Article Text

Abstract
Aims: To study the regulation of type 1 insulin like growth factor receptor (IGF-1R) tyrosine kinase activity using the fission yeast Schizosaccharomyces pombe and a green fluorescent protein (GFP) tagged, full length IGF-1R.
Methods: The β chain of the IGF-1R (βwt) was expressed under inducible conditions in the fission yeast S pombe. Western blot analysis with antiphosphotyrosine antibodies was used to assess the kinase activity of βwt. A GFP tagged IGF-1R (GFP–IGF-1R) was constructed to study the tyrosine kinase activity of the full length IGF-1R. The signalling capabilities of GFP–IGF-1R in response to IGF-1 stimulation were investigated in transiently transfected fibroblasts. Immunofluorescent staining for cellular phosphotyrosine content was used to assess the localisation and tyrosine kinase activity of GFP–IGF-1R.
Results: The βwt protein displayed functional tyrosine kinase activity in S pombe and phosphorylated endogenous yeast proteins. In response to IGF-1 stimulation, the GFP–IGF-1R became autophosphorylated and also activated the phosphatidylinositol 3-kinase and mitogen activated protein kinase pathways. Tyrosine phosphorylation and kinase activity of the GFP–IGF-1R could be visualised by immunofluorescence with antiphosphotyrosine antibodies. Coexpression of a mammalian tyrosine phosphatase PTP1B with βwt completely inhibited this tyrosine kinase activity in yeast and also reduced the tyrosine phosphorylation in COS cells transfected with the GFP–IGF-1R.
Conclusions: Schizosaccharomyces pombe can be used to analyse the tyrosine kinase activity of the IGF-1R β chain and its regulation by tyrosine phosphatases. In addition, the regulation of IGF-1R tyrosine kinase activity can be studied using a GFP tagged IGF-1R. Using both of these methods, IGF-1R kinase activity was shown to be inhibited by the protein tyrosine phosphatase, PTP1B.
- βwt, β chain of the type 1 insulin-like growth factor receptor
- GFP, green fluorescent protein
- IGF-1R, type 1 insulin-like growth factor receptor
- PI3K, phosphatidylinositol 3-kinase
- IR, insulin receptor
- MAPK, mitogen activated protein kinase
- PAGE, polyacrylamide gel electrophoresis
- PDGF, platelet derived growth factor
- PTP, protein tyrosine phosphatase
- SDS, sodium dodecyl sulphate
Statistics from Altmetric.com
- βwt, β chain of the type 1 insulin-like growth factor receptor
- GFP, green fluorescent protein
- IGF-1R, type 1 insulin-like growth factor receptor
- PI3K, phosphatidylinositol 3-kinase
- IR, insulin receptor
- MAPK, mitogen activated protein kinase
- PAGE, polyacrylamide gel electrophoresis
- PDGF, platelet derived growth factor
- PTP, protein tyrosine phosphatase
- SDS, sodium dodecyl sulphate
The type 1 insulin-like growth factor receptor (IGF-1R) is a member of the family of transmembrane, tyrosine kinase receptors that includes the insulin receptor (IR) and the orphan insulin receptor related receptor. It is expressed in a variety of cell types and is activated upon binding of the secreted ligands, IGF-1 or IGF-2, triggering activation of the intracellular tyrosine kinase domain and autophosphorylation of the receptor. The physiological responses to IGF-1R tyrosine kinase activation are diverse and include differentiation, proliferation, protection from apoptosis, and transformation (for reviews see O'Connor1). The pathways leading from the receptor that mediate these effects have been studied extensively and at least partly elucidated in recent years.2 However, very little is known about the regulation of IGF-1R activity at the receptor level. Most notably, no conclusive evidence exists for the role of a particular protein tyrosine phosphatase (PTP) in the regulation of IGF-1R tyrosine kinase activity, despite the fact that the balance between tyrosine phosphorylation and dephosphorylation is a crucial control mechanism in the regulation of tyrosine kinase signalling.3 In contrast, the identification of PTPs involved in insulin signalling has been the focus of extensive research because these molecules could provide potential therapeutic targets for the treatment of type 2 diabetes. Although in vitro studies and the use of substrate trapping mutants have implicated several PTPs involved in insulin signalling,4 the most compelling evidence for the role of a particular PTP comes from studies in transgenic mice with a targeted disruption for the PTP1B gene.5 These mice display increased sensitivity to insulin and prolonged insulin receptor phosphorylation compared with their wild-type littermates. The authors speculate that this is probably the result of a direct interaction between PTP1B and the IR. However, the demonstration of a direct interaction between PTPs and their targets in vivo is difficult because of the number of tyrosine phosphorylation events taking place at any one time in the cell and the crosstalk and compensation that occurs between various tyrosine kinase signalling pathways.
“The physiological responses to IGF-1R tyrosine kinase activation are diverse and include differentiation, proliferation, protection from apoptosis, and transformation”
The fission yeast Schizosaccharomyces pombe is an excellent model organism in which to study mammalian signalling systems because of the high degree of homology between this yeast and mammalian cells. In particular, studies in this organism have contributed greatly towards our understanding of the complexities of the cell cycle (for recent review see Nurse6). Despite this homology, and in contrast to Saccharomyces cerevisiae, S pombe lacks detectable tyrosine kinase activity and it is this fact that makes it particularly amenable to studying the activity and modulation of mammalian tyrosine kinases.7 Schizosaccharomyces pombe has been used to study the activity and regulation of the cytoplasmic tyrosine kinases Src and c-Abl,7 and in the identification of PTP2C as a negative regulator of platelet derived growth factor (PDGF) receptor signalling.8
Another tool that has recently been shown to be applicable to the study of receptor tyrosine kinases is the reporter molecule green fluorescent protein (GFP). Originally purified from the jellyfish Aequorea victoria, GFP is now widely used in the functional analysis and intracellular localisation of a variety of proteins.9 GFP chimaeras are created by fusing the cDNA for the 26 kDa GFP to either the C-terminal or N-terminal coding sequence of the target protein. Proteins tagged with GFP exhibit green fluorescence when excited with blue light, and cells expressing these proteins can be unambiguously identified by fluorescence light microscopy. The epidermal growth factor receptor (EGFR) was the first tyrosine kinase receptor to be tagged with GFP, and studies with this chimaera have helped elucidate the dynamics of EGFR trafficking10 and receptor tyrosine kinase activity.11 Biochemical analysis of the EGFR–GFP chimaera revealed little difference in signalling between this protein and the wild-type receptor, despite the C-terminal addition of this 26 kDa protein.10
We set out to determine whether S pombe could be used to study the tyrosine kinase activity of the IGF-1R and be used as a tool for the identification of IGF-1R regulatory tyrosine phosphatases. This involved the initial characterisation of the tyrosine kinase activity of the IGF-1R β chain (βwt) when expressed under inducible conditions in S pombe. To extend the findings from these studies to mammalian cells, we turned to immunofluoresence methods and constructed a GFP tagged IGF-1R to study IGF-1R regulation in the context of the full length receptor. We then investigated whether βwt activity could be modulated by coexpression with the mammalian tyrosine phosphatase, PTP1B. Overall, we show that the tyrosine kinase activity of the IGF-1R β chain can be studied in S pombe, and that this activity can be inhibited by PTP1B. We describe the use of a GFP tagged IGF-1R to study IGF-1R function, and use this to demonstrate that PTP1B can also inhibit the tyrosine kinase activity of the full length IGF-1R in mammalian cells.
MATERIALS AND METHODS
Subcloning and GFP–IGF-1R construction
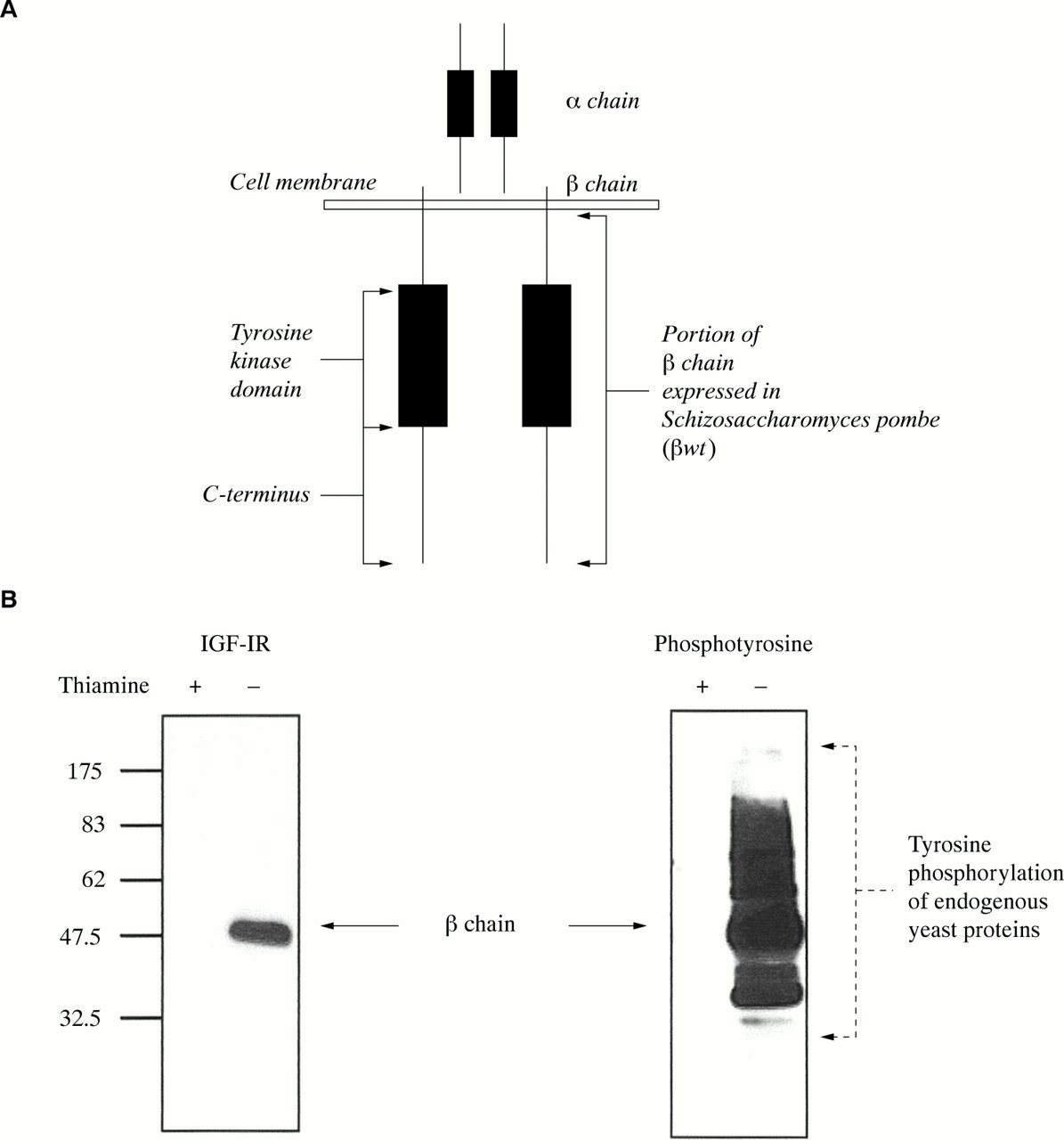
The IGF-1R β chain (residues 931–1337; βwt) was subcloned from pKS via XhoI/BamHI into the S pombe expression vector pRSP,7 which contains the inducible nmt1 promoter.12 The full length cDNA of PTP1B was excised from pKS by SalI/XbaI digestion and ligated into the constitutive expression vector, pADH, which has been described previously.7 For mammalian transfection studies, PTP1B was liberated from pADH and ligated into pIRES (Clontech Laboratories, Palo Alto, California, USA) after EcoRI digestion. For expression of full length IGF-1R (wild-type), pCDNA3–IGF-1R was used.13 GFP was fused to the C-terminus of the full length protein coding region of the IGF-1R (fig 1A) as follows: an approximately 1 kb region (corresponding to residues 1020–1337) was amplified by the polymerase chain reaction using the following primers, HinK (ctcaacgaagcttctgtgatg) and Nostop (acggtccaaggatccgcaggtcgaagactgg). Primer HinK flanks the endogenous HindIII site (in bold) whereas primer Nostop flanks the endogenous BamHI site (in bold) and also incorporates a mutated stop codon. The amplicon was digested with HindIII and BamHI and cloned into HindIII/BamHI digested full length IGF-1R (pKS–IGF-1R). The mutated IGF-1R was then released from pKS upon digestion with SalI and BamHI, blunt ended by incubating with Taq polymerase for 30 minutes at 72°C, then cloned into the pT-Adv vector (Clontech). Digestion with EcoRI released the full length mutated IGF-1R from pT-Adv, which was then ligated into EcoRI digested pEGFP (Clontech) to yield pIGF-1R–GFP.

(A) Illustration of the heterodimeric type 1 insulin-like growth factor receptor (IGF-1R) indicating subdomains. The cytoplasmic region of the IGF-1R β chain (βwt; residues 931–1337), which contains the tyrosine kinase domain, was expressed inducibly in Schizosaccharomyces pombe. The C-terminus of the full length IGF-1R was tagged with green fluorescent protein (GFP) to generate GFP–IGF-1R. (B) βwt expression and tyrosine kinase activity in S pombe. Schizosaccharomyces pombe cells transformed with pRSP βwt were induced to express βwt by removal of thiamine from the media (thiamine −), or left uninduced (thiamine +). After 24 hours, cultures were harvested, lysates were prepared, and 10 μg of protein was separated by 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis. Western blot analysis was performed with anti-IGF-1Rβ and antiphosphotyrosine antibodies. The βwt protein is indicated at 47 kDa in the induced culture (thiamine −). A range of antiphosphotyrosine reactive bands in the right panel indicates tyrosine phosphorylation of endogenous yeast proteins
Schizosaccharomyces pombe expression system
Schizosaccharomyces pombe strain SP200 (h−s leu1–32 ura4 ade6–210) was used in these studies and growth conditions were according to Moreno et al.14 Transformation was performed using either the lithium acetate method15 or by electroporation.16 Transformants were selected based on their ability to grow on selective media, leu− and ura− for pRSP and pADH, respectively. To induce the expression of βwt, transformants were seeded in selective medium containing thiamine (5 μg/ml), grown to mid-log phase, then washed five times in thiamine free medium, followed by resuspension in medium with or without thiamine. Cultures were maintained in log phase for 24 hours after induction, at which time cultures were harvested by centrifugation for western blot analysis.
Cell culture and transfection
R− cells are 3T3-like fibroblasts derived from mouse embryos with a targeted disruption of IGF-1R genes and have been described previously.17 COS7 cells are monkey kidney cells transformed with the SV40 T-antigen. All cell lines were maintained in Dulbecco's modified Eagle's medium (DMEM; Biowhittaker UK, Berkshire, UK) containing 10% fetal calf serum, glutamine, and antibiotics. Transient transfections were performed using a liposomal based method (LipofectAMINE; Life Technologies, Paisley, Scotland). To compare the activity of non-GFP tagged versus GFP tagged IGF-1R, R− cells were transfected in a 10 cm plate with 8.0 μg of either pIGF-1R–GFP or pCDNA3–IGF-1R. After 24 hours, each plate was split into two wells on a six well plate and 48 hours after transfection the cells were serum starved for four hours and then stimulated with IGF-1 (100 ng/ml) for 15 minutes, or left unstimulated. Cells were washed once in ice cold phosphate buffered saline and then lysed for western blot analysis. For immunofluoresence studies, COS7 cells were plated in a 24 well plate and transiently transfected with 0.5 μg of either pIGF-1R–GFP + pIRES, pEGFP + pIRES–PTP1B, or pIGF-1R–GFP + pIRES–PTP1B. Forty eight hours after transfection cells were stimulated with IGF-1 as before.
Preparation of lysates and western blotting
Cell pellets from S pombe induction experiments were washed once in ice cold Tris buffered saline containing the protease inhibitors, PMSF (1mM), pepstatin (1μM), and aprotinin (1.5 μg/ml) and the tyrosine phosphatase inhibitor Na3VO4 (1mM). Cells were then resuspended in an appropriate volume of modified RIPA buffer (10mM Na phosphate buffer, pH 7.0, 150mM NaCl, 50mM NaF, 0.1% sodium dodecyl sulphate (SDS), 1% Triton X-100, 0.5% deoxycholate) plus the above inhibitors. Lysis was achieved by the addition of glass beads (425–600 μm; Sigma-Aldrich Ireland Ltd, Wexford, Ireland) and rigorous agitation at 4°C for two hours. Lysates were retrieved from the beads by aspiration and one wash with the starting volume of RIPA buffer, and then cleared of debris and unlysed cells by centrifugation at 2700 ×g for two minutes. R− cells were lysed by scraping into 60 μl of lysis buffer, consisting of 50mM Tris/HCl, pH 7.6, 150mM NaCl, 1% Triton X-100 plus the above inhibitors. After a 15 minute incubation on ice, lysates were centrifuged at 20 800 ×g for 15 minutes. Protein concentrations in lysates were measured by means of the Bradford microassay (BioRad Laboratories GmbH, Munich, Germany). Aliquots of the of S pombe lysates (10 μg) were separated by 10% SDS polyacrylamide gel electrophoresis (SDS-PAGE), followed by transfer to nitrocellulose. Blots were initially probed with antiphosphotyrosine (PY20; Transduction Laboratories, Lexington, Kentucky, USA), then stripped and re-probed with anti-IGF-1Rβ (Santa Cruz Biotechnology, Santa Cruz, San Francisco, USA). PTP1B expression was confirmed with anti-PTP1B (Upstate Biotechnology, Lake Placid, New York, USA). Duplicate 4–15% gradient gels were used to separate 25 μg of transfected R− lysates. After transfer to nitrocellulose membranes, one membrane was probed for phosphotyrosine and IGF-1R as above. The second membrane was probed with anti-phospho-p42/44 mitogen activated kinase (MAPK), stripped and reprobed with anti-phospho-AKT, followed by re-probing with anti-AKT (all from Cell Signalling Technology, Beverly, Massachusetts, USA) and with anti-MAPK2 (Upstate Biotech). Secondary antibodies labelled with horseradish peroxidase (Dako, Glostrup, Denmark) were used and detection was by chemiluminescence (Super Signal from Pierce, Rockford, Illinois, USA for S pombe experiments; ECL+ from Amersham Pharmacia Biotech, Buckinghamshire, UK for R− experiments).
Immunofluoresence to examine the activity of IGF-1R–GFP and modulation by PTP1B
Following IGF-1 stimulation, cells were washed three times with ice cold PHEM buffer (60mM Pipes, 25mM Hepes, 10mM EGTA, 2mM MgCl2, pH 6.9) then fixed and permeabilised with methanol at −20°C for five minutes. The methanol was then removed by aspiration and PHEM was added to rehydrate the cells for 10 minutes at room temperature. Blocking was performed for 30 minutes with 2.5% goat serum in PHEM followed by incubation for 90 minutes with primary antibody (anti-PTP1B or PY20) in blocking solution. Cells were then washed three times for 10 minutes with PHEM and incubated for one hour with secondary antibody solution (donkey antimouse IgG labelled with Texas red in blocking solution, also containing Hoescht dye). Cells were then washed as before with PHEM and examined and photographed using a SPOT-RT digital camera mounted on a Nikon TE-300 inverted microscope equipped for epifluorescence.
RESULTS
IGF-1R β chain kinase activity in S pombe
In mammalian cells, the full length IGF-1R localises to the cell surface and consists of the extracellular ligand binding α chain and the membrane spanning β chain, which contains the tyrosine kinase domain (fig 1A1). Upon ligand binding, the IGF-1R is thought to undergo a conformational change resulting in activation of the tyrosine kinase domain and autophosphorylation of the receptor. To study IGF-1R tyrosine kinase activity in S pombe, we initially had to establish whether the IGF-1R β chain could be expressed as a functional tyrosine kinase in this organism. The cytoplasmic domain of the IGF-1R β chain encoding residues 931–1337 (βwt) was first subcloned into an S pombe expression vector, under the control of the nmt1 promoter.12 This promoter is repressed in the presence of thiamine and subsequently induced by removal of thiamine from the media, to yield protein expression levels significantly higher than alternative constitutive expression systems.18 Preliminary experiments demonstrated that expression of the 47 kDa βwt was maximal at 24 hours after removal of thiamine. As shown in fig 1B by western blot analysis with antiphosphotyrosine antibodies, βwt autophosphorylates in S pombe and exhibits extensive tyrosine kinase activity towards endogenous yeast proteins. This activity is specific for the wild-type IGF-1R β chain, because phosphotyrosine staining is not observed in lysates from the uninduced culture (thiamine +) (fig 1B). These results demonstrate that in the absence of background tyrosine kinase activity in S pombe, the IGF-1R β chain exhibits an easily measurable phenotype, hence providing a system in which to study modulation of IGF-1R tyrosine kinase activity.
PTP1B inhibits the tyrosine kinase activity of the IGF-1R β chain in S pombe
Several studies have implicated PTP1B as an important regulator of signalling by a variety of tyrosine kinases including the IR. Initial experiments investigated whether PTP1B could also inhibit the tyrosine kinase activity of the IGF-IR β chain, by coexpression of this protein with βwt in S pombe. The full length cDNA for human PTP1B was subcloned into the pADH vector, under the control of the constitutive ADH promoter. pADH–PTP1B or pADH alone were then co-transformed with pRSP–βwt in S pombe cells. βwt expression was induced as before and lysates from induced or uninduced cultures were analysed by immunoblotting and compared with the appropriate vector control. As seen in fig 2, PTP1B completely inhibits βwt induced phosphorylation of endogenous yeast proteins in thiamine deficient cultures. The bottom panels confirm expression of PTP1B and show that βwt expression is unaffected by the expression of PTP1B. Tyrosine phosphorylation of βwt itself is also dramatically reduced in the presence of PTP1B (fig 2). This indicates that PTP1B acts directly on the βwt protein and not merely by dephosphorylation of βwt tyrosine phosphorylated substrates. These results demonstrate that in an S pombe background PTP1B is a potent inhibitor of IGF-1R β chain tyrosine kinase activity.

Effects of protein tyrosine phosphatase 1B (PTP1B) on type 1 insulin-like growth factor receptor (IGF-1R) β chain (βwt) tyrosine kinase activity in Schizosaccharomyces pombe. Schizosaccharomyces pombe cells were transformed with pRSP–βwt and pADH (control) or pRSP–βwt and pADH–PTP1B. These transformants were induced to express βwt, and lysates were analysed by western blotting as in fig 1B. Antiphosphotyrosine staining indicates tyrosine phosphorylation of endogenous yeast proteins when βwt is expressed with the control vector. Tyrosine phosphorylation of endogenous yeast proteins is not observed when both βwt and PTP1B are expressed. Expression of βwt is equal in both induced cultures (thiamine −). Constitutive expression of PTP1B (thiamine + and −) was confirmed with an anti-PTP1B antibody.
Construction and characterisation of GFP–IGF-1R
We were interested in determining which phosphatases could inhibit the kinase activity of the IGF-1R in mammalian cells. This was initially tested by transient transfection of PTP1B into a cell line stably overexpressing the IGF-1R and examination of the tyrosine phosphorylation status of the receptor at various time points after IGF-1 stimulation by western blotting. However, owing to background tyrosine kinase activity and a masking of any alterations in IGF-1R phosphorylation by non-transfected IGF-1 stimulated cells, no detectable difference was observed. Therefore, it was necessary to develop a method for analysing IGF-1R phosphotyrosine activity in individual cells using a transient transfection system and immunofluorescence. Analysis of cellular phosphotyrosine content by immunofluoresence provides a means of studying tyrosine kinase activity in individual cells in a transiently transfected population.
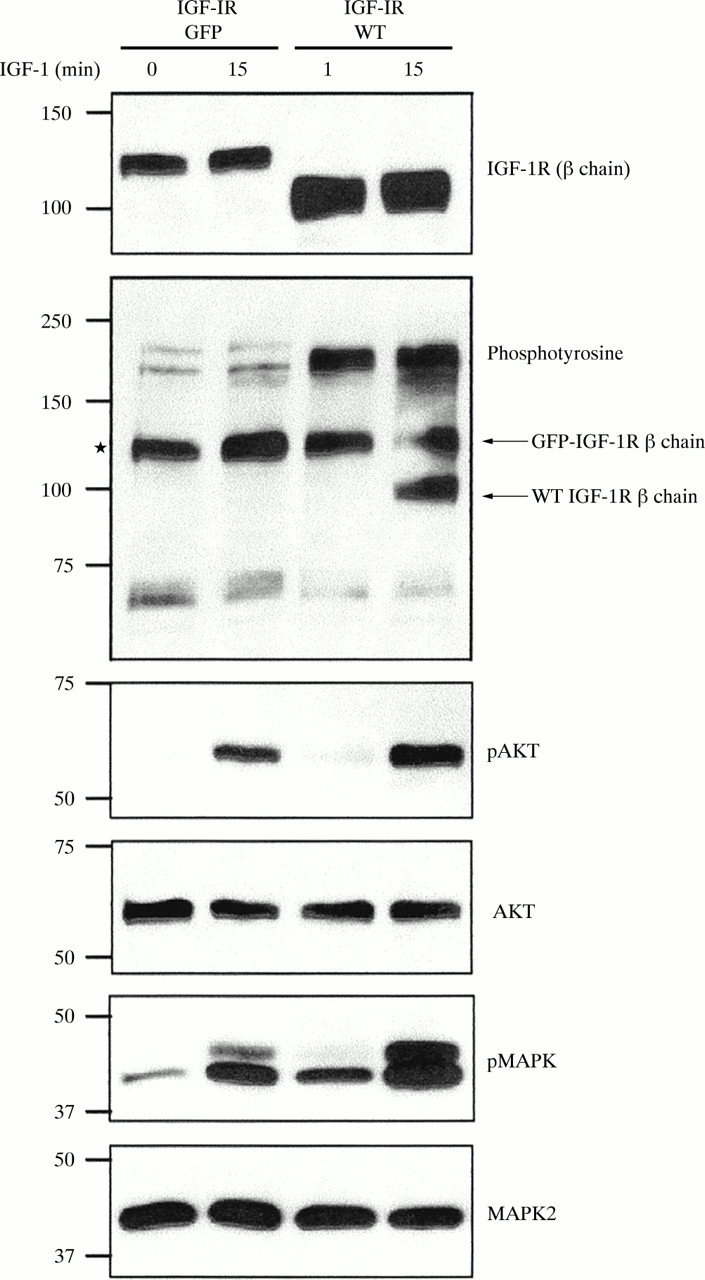
To study the effect of phosphatases on the IGF-1R using immunofluoresence, we first constructed a GFP tagged IGF-1R, as described in the materials and methods section. To determine whether addition of the 26 kDa GFP moiety to the C-terminus of the receptor had an effect on IGF-1R signalling, we assessed the ability of GFP–IGF-1R to autophosphorylate in response to IGF-1 stimulation and subsequently activate the phosphatidylinositol 3-kinase (PI3K) and MAPK signalling pathways. R− cells were transiently transfected with either GFP–IGF-1R or wild-type IGF-1R and 48 hours after transfection cells were stimulated with IGF-1 for 15 minutes. Expression of the GFP–IGF-1R was confirmed with an antibody specific for the C-terminus of the IGF-1R β chain. The β chain subunit migrates on SDS-PAGE at the expected molecular weight, which is approximately 26 kDa greater than the 100 kDa wild-type receptor β chain (fig 3). The wild-type IGF-1R becomes autophosphorylated on tyrosine residues in response to IGF-1. Tyrosine phosphorylation of the GFP–IGF-1R was also seen following IGF-1 stimulation, although visualisation of this is impeded by a constitutively phosphorylated protein of similar molecular weight, indicated by the asterisk in fig 3. Western blot analysis with antibodies for phospho-AKT and phospho-MAPK show that these proteins are phosphorylated in response to IGF-1 (fig 3), thus demonstrating that the GFP–IGF-1R can activate the PI3K and MAPK pathways at levels comparable to the wild-type IGF-1R.

Green fluorescent protein–type 1 insulin-like growth factor receptor (GFP–IGF-1R) signalling compared with wild-type IGF-1R. R− cells were transiently transfected with pIGF-1R–GFP or pCDNA3–IGF-1R (WT). After 24 hours, each transfection was split into two wells. After a further 24 hours one well from each transfection was treated with IGF-1 (100 ng/ml) for 15 minutes, whereas the other was left untreated (0 min IGF-1). Lysates were made and 25 μg of protein was separated using 4–15% gradient sodium dodecyl sulphate polyacrylamide gel electrophoresis, followed by western blot analysis. Expression of both GFP–IGF-1R and WT IGF-1R was confirmed with an anti-IGF-1R β chain antibody. Antiphosphotyrosine staining indicates phosphorylation of the GFP–IGF-1R β chain (*, discussed in results), and of the IGF-1R wild-type β chain. Anti-phospho-AKT and anti-phospho-MAPK2 antibodies demonstrate phosphorylation of these proteins by GFP–IGF-1R and WT IGF-1R in response to IGF-1 stimulation. Equal loading was shown by immunoblotting with anti-AKT and anti-MAPK2 antibodies.
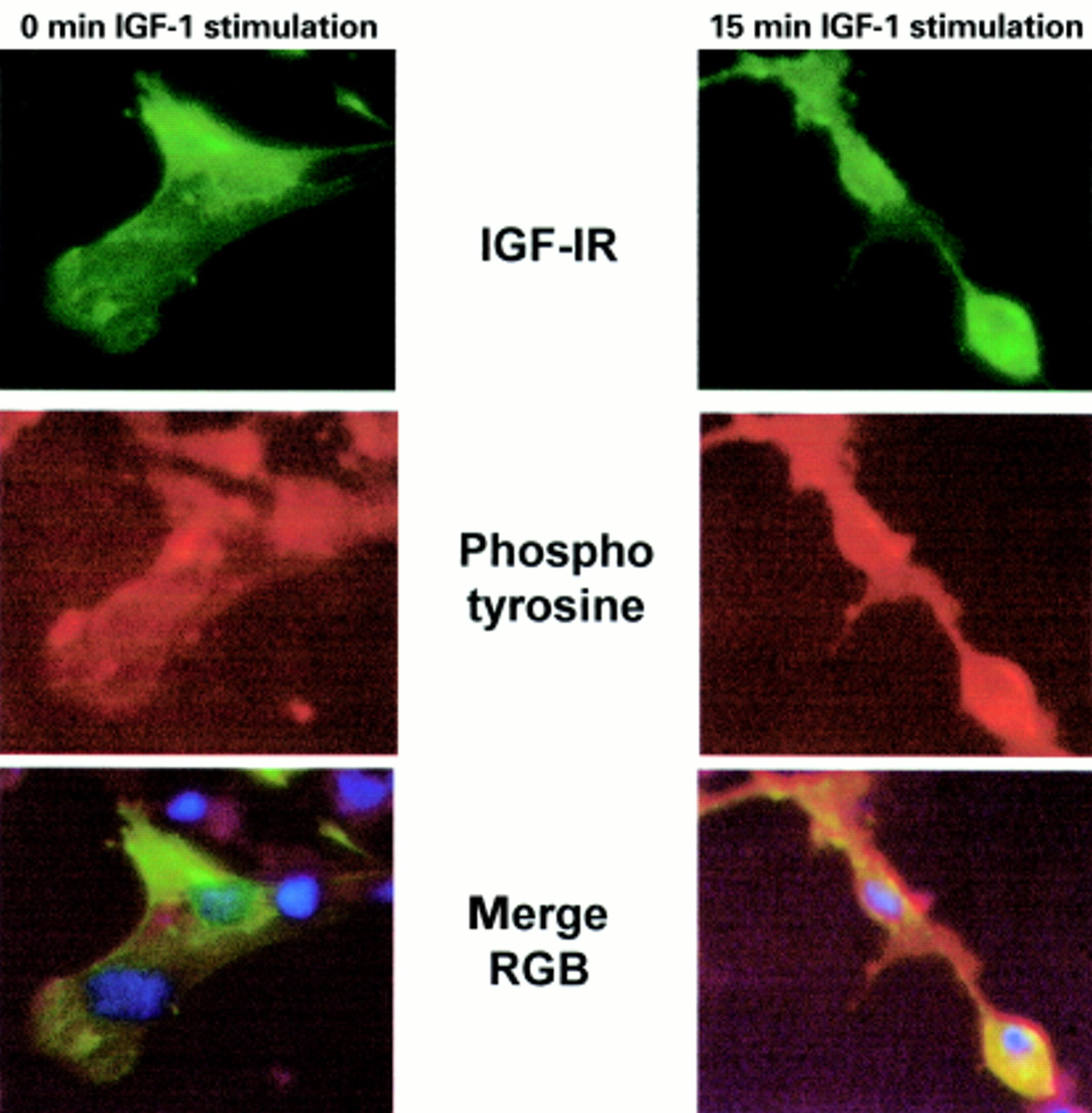
We next investigated whether we could detect GFP–IGF-1R phosphorylation by immunofluorescent staining for phosphotyrosine in the cell. R− cells were transfected with GFP–IGF-1R and 48 hours later cells were stimulated with IGF-1, or left unstimulated, followed by staining with an antiphosphotyrosine antibody and a Texas red labelled secondary antibody. As fig 4 shows, there is increased phosphotyrosine staining in transfected cells in response to IGF-1. Examination of the merged image shows that this red staining colocalises with the GFP–IGF-IR and is found predominately at the cell surface in response to IGF-1 stimulation. Altogether, these data demonstrate that the addition of a C-terminal GFP moiety does not impede IGF-1R signalling and localisation, and that the GFP–IGF-1R can be used to study IGF-1R signalling.
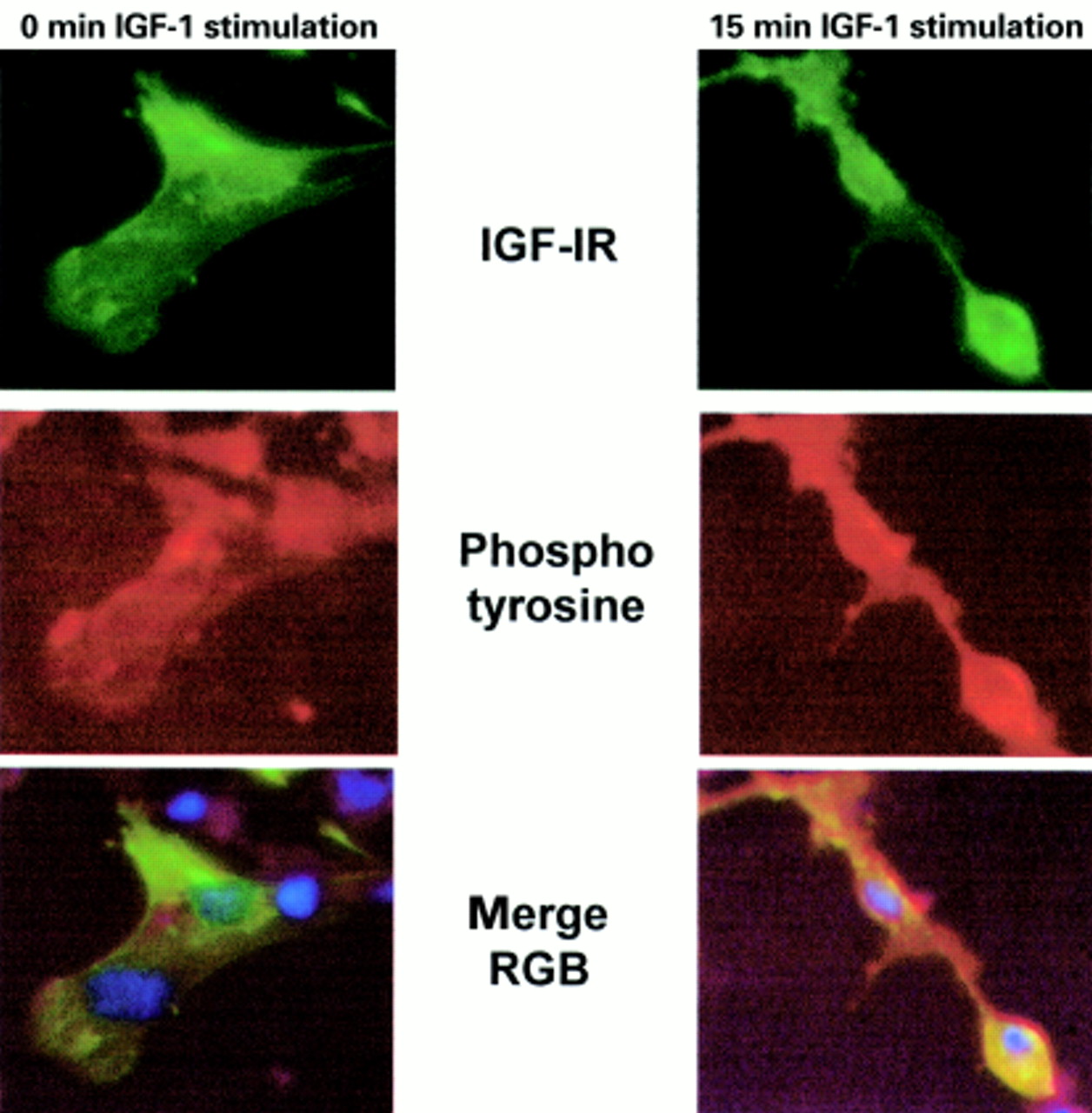
Immunofluorescent detection of type 1 insulin-like growth factor (IGF-1) induced tyrosine phosphorylation in cells transfected with green fluorescent protein –IGF-1 receptor (GFP–IGF-1R). R− cells were transiently transfected with GFP–IGF-1R and 48 hours after transfection cells were stimulated with IGF-1 for 15 minutes, or left untreated. Cells were fixed and stained for phosphotyrosine as described in the materials and methods. Stained cells were examined and photographed under ×40 magnification, using the blue filter to visualise GFP–IGF-1R (green) and the fluorescein isothiocyanate filter to visualise phosphotyrosine staining (red). Untreated cells are shown in the left panel and colocalisation of red (phosphotyrosine) and green (GFP–IGF-1R) is not seen in the merged image. In IGF-1 stimulated cells, shown in the right panel, colocalisation of GFP–IGF-1R with phosphotyrosine staining is observed, and appears yellow in the merged image.
Analysis of the effects of PTP1B on GFP–IGF-1R activity
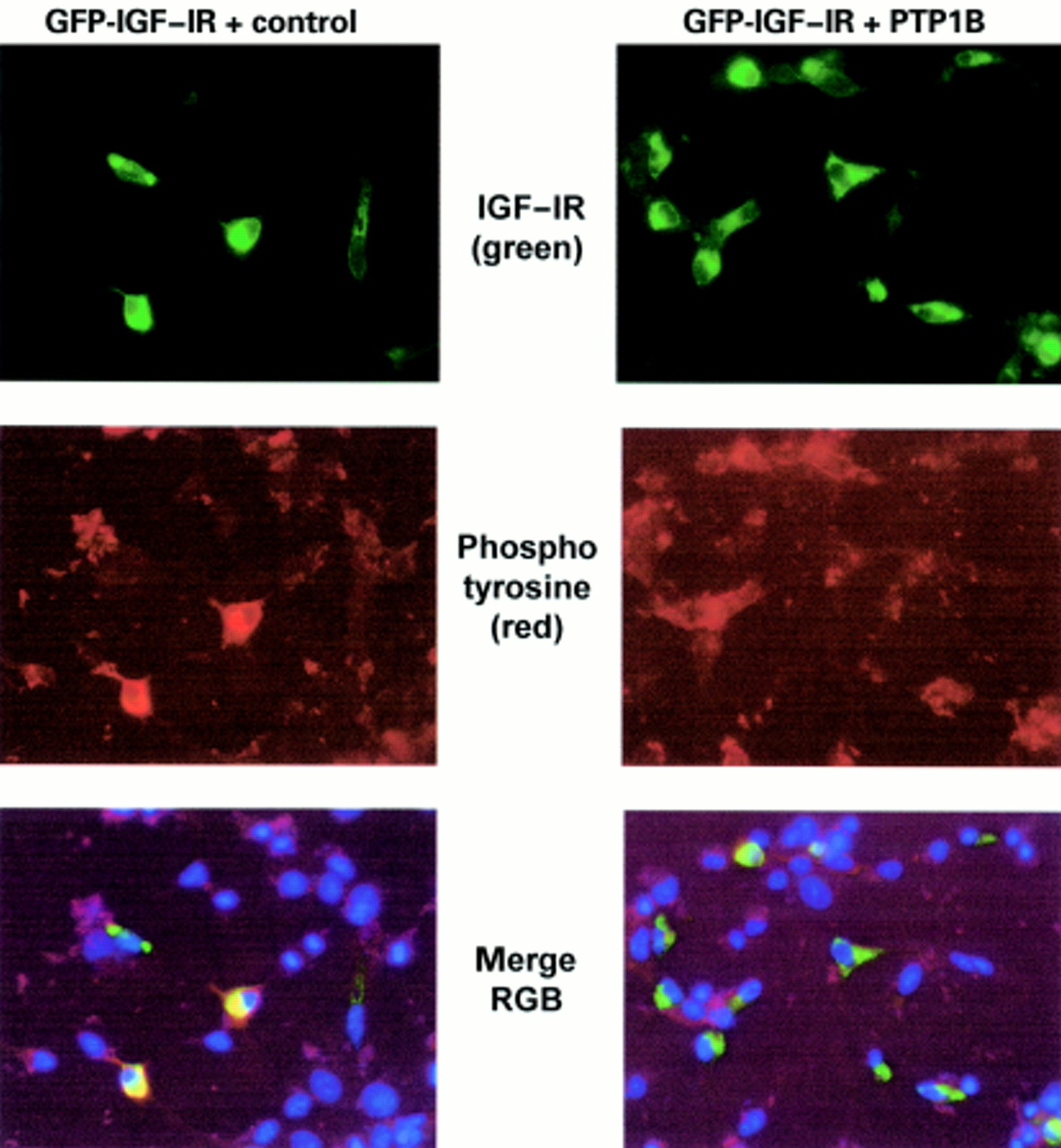
As shown above, GFP–IGF-1R kinase activity in response to IGF-1 stimulation can be visualised via colocalisation with a phosphotyrosine signal. Therefore, we wanted to know whether the inhibition of kinase activity by a phosphatase could also be measured by imunofluorescence. To do this we investigated the effect of coexpressing PTP1B on GFP–IGF-1R tyrosine kinase activity. COS7 cells were transfected with GFP–IGF-1R and either PTP1B or the appropriate control vector (fig 5, right and left panels, respectively). As before, cells were serum starved and stimulated with IGF-1 for 15 minutes. Duplicate transfected populations were stained for PTP1B or phosphotyrosine and examined by immunofluoresence under reduced magnification (×10) to allow visualisation of a range of cells in each field. In Fig 5, GFP–IGF-1R is strongly expressed in a subpopulation of cells (top panel). PTP1B cellular protein is stained with an anti-PTP1B antibody and a red secondary antibody. Upon transient transfection, significantly higher amounts of PTP1B are expressed in transfected cells than are detectable in the surrounding non-transfected cells (middle panel). Examination of the merge clearly indicates colocalisation of GFP–IGF-1R with overexpressed PTP1B, as indicated by yellow staining in the PTP1B transfected cells (bottom panel). This confirms that a significant subpopulation of cells are transfected with both proteins. Figure 6 demonstrates that when a duplicate population of cells is stained for phosphotyrosine, this pattern of red colocalisation with GFP is altered. In cells transfected with GFP–IGF-1R and the control vector, there is an increase in phosphotyrosine staining in the GFP–IGF-1R expressing subpopulation. However, in cells overexpressing PTP1B, cellular phosphotyrosine staining is significantly reduced. Comparison of the merged images clearly demonstrates that, when overexpressed with the GFP–IGF-1R, PTP1B has the ability to inhibit the tyrosine kinase activity of the IGF-IR.

Coexpression of protein tyrosine phosphatase 1B (PTP1B) with type 1 insulin-like growth factor (IGF-1) stimulated green fluorescent protein–IGF-1 receptor (GFP–IGF-1R). COS7 cells were transiently transfected with pIGF-1R–GFP + pIRES (left panel) or pIGF-1R–GFP + pIRES-PTP1B (right panel) and stimulated with IGF-1 as before. The top panels show GFP–IGF-1R expression (green). Immunofluorescent staining with an anti-PTP1B antibody indicates expression of endogenous PTP1B in the control vector transfected cells (middle left panel: red) and overexpression of PTP1B in cells transfected with pIRES–PTP1B (middle right panel: brighter red fluorescence). Examination of the merged images indicates colocalisation of GFP–IGF-1R with overexpressed PTP1B in a subpopulation of cells (yellow).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Duplicate transfection to fig 5 but stained with antiphosphotyrosine antibodies. Expression of type 1 green fluorescent protein–type 1 insulin-like growth factor receptor (GFP–IGF-1R) is shown in the top panels. Phosphotyrosine staining (red) is shown in the middle panels and is decreased in cells transfected with pIGF-1R–GFP and pIRES–protein tyrosine phosphatase 1B (PTP1B) (middle right panel). In the bottom panels the merged images show that colocalisation of cellular phosphotyrosine content with GFP–IGF-1R is reduced in cells also expressing PTP1B, as indicated by a decrease in yellow staining.
DISCUSSION
Despite the importance of the IGF-1R in normal cellular function and its implications as a tumorigenic agent, very little is known about proteins that downregulate the tyrosine kinase activity of the receptor. To a certain extent, downregulation of receptor tyrosine activity is achieved during endosomal processing of the receptor and by degradation following ubiquitination. However, it is well established that the activity of many tyrosine kinases, including the tyrosine kinase receptors for PDGF, EGF, and insulin, are regulated by the action of protein tyrosine phosphatases.19 We were interested in developing a system and tools for the identification of IGF-1R regulatory phosphatases.
The fission yeast S pombe provides an ideal model system in which to study the activity of protein tyrosine kinases because it signals mainly through serine/threonine or dual specificity kinases and phosphatases. Hence, mammalian tyrosine kinases can be studied in the absence of background tyrosine kinase activity. The activity displayed by IGF-IR βwt is comparable to Src tyrosine kinase activity in S pombe7 and is far greater than that displayed by the PDGF receptor β.8 We have also shown that this system can be used to identify potential regulators, including PTPs, which could downregulate the tyrosine kinase activity of the full length IGF-1R. From the results presented here, it is clear that PTP1B can have a dramatic affect on the activity of the IGF-1R β chain. Bwt induced tyrosine phosphorylation of endogenous yeast proteins is completely abolished by coexpression of PTP1B and autophosphorylation of βwt is also dramatically reduced. PTP1B was expressed from the constitutive ADH promoter, which has previously been shown to result in reduced expression compared with the nmt1 promoter (N Spellacy and D Buckley, 1999 unpublished observations).18 It is tempting to speculate that PTP1B, if expressed at higher or similar levels to βwt, would completely inhibit autophosphorylation and subsequent tyrosine kinase activity of the IGF-1R β chain.
PTP1B did not cause a reduction in IGF-1R activity as measured by conventional biochemical analysis in a cell line overexpressing the IGF-1R and transiently expressing PTB1B. This is a common problem associated with transient transfection because differences in signalling are often masked by signals from the untransfected population, which makes up more than 50% of the cells.
“PTP1B was the first identified protein tyrosine phosphatase and has been implicated in several studies as a negative regulator of insulin signalling”
To analyse the effect of overexpression of PTP1B or other phosphatases on IGF-1R activity in a transiently transfected population, we have developed a method for analysing IGF-1R tyrosine kinase activity by immunofluoresence. A GFP tagged IGF-1R was constructed and characterised for expression and signalling activities. It appears that the GFP–IGF-1R is expressed at lower levels than wild-type IGF-1R, even though both are expressed under the control of the cytomegalovirus promoter. Possible explanations for this could be a reduction in antibody affinity for this receptor or a decrease in protein stability, either of which could result from the addition of the 26 kDa GFP tag at the C-terminus. However, western blot analysis showed that the GFP–IGF-1R is phosphorylated in response to IGF-1 stimulation and can activate the PI3K and MAPK pathways at levels comparable to the wild-type receptor. This shows that the addition of a GFP tag does not impede IGF-1R activity and that the GFP–IGF-1R can be used to study IGF-1R signalling. Immunofluorescent staining for cellular phosphotyrosine content showed that the GFP–IGF-1R is activated in response to IGF-1 stimulation and is localised predominantly at the cell surface, again supporting the use of this chimaera in IGF-1R studies. GFP–IGF-1R was then used to demonstrate that PTP1B can inhibit the tyrosine kinase activity of the full length IGF-1R.
PTP1B was the first identified protein tyrosine phosphatase20 and has been implicated in several studies as a negative regulator of insulin signalling. PTP1B has also been shown to play an important role in integrin signalling21 and to be involved in regulation of Src activity.22 PTP1B coexpression results in a clear reduction of IGF-1 induced cellular tyrosine phosphorylation by GFP–IGF-1R. The functional consequences of PTP1B inhibition of IGF-IR kinase activity is a current focus of study in our laboratory and is of particular interest because of the knockout mice studies suggesting that PTP1B can specifically regulate IR signalling without affecting the IGF-1R.5 These initial findings also suggest that further analysis of the role of PTP1B in IGF-1R signalling will be necessary before PTP1B can be widely used as a therapeutic target in the treatment of type 2 diabetes, and that overexpression of PTP1B could block IGF-1R signalling in IGF-1R transformed cells.
In conclusion, we have presented two new methods that can be used in the study of IGF-1R kinase regulation by phosphatases. Using these methods we have also shown that PTP1B can negatively regulate IGF-1R tyrosine kinase activity.
Take home messages
-
Schizosaccharomyces pombe can be used to analyse the tyrosine kinase activity of the insulin-like growth factor type 1 receptor (IGF-1R) β chain and its regulation by tyrosine phosphatases
-
The regulation of IGF-1R tyrosine kinase activity can be studied using a green fluorescent protein tagged IGF-1R
-
Using both of these methods, we showed that IGF-1R kinase activity can be inhibited by the protein tyrosine phosphatase, PTP1B
Acknowledgments
We are grateful to G Superti-Furga for providing the S pombe strain and plasmids, B Neel for the pKS PTP1B plasmid, and P Walsh for critically reading the manuscript. This work was supported by research grants from Enterprise Ireland, The Wellcome Trust, and Tanaud Ireland, Inc.