase, cyclooxygenase-2 (COX-2), is an immediate early gene enriched within dendritic spines and postulated to play a role in synaptic plasticity (Kaufmann et al., 1997). Cyclooxygenase-2 expression is strongly regulated during normal postnatal development in the rat, following well known histiogenetic gradients and coinciding with the critical period for activity-dependent synaptic remodeling (Kaufmann et al., 1996). Of the prostanoids synthesized by COX-2, PGE2 is one of the most biologically active of the family and has long been recognized as a diffusible factor capable of regulating several second messenger pathways in a variety of tissues (Villani et al., 1974; Luczak et al., 1979; Ruwe and Myers, 1979).
Astrocytes are important in synapse formation and efficacy (Pfrieger and Barres, 1997; Ullian et al., 2001), including that of E2-induced dendritic spine synapses (Mong et al., 1999, 2001). Recent and emerging evidence implicates PGE2 as a mediator of dynamic cell-to-cell communication involving cross talk between astrocytes and neurons (Ma et al., 1997; Rage et al., 1997; Bezzi et al., 1998; Sanzgiri et al., 1999). Astrocyte-to-neuron signaling regulates the number of dendritic spines available for synaptogenesis in regions of the developing brain (Baloyannis and Kim, 1979; Meshul and Seil, 1988; Seil et al., 1992;Seil, 1997). Astrocytes release glutamate in response to PGE2 (Nicol et al., 1992; Bezzi et al., 1998), which can activate the glutamate receptors of neighboring neurons and modulate their dendritic spine density (McKinney et al., 1999a,b; Luthi et al., 2001). A relationship between PGE2 and neuronal morphology, however, has not previously been demonstrated. We report here a novel role for PGE2 in synaptic plasticity, the induction of dendritic spines in response to the gonadal steroid E2. This effect is region specific in that it is observed in the preoptic area (POA) but not the hippocampus, and it is dependent on the activation of AMPA–kainate receptors by glutamate that may originate from astrocytes.
MATERIALS AND METHODS
Animals. Female Sprague Dawley rats (Charles River Laboratories, Wilmington, MA) were mated in our animal facility, and pregnancy was confirmed by vaginal smear. Pregnant females were isolated and allowed to deliver normally. Animals were maintained on a reverse 12 hr light/dark cycle and provided ad libitumaccess to food and water. Cages were checked regularly for the presence of pups to determine the day of birth [postnatal day 0 (P0)], and litters that were found the morning of delivery were used for experiments. Female pups were used for experiments involving hormonal manipulation to avoid the endogenous hormonal exposure that occurs in males.
Microdissection and culture conditions. Under sterile conditions, brains were removed within 1–2 hr after birth, placed in aZivic Miller brain mold, and sectioned at 1 mm. The POA and rostral hippocampal formation (including the rostral portions of the differentiating CA1 and CA3 fields as well as the dentate gyrus) were microdissected using the guidelines illustrated in Figure1, A and B. Briefly, the optic chiasm appears near the rostral portion of the diencephalon and was used as a landmark to guide the rostral-to-caudal dissection within the brain mold. The POA was then microdissected from the 1-mm-thick section using the perimeter of the anterior commissure for both the dorsal and lateral incisions (Fig. 1B). Another 1 mm slice was taken immediately posterior to the section containing the POA. This segment contained the rostral portion of the hippocampal formation, which was microdissected using the lateral ventricle to delineate the lateral limits, the corpus callosum, the dorsal limit, and the dorsal portion of the third ventricle the ventral limit (Fig. 1A). Tissue chunks were placed in HBSS+ [HBSS, HEPES buffer, antibiotic/antimycotic (10,000 U of penicillin G sodium, 10,000 μg of streptomycin sulfate, and 25 μg of Amphotericin B)] (Invitrogen, Grand Island, NY) and 0.25% trypsin. After 15 min at 37°C, 1 ml of 10% dextran-treated charcoal-stripped fetal bovine serum (Invitrogen) and 0.5 ml of antibiotic/antimycotic (FBS+) was added to deactivate the trypsin. The solution was dissociated by mechanical triturating and centrifuged at 1000 rpm for 5 min, the supernatant was discarded, and the pellet was resuspended in phenol red-free, sterile culture medium (SCM) [DMEM-F12 (Invitrogen) and FBS+]. Density was established, and cells were seeded onto poly-l-lysine (MW 70,000; Sigma, St. Louis, MO)-coated glass coverslips (number 1, 25 mm diameter; Fisher Scientific, Pittsburgh, PA) at a density of 150,000. Plates were maintained in an incubator at 37°C and 5% CO2for 4 d in vitro (DIV; time of plating, DIV0). Cells were allowed to acclimate for 24 hr before first treatment and were treated on DIV1 and DIV2 and collected in lysis buffer on DIV3 for Western analysis, ∼24 hr after the second treatment. Treatments were added to existing culture medium, and depending on the specific experiment, cells were exposed to the following either individually or in combination: (1) sesame oil vehicle, (2) 0.5 nm PGE2 in 0.9% saline, (3) 0.5 nm PGF2α in 0.9% saline, (4) 5 nm estradiol benzoate (E2) in 0.001% DMSO in sesame oil, (5) 1 μm indomethacin in sesame oil, (6) 10 μml-glutamic acid in 0.9% saline, (7) 40 μm (+)-MK801 in 0.9% saline, (8) 100 μmdl-AP-5 in 0.9% saline, and (9) 40 μm DNQX in 0.9% saline (n = 6/group for all culture experiments). Doses of the PGE2 and estradiol were based on published literature (Whalen and Nadler, 1963; Morimoto et al., 1991; Mouihate and Lestage, 1995; Caggiano and Kraig, 1998; Southall and Vasko, 2000) as well as our own experience (Mong et al., 1999). Doses of estradiol were 5 nmin vitro and 100 μgin vivo. This seemingly large in vivo dose was used to overwhelm α-fetoprotein, an endogenous steroid-binding globulin found in the fetal circulation. This protein binds estradiol, preventing the hormone from crossing the blood–brain barrier and from entering cells, thereby protecting the female fetuses from being masculinized by estradiol originating in the dam. Levels of α-fetoprotein reach a maximum during the same perinatal period that testosterone surges and remain at significant levels in the bloodstream of pups for several days after birth (Montano et al., 1995).

Microdissection of the preoptic area and the rostral hippocampal formation. A, Sagittal drawing of the developing rat brain demonstrating the areas collected (shaded regions) and the landmarks used to guide the microdissection, including the anterior commissure (ac) and the optic chiasm (ox) for the preoptic area (POA), and the corpus callosum and the dorsal portion of the third ventricle (d3v) for the hippocampal formation (hf). B,Coronal drawing of the neonatal brain at the level of the POA.Top left, Three-dimensional reconstruction of the neonatal brain at the level of the section (taken from Altman and Bayer, 1995). Top right, Sagittal view of the neonatal brain displaying the rostral to caudal coordinates used for collecting the POA. Shaded region represents the area collected for microdissections that were guided by the anterior commissure and the optic chiasm. cc, Corpus callosum;BST, bed nucleus of the stria terminalis;3v, third ventricle. Grid scale, 0.5 mm.
Immunocytochemistry: microtubule-associated protein-2 and spinophilin immunoreactivity. Coverslips with adhered cells were immersed in 4% paraformaldehyde and 5% sucrose in 0.1m PBS, pH 7.5, warmed to 37°C for 10 min, washed three times in PBS, and permeabilized with 50% ethanol for 60 min at 4°C. The cells were then washed three times in PBS and incubated in 10% normal goat serum in 0.4% Triton X-100 PBS (PBST) for 60 min at room temperature with agitation. After blocking, either a mouse monoclonal anti-microtubule-associated protein-2 (MAP-2) primary antibody (Sigma) (1:3000) or a rabbit polyclonal anti-spinophilin/neurabin II primary antibody (Upstate Biotechnology, Lake Placid, NY) (1:8000) was applied in 10% normal goat serum PBST, and the cultures were incubated overnight at 4°C. The coverslips were washed three times in PBST and incubated in biotinylated secondary antibody (1:500; goat anti-mouse or goat anti-rabbit; Vector Laboratories, Burlingame, CA) for 60 min at room temperature with agitation, followed by three washes in PBST. The cultures were then incubated with an avidin–biotin horseradish-peroxidase complex (Vectastain ABC, Elite Kit; Vector Laboratories) for 60 min at room temperature, washed twice with PBST, and visualized with 0.05% 3,3′-diaminobenzidine tertrahydrochloride (Polysciences, Warrington, PA) and 0.005% H2O2. After visualization, the coverslips were mounted on 2% gelatin-coated glass slides.
In vivo hormonal manipulations and comparison of spinophilin levels between sexes. Female pups received two hormonal treatments 24 hr apart with the first within 6 hr of birth. Animals were assigned to one of six treatment regiments including: (1) subcutaneous injections of 0.1 cc sesame oil, (2) subcutaneous injections of 100 μg of E2 in 0.1 cc sesame oil, (3) subcutaneous injections of 25 μg of indomethacin and 100 μg of E2 in 0.1 cc sesame oil, (4) intracerebroventricular injections of 2 μg of PGE2 in 0.1 cc of 0.9% saline and subcutaneous injections of 0.1 cc oil, (5) intracerebroventricular injections of 2 μg of PGE2 in 0.1 cc of 0.9% saline and subcutaneous injections of 25 μg indomethacin in 0.1 cc sesame oil, or (6) intracerebroventricular injections of 0.9% saline and subcutaneous injections of 0.1 cc sesame oil (n = 4/group). Intracerebroventricular injections were performed by hand on cold-anesthetized pups. Under bright light, the cranial landmark Bregma is visible through the skin and was used to locate the lateral ventricles. Injections were performed by penetrating the skin and skull with a 23 gauge 1 cc Hamilton syringe stereotaxically lowered to a predetermined depth. Infusion volume was 0.1 cc and extended for a 60 sec period. Brains were collected on P2, ∼24 hr after the second treatment. To determine the levels of spinophilin within the POA on the day of birth, POA tissue was collected within 2 hr of birth from male (n = 5) and female (n = 5) littermates and immediately flash frozen in isopentane and stored at −70°C.
Western immunoblots. For cultured cells, coverslips were rinsed with 250 μl of TBS and then immersed in 75 μl of lysis buffer, pH 7.5, consisting of 0.8% Tris-HCl, 0.9% NaCl, 1% Tergitol NP-40 (Sigma), 1 mm phenylmethylsulfonyl fluoride, and peptidases including leupeptin, aprotinin, and pepstatin (1 μg/ml) for 5 min at 4°C. The coverslips were grazed with a cell scraper, and the resulting homogenate was collected. For analysis ofin vivo brain tissue, the POA and rostral hippocampal formation were microdissected as described above, flash frozen in isopentane, and stored at −70°C until being homogenized in lysis buffer as described above with 0.5% Triton X-100 (Sigma) substituting for NP-40. The homogenate was then subjected to a Bradford protein assay to determine and standardize protein levels in subsequent Western analyses. Protein (10 μg) was electrophoresed in separate lanes on an 8–16% precast SDS-polyacrylamide gel (Novex, San Diego, CA) and transferred to a polyvinyl difluoride membrane (Bio-Rad, Hercules, CA). Membranes were blocked in 5% nonfat milk in 0.1% Tween TBS (M-TTBS) for 1 hr at room temperature and then incubated in anti-spinophilin/neurabin II rabbit polyclonal IgG (Upstate Biotechnology) at 1 μg/ml in M-TTBS. A 30 min incubation of a goat anti-rabbit HRP (New England BioLabs, Beverly, MA) conjugated IgG at a 1:10,000 dilution followed. The Phototope chemiluminescence system (New England BioLabs) was used for detection of the protein recognized by the antisera. The blots were exposed on Hyperfilm-ECL (Amersham, Arlington Heights, IL) for varying exposure times (30–60 sec). The protein was detected as a band of relative molecular mass of 120 kDa, and the integrative grayscale pixel area-density (iad) was captured with a CCD camera and quantified using NIH Image software.
Reverse transcription-PCR. Brains were removed and placed in a Zivic Miller brain mold and immediately sectioned at 1 mm. The POA was microdissected as described above, flash frozen in isopentane, and stored at −70°C until being homogenized. Tissue from the kidney was also collected to verify the detection of EP4. Tissue was used for isolation of total RNA using an RNeasy Mini Kit (Qiagen, Valencia, CA). RNA concentration was determined by spectrophotometery, and 1 μg of RNA was reverse translated to DNA and then amplified using the one step protocol for Ready-To-Go RT-PCR beads (Amersham Pharmacia Biotech, Buckinghamshire, UK). DEPC-treated water and 20 pmol of each primer was added to the RNA and RT-PCR bead at 4°C. This solution was transferred to the thermal cycler and cycled 35 times at annealing temperatures appropriate for each primer pair (55–62°C). All oligonucleotides used were synthesized by Invitrogen (Rockville, MD) with standard purity. Subsequent PCR product was electrophoresed on a 2% agarose gel in Tris-acetate, and DNA product was visualized with chelation of 0.5% ethidium bromide.
Gas chromatography-mass spectrometry. Female pups received two treatments 24 hr apart with the first within 6 hr of birth. Females were assigned to one of two regimes: two subcutaneous injections of 0.1 cc sesame oil (n = 5) or two subcutaneous injections of 100 μg of E2 in 0.1 cc sesame oil (n = 6). The POA was collected and immediately prepared for mass spectrophotometery 12 hr after the second treatment. Briefly, each sample, consisting of ∼30 mg of tissue, was rinsed with HBSS (containing 138 nm NaCl, 5 mm KCl, 4 mmNaHCO3, 5.6 mmd-glucose, 0.3 mmNaHPO4, and 0.3 mmKH2PO4; Invitrogen) and manually homogenized in a glass-microhomogenizer with 75 μl of HBSS containing 1 mm CaCl2. An additional 75 μl of HBSS was added, and the solution was sonicated for 20 sec. Any tissue debris was removed by centrifugation at 12,000 rpm for 15 sec. Ten microliters of the supernatant was used for Bradford protein assay. Fifty microliters of denatured prostanoid standards and 250 μl of acetone were added, and the solution was vortexed and centrifuged for 5 min. The supernatant was divided equally and dried under a steady stream of nitrogen gas. Once completely dry, 25 μl of 2% 0-methoxylamine HCl in pyridine was added to each sample. The pyridine solvent evaporated under a nitrogen stream, and the residue in each vial was treated with reagents to synthesize the pentafluorobenzyl ester-trimethylsilyl ether derivatives of the prostanoids for gas chromatography–mass spectrometric (GC–MS) analysis at the Asthma and Allergy Center of Johns Hopkins University School of Medicine (Baltimore, MD). Levels of prostanoids were subsequently normalized to the amount of protein in each sample.
Data collection and analyses. Analysis of MAP-2 immunocytochemical cellular morphology was performed under a Nikon 100× oil-immersion objective using the Neurolucida system (MicroBrightField Inc., Colchester, VT). Ten neurons from six coverslips of each treatment (n = 6/group) were examined. Criteria for cell selection included the ability of the experimenter to discern individual neurons stained evenly from soma to distal processes. Neurons entangled with one another were therefore excluded. Neurons were chosen from areas of even cell survival, thus neurons growing within dense or sparse pockets were also excluded. The experimenter was blind to the treatment group throughout analysis. The following morphological features were measured: somal area (in square micrometers), neurite length (in micrometers), neurite number, primary and secondary neurite branching, number of spine-like processes (per neurite), and density of spine-like processes (spine number per micrometer), with a spine defined as any protrusion <5 μm. Analyses of the spine-like processes incorporated the complete neurite from soma to distal end and included all processes extending from each neuron examined. Typically a 1:2 ratio of neurons to astrocytes was observed in these 3-d-old cultures. Western blots were compared within each gel, with the exception of the in vitro spinophilin experiments where results were across several immunoblots with an nvalue of two or four for each group within each blot. For these, bands were quantified as a ratio of iad with the iad of the DMSO controls of the same gel serving as the comparison group. This standardization allowed for the analysis of multiple groups (each with ann = 6) across multiple films. For the comparison between sexes on the day of birth, males were compared with females on the same gel and represented in arbitrary units. All results except those of the GC–MS and sex dimorphism experiments were analyzed using a one-way ANOVA followed by the Student–Neumann–Keulspost hoc multiple comparison test to determine significance between groups. Results of the GC–MS were analyzed using Student'st test between treatment groups, as were those of the sex dimorphism experiment, data from which were expressed in arbitrary units. All statistical tests used α <0.05 as the criterion for significance.
GenBank accession numbers. The accession numbers for the sequences of the previously identified prostaglandin-E2 receptor subtypes are as follows: EP1 (D88751); EP2 (U94708); EP3 (NM-012704); and EP4 (U94709).
RESULTS
Estradiol or PGE2 increase spine-like processes in cultured POA neurons
To explore the possible role of E2 in mediating changes in the morphology of neurons, POA tissue from newborn females was dispersed and plated. Cells acclimated for 24 hr and were then exposed to physiologic levels of E2 (5 nm) for 48 hr before postfixing and immunocytochemical detection of neuron-specific MAP-2. Cell morphology was assessed at the light level using the Neurolucida image analysis system. Estradiol-treated cultures showed an almost threefold increase in the mean number per neurite (F(3,12) = 21.8; p < 0.001), and in a separate experiment, density (F(5,24) = 26.2;p < 0.001) of spine-like processes, with a spine defined as any neurite protrusion <5 μm in length (Fig.2A–D).

E2 or PGE2 increase spine-like processes of preoptic area neurons in vitro. A, Representative photomicrograph of cultured neurons treated with 0.5 nm PGE2 on DIV1 and DIV2 and visualized on DIV3 after immunocytochemical detection of MAP-2. Numerous aspects of neuronal morphology were assessed, but the only changes observed were in the number and density of spine-like processes, defined as any protrusion <5 μm in length. Scale bar, 25 μm. B, Representative photomicrographs of the neurites of cultured neurons treated with vehicle, 5 nmE2, or 5 nm E2coadministered with 1 μm of the cyclooxygenase inhibitor indomethacin, and visualized after MAP-2 immunocytochemistry. Scale bar, 5 μm. C, Treatment with E2 (5 nm) or PGE2 (0.5 nm) for 48 hr resulted in a significant increase in the number of spine-like processes on neurites. PGF2α (0.5 nm) had no effect compared with vehicle-treated controls (10 neurons from each coverslip, n = 6 per group, ANOVA; *p < 0.001 compared with PGF2α and vehicle). D, Similar effects of E2 and PGE2 were seen on mean spine density in a separate experiment. The combination of E2 and PGE2 was not different from either one alone. The induction of spine-like processes by E2 was blocked by simultaneous administration of indomethacin (1 μm) but had no effect on the actions of exogenous PGE2 (10 neurons from each coverslip,n = 6 per group, ANOVA; *p < 0.001 compared with E2 + Indo and vehicle).
Spines were visualized by immunocytochemistry for MAP-2 using diaminobenzidine as the chromagen. Microtubule proteins, such as MAP-2, are localized to the dendritic shaft of hippocampal neurons and are associated with the relative stability of this structure as compared with the plasticity of dendritic spines (Kaech et al., 2001). Our ability to visualize spine-like processes on cultured POA neurons with MAP-2 immunocytochemistry may be a result of intracellular spread of the reaction product or may be functionally related to a process of spine induction different from that seen in hippocampus. Moreover, the plasticity of the dendritic architecture may vary between the two regions.
We next tested the hypothesis that PGE2 is a mediator of E2-induced spine induction. Treatment with PGE2 (0.5 nm) for 48 hr precisely mimicked the effect of E2, causing a significant increase in both spine number per neurite (p < 0.001) and density (p < 0.001). PGF2α, a biologically active eicosanoid converted from the same immediate precursor as PGE2, had no effect on spine number. Simultaneous administration of the cyclooxygenase inhibitor, indomethacin (1 μm), blocked the induction of spine-like processes by E2 but had no effect on the actions of exogenous PGE2(p < 0.001 from vehicle and E2 and indomethacin). The combination of E2 and PGE2 was not significantly different from either one alone. There were also no significant differences between E2-treated and control cultures in somal size, neurite length, neurite number, or neurite branching. Similarly, the effects of PGE2treatment were limited solely to changes in spines, with no other effects on neuronal morphology observed (data not shown).
Estradiol or PGE2 increase spinophilin expression in cultured POA neurons
Spinophilin, a protein highly enriched within dendritic spines, has been implicated in regulating spine formation and function (Allen et al., 1997; Feng et al., 2000). We therefore used it as a protein marker of spines (Fig. 3A,B), with the assumption that increases in spinophilin expression would result in proportional increases in spine density. POA cell cultures were treated as above, the cells collected and protein levels analyzed by Western blot. Consistent with our morphological analysis, both E2 or PGE2 induced a threefold increase in spinophilin expression (F(6,35) = 66.9; p < 0.001). Indomethacin again blocked the effect of E2 but did not alter the effect of exogenous PGE2. As with spine-like processes, the combination of E2 and PGE2did not increase spinophilin expression above that seen with either treatment alone (Fig. 3C,D). Although there was a striking similarity in the magnitude and direction of treatment effects on spinophilin levels detected by Western blot and spine-like processes detected by MAP-2 immunocytochemistry, a direct relationship between these two variables has not been established.
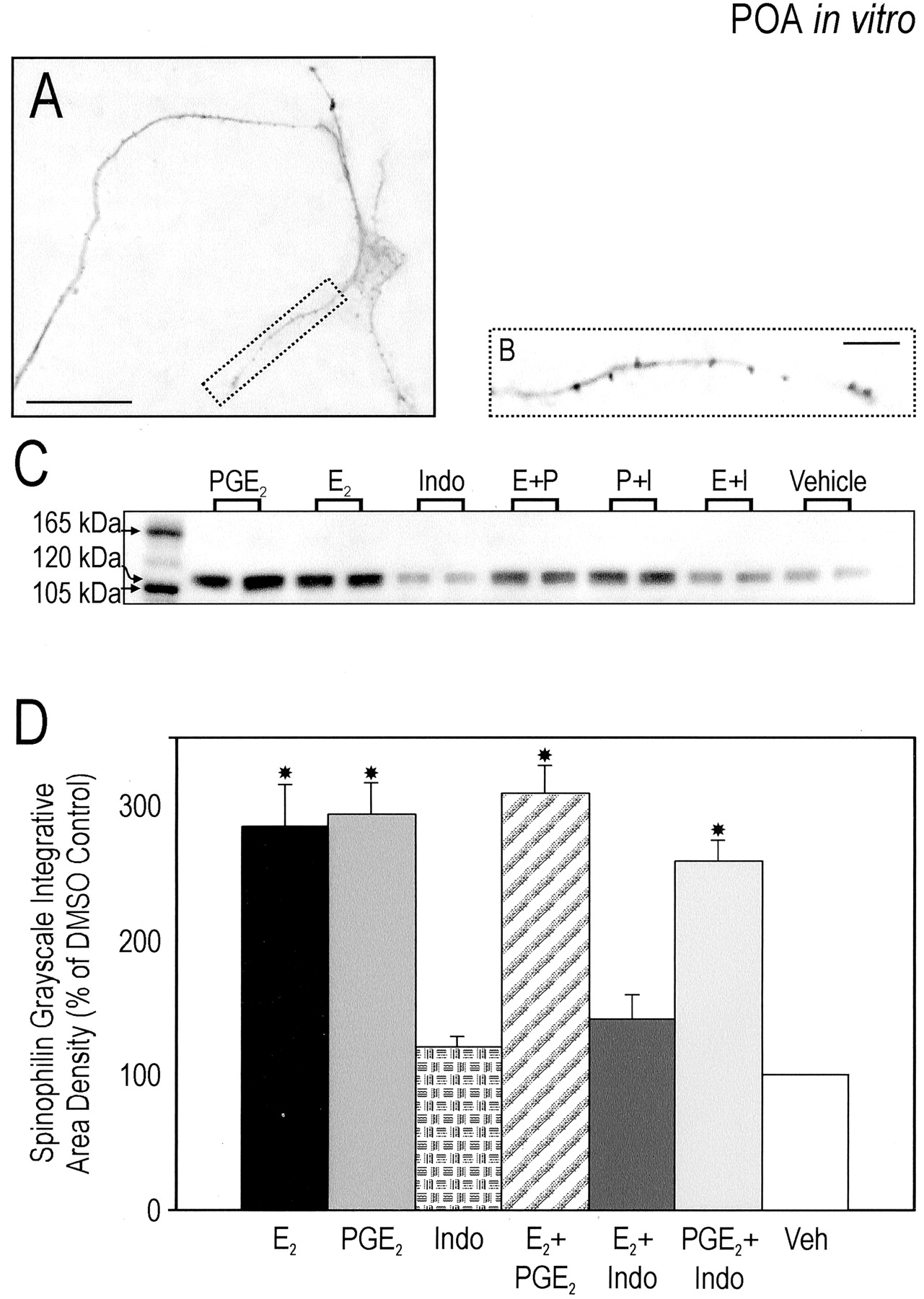
E2 or PGE2 increase expression of spinophilin protein in the preoptic area in vitro. A, Representative photomicrograph of the neurites of cultured neurons treated with 5 nmE2 and visualized after spinophilin immunocytochemistry. Scale bar, 25 μm. B, Enlargement of process encapsulated in A. Each punctum corresponds to one locus of concentrated protein. Scale bar, 5 μm. C,Representative Western immunoblot from treated cultures loaded onto each lane 3 d after plating. Cultures were exposed to the following either individually or in combination for 48 hr: (1) 0.5 nm PGE2, (2) 5 nmE2, (3) 1 μm indomethacin, or (4) vehicle. The blot was probed with an anti-spinophilin/neurabin II rabbit polyclonal antibody that recognized an appropriate band at 120 kDa. D, Analysis of Western immunoblots of cultured newborn preoptic area neurons treated for 48 hr in vitro. Data represent a ratio of iad with the iad of the vehicle controls of the same gel serving as the comparison group (n = 6 per group). This standardization allowed for the analysis of seven groups across three gels. Treatment with E2 or PGE2 resulted in a significant increase in expression of spinophilin. Coadministration of both did not augment effects seen when applied individually. Indomethacin blocked the estradiol-induced effect on spinophilin expression and had no effect alone (ANOVA; *p < 0.001 compared with Indo, E2 + Indo, or vehicle).
Antagonism of the AMPA–kainate receptor reduces both E2- and PGE2-induced spinophilin expression
Localization of ionotropic glutamate receptors to dendritic compartments allows for selective regulation of spine formation by glutamate released from axonal growth cones, presynaptic terminals, and neighboring astrocytes (Mattson, 1988; Goldin et al., 2001). Expression of glutamate receptors peaks during a developmental time window within which synaptogenesis occurs (Tremblay et al., 1988; Herrmann, 1996;Zhang et al., 1996). We therefore tested the hypothesis that ionotropic glutamate receptor activation participates in the estradiol-mediated increases in spinophilin. Effects of glutamate (10 μm) exposure paralleled those of E2 and PGE2, demonstrating a threefold increase in spinophilin expression (F(3,18) = 113.8; p < 0.001) in POA cell cultures (Fig.4A,B). The selective AMPA–kainate antagonist DNQX (40 μm) attenuated estradiol-mediated increases in spinophilin expression by 56.6% and PGE2-mediated increases by 66.2% (F(4,23) = 106.5; p < 0.001), establishing the role of glutamate subsequent to that of E2 and the prostanoid (Fig. 4C). Both the noncompetitive NMDA antagonist (+)-MK801 (40 μm) and the selective NMDA antagonistdl-AP-5 (100 μm) had no effect on induction of spinophilin by estradiol. Application of the any of the antagonists alone had no effect on baseline spinophilin expression (data not shown), further supporting an active role for E2 in the induction of PGE2synthesis and subsequent glutamate release.

Antagonism of the AMPA–kainate receptor attenuates E2- or PGE2-induced spinophilin expression. A, Representative Western immunoblot from treated POA cultures 3 d after plating. Cultures were exposed to the following either individually or in combination for 48 hr: (1) 0.5 nm PGE2, (2) 5 nmE2, (3) 10 μm glutamate, (4) 100 μm AP-5, (5) 40 μm MK801, (6) 40 μm DNQX, or (7) vehicle. The blot was probed with an anti-spinophilin/neurabin II rabbit polyclonal antibody that recognized an appropriate band at 120 kDa. B, C, Analysis of Western immunoblots of cultured newborn preoptic area neurons treated for 48 hr in vitro. Data represent a ratio of iad with the iad of the vehicle controls of the same gel serving as the comparison group (n = 6 per group). Treatment with glutamate resulted in an increase in the expression of spinophilin comparable with that induced by either E2 or PGE2. Coadministration of the AMPA–kainate antagonist DNQX significantly reduced both the E2 and PGE2-induced increase in expression, whereas coapplication of NMDA receptor antagonists had no effect on expression (ANOVA; *p < 0.001 compared with vehicle; **p < 0.001 compared with E2 + DNQX, PGE2 + DNQX, and vehicle; ***p < 0.001 compared with E2 + AP-5, E2 + MK801, and vehicle).
Prostaglandin receptor mRNA is expressed in the developing POA
For prostaglandins to promote dendritic spine formation early in development, the cognate receptors must be expressed perinatally. RT-PCR was used to identify the population of prostaglandin receptors (EP) expressed in the POA on the day of birth and amplified the mRNA of three of the four subtypes, EP1, EP2, and EP3 (Fig.5A). EP4 mRNA was not detected in the POA, but was amplified in kidney as a positive control.
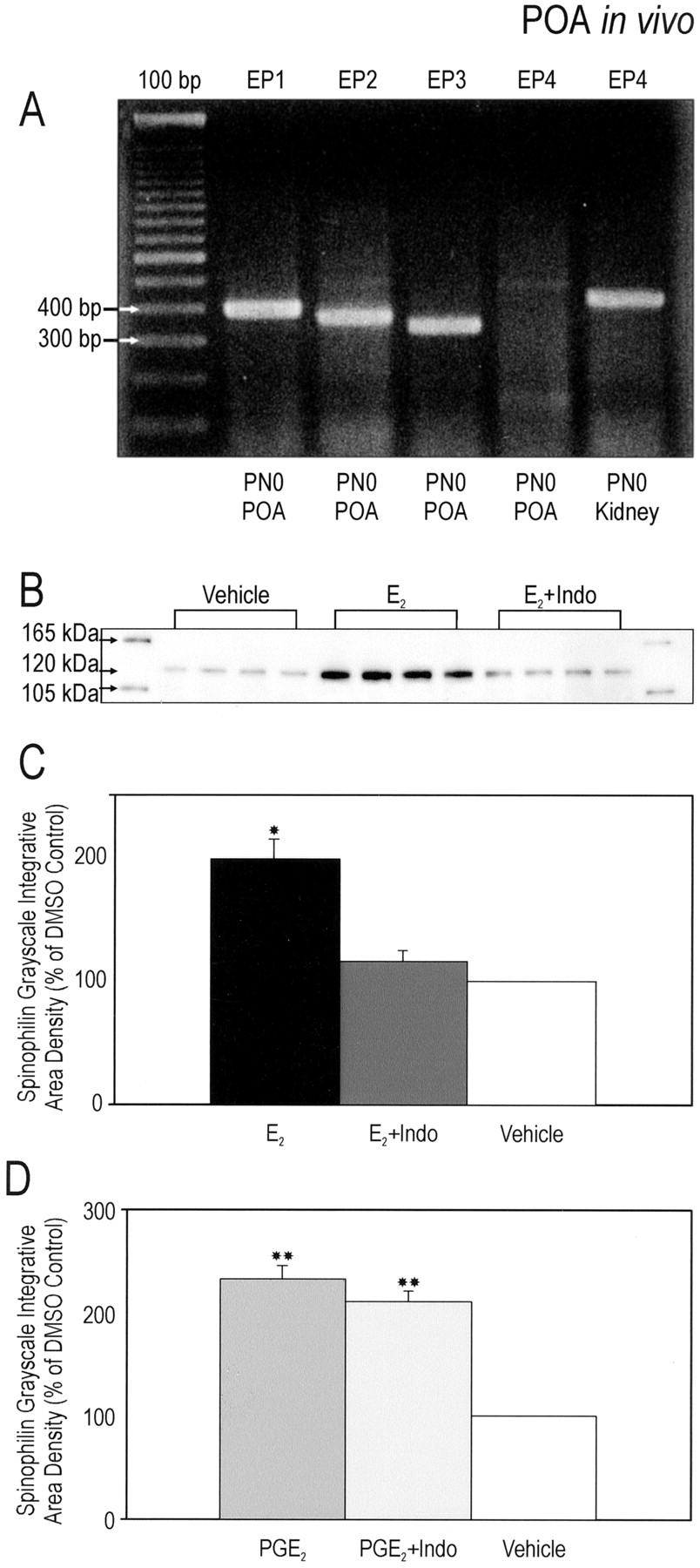
The in vitro effects of E2 and PGE2 on spinophilin expression are maintained in vivo. A, The mRNA for three of the four PGE2 receptor (EP1–4) subtypes are detectable within the POA as early as 4 hr post-utero. Tissue from rat kidney was used as a positive control for EP4. PN0, Postnatal day 0.B, Representative Western immunoblot of tissue from the POA collected from animals treated for 48 hr beginning on P0. Animals were exposed to the following either individually or in combination: (1) 0.1 cc sesame oil (subcutaneously), (2) 100 μg of E2in 0.1 cc sesame oil (subcutaneously), (3) 25 μg of indomethacin in 0.1 cc sesame oil (subcutaneously), (4) 2 μg of PGE2 in 0.1 cc in 0.9% saline (intracerebroventricularly), or (5) 0.1 cc of 0.9% saline (intracerebroventricularly). The blot was probed with an anti-spinophilin/neurabin II rabbit polyclonal antibody that recognized an appropriate band at 120 kDa. C, D, Quantification of Western immunoblots of preoptic area tissue treated for 48 hr in vivo. Each analysis represents one immunoblot quantified as a ratio of iad to that of controls of the same gel (n = 4 per group). Treatment with E2 or PGE2 resulted in a significant increase in levels of spinophilin. Indomethacin blocked the estradiol-induced effect on spinophilin, whereas its coapplication with PGE2 had no effect (ANOVAs; *p < 0.001 compared with E2 + Indo and vehicle; **p < 0.001 compared with vehicle).
Estradiol or PGE2 increase spinophilin expressionin vivo in the POA
We next asked whether PGE2 is required for the induction of dendritic spines by E2 in the POA in vivo. Newborn female rat pups received subcutaneous injections of E2 (100 μg) or intracerebroventricular infusions of PGE2 (2 μg). Forty-eight hours later, the brains were collected, and the POA was rapidly dissected and flash frozen until assay by Western blot. The expression of spinophilin was significantly increased after E2 (F(2,9) = 23.5; p = 0.001) (Fig. 5B,C) and PGE2 (F(2,9) = 77.6; p < 0.001) (Fig. 5D) in the developing POA as compared with the appropriate controls. In a separate group of animals receiving coadministration of indomethacin (25 μg, s.c.) with E2 or PGE2, the E2-induced increase in spinophilin expression was fully blocked (p < 0.001) (Fig.5C,D), with no change in the effect of exogenous PGE2.
Estradiol increases PGE2 levels in the POA
Coapplication of E2 and PGE2 showed neither an additive nor a synergistic response, further suggesting that these compounds are inducing spine induction through a serial rather than parallel process (although the possibility of a ceiling effect of each compound cannot be ruled out). Coadministration of the prostaglandin synthesis inhibitor indomethacin blocked the effect of E2, implying that PGE2 acts downstream of E2. We therefore tested the hypothesis that E2increases PGE2, while also investigating whether E2 promotes the synthesis of the other metabolites of arachidonic acid. We used GC–MS analysis of POA tissue collected from pups 36 hr after being treated immediately postpartum with either E2 or vehicle. The homogenate was evaluated for differences in the arachidonic acid metabolites PGD2, PGE2, PGF2α, and the two nonenzymatic byproducts of prostacyclin (PGI2), and thromboxane-A2 (TXA2), 6-keto-PGF1α and TXB2, respectively. Of the lipids analyzed, only PGE2showed a significant difference (p < 0.002) with a threefold increase from ∼70 pg/mg of protein to ∼280 pg/mg of protein after exposure to E2 (Fig.6). Two of the five control animals had PGE2 levels so low as to be undetectable above background and were scored as zero. Whereas PGD2, PGF2α, and 6-keto-PGF1αlevels were significantly greater than that of PGE2, no differences were found between treatment groups. TXB2 levels were undetectable above background (data not shown) in both groups.

E2 increases PGE2 levels in the developing preoptic area. Quantitative GC–MS analysis was used to measure the levels of various prostanoids and their metabolites in the preoptic area of newborn female rats treated with E2 or vehicle for 36 hr. A, PGE2 levels were increased threefold in rats treated with 100 μg of E2 in 0.1 cc sesame oil (subcutaneously) (t test, *p < 0.002). B, There was no change in PGD2, PGF2α, or 6-keto-PGF1α. TXB2 was undetectable above background. C, The pathway of arachidonic acid conversion to the prostanoids via cyclooxygenase suggests the effects of E2 may be acting directly at PGE synthase or indirectly via COX-2.
Estradiol, but not PGE2, increases spinophilin in the rostral hippocampal formation
Dendritic spine density is markedly increased on the pyramidal neurons of the adult hippocampus after E2 exposure (Woolley et al., 1990; Woolley and McEwen, 1992; Woolley, 1998), but this potential has not been investigated in immature hippocampal neurons. We therefore explored the possible involvement of both E2 and PGE2 in the induction of spines in developing hippocampus. The rostral portion of the perinatal hippocampus was plated and allowed to acclimate for 24 hr before exposure to either E2 or PGE2 for 48 hr. As compared with vehicle-treated cells, E2 but not PGE2 induced a significant increase in spinophilin expression (F(2,9) = 58.5;p < 0.001) (Fig.7A,B). E2 also induced a significant increase in spinophilin expression in vivo as compared with control tissue (F(2,9) = 66.9;p < 0.001) (Fig. 7C). Unlike our observations in the POA, however, coadministration of indomethacin with E2 did not inhibit this increase. Furthermore, neither PGE2 alone nor PGE2coapplied with indomethacin had any effect on spinophilin expression in hippocampal neurons (Fig. 7D).

Different mechanisms appear involved in the regulation of spinophilin protein in hippocampal tissue.A, Representative Western immunoblot of tissue from the POA collected from animals treated for 48 hr beginning on P0. Animals were exposed to the following either individually or in combination: (1) 0.1 cc sesame oil (subcutaneously), (2) 100 μg of E2in 0.1 cc sesame oil (subcutaneously), (3) 25 μg of indomethacin in 0.1 cc sesame oil (subcutaneously), (4) 2 μg of PGE2 in 0.1 cc in 0.9% saline (intracerebroventricularly), or (5) 0.1 cc of 0.9% saline (intracerebroventricularly). The blot was probed with an anti-spinophilin/neurabin II rabbit polyclonal antibody that recognized an appropriate band at 120 kDa. B, Analysis of Western immunoblots from cultured newborn rostral hippocampal tissue treated for 48 hr in vitro. The analysis represents one immunoblot quantified as a ratio of iad to that of controls of the same gel (n = 4 per group). Treatment with E2 (5 nm) but not PGE2 (0.5 nm) resulted in a significant increase in expression of spinophilin (ANOVA; *p < 0.001 compared with PGE2 or vehicle). C, D, Quantification of Western immunoblots from animals treated 48 hr in vivoas described in A. Each analysis represents one immunoblot quantified as a ratio of iad to that of controls of the same gel (n = 4 per group). Treatment with E2alone or in combination with indomethacin resulted in a significant increase in expression of spinophilin. PGE2 alone or in combination with indomethacin had no effect on spinophilin expression (ANOVA; **p < 0.001 compared with vehicle).
Males exhibit greater spinophilin protein levels on the day of birth in the POA
To investigate the possibility of a sexually dimorphic expression of spinophilin in the developing POA, tissue was collected within 2 hr of birth from male and female littermates and analyzed by Western blot. Males had two and a half times the level of spinophilin protein in the POA as that of females (t test, p < 0.001) (Fig. 8). The magnitude of this sex difference is consistent with the effects on spinophilin levels induced by estradiol and PGE2 treatment both in vivo and in vitro and suggests that the naturally occurring sex difference in spinophilin is the result of higher endogenous estradiol in the newborn male POA.

Sexually dimorphic expression of spinophilin on the day of birth in the POA. A, Western immunoblot of preoptic area tissue collected from males and females on the day of birth. The blot was probed with an anti-spinophilin/neurabin II rabbit polyclonal antibody that recognized an appropriate band at 120 kDa.B, Quantification of the above Western immunoblot of male and female preoptic area tissue. We determined iad for each band in arbitrary units, and males were compared to females on the same gel (n = 5/group). Males displayed two and a half times the level of spinophilin protein in the POA as compared with that of females (t test, ∗p < 0.001). The magnitude of difference is consistent with the effects on spinophilin levels induced by estradiol or PGE2 treatment bothin vivo and in vitro.
DISCUSSION
We report here a novel mechanism for induction of dendritic spines completely dependent on E2-induced increases in the prostanoid, PGE2, and subsequent glutamatergic activation of AMPA–kainate receptors. Cultured POA neurons exposed to either E2 or PGE2 exhibited an almost threefold rise in the number and density of spine-like processes. The effects of E2 were blocked by the prostanoid synthesis inhibitor, indomethacin, and the combination of E2 and PGE2 was equally effective as either one alone. The related but distinct prostaglandin, PGF2α, had no effect on spine-like processes of cultured preoptic neurons. Using spinophilin as a marker for spinesin vivo, we found parallel responses to those seen in vitro. Although a ceiling effect cannot be ruled out, coapplication of E2 and PGE2 showed neither an additive nor a synergistic response, suggesting that these compounds are acting through a serial rather than parallel process. Quantitative GC–MS analysis demonstrated a threefold increase in the levels of PGE2 within the POA of neonates exposed to E2 as compared with vehicle-treated controls. There was no change in the levels of the other prostanoids, suggesting the effect of E2 is specific to PGE2. A comparison of spinophilin between the sexes found neonate males to express 2.5-fold greater levels than that of neonate females in the perinatal POA, a difference similar in magnitude to that induced by estrogen treatment of females. Glutamatergic activation of AMPA–kainate receptors significantly increased spinophilin expression, and antagonism of these receptors significantly reduced both the E2 and PGE2-induced increases in protein expression. Finally, the ability of E2 to induce spinophilin was also observed in hippocampal neurons, however these effects appear to be mediated through a mechanism not involving PGE2.
In rodents, the newborn brain is essentially bipotential, equally capable of becoming masculinized or feminized. Resolution of the ultimate phenotype is a function of the hormonal milieu established by the gonads. In males, the neonatal testes secrete a copious quantity of testosterone during a restricted perinatal developmental window. This testosterone gains access to the brain where it is converted locally to E2 by neuronal aromatase (Naftolin et al., 1971;Ryan et al., 1972; MacLusky and Naftolin, 1981). As a result, local levels of E2 in the developing hypothalamus can be as high as 5 ng/gm brain tissue (Rhoda et al., 1984). Estradiol induces permanent effects on the developing neuronal substrate, most notably the establishment of sexually dimorphic synaptic patterning. The frequency of dendritic spine synapses can be twice as high in some regions of the male brain compared with that of the female, and yet be the exact opposite in an adjacent region (Matsumoto and Arai, 1980,1986; Pozzo Miller and Aoki, 1991; Mong et al., 1999, 2001). Despite the fundamental significance of the establishment of sexually dimorphic synaptic profiles, the mechanism or mechanisms by which E2 achieves this end has remained virtually unknown.
The POA, a subdivision of the hypothalamus, is centrally involved in the control of sex-typic reproductive behavior, including sexual and maternal behaviors (Meisel and Sachs, 1994; Numan, 1994). Sex differences in morphometry of this brain region include volumetric variance in particular nuclei (Bloch and Gorski, 1988) and synaptic patterning (Raisman and Field, 1973; Larriva-Sahd, 1991). Estrogen receptors are highly expressed by preoptic neurons throughout life (Fuxe et al., 1987; DonCarlos and Handa, 1994; DonCarlos, 1996), and synaptic patterns of the adult brain are established by the neonatal hormonal profile (Raisman and Field, 1973), although additional plasticity is found in the female at puberty (Anderson, 1982). Although we have demonstrated a role for prostaglandins in establishing the neuroarchitecture of the developing brain, it is clear the adult POA remains responsive to PGE2 and may involve a recapitulation of developmental processes. Sexually unresponsive males can be induced to copulate after POA infusions of PGE2 (Clemens and Gladue, 1977), and prostaglandins within the POA are critically involved in control of body temperature (Adair, 1977; Gordon and Heath, 1980).
A critical but limiting variable in studies of synaptic patterning is the ability to quantify synapses. Traditional and reliable methods include Golgi impregnation, which allows for analysis of overall cell morphology but not the presence of synapses, and electron microscopy, which allows for the latter but not the former. Although highly informative, both of these techniques are labor intensive and can become prohibitive when multiple manipulations are compared. Spinophilin was used here as an indirect marker for spines. Spinophilin localizes to the head and neck region of dendritic spines and interacts with several proteins, including actin (Stephens and Banting, 2000). Actin is central to the formation, maintenance, morphology, and activity of spines (Fifkova, 1985; Matus, 2000; Kaech et al., 2001), and spinophilin bundles actin filaments, suggesting a role as one of the organizers of the actin-based cytoskeleton in dendritic spines (Nakanishi et al., 1997; Feng et al., 2000; Stephens and Banting, 2000). Such properties make spinophilin not only a marker of dendritic spine density, but also suggest it may be a mediator of dynamic changes in spine morphology. Others have also observed a comparable relationship between the magnitude of synapse-related proteins and changes in spinophilin (Brake et al., 2001), suggesting that this biomarker is a valuable tool for assaying changes in synaptic profiling under a myriad of conditions. In the hippocampus we detected a 30% increase in spinophilin expression in response to E2, a result consistent in both scale and direction with previous studies investigating the effects of E2 on hippocampal spines and spine synapses (Woolley et al., 1990; Woolley and McEwen, 1992; Woolley, 1998). Although extremely powerful, using spinophilin as a marker of dendritic spines does have its limitations. Analyses of protein levels cannot discern if a treatment selectively promotes the construction of some spines versus the dismantling of others. Furthermore, assays for spinophilin do not reveal information regarding the status of spine function or activity. Nonetheless, the striking correlation we observed in the effects of treatment on spine-like processes and spinophilin levels measured by Western blot both in vivo and in vitro suggest this protein is a reliable predictor of spine density.
A link between glutamate receptor activation, which initiates most changes in synaptic strength, and subsequent modifications of dendritic density has been difficult to establish in the vertebrate CNS. Combined electrophysiologic and morphometric analyses have demonstrated the necessity of AMPA receptor activation in the maintenance and structure of postsynaptic dendritic spines (McKinney et al., 1999a,b). NMDA receptor activation, on the other hand, appears critical to synaptogenesis and the pruning of dendritic spines (Luthi et al., 2001). Although our investigations do not explore synaptogenesis per se, they do demonstrate that AMPA–kainate receptor activation is critical and sufficient to increase a dendritic protein, spinophilin, within the developing POA in response to estradiol exposure. Of great interest is the finding that AMPA—kainate-specific antagonists alone had no effect on baseline spinophilin expression, suggesting that in the POA, glutamatergic activation of these receptors are not involved in maintenance, but rather creation of dendritic spines.
A central question raised by the current findings is whether E2-induced increases in PGE2 are manifest in POA neurons, astrocytes, or a combination of the two. Although it is clear that neurons of the POA express significant concentrations of estrogen receptor throughout life (Fuxe et al., 1987; Yuri and Kawata, 1991; DonCarlos, 1996), there is ambiguity regarding the potential for the same receptors in astrocytes of the POA (Langub and Watson, 1992; Ojeda and Ma, 1999). Similarly, there is uncertainty regarding whether COX-2 is exclusively neuronal or also present in astrocytes under restricted conditions (Kaufmann et al., 1996; O'Banion et al., 1996). Given that the preponderance of evidence suggests neuronal localization for both the estrogen receptor and COX-2, we have developed a working model in which E2 increases PGE2 in neurons which then diffuses to act on neighboring astrocytes. Evidence suggests that neighboring astrocytes respond to PGE2 by releasing glutamate (Nicol et al., 1992;Bezzi et al., 1998), which in our model back-communicates to trigger neuronal AMPA–kainate receptors, activation of which has been implicated in dendritic spine modification (McKinney et al., 1999a; Goldin et al., 2001; Luthi et al., 2001). Several components of this model remain to be tested.
Our data suggest a regionally specific effect of E2 in the POA involving the synthesis of PGE2 because this mechanism was not evident in the developing hippocampus. Interestingly, outside of the cortex, the densest collection of COX-2-expressing neurons is found in the POA, with only a few in the CA1 field of the hippocampus (Breder et al., 1995). Regional differences in either the receptors or synthetic enzymes for PGE2 may underlie the specificity of the effects of PGE2. The localization of COX-2 to dendritic spines and its rapid induction with synaptic activity previously led to the hypothesis that this immediate early gene is a modulator of neuronal plasticity (Kaufmann et al., 1997). The observation that pharmacological inhibition of COX-2 does not result in disturbances in learning and memory seemed to contradict this view (Kaufmann et al., 1997). Our findings may shed light on this discrepancy. Estradiol-induced increases in hippocampal dendritic spine density have been correlated with enhanced NMDA-mediated currents (Woolley and McEwen, 1994; Woolley et al., 1997) and exaggerated LTP (Warren et al., 1995; Cordoba Montoya and Carrer, 1997; Gupta et al., 2001), a cellular correlate of learning and memory. Performance on various hippocampal-dependent spatial tasks are enhanced by estradiol pretreatment, and the increase in dendritic spines induced by estradiol has been directly correlated with enhanced performance (Sandstrom and Williams, 2001). In contrast, E2 induction of dendritic spines in the POA is likely related to sex differences in reproductive behavior and would be predicted to have no effect on learning and memory. This divergence in the mechanism of E2-mediated neuronal plasticity demonstrates how epigenetic control of behavior is achieved through regionally specific signal transduction pathways.
Footnotes
This work was supported by a predoctoral National Research Service Award (MH12862) to S.K.A. and a grant from the National Institutes of Mental Health (MH52716) to M.M.M. We thank J. J. Alt for his critical and conscientious technical assistance with culture preparation as well as A. Z. Murphy, S. M. Thompson, A. Keller, and D. Weinreich for their constructive comments on this manuscript.
Correspondence should be addressed to Stuart K. Amateau, Program in Neuroscience, University of Maryland at Baltimore, School of Medicine, 655 West Baltimore Street, BRB 5020, Baltimore, MD 21201. E-mail:samat001{at}umaryland.edu.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}