


Abstract
The activity of large conductance, Ca2+-sensitive K+ (BKCa) channels, known to control neuronal excitability, is increased by ethanol (EtOH) exposure. Moreover, brain cholesterol (CHS) is elevated after chronic exposure to EtOH, suggesting that membrane CHS may play a role in drug tolerance. Here, we use BKCa channels from human brain (hslo subunits), reconstituted into 1-palmitoyl-2-oleoyl phosphatidylethanolamine/1-palmitoyl-2-oleoyl phosphatidylserine (POPS) bilayers, to examine CHS modulation of EtOH sensitivity. Acute exposure to clinically relevant EtOH levels increases channel activity without modifying conductance. In this minimal system, increases in CHS content within the range found in neuronal membranes lead to progressive antagonism of EtOH action. Furthermore, CHS inhibits basal channel activity with an affinity similar to that of CHS blunting of the alcohol effect. Modification of channel gating by either EtOH or CHS is reduced dramatically by removal of POPS from the bilayer, suggesting a common mechanism(s) of action. Indeed, channel dwell-time analysis indicates that CHS and EtOH exert opposite actions on the stability of channel closed states. However, each agent also acts on distinct dwell states not mirrored by the other, which contribute to the opposite effects of CHS and EtOH on channel gating.
Ion channels reside in a heterogeneous lipid matrix. Lipid species partition asymmetrically both within and across biological membrane leaflets (Devaux, 1991). These nonrandom lipid associations produce domains that differ in composition and physicochemical properties from the bulk membrane (Welti and Glaser, 1994), resulting in distinct microenvironments for ion channels. An example is the cholesterol (CHS) and sphingomyelin-rich lipid raft, thought to participate in many aspects of cell function (Brown and London, 1998). Large conductance, Ca2+-activated K+ (BKCa) channels cloned from human brain (hslo) and expressed in Madin-Darby canine kidney cells associate with these lipid microdomains (Bravo-Zehnder et al., 2000). The biological implications of channel association with distinct lipid domains are not well understood, but it is likely that the domain physical properties influence channel activity.
Cholesterol is a significant component of lipid rafts and a major determinant of overall membrane physical properties (Bloch, 1983), which may contribute to its effects on the activity of native ion channels (Bolotina et al., 1989; Barrantes, 1993; Chang et al., 1995; Lundbaek et al., 1996; Levitan et al., 2000). Modulation of membrane CHS content and distribution may play a role in cellular adaptation to ethanol (EtOH) (Wood et al., 1990). Increased content (Chin et al., 1978; Omodeo-Sale et al., 1995) and altered distribution of CHS between membrane leaflets (Wood et al., 1990) are observed in response to EtOH exposure in both animal and cell culture models. Ethanol differentially increases the diffusion of various lipid probes in Aplysia californica neurons, suggesting that its actions on ion channels might depend upon the existence of dissimilar lateral domains (Treistman et al., 1987). In model membranes and computer simulations, EtOH affects lateral lipid domains (Chin and Goldstein, 1981; Harris et al., 1984; Jorgensen et al., 1993; Slater et al., 1993), suggesting that specific lipid species might modify the sensitivity of a domain to EtOH action. Cholesterol, in particular, counteracts EtOH's disordering action in mouse synaptosomal membranes and phospholipid bilayers (Chin and Goldstein, 1981). Interestingly, EtOH increases the fluidity of the extracellular leaflet of synaptic plasma membranes to a larger extent than that of the cytoplasmic leaflet, an effect attributed to the larger CHS content in the inner leaflet. After chronic EtOH treatment, however, transbilayer differences in fluidity and CHS content are reduced in concert (Wood et al., 1990).
Ethanol reversibly potentiates BKCa channels in excised membrane patches from rat neurohypophysial terminals, an action that may contribute to EtOH inhibition of neuropeptide release (Dopico et al., 1996). EtOH potentiation persists after expression of BKCa (mslo) channels in Xenopus laevis oocytes (Dopico et al., 1998) and incorporation of native BKCa channels into 1-palmitoyl-2-oleoyl phosphatidylethanolamine (POPE)/1-palmitoyl-2-oleoyl phosphatidylserine (POPS) bilayers (Chu et al., 1998). Thus, neither cytosolic second messengers nor complex cytoskeletal architecture is required for EtOH action on BKCa channels.
Here, we use hslo channels expressed in HEK 293 cells and incorporated into bilayers made of only one or two phospholipid species to study CHS modulation of EtOH action on channel function. This preparation allows near complete control of both protein and lipid constituents. Results indicate that increased bilayer CHS antagonizes EtOH potentiation of channel activity in a concentration-dependent manner. The reciprocal actions of EtOH and CHS on commonly targeted channel dwell states are the major determinants of CHS antagonism of EtOH effect on channel activity. Furthermore, the effect of each of these modulators on channel activity is drastically reduced in the absence of phosphatidylserine in the bilayer.
Materials and Methods
HEK 293 Membrane Preparation. HEK 293 membrane fragments were isolated using a protocol for COS cells (Sun et al., 1994), modified slightly. Briefly, HEK 293 cells stably transfected with hslo cDNA (a gift from Dr. P. Ahring, NeuroSearch A/S, Ballerup, Denmark) were grown to confluence, pelleted, and resuspended on ice in 10 ml of buffer: 30 mM KCl, 2 mM MgCl2, 10 mM HEPES, and 5 mM EGTA, pH 7.2. The cell suspension was forced through a 27-gauge needle four times and sonicated at 30% maximum power for 30 s, twice. The suspension was layered on a 20 to 38% sucrose density gradient (in 20 mM MOPS, pH 7.1) and centrifuged at 25,000 rpm for 60 min at 4°C. The band at the 20 to 38% interface was collected with a syringe, diluted with bidistilled H2O, and centrifuged in a 50.2 Ti rotor at 45,000 rpm for 60 min at 4°C. The resulting pellet was resuspended in 200 μl of buffer: 250 mM sucrose and 10 mM HEPES, pH 7.3. Aliquots were stored at -80°C.
Electrophysiology. Channels were incorporated by dropping 0.5 μl of the membrane preparation onto bilayers consisting of POPE/POPS and differing concentrations of CHS. Lipids were dried under N2 gas and resuspended in decane in a 3:1 (w/w) POPE/POPS ratio, with 0 to 49 mol% CHS. The final lipid concentration was 25 mg/ml. Bilayers were formed by painting the lipid mixture across a 100 μm hole formed in a plastic coverslip (Wonderlin et al., 1990). Capacitance was monitored by the capacitive current generated by a triangle pulse (20 mV/25 ms). Vesicle fusion was promoted by an osmotic gradient, with the cis chamber (to which the vesicles were added) hyperosmotic to the trans. Only channels with their Ca2+-sensor facing the cis chamber were studied. Solutions consisted of the following: cis, 300 mM KCl, 10 mM HEPES, 1.10 mM N-(2-hydroxyethyl) ethylenediaminetriacetic acid (HEDTA) ([Ca2+]free ≈ 50 μM) or 1.45 mM HEDTA ([Ca2+]free ≈ 15 μM), and 1.05 mM CaCl2, pH 7.2; and trans, 30 mM KCl, 10 mM HEPES, and 0.1 mM HEDTA, pH 7.2. CHS action on baseline channel function was studied at 50 μM [Ca2+]free, and 0 mV, which promotes high channel activity from which the inhibitory effects of CHS can be effectively quantitated. EtOH sensitivity was tested at 10 μM [Ca2+]free, at potentials between -10 and +60 mV, which yields lower activity and prevent a “ceiling effect” when measuring EtOH potentiation. EtOH modification of slo activity is independent of voltage within this range (Dopico et al., 1998). [Ca2+]free in the cis chamber was adjusted using aliquots from a 1 M stock solution of HEDTA, pH 7.2. [Ca2+]free values given are nominal, calculated using the Max Chelator Sliders program (C. Patton, Stanford University, Palo Alto, CA).
Experiments were performed at room temperature (20–25°C) to facilitate comparison with our previous studies examining EtOH action on BKCa channels (Dopico et al., 1996, 1998; Chu et al., 1998). Single channel events were recorded at a bandwidth of 10 kHz using a patch-clamp amplifier (model 8900; Dagan, Minneapolis, MN) and stored on videotape using pulse code modulation (model DMP-100; Nakamichi, Tokyo, Japan). Data were low-pass filtered at either 3 (for dwell-time analysis) or 1 kHz (for display and NPo determination) using an eight-pole Bessel filter (model 902; Frequency Devices, Haverhill, MA) and digitized at 10 kHz.
Data Analysis. Data were acquired and analyzed using pClamp 6.0.2 (Axon Instruments, Inc., Union City, CA). As an index of steady-state channel activity, we used the product of the number of channels in the bilayer during recording (N) and the open channel probability (Po). N was monitored pre- and post-EtOH by stepping to positive potentials to maximize Po. Experiments showing an increase in N after EtOH addition were discarded. NPo was determined as described elsewhere (Dopico et al., 1996) from periods of at least 20 s of continuous recording. Dwell-time histograms were constructed using the half-amplitude threshold criterion, events shorter than 0.3 ms being excluded. An F table (p < 0.01) was used to determine the minimum number of exponential components to appropriately fit dwell-time histogram data. Fifty percent of maximal effect (EC50 or IC50) was obtained from concentration-response curves by extrapolation. Data are shown as mean ± S.E.M. The significance of the difference between means was determined by ANOVA and an a posteriori test (Dunnett's).
Chemicals. All solutions were prepared with Milli-Q water and ultrapure grade salts. Ethanol (100%, anhydrous) was purchased from American Bioanalytical (Natick, MA), decane (>99% pure, anhydrous) from Sigma-Aldrich Chemical Co. (Milwaukee, WI), and POPE, POPS, and CHS from Avanti Polar Lipids (Alabaster, AL).
Results
Ethanol Increases hslo Channel Activity in Binary Bilayers. We first determined whether EtOH modifies hslo steady-state activity (NPo) in a binary phospholipid mixture. We chose a 3:1 POPE/POPS (w/w) planar bilayer, where EtOH sensitivity of native skeletal muscle BKCa channels was initially explored (Chu et al., 1998). Thus, cloned hslo subunits were incorporated into this bilayer type, where they displayed characteristic features of BKCa channels: large unitary current amplitude (Fig. 1) and increases in Po as the applied voltage is made more positive (9.8 mV ± 0.4 mV/e-fold change in NPo) and/or [Ca2+] at the cytosolic side of the channel is increased (not shown). Figure 1 shows traces of hslo activity before and during application of 50 mM EtOH to the “intracellular” side of the POPE/POPS bilayer. The EtOH-induced increase in Po shown in the figure, which occurred within 1 to 5 min of drug exposure, was observed in 8 of 10 bilayers, the average NPo showing a 5.2 ± 1.5-fold increase over pre-EtOH values. EtOH-induced potentiation of hslo channel activity in the POPE/POPS bilayer is similar to our previously reported results in complex lipid/protein systems with mslo subunits expressed in oocytes (Dopico et al., 1998), native BKCa channels studied in situ (Dopico et al., 1996), and rat skeletal muscle t-tubule BKCa channels reconstituted into this bilayer type (Chu et al., 1998). Thus, hslo subunits, POPE, POPS, and the system interfaces are sufficient for EtOH potentiation of BKCa channel activity.

Ethanol (50 mM) increases hslo channel activity in planar POPE/POPS bilayers. Traces of hslo activity before (left) and during (right) application of EtOH to the intracellular (cis) side of the same POPE/POPS 3:1 (w/w) bilayer. The potential across the bilayer was set at 0 mV and free [Ca2+]ic ≈ 10 μM. Data were low pass filtered at 1 kHz and sampled at 10 kHz. Arrows denote the current level corresponding to channel closed states. Steady-state channel activity (NPo) was determined from continuous recording (see Materials and Methods).
In contrast to its action on steady-state activity, 50 mM EtOH consistently failed to modify other parameters of hslo function, such as unitary amplitude (14.9 versus 14.8 pA, recorded at 0 mV and 300/30 mM [K+]i/[K+]o in the presence and absence of EtOH, respectively) (Fig. 1), suggesting that EtOH actions on hslo channels are restricted to modification of channel gating. This is also in agreement with data obtained with BKCa channels in more complex lipid/protein systems (Dopico et al., 1996, 1998; Jakab et al., 1997; Chu et al., 1998), validating our minimal system for studies on CHS modulation of EtOH sensitivity.
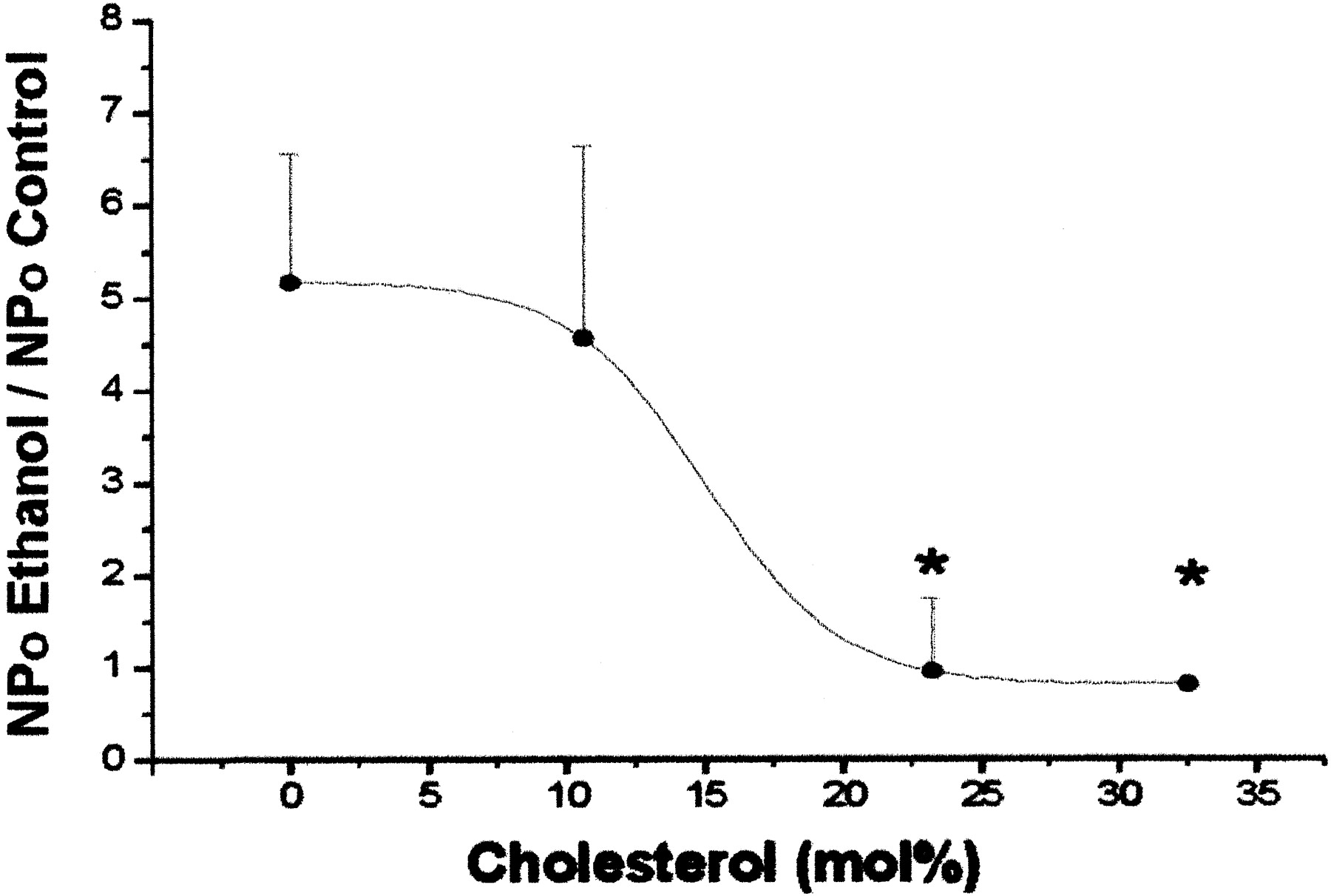
Cholesterol Blunts Alcohol Potentiation of hslo Channel Activity. We next tested whether CHS incorporation into this binary bilayer could modify EtOH-induced increases in hslo activity. Figure 2 shows that CHS incorporation (11–33 mol%) into POPE/POPS bilayers diminishes EtOH-induced potentiation in a concentration-dependent manner (IC50 = 15 mol%). CHS content of 23 mol% resulted in almost total prevention of channel potentiation by 50 mM EtOH. Under these conditions, EtOH slightly increased channel activity in two of five experiments, and slightly inhibited activity in the remaining three cases, yielding no net effect on channel activity (average NPo value = 0.96 ± 0.38 of controls). This lack of a major alcohol effect in the presence of CHS may be explained by 1) reduced EtOH partitioning into CHS-containing bilayers; 2) CHS antagonism of EtOH at EtOH's recognition site(s) on the hslo subunit or its immediate phospholipid microenvironment (see Discussion); and/or 3) the actions of CHS and EtOH on channel kinetic states are of opposite sign, and balanced, which results in reciprocal modulation of basal channel activity.
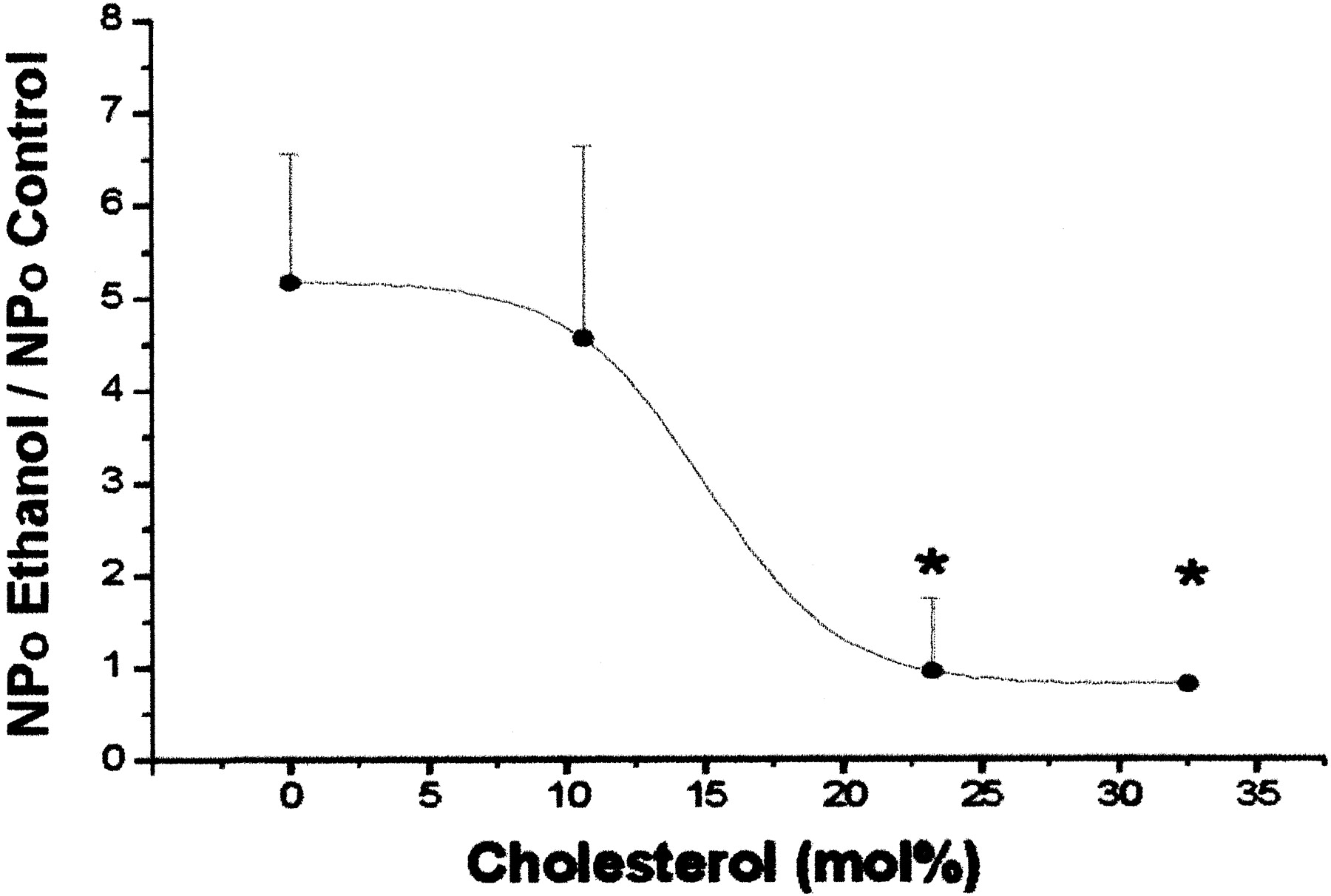
Cholesterol antagonizes EtOH-induced potentiation of hslo channel activity in POPE/POPS 3:1 (w/w) bilayers. The plot shows that average ratios of hslo NPo in the presence and absence of 50 mM EtOH are progressively decreased as the bilayer CHS content is increased. EtOH potentiation is significantly reduced at ∼23 mol% [CHS] (IC50 = 15 mol%). The potential at the cis side of the bilayer was set between -10 and +20 mV, and free [Ca2+]ic ≈ 10 μM. Data were low pass filtered at 1 kHz and sampled at 10 kHz. NPo values were determined from continuous recording (see Materials and Methods). Data are mean ± S.E.M.; n = 2 to 8 bilayers; *, significantly different from POPE/POPS (control) values (p < 0.05; ANOVA and Dunnett's test).
Cholesterol Reduces Basal hslo Channel Activity in POPE/POPS Bilayers. A reduction in hslo activity caused by the presence of CHS in the bilayer could explain part or all of the CHS antagonism of EtOH potentiation of hslo activity in POPE/POPS bilayers. Figure 3A shows hslo single channel activity recorded under identical conditions of voltage and free [Ca2+]ic at the cis side of POPE/POPS bilayers in the absence and presence of CHS in this bilayer. Increases from 11 to 49 mol% inhibit hslo activity in a concentration-dependent manner. Maximal inhibition is reached at 33 to 49 mol% CHS with an IC50 = 15.5 mol% (Fig. 3B). Both the concentration for maximal effect and the IC50 are similar to those for CHS blunting of EtOH-induced increases in hslo activity (see above), which suggests that CHS inhibition of channel basal activity contributes to the sterol modulation of alcohol-induced potentiation of hslo activity.

Inhibition of basal hslo channel activity and its underlying changes in mean open and closed times, as a function of cholesterol content in POPE/POPS 3:1 (w/w) bilayers. A, single current traces displaying 2 s of continuous recording obtained in the absence (top) and presence (bottom) of 49 mol% CHS. The potential at the cis side of the bilayer was set at 0 mV, and free [Ca2+]ic ≈ 50 μM. Data were low pass filtered at 1 kHz and sampled at 10 kHz. Arrows denote the current level corresponding to channel closed state(s). B, increases in CHS content (11–49 mol%) inhibit hslo activity (Po) in a concentration-dependent manner. Maximal inhibition is reached at 33 to 49 mol% CHS (IC50 = 15.5). C, channel mean open time (to) is maximally inhibited at 23 mol% CHS (IC50 = 4.3), D, channel mean closed time (tc) fails to reach a well defined maximum but progressively increases as CHS content in the bilayer increases. If a maximal effect is assumed to be reached at the maximal [CHS] tested (49 mol%), data extrapolation renders EC50 ≥ 36 mol%. Both to and tc were obtained by weighting the different exponential components of respective dwell-time distributions (Fig. 7). For B to D, data are mean ± S.E.M.; n = 2 to 8 bilayers; *, significantly different from POPE/POPS (control) values (p < 0.05); **, significantly different from POPE/POPS (control) values (p < 0.01) (ANOVA and Dunnett's test).
Inhibition of basal Po could result from a CHS-induced decrease in channel mean open time (to), an increase in channel mean closed time (tc), or a combination of both. Figure 3, C and D, clearly demonstrates that the third possibility is the case. Furthermore, single channel analysis reveals differential CHS effects on to and tc, dependent upon concentration. Although inhibition of channel to is maximal at 23 mol% CHS (IC50 = 4.3 mol%) (Fig. 3C), channel tc failed to reach a well defined maximum at the concentrations tested (Fig. 3D). If a maximal effect is assumed at the highest [CHS] tested (49 mol%), tc data extrapolation yields an EC50 = 36.1 mol%, representing a minimum for this value. Thus, at concentrations of CHS below the IC50 for the reduction in Po, decreases in channel activity are determined by a major reduction in to and a minor increase in tc. In contrast, above the IC50, further reduction in Po is caused primarily by a progressive increase in tc.
As with EtOH (see above), CHS-induced modifications of hslo channel Po were not accompanied by significant changes in conductance (pS): 323.3 ± 5 in POPE/POPS (n = 12), 329.2 ± 4 in POPE/POPS + 13% CHS (n = 11), and 335.3 ± 7.3 in POPE/POPS + 33% CHS (n = 4); all measurements obtained in 300/30 mM [K+]i/[K+]o (r2> 0.95 for linear fits of unitary current amplitude/voltage data). This result is in agreement with studies of CHS action on BKCa channels in rabbit aorta (Bolotina et al., 1989). A subtle 7% change in slope conductance has been reported for rat brain BKCa channels incorporated into POPE/POPS (55:45) bilayers containing 11% CHS (w/w) (Chang et al., 1995), an effect not apparent in our system. Those experiments were conducted in the presence of a 300/100 mM (cis/trans) KCl gradient, whereas ours were performed with a larger 300/30 mM KCl gradient. This lower trans chamber [K+] in our experiments probably created a driving force large enough to mask subtle changes in slope conductance caused by CHS. In summary, CHS-induced reduction of hslo channel Po occurred without major changes in unitary conductance, as in more complex systems. Thus, as for EtOH, the channel pore-forming hslo subunits reconstituted into a minimal bilayer system are sufficient for CHS modulation of BKCa channel activity.
Role of Phosphatidylserine for the Actions of EtOH and CHS on hslo Channels. We next tested the role of POPS in the modulation of channel activity by EtOH by casting bilayers from 100% POPE. The amount of PS is altered in synaptic plasma membranes after EtOH treatment (Sun and Sun, 1985). Removal of POPS may alter lateral domain formation in the bilayer resulting from POPE-POPS and POPS-POPS headgroup interactions. Headgroup interactions and lateral domains may influence EtOH action (Treistman et al., 1987; Jorgensen et al., 1993), as well as CHS miscibility in the membrane (see Discussion).
POPS carries a net negative charge at physiological pH, at which our experiments were performed. Negative surface charge promotes adsorption of cations to the membrane (McLaughlin et al., 1981), influencing BKCa channel conductance and Po by the accumulation of K+ and Ca2+ at the mouth of the channel (Moczydlowski et al., 1985; Turnheim et al., 1999). As expected, hslo channels displayed changes in slope conductance consistent with the loss of negative charge carried by POPS: 323.3 ± 5.8 versus 281.8 ± 11.1 pS (p < 0.004), in POPE/POPS [3:1 (w/w)] versus POPE bilayers, respectively (data obtained at 0 mV and 50 μM [Ca2+]free). In addition, hslo channel steady-state activity followed a monotonic function of the POPS concentration in the bilayer (Fig. 4). It is important to note, however, that CHS inhibition of hslo activity cannot be attributed solely to a dilution of the POPS concentration caused by addition of the sterol to the lipid mixture. Although the addition of 32 mol% CHS to the POPE/POPS (3:1) mixtures causes the POPS concentration to drop from 25 to 20% of total lipid weight, the decrease in Po caused by CHS (Fig. 3B) is far greater than the corresponding dilution of POPS (Fig. 4).
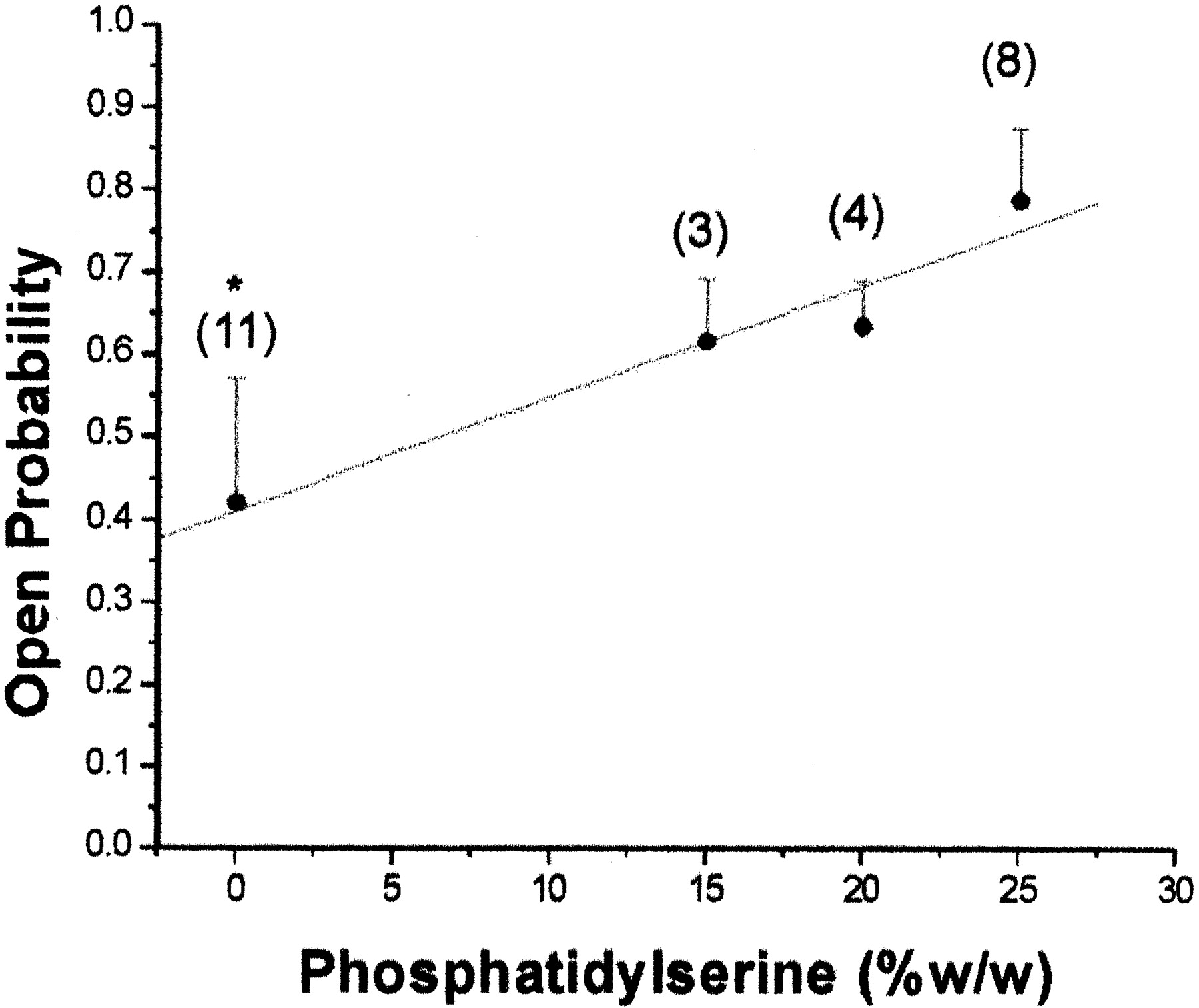
Dilution of POPS in the binary POPE/POPS mixture influences hslo Po in a concentration dependent manner. Po values were calculated from hslo channels reconstituted into bilayers with varying weight percentages of POPS in a binary mixture with POPE. Current records were obtained at 0 mV and [Ca2+]free ≈ 50 μM on the intracellular side of the bilayer. Data were low-pass filtered at 1 kHz and sampled at 10 kHz. Po values were obtained from continuous recording (see Materials and Methods). The number of experiments performed for each condition is shown in parentheses above each point. *, significantly different from POPE/POPS [3:1, or 25% w/w] values (p < 0.05).
Figure 5A shows that application of either 50 or 100 mM EtOH fails to significantly increase the NPo of hslo channels incorporated into pure POPE bilayers. The average NPo during EtOH exposure reached 1.46 ± 0.63 of control (n = 11) at 50 mM and 0.60 ± 0.09 of control (n = 4) at 100 mM, a concentration near lethal blood levels in naïve mammals (Diamond, 1992). For comparison, potentiation by 50 mM ethanol reached (values compared with control) 5.2 ± 1.5 in 75:25 (w/w) POPE/POPS, 3.3 ± 1.3 in 85:15 (w/w) POPE/POPS, and 1.5 ± 0.6 in 100% POPE, suggesting that the alcohol response may be a monotonic function of POPS concentration in the bilayer.

Both ethanol and cholesterol fail to markedly modify the activity of hslo channels incorporated into a single (POPE) bilayer. A, EtOH concentrations (50 and 100 mM) that increase hslo channel activity in POPE/POPS bilayers fail to potentiate activity in 100% POPE bilayers. Ratios of NPo values obtained in the presence and absence of EtOH (left, 50 mM; right, 100 mM) are shown in a scatter graph, where each data point represents an individual bilayer (n). Mean ± S.E.M. of data are shown at the top of the graph. The dotted line highlights the point at which NPo EtOH/NPo control = 1. The potential at the cis side of the bilayer was set between -20 and +60 mV and free [Ca2+]ic ≈ 10 μM. Data were low-pass filtered at 1 kHz and sampled at 10 kHz. NPo values were obtained from continuous recording (see Materials and Methods). B, significant inhibition of hslo activity by CHS in POPE/POPS bilayers is not observed when CHS action is evaluated in POPE bilayers. Scatter graph of hslo NPo from POPE bilayers in the absence (left) and presence of 23 mol% (middle) or 32 mol% (right) CHS. Data points represent individual bilayers. Mean ± S.E.M. of data are shown at the top of the graph. The potential at the cis side of the bilayer was set at 0 mV and free [Ca2+]ic ≈ 50 μM. Data were low-pass filtered at 1 kHz and sampled at 10 kHz. NPo values were obtained from continuous recording (see Materials and Methods).
Next, we determined whether the absence of POPS from the planar bilayer affected CHS inhibition of hslo channels. Figure 5B demonstrates that, despite a large variability in Po (as with native BKCa channels; Chu et al., 1998), average hslo activity in POPE + 23 mol% CHS (p > 0.3) or POPE + 33 mol% CHS (p > 0.2) was not significantly different from that in pure POPE bilayers. Thus, CHS-induced hslo inhibition, like EtOH-induced hslo activation, is significantly impaired in a single species POPE bilayer, suggesting that a negatively charged headgroup or other structural requirement associated with the presence of POPS in the bilayer favors CHS and EtOH modulation of hslo channel function (see Discussion).
Cholesterol and Ethanol Target Both Common and Distinct hslo Channel Dwelling States. Modulation of hslo channel function by CHS and EtOH is characterized by striking similarities: both agents modify channel Po with minor, if any, modification of conductance. Furthermore, CHS and EtOH effects on hslo baseline activity are determined by the amount of POPS in the bilayer (Fig. 5). Thus, we considered the possibility that CHS and EtOH actions on hslo channels are mediated by a single, common mechanism. In this case, we might expect channel dwell-time histograms in the presence of each agent to show a mirroring profile of actions.
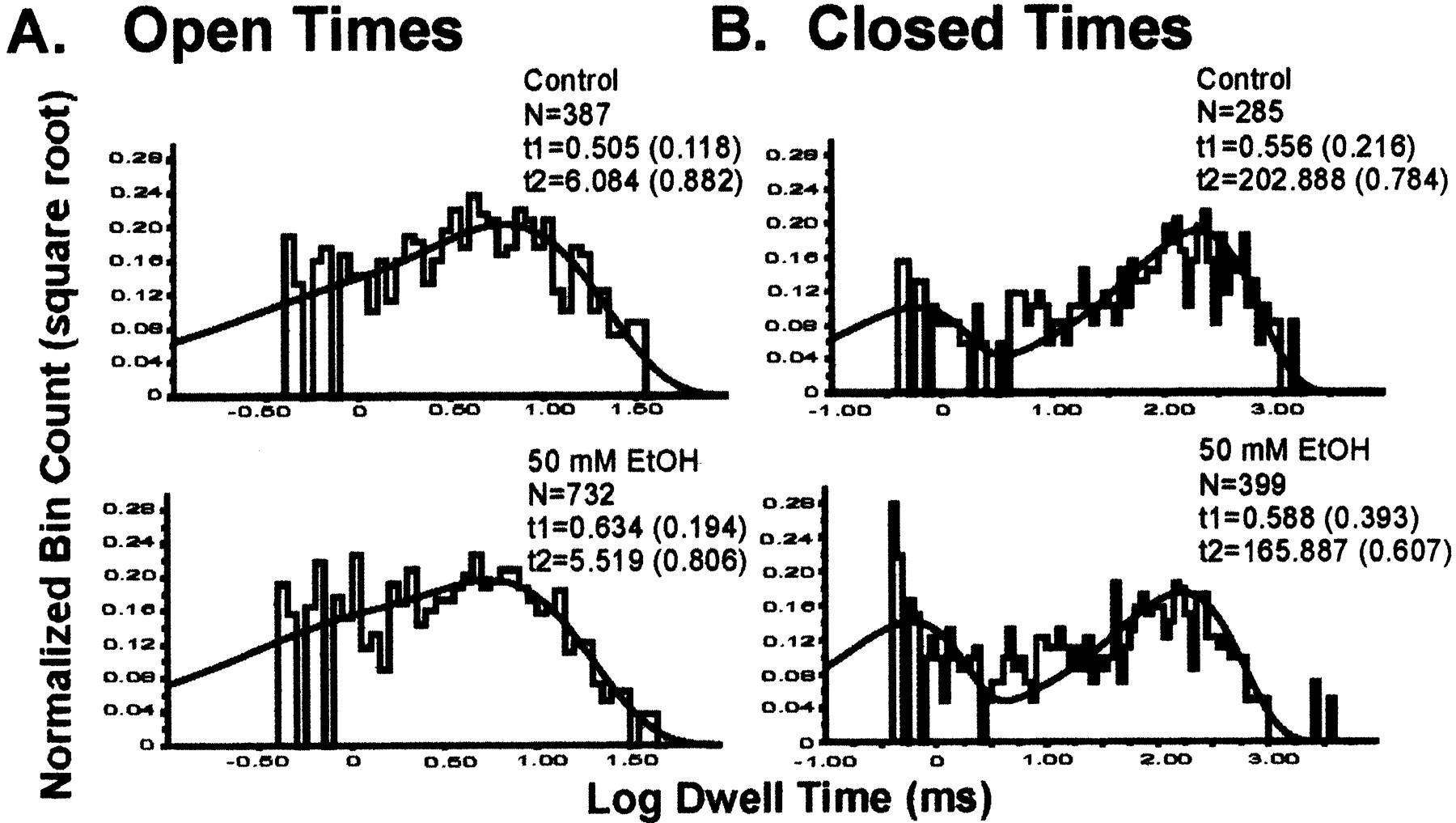
Dwell times in the absence and presence of 50 mM EtOH were evaluated in several POPE/POPS (3:1) bilayers (V = 0–20 mV; [Ca2+]free = 15 μM; n = 3). In both the absence and presence of EtOH, the open times distribution could be well fitted with a double exponential (Fig. 6A), indicating the existence of at least two open states. Although 50 mM EtOH characteristically increases channel NPo to ∼5-fold of control values, on average, it mildly increased the duration of short and long openings and slightly shifted the channel population from long to short openings. These drug-induced changes in open channel populations result only in a minor change in mean open time. Thus, a major increase in hslo steady-state activity induced by acute EtOH could be obtained in the absence of a significant increase in channel mean open time.

Representative dwell-time distributions of hslo channel activity reconstituted into POPE/POPS 3:1 (w/w) bilayers in the absence (top) and presence (bottom) of 50 mM EtOH, which show the targeting of open (A) and closed (B) times by the drug that lead to potentiation of channel activity. A, open time distributions in both the absence and the presence of EtOH were well fitted with a double exponential. EtOH slightly increased the duration of short openings while slightly shifting the channel population from long to short openings. B, closed time distributions in both absence and presence of EtOH were also well fitted with a double exponential. EtOH decreased the duration of long closures and markedly shifted the channel population from long to short closures. The potential at the cis side of the bilayer was set between 0 and 20 mV and free [Ca2+]ic ≈ 15 μM. In both A and B are shown the total number of events (N), the duration of each particular component (t, in milliseconds), and the relative contribution of each component to the total fit (in parentheses). The solid line represents the composite fit. Data were low pass filtered at 3 kHz and sampled at 10 kHz. The following are mean ± S.E.M. values (n = 3) for the time constants (in milliseconds) and the fractional contribution of each component to the total fit, respectively: control open states, τfast = 0.768 ± 0.142 and 0.150 ± 0.016, τslow = 7.917 ± 0.962 and 0.850 ± 0.016; open states in EtOH, τfast = 0.963 ± 0.178 and 0.176 ± 0.014, τslow = 8.643 ± 1.572 and 0.824 ± 0.014; control closed states, τfast = 0.712 ± 0.178 and 0.206 ± 0.008, τslow = 218.671 ± 8.671 and 0.794 ± 0.008; and closed states in EtOH, τfast = 0.588 ± 0.015 and 0.335 ± 0.046, τslow = 151.996 ± 7.811 and 0.659 ± 0.049.
The closed times distribution was also well fitted with a double exponential function, in both the absence and presence of EtOH (Fig. 6B), indicating the existence of at least two closed states. Two actions of EtOH are evident: a decrease in the average duration of long closures, and a marked shift in the closed channel population from long to brief events. EtOH-induced changes in the closed times distribution result in a significant change in mean closed time (∼60% of control), the major contributor to EtOH-induced increase in channel Po. In summary, EtOH markedly increases hslo steady-state activity by producing a marked reduction in the average duration of channel long closures and their relative contribution to the total time spent in closed states, without causing a major change in the channel mean open time.
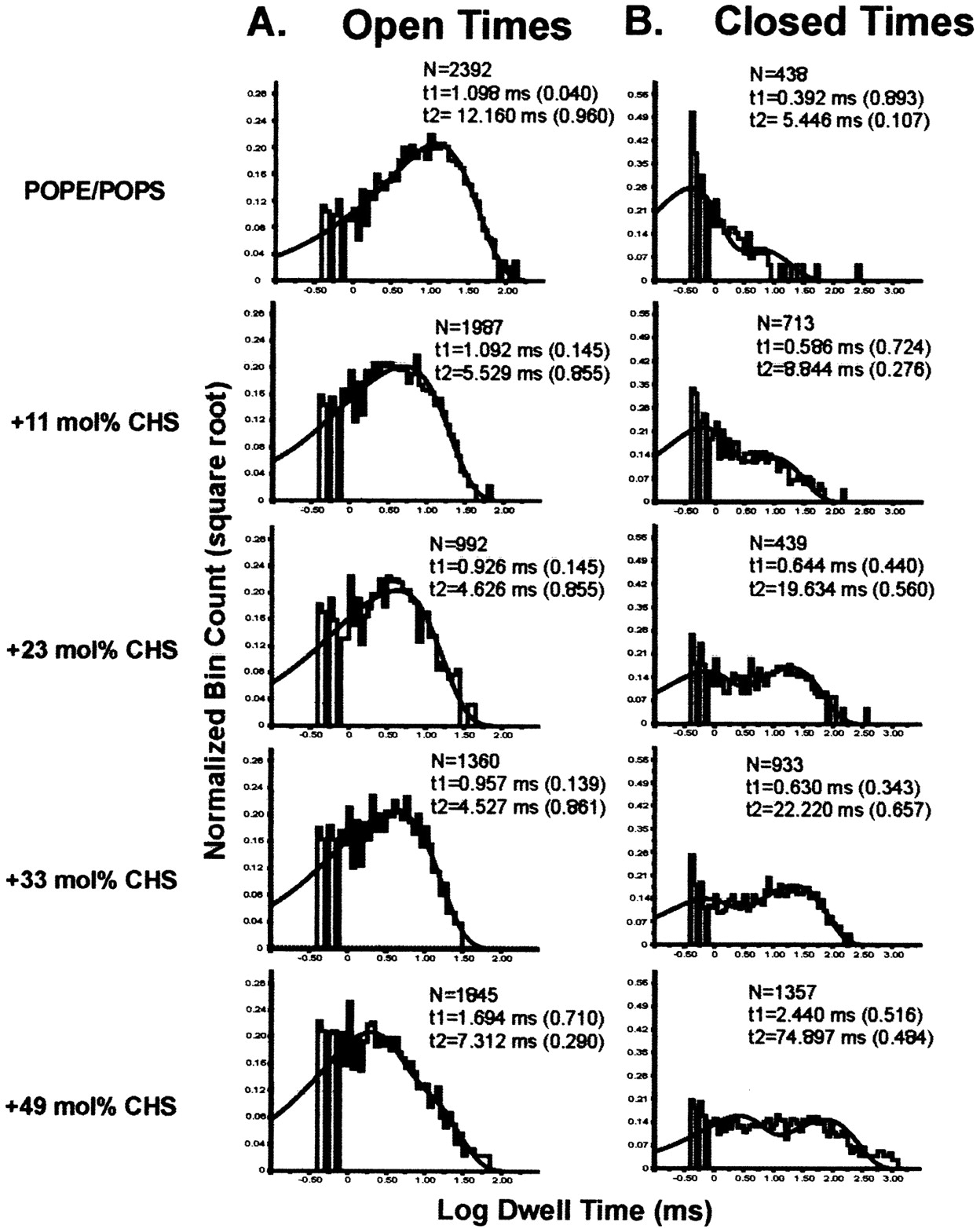
Were CHS and EtOH having opposite effects on channel Po solely by targeting common kinetic states in opposite manner, we might expect a profile of changes in open and closed times distributions by CHS mirroring those caused by EtOH. This complementarity was indeed present, but each of these agents also had unique effects not mirrored by the other. The dwell-time data for hslo channels in POPE/POPS (3:1) bilayers with increasing amounts of CHS (n = 2–8 for each CHS concentration) were obtained under identical conditions of voltage (0 mV) and [Ca2+]free (50 μM). The closed times distributions in the presence and absence of CHS could be well fitted with two exponentials in the representative example shown (Fig. 7B), which is particularly useful for a comparison with the closed time distribution in the presence of EtOH (Fig. 6B, right). These data demonstrate that CHS increases the average duration of longer closures and shifts the closed channel population from short to longer closures, these two changes being a mirror of EtOH actions that lead to channel activation. However, CHS also increases the average duration of short closures, an action not mirrored by EtOH.

Representative dwell-time distributions of hslo channel activity reconstituted into POPE/POPS 3:1 (w/w) bilayers in the absence (top) and presence of increasing amounts of cholesterol. A, open time distributions in both absence and presence of CHS were well fitted with a double exponential. Increases in CHS content up to 23 mol% progressively decrease the average duration of long openings. B, closed time distributions in both absence and presence of CHS were also well fitted with a double exponential function. Increases in CHS content (11–49 mol%) progressively shift the channel population from short to long closures and increase the duration of long closures. The potential at the cis side of the bilayer was set at 0 mV and free [Ca2+]ic ≈ 50 μM. In both A and B are shown the total number of events (N), the duration of each particular component (t, in milliseconds) and the relative contribution of each component to the total fit (in parentheses). The solid line represents the composite fit. Data were low pass filtered at 3 kHz and sampled at 10 kHz. The average values for the time constants (in milliseconds) and the fractional contribution of each component to the total fit for the open states were as follows: POPE/POPS, τfast = 1.61 ± 0.393 and 0.139 ± 0.041, τslow = 11.765 ± 2.111 and 0.861 ± 0.041 (n = 8); POPE/POPS + 10 mol% CHS, τfast = 1.557 ± 0.356 and 0.185 ± 0.09, τslow = 5.773 ± 0.333 and 0.815 ± 0.09 (n = 5); POPE/POPS + 23 mol% CHS, τfast = 1.242 ± 0.309 and 0.239 ± 0.073, τslow = 4.194 ± 0.77 and 0.704 ± 0.108 (n = 7); POPE/POPS + 32 mol% CHS, τfast = 1.046 ± 0.218 and 0.22 ± 0.079, τslow = 4.628 ± 0.246 and 0.779 ± 0.079 (n = 4); and POPE/POPS + 49 mol% CHS, τfast = 1.066 and 0.28, τslow = 6.981 and 0.72 (n = 2). The average values for the time constants (msec) and fractional contribution of each component of the distribution to the total fit for the closed states were as follows: POPE/POPS, τfast = 0.436 ± 0.094 and 0.835 ± 0.033, τslow = 12.994 ± 5.904 and 0.165 ± 0.033 (n = 8); POPE/POPS + 10 mol% CHS, τfast = 0.694 ± 0.053 and 0.671 ± 0.07, τslow = 19.152 ± 4.341 and 0.329 ± 0.07 (n = 5); POPE/POPS + 23 mol% CHS, τfast = 0.587 ± 0.113 and 0.533 ± 0.078, τslow = 21.022 ± 4.11 and 0.467 ± 0.077 (n = 7); POPE/POPS + 32 mol% CHS, τfast = 1.155 ± 0.126 and 0.566 ± 0.079, τslow = 42.971 ± 13.052 and 0.434 ± 0.079 (n = 4); and POPE/POPS + 49 mol% CHS, τfast = 1.462 and 0.67, τslow = 146.06 and 0.33 (n = 2).
In addition, representative open times distributions (Fig. 7A), fitted by double exponential functions, indicate that CHS at all concentrations tested decreased the average duration of long openings, a dwell state basically unmodified by EtOH. Figure 7A also shows that CHS produces a minor shift from long to short openings, this shift being another contributor to the decrease in mean open time induced by CHS. In summary, overall antagonism between CHS and EtOH on hslo steady-state activity results from the targeting of both common and distinct channel dwelling states by these modulators, probably reflecting their common and distinct sites of action (see Discussion).
Discussion
The role of the lipid environment in the function of embedded ion channel proteins and their drug sensitivity is difficult to assess in complex biological systems but is more approachable in reduced preparations such as with cloned channels incorporated into planar lipid bilayers. Our data demonstrate the feasibility of this approach, because hslo channels in POPE and POPE/POPS bilayers retain basic BKCa channel characteristics and respond to the change in bulk phospholipid composition. Of course, this reductionist approach ignores many of the potential interactions present in a rich, heterogeneous natural membrane. However, the model used successfully addresses the specific questions that are being asked.
We demonstrated that both CHS and EtOH modulation of basal hslo activity are dramatically impaired in the absence of POPS in the bilayer. The fact that these agents modulate hslo channel activity in POPE/POPS, but fail to do so in pure POPE bilayers, could be attributed to the loss of headgroup negative charge, differing headgroup size, and/or altered headgroup interactions. Neutral phosphatidylethanolamine bilayers have a high propensity to transition from the lamellar phase into the HII (inverted hexagonal) phase, a transition directly attenuated by negative membrane surface charge carried by PS (Lewis and McElhaney, 2000). Although this transition for POPE generally occurs at higher temperatures (70°C; Epand, 1985) than those used in this study, we cannot rule out that such a tendency may mask or alter the actions of CHS or EtOH under the exact conditions used in our system. Both PE and CHS are nonlamellar phase-preferring lipids, which can presumably increase curvature stress when incorporated into a lamellar bilayers and, eventually, modify channel function (Lundbaek et al., 1996). It is possible that a pure POPE bilayer with a high initial degree of curvature stress masks sterol modulation of this parameter. The inability of CHS to modify hslo function when added to POPE bilayers might also reflect a relatively low miscibility of CHS in pure PE (McMullen et al., 1999), perhaps alleviated by the headgroup structure and charge in the POPE/POPS mixture.
In both cell culture and animal models of chronic EtOH exposure, alterations in both content (Chin et al., 1978; Omodeo-Sale et al., 1995) and distribution (Wood et al., 1990) of membrane CHS have been reported. These alterations might represent a compensatory response (i.e., “tolerance”) to counteract the effects of EtOH on relevant targets, such as defined ion channel populations. Here, we demonstrate that increases in the CHS content of POPE/POPS bilayers, indeed, reduce EtOH potentiation of hslo channel Po.
Increases in bilayer/membrane CHS content diminish the lipid/membrane partition coefficient of a variety of small anesthetics, such as halothane (Lechleiter et al., 1986), un-charged pentobarbitone (Miller and Yu, 1977), and benzyl alcohol (Colley and Metcalfe, 1972). Isothermal titration calorimetry data show that EtOH partitioning into phosphatidylcholine (PC) liposomes is also reduced by CHS, when present in the bilayer at concentrations >10 mol% (Trandum et al., 2000). Consistent with these findings, Fig. 2 demonstrates that CHS effects on EtOH sensitivity of hslo channels is largely absent at 10 mol% CHS but evident at concentrations above 23 mol%.
The effect of CHS on EtOH partitioning may be explained by bilayer phase behavior. Isothermal titration calorimetry (Trandum et al., 1999) and computer stimulation (Jorgensen et al., 1993) studies strongly suggest that EtOH preferentially partitions into bilayers at the interfaces between the gel and the liquid crystalline domains that form as the bilayer approaches the transition temperature. High bilayer CHS concentrations abolish the gel-to-liquid crystalline transition, causing the bilayer to exist in a liquid-ordered state (Trandum et al., 2000). The resulting disappearance of the gel/liquid crystalline interfaces preferentially targeted by EtOH would serve to decrease its partitioning into the membrane. In fact, X-ray diffraction studies of POPE/POPS mixtures suggest that at 25°C, multilayer samples exist as a combination of both gel and liquid crystalline phases (Chang et al., 1995). The coexistence of these lateral domains would support the partitioning of EtOH into this lipid mixture. We show here that channels are sensitive to EtOH in POPE/POPS bilayers (Fig. 1). However, in bilayers containing >20 mol% CHS that lack these domain interfaces (Chang et al., 1995), EtOH potentiation of hslo activity is markedly reduced (Fig. 2). Thus, our data are consistent with a CHS-induced reduction of EtOH partitioning in the bilayer. In our system, however, the hydrocarbon interior of the bilayer probably contains contaminant decane in equilibrium with that in the torus (Gruen, 1981), so we cannot rule out some contribution of this solvent to the phase behavior of the bilayers used in our study. The manipulation of parameters such as temperature and acyl chain saturation will yield further insight into the importance of bilayer phase behavior on BKCa ethanol sensitivity.
Further evidence that CHS reduces EtOH interaction with the membrane comes from NMR spectroscopy data demonstrating that EtOH resides at the lipid-water interface in phospholipid bilayers. The carbonyl groups in the glycerol backbone are specifically favored hydrogen bonding sites for EtOH (Barry and Gawrisch, 1994), this binding being decreased by increasing amounts of CHS (Barry and Gawrisch, 1995). Because CHS interacts with PC at the same carbonyl groups in the glycerol backbone (Worcester and Franks, 1976), this was interpreted as CHS directly competing for EtOH's favored binding sites (Barry and Gawrisch, 1995). CHS location at the phospholipid carbonyl groups would also increase the packing density of the phospholipids and antagonize the increase in acyl chain motion (“disordering”) introduced by EtOH (Sun and Sun, 1985), which may contribute to functional antagonism on hslo kinetics (see below).
Apart from effects on EtOH partitioning, CHS may directly antagonize the action of the drug on the bilayer or the hslo channel itself. If CHS and EtOH act through a single, common mechanism, we might expect them to exert reciprocal actions on common dwell states of the channel. Indeed, EtOH and CHS produce a mirrored shift between the long and short channel closed states and have opposite actions on the mean duration of long closures. This suggests that CHS and EtOH share a common target on the channel protein or in the phospholipid bilayer, which is important in determining the stability of the channel closed state(s). A similar increase in the average duration of native BKCa channel long closures after CHS enrichment of myocyte membranes has been reported (Bolotina et al., 1989). Channel Po was approximately halved, coincident with a similar decrease in the rotational diffusion coefficient of DPH (Bolotina et al., 1989), indicating that a decrease in both rate and range of motion of phospholipid acyl chains accompanies the reduction in channel activity. Spin-labeling experiments also demonstrate that CHS increases, whereas EtOH decreases bilayer order (Chin and Goldstein, 1981). Interestingly, the magnitude of CHS and EtOH effects on multilayer PC vesicle order are similar to those in brain synaptosomal membranes (Chin and Goldstein, 1981). Thus, we postulate that opposite actions of CHS and EtOH on acyl chain order may underlie or, at least, contribute to CHS and EtOH opposing effects on common channel dwell states, such as the long closed state.
In addition to their common modulation of channel long closures, CHS and EtOH exhibit individual effects on channel dwell times, which contribute to their opposite actions on Po. These distinct effects on channel dwell states may represent independent actions of these modulators on specific bilayer characteristics that modify channel function. For example, EtOH increases the rate of phospholipid desorption, displacing water from the hydrogen-bonded network of water molecules in the hydration layer, whereas CHS has little, if any, effect on phospholipid desorption (Slater et al., 1993). Changes in phospholipid desorption alter not only lipid-lipid interactions but also protein-lipid interactions, with eventual modification of ion channel function. Interestingly, EtOH desorption is more marked in PS than PE (Slater et al., 1993), consistent with the EtOH activation of hslo channels (Figs. 1 and 5).
Monolayer or bilayer properties modified by CHS, but not EtOH, include a broadening and eventual elimination of the gel-to-liquid crystalline phase transition, a decrease in the cross-sectional area occupied by the phospholipid in the liquid crystalline state, increases in both bilayer thickness and mechanical strength, and increases in the lateral stress and stiffness of the phospholipid monolayer or bilayer in the physiologically relevant fluid phase (McMullen et al., 1999; Nielsen et al., 1999). In particular, changes in lateral stress and bilayer stiffness have been causally related to modification of ion channel function. Cholesterol and other compounds promoting negative monolayer curvature increase stiffness and decrease channel activity, whereas compounds promoting positive monolayer curvature have opposite effects on both stiffness and channel activity (Lundbaek et al., 1996; Bezrukov et al., 1998). Furthermore, CHS inhibition of native BKCa channels in PE/PS bilayers has been linked to an increase in bilayer lateral stress caused by the presence of the steroid (Chang et al., 1995). A major consequence of increases in lateral stress is a reduction in the activation energy for the transition from open to closed state(s). This reduces the average duration of long openings, as we report here (Fig. 7A). Thus, we postulate that the distinct decrease in the average duration of long openings observed with CHS, an effect not mirrored by EtOH, may be related to the increase in lateral stress caused by the steroid.
In our demonstration of CHS blunting of alcohol potentiation of hslo activity we used 50 mM EtOH, close to legal intoxication (∼20 mM) and below lethal blood levels in naïve subjects (>90 mM), and a range of CHS content similar to that found in cell membranes (5.6–44 mol%). Thus, modification of hslo function by EtOH may depend on the membrane CHS content where the channel resides. Membrane CHS content and/or distribution might contribute to differential EtOH sensitivity of BKCa in different cell types and in similar channel subtypes from different neuronal domains, as in supraoptic neurons (Dopico et al., 1999). Present results might also help to explain recent findings that rats undergoing extending feeding with EtOH display not only reduced BKCa current density in neurohypophysial terminals but also reduced BKCa sensitivity to acute EtOH (Knott et al., 2002).
Our findings suggest that manipulation of membrane lipid composition may represent a mechanism for plasticity responsible for alteration of channel basal Po as well as sensitivity to small amphiphiles such as EtOH. This hypothesis becomes particularly attractive when coupled with data demonstrating alterations in lipid composition after EtOH exposure, and more generally with the emerging theme of lipid domains. Here, we demonstrate that alcohol action on the activity of a human neuronal ion channel depends on the lipid environment of the channel protein.
Acknowledgments
We thank Andy Wilson for excellent technical assistance.
Footnotes
-
This study was supported by National Institute on Alcohol Abuse and Alcoholism grants AA12054 and AA11560 and predoctoral fellowship AA05548.
-
Preliminary data were presented at the 46th Annual Meeting of the Biophysical Society; 2002 Feb 23–27; San Francisco, CA and published in abstract form [Crowley JJ, Treistman SN, and Dopico AM (2002) Phospholipid and cholesterol modulation of hslo channel activity and ethanol sensitivity in lipid bilayers. Biophys J 82:586a].
-
ABBREVIATIONS: CHS, cholesterol; BKCa, large conductance Ca2+-activated K+ channel; EtOH, ethanol; POPE, 1-palmitoyl-2-oleoyl-phosphatidylethanolamine; POPS, 1-palmitoyl-2-oleoyl-phosphatidylserine; HEK, human embryonic kidney; MOPS, 3-(N-morpholino)propanesulfonic acid; HEDTA, N-(2-hydroxyethyl) ethylene-diaminetriacetic acid; ANOVA, analysis of variance; PC, phosphatidylcholine; Po, open probability; NPo, steady-state activity.
- Received January 3, 2003.
- Accepted April 24, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}