


Abstract
Hetero-oligomerization among G protein-coupled receptors has been proposed to contribute to signal integration. Because sensory neuron-specific receptors (SNSRs) and the opioid receptors (OR) share a common ligand, the bovine adrenal medulla peptide (BAM) 22, and have opposite effects on pain modulation, we investigated the possible consequences of δOR/SNSR-4 hetero-oligomerization on the signaling properties of both receptor subtypes. Bioluminescence resonance energy transfer revealed that the human δOR has similar propensity to homo-oligomerize and to form hetero-oligomers with human SNSR-4 when coexpressed in human embryonic kidney 293 cells. The hetero-oligomerization leads to a receptor form displaying unique functional properties. Individual activation of either δOR or SNSR-4 in cells coexpressing the two receptors led to the modulation of their respective signaling pathways; inhibition of adenylyl cyclase and activation of phospholipase C, respectively. In contrast, the δOR/SNSR-4 bivalent agonist BAM22, which could activate each receptor expressed individually, fully activated the SNSR-4-dependent phospholipase C but did not promote δOR-mediated inhibition of adenylyl cyclase in δOR/SNSR-4-coexpressing cells. Likewise, concomitant activation of the δOR/SNSR-4 hetero-oligomer by selective δOR and SNSR-4 agonists promoted SNSR-4 but not δOR signaling, revealing an agonist-dependent dominant-negative effect of SNSR-4 on δOR signaling. Furthermore, the δOR selective antagonist naltrexone trans-inhibited the SNSR-4-promoted phospholipase C activation mediated by BAM22 but not by the SNSR-4-selective agonists, suggesting a bivalent binding mode of BAM22 to the δOR/SNSR-4 hetero-oligomer. The observation that BAM22 inhibited the Leu-enkephalin-promoted cAMP inhibition in rat dorsal root ganglia neurons supports the potential physiological implication of such regulatory mechanism.
Sensory neuron-specific G protein-coupled receptors (SNSRs) have recently been identified as a new family of human G protein-coupled receptors (GPCRs), consisting of seven receptor subtypes (SNSR-1 to -7), exclusively expressed in dorsal root ganglia cells (Dong et al., 2001; Lembo et al., 2002). Based on their unique localization, it was predicted that they could play a role in the control of pain perception in response to external stimuli received by peripheral somato- or chemosensory neurons. In agreement with this prediction, activation of SNSRs elicited nociceptive responses in rats (Grazzini et al., 2004; Hong et al., 2004). It is noteworthy that the proposed endogenous ligands for SNSRs originate from a precursor molecule, proenkephalin, that also gives rise to the antinociceptive peptides Leu- and Met-enkephalins. In addition to enkephalins, proteolytic cleavage of proenkephalin by prohormone convertases produces a family of biologically active compounds named bovine adrenal medulla (BAM) peptides that bind SNSRs with high affinity (Lipman, 1996; Lembo et al., 2002). One of these peptides, BAM22, has the ability to bind and activate both SNSR-4 and the δ opioid receptor (OR) (Lembo et al., 2002). Its extreme N terminus harbors an enkephalin motif (YGGF) that is responsible for high-affinity binding to δOR (Quirion and Weiss, 1983; Swain et al., 1994), whereas the C-terminal portion of BAM22 binds and activates SNSR-4 (Lembo et al., 2002). Removal of the N-terminal tyrosine residue (BAM2-22) is sufficient to generate a selective SNSR-4 agonist that cannot bind to δOR. The SNSR-4 agonistic activity of the peptide is preserved following removal of up to 12 residues, as indicated by the full SNSR activity of the shorter C-terminal fragments BAM8-22 and BAM13-22. The N-terminal fragments BAM1-12 and BAM1-20, for their part, exhibit selective agonistic activity for the δOR (Lembo et al., 2002). It is noteworthy that the fragments BAM1-12 and BAM8-22 as well as the entire BAM22 peptide can be found endogenously (Hollt et al., 1982; Dores et al., 1990), indicating that coexisting peptides originating from the same precursor can either bind selectively to SNSR-4 or δOR or to both receptors at the same time.
In addition to their ligand-sharing properties, the overlapping localization of SNSR-4 and δOR within the same dorsal root ganglia compartments (lamina I and II) raises the possibility of functional interactions between the two receptors. This is particularly relevant when considering their opposite roles in pain perception (SNSR-4 being pronociceptive and δOR antinociceptive) (Puttfarcken et al., 1988; Grazzini et al., 2004).
Interactions between subtypes of the same or even among distinct GPCR families have been described at different molecular levels. Receptor-oligomerization (Salahpour et al., 2000) as well as heterologous cross-talk between signaling pathways (Salahpour et al., 2000) have been shown to contribute to signal integration. Direct receptor-receptor interactions between the β2-adrenergic receptor (Jordan et al., 2001), the μOR (George et al., 2000; Martin and Prather, 2001), or the κOR (Jordan and Devi, 1999) have been shown to alter agonist-promoted receptor endocytosis or ligand binding properties of the δOR. Signaling efficacy of the δOR can also be regulated by a heterologous cross-talk with signaling pathways activated by other receptors or drugs. For example, specific activation of protein kinase C by phorbol ester has been shown to induce phosphorylation of the δOR, which in turn results in the desensitization and internalization of the receptor (Xiang et al., 2001).
To date, no functional interaction between members of the OR and SNSR families have been reported. The present study was therefore initiated to examine the possibility that δOR and SNSR-4 may interact when expressed in the same cells. Here, using bioluminescence resonance energy transfer (BRET) approaches, we report that the δOR exhibits similar propensity to homo-oligomerize as to form hetero-oligomers with SNSR-4. The functional consequences of such hetero-oligomerization were then characterized in HEK293 cells stably expressing the human δOR and SNSR-4. We found that SNSR-4 signaling promoted by the mixed, bivalent agonist BAM22 could be trans-inhibited by the δOR-selective antagonist naltrexone. Moreover, simultaneous activation of the δOR and SNSR-4 by BAM22 or by various combinations of selective agonists for each receptor dramatically decreased the efficacy of δOR agonist to inhibit adenylyl cyclase activity, suggesting a dominant-negative effect of SNSR-4 on the δOR activity. Together, these results demonstrate that hetero-oligomerization between δOR and SNSR-4 generates a novel signaling unit with a unique pharmacological profile that may contribute to the signal integration involved in pain perception.
Materials and Methods
Materials. Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum, penicillin/streptomycin, l-glutamine, and G418 (Geneticin) were obtained from Wisent Inc. (St-Bruno, QC, Canada). Neurobasal medium, B27 supplement, l-glutamine, trypsin HEPES buffer, and Zeocin were purchased from Invitrogen (Carlsbad, CA). 3-Isobutyl-1-methylxanthine (IBMX), Leu-enkephalin (Leu-enk), naltrexone, and [d-Pen2,d-Pen5]enkephalin (DPDPE) were purchased from Sigma-Aldrich (St. Louis, MO). FuGENE 6 was obtained from Roche Diagnostics (Laval, QC, Canada). Coelenterazine H was from Molecular Probes (Eugene, OR), whereas DeepBlueC coelenterazine, [3H]adenine, [myo-3H]inositol, [3H]naloxone, and [3H]DPDPE were from PerkinElmer Life and Analytical Sciences (Boston, MA). cAMP antiserum and 125I-cAMP were purchased from Biomedical Technologies (Stoughton, MA). Nerve growth factor was purchased from Harlan (Madison, WI). BAM22 was purchased from the American Peptide Co., Inc. (Sunnyvale, CA), whereas BAM8-22 was from Tocris Cookson, Inc. (Ellisville, MO). BAM1-12 and BAM2-22 were kindly provided by the AstraZeneca Research Center (Montréal, QC, Canada).
Eukaryotic Expression Vectors. For BRET experiments, expression vectors encoding the human δOR and SNSR-4 fused to either the green fluorescent protein (GFP2) (where GFP2 is a variant of the wild-type GFP containing the following mutations: P64L, Y100F, S108T, M141L, S147P, S202F, I219V, and H213L) or Renilla reniformis luciferase (Rluc) were generated as follows: PCR fragments containing the entire coding sequences excluding the stop codons of the human δOR or of the human SNSR-4 were subcloned into the humanized pGFP2-N2 vector (PerkinElmer Life and Analytical Sciences) in a way that fused the 3′ end of the receptors onto the 5′ end of the GFP2-cDNA using the restriction sites HinDIII and BamHI. This resulted in an in frame fusion of the receptors with GFP2 separated by a seven-amino acids linker. The pcDNA3.1-δOR-Rluc cDNA was obtained by subcloning a PCR fragment containing the human δOR into the pcDNA3.1-Rluc vector using the restriction sites HinDIII and BamHI in a way that fused the receptor sequence onto the 5′ end of Rluc. This resulted in an in frame fusion of the receptor with Rluc separated by a 4-amino acid linker. The pcDNA3.1-SNSR-4-Rluc cDNA containing the human SNSR-4 fused to the 5′ end of Rluc with a 3-amino acid linker was kindly provided by AstraZeneca Research Center. The pcDNA3.1 GABABR1b-Rluc construct was generated as described previously (Villemure et al., 2005).
The cell lines stably expressing δOR, SNSR-4, or the two receptors together were generated using the following cDNAs: The pcDNA4-SNSR-4 was created by subcloning a PCR fragment containing the entire coding sequence, including the stop codon of the human SNSR-4 into the pcDNA4 vector (Invitrogen) using the EcoRI and EcoRV restriction sites. The pcDNA3.1-δOR-FLAG construct was generated as described previously (Petaja-Repo et al., 2000).
Cell Culture and Transfection. HEK293 cells were cultured in DMEM supplemented with 5% fetal bovine serum and 2 nM l-glutamine. For transient expression of recombinant proteins, cells were seeded at a density of 2 × 105 cells in a six-well dish, cultured for 24 h, and then transfected with the appropriate vectors using FuGENE 6 reagent according to the manufacturer's protocol. Forty-eight hours after transfection, cells were washed twice with phosphate-buffered saline, detached with 5 mM EDTA in phosphate-buffered saline, and used immediately. HEK293 cell clones stably expressing human SNSR-4 (HEK293-SNSR-4) were obtained by selecting cells (50 μg/ml zeocin) transfected with the pcDNA4-SNSR-4 construct. The expression of SNSR-4 was controlled by measuring BAM22-induced accumulation of total phosphoinositols (IPn). HEK293 cells stably expressing the δOR-FLAG fusion protein (HEK293-δOR) were obtained as described previously (Petaja-Repo et al., 2000) by transfection of the pcDNA3.1-δOR-FLAG construct and selection with 400 μg/ml G418. To obtain HEK293 cells stably coexpressing SNSR-4 and the human δOR (HEK293-δOR/SNSR-4), a cell clone already expressing the δOR was transfected with the pcDNA4-SNSR-4 construct. After selection with 50 μg/ml zeocin, in the presence of 250 μg/ml G418, cell clones were isolated and coexpression of δOR-FLAG and SNSR-4 monitored by ligand binding and BAM22-promoted IPn accumulation, respectively.
Dorsal Root Ganglia Cell Culture and Stimulations. Dorsal root ganglia were dissected from embryonic day 16 embryos of Long-Evans rats. Cells were obtained by mechanical dissociation in 0.25% trypsin and plated on poly-l-lysine/laminin coated 24-well plates (∼3 ganglia/well). Cells were grown in “culturing medium” consisting of neurobasal medium supplemented with B27, l-glutamine, and nerve growth factor. For the selection of neurons, cultures were cycled between culturing medium and medium containing fluorodeoxyuridine/uridine every 2 days for the fist week of culture. Two-week-old cultures were used for the study. Cells were washed with fresh neurobasal medium containing 20 mM HEPES, 0.1% BSA, and 250 μM IBMX. Cells were stimulated for 30 min at 37°C in the same buffer containing 20 μM forskolin along with the indicated agonists. The reaction was terminated by removing the medium and adding ice-cold 5% trichloroacetic acid to the cells. After 30-min incubation at 4°C, the solution was neutralized by the addition of 50 mM sodium acetate buffer, pH 6.2.
BRET Measurement. To monitor receptor-receptor interactions in living cells, BRET2 assays were performed using a TopCount NXT (customized version purchased from BioSignal Packard, Inc., now PerkinElmer Life and Analytical Sciences) as described previously (Mercier et al., 2002). In brief, after catalytic degradation of the substrate DeepBlueC by the energy donor Rluc, light is emitted with a peak at 400 nm. The energy acceptor GFP2 is excited by nonradiative energy transfer if GFP is located within a distance of less than 100 Å from the energy donor. As a result, fluorescence is re-emitted by GFP with a peak at 510 nm. The ratio of the light intensity emitted at 500 to 530 nm over 370 to 450 nm is defined as the BRET2 signal. BRET signals were then corrected by subtracting the BRET background detected in cells expressing only the energy donor fusion protein. The expression level of the energy donor (Rluc) or acceptor (GFP) was controlled by measuring total luminescence and fluorescence for each BRET experiment, as described previously (Mercier et al., 2002). To increase the relative amount of δOR fusion proteins expressed at the cell surface, we took advantage of the pharmacological chaperone effect of hydrophobic δOR antagonist (Petaja-Repo et al., 2002) by treating δOR-Rluc- or δOR-GFP2-expressing cells with 1 μM naltrexone for 12 to 16 h before harvesting and washing the cells.
Radioligand Binding Assay. For ligand binding studies, ∼100,000 cells were seeded in 24-well dishes, coated with 0.1% poly-d-lysine. After 24 h, specific binding of [3H]naloxone (5-10 nM) or [3H]DPDPE (5 nM) was determined in the presence of increasing concentrations of the competing ligands. These radioligand binding assays were carried out on attached cells for 2 h on ice in DMEM with 100 mM HEPES, pH 7.4, and 0.1% of BSA. The amount of cell-bound radioactivity was measured after washing the cells twice with 2 ml of ice-cold binding buffer and lysing them with 0.1% SDS and 0.1 N NaOH.
Internalization Assay. To induce endocytosis of the δOR, cells were stimulated at 37°C for 30 min with 1 μM BAM22 or Leu-enkephalin in DMEM containing 100 mM HEPES and 0.1% BSA. Cells were then washed extensively, and the ligands were allowed to dissociate for 10 min on ice in 2 ml of binding buffer. Receptor sequestration was measured by detecting both total and cell surface binding using 50 nM [3H]naloxone. Specific cell surface binding of [3H]naloxone was defined as the binding inhibited by 1 μM DPDPE, a hydrophilic ligand, whereas total specific binding sites were defined as the binding inhibited by 10 μM naltrexone, a hydrophobic ligand. The amount of total specific binding sites of untreated and agonist pretreated cells was similar, indicative of complete dissociation of the agonist used to induce receptor endocytosis. Receptor sequestration is given as the percentage of receptor binding sites that were no longer accessible by the hydrophilic ligand DPDPE after agonist stimulation.
cAMP Accumulation. To determine δOR agonist-promoted inhibition of forskolin-induced cAMP accumulation in HEK293 cells, ∼200,000 cells were seeded in 12-well dishes coated with 0.1% poly-d-lysine 24 h before the experiment and labeled for 2 to 4 h in serum-free DMEM containing 2 μCi/ml [3H]adenine. Cells were stimulated for 30 min at 37°C in DMEM containing 100 mM HEPES, pH 7.4, 2.5 μM IBMX, 0.1% BSA, and 20 μM forskolin along with the different drugs at the indicated concentrations. The reaction was terminated by removing the medium and adding ice-cold 5% trichloroacetic acid to the cells. [3H]cAMP was then purified by sequential chromatography over DOWEX resin and aluminum oxide, and the cAMP accumulation was expressed as the total amount of [3H]cAMP in picomoles per hour per well. cAMP Radioimmunoassay. For dorsal root ganglia cells, cAMP levels were measured by incubating samples with anti-cAMP anti-serum and 125I-cAMP in sodium acetate buffer at 4°C for 18 h and separating the antibody-bound radioligand by precipitation with 17.5% polyethylene glycol as described previously (Gomes et al., 2003). The radioactivity in the pellet was measured in a gamma counter (PerkinElmer Life and Analytical Sciences).
IPn Production. To determine the accumulation of IPn, ∼105 cells/well were seeded 48 h before the experiment in 24-well dishes coated with 0.1% poly-d-lysine and labeled for 24 h with 1 μCi/ml [myo-3H]inositol. IPn production was then measured after stimulation of the cells for 1 h at 37 °C with the indicated concentrations of the various drugs in DMEM containing 100 mM HEPES, pH 7.4, 0.1% BSA, and 20 mM LiCl. Total IPn was isolated by chromatography over DOWEX (OH-) as described previously (Berridge et al., 1983) and expressed as -fold over basal of nonstimulated cells.
Data Analysis. Data obtained in binding experiments and cAMP or IPn accumulation assays were analyzed using Prism 3.0 (Graph-Pad Software, San Diego, CA). Isotherms were plotted for one or two binding sites; the best fit was then used to calculate Ki or EC50 values. Data obtained in the BRET titration experiments were plotted using a single binding site saturation isotherm. Statistical significance of the differences was assessed by two-tailed Student's t test.
Results
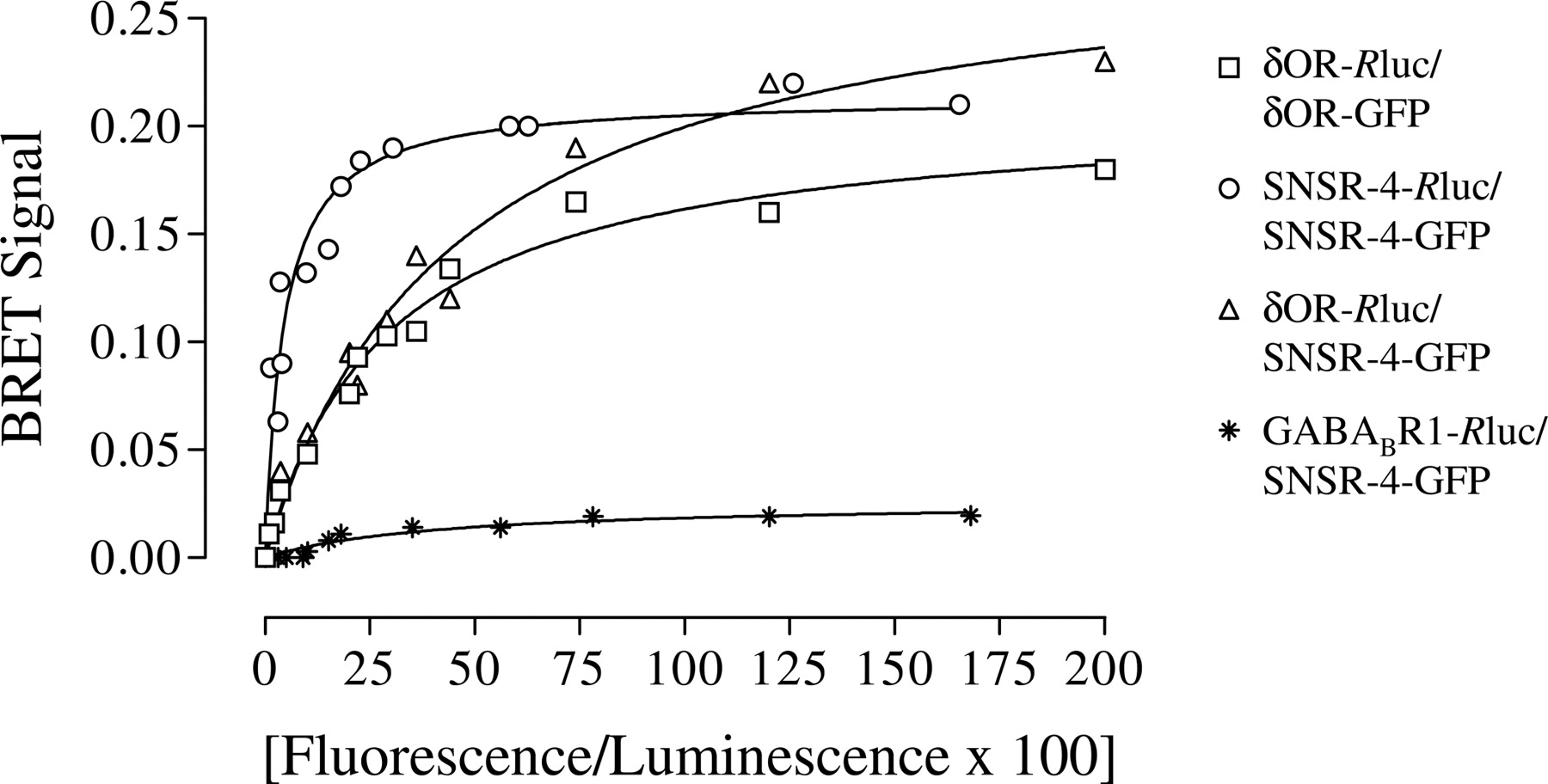
Hetero-Oligomerization between the δOR and the SNSR-4 Monitored by BRET2. The BRET technique has been successfully used to monitor receptor-receptor interactions in living cells (Angers et al., 2000; McVey et al., 2001; Ayoub et al., 2002). To determine whether the δOR and the SNSR-4 can form hetero-oligomers, we fused both receptors on their C terminus to Rluc (energy donor) or GFP2 (energy acceptor) and applied the BRET2 technology. We took advantage of the quantitative features of BRET2 and performed BRET titration experiments to determined BRET50 values for both homo-oligomers and the δOR/SNSR-4 hetero-oligomer. It has been proposed that BRET50 values reflect the relative affinity of a receptor-receptor complex (Mercier et al., 2002) and therefore allow to compare the propensity of a receptor to form homo- or hetero-oligomeric complexes. As shown in Fig. 1, BRET titration curves for both homo- and hetero-oligomers were best fitted to a hyperbolic function, indicating the saturation of the energy donor by the energy acceptor as a result of specific interactions of the proteins fused to the BRET pair. In BRET titration experiments between the unrelated GABABR1-Rluc fusion protein and the SNSR-4-GFP fusion protein, no detectable BRET signal was observed (Fig. 1), providing further evidence for the specific interactions between the SNSR-4-Rluc or δOR-Rluc fusion protein and the SNSR-4-GFP. To characterize the potential functional consequences of such hetero-oligomeric complexes between δOR and SNSR-4, HEK293 cell lines stably expressing each of the receptor individually (HEK293-δOR and HEK293-SNSR-4) or coexpressing the two receptors (HEK293-δOR/SNSR-4) were generated. To limit the differences between cell lines, the HEK293-δOR/SNSR-4 cell line was obtained by transfection of the SNSR-4 construct into the HEK293-δOR cells used in the study (see Materials and Methods).

Detection of δOR/SNSR-4 hetero-oligomers by BRET2. A constant cDNA amount of the energy donor δOR-Rluc, SNSR-4-Rluc, or GABABR1-Rluc and increasing amounts of the energy acceptor δOR-GFP2 or SNSR-4-GFP2 were transiently coexpressed in HEK293 cells, and 48 h after transfection, BRET signals were measured after the addition of the luciferase substrate DeepBlueC as described under Materials and Methods. BRET signals are plotted as a function of the expression ratio of receptor-GFP2 over receptor-Rluc (×100) obtained by measuring the total luminescence and fluorescence in each condition. The expression level of the receptor-Rluc fusion protein was not significantly affected (±20%) by the coexpression of the receptor-GFP2 fusion proteins. The BRET titration curves were drawn from data obtained in three independent experiments.

Effects of SNSR-4 coexpression on agonist-promoted δOR endocytosis. HEK293-δOR and HEK293-δOR/SNSR-4 cells were incubated with 1 μM Leu-enkephalin or BAM22 for 30 min at 37°C. After treatment with the corresponding drugs, cell surface receptors were quantified by assessing the proportion of [3H]naloxone (50 nM) binding sites accessible to the hydrophilic ligand DPDPE and are expressed on the y-axis as the percentage of total receptor number. Numbers above the columns indicate the extent of endocytosis in percentage after treatment with the corresponding drug. Results are expressed as the mean ± S.E. of three independent experiments carried out in triplicates. Asterisk indicates a significant (p < 0.05) difference of the δOR endocytosis observed in δOR/SNSR-4-versus δOR-expressing cells.
Coexpression of δOR and SNSR-4 Inhibits Agonist-Induced δOR Endocytosis. After sustained exposure to agonists, most GPCRs undergo rapid endocytosis that results in a loss of receptor sites from the cell surface (Von Zastrow, 2001, 2003). Coexpression of two distinct receptors has often been reported to modulate this regulatory process either as a result of hetero-oligomerization (George et al., 2000) or heterologous cross-talk between the two receptor signaling cascades (Yoshimasa et al., 1987; Klein et al., 2001; Xiang et al., 2001). In particular, the endocytotic properties of the δOR were reported to be affected by the coexpression of several receptors (Jordan and Devi, 1999; George et al., 2000; Jordan et al., 2001). To determine whether δOR endocytosis could be affected by its hetero-oligomerization with SNSR-4, endocytosis of the δOR promoted by the mixed δOR/SNSR-4 agonist BAM22 or δOR-selective agonist Leu-enkephalin was examined in HEK293-δOR and HEK293-δOR/SNSR-4 cells. As seen in Fig. 2, BAM22-promoted endocytosis of the δOR was significantly lower in cells coexpressing SNSR-4 (21 ± 4% in HEK293-δOR/SNSR-4 compared with 56 ± 6% in HEK-δOR cells), indicating an inhibitory effect of the SNSR-4 on the δOR endocytosis when both receptors are activated. It is noteworthy that δOR endocytosis induced by the δOR-selective agonist Leu-enkephalin was also lower in cells coexpressing SNSR-4 (52 ± 5% in HEK293-δOR/SNSR-4 compared with 91 ± 7% in HEK293-δOR cells), indicating that the inhibitory action did not require agonist-promoted SNSR-4 signaling and that the presence of SNSR-4 is sufficient to block δOR endocytosis. We therefore conclude that the reduced agonist-promoted endocytosis of the δOR in HEK293-δOR/SNSR-4 is probably due to hetero-oligomerization of the two receptors. Consistent with this idea, no SNSR-4 endocytosis could be detected in either HEK293-SNSR-4 or HEK293-δOR/SNSR-4 cells in response to BAM22 (data not shown), indicating that, under these particular conditions, SNSR-4 cannot undergo robust internalization in these cells.
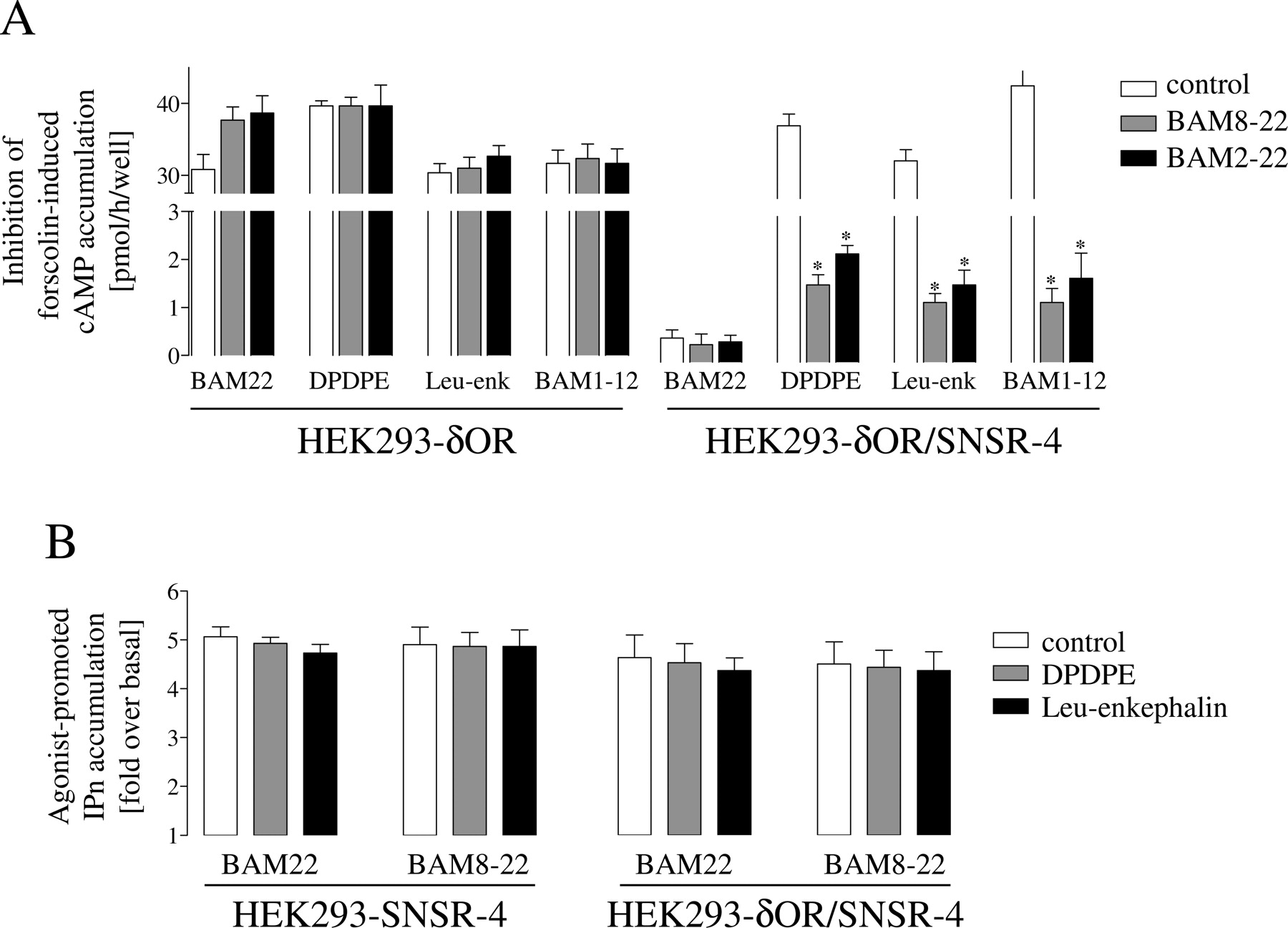
Signaling Properties of the δOR/SNSR-4 Hetero-Oligomer. Several studies have recently suggested the occurrence of receptor trans-activation within receptor hetero-oligomers (Galvez et al., 2001; Lee et al., 2002; Carrillo et al., 2003). Based on the distinct ligand binding profile and different G protein coupling selectivity of δOR (binding of Leu-enkephalin and BAM1-12 and coupling to Gαi/o that leads to adenylyl cyclase inhibition) and SNSR-4 (binding of BAM2-22 and BAM8-22 and coupling to Gαq that promotes phospholipase C activation), the δOR/SNSR-4 hetero-oligomer represents an excellent system to consider potential trans-activation. Hence, the ability of δOR and SNSR-4 ligands to activate Gαi/o and Gαq was assessed in HEK293-δOR, HEK293-δOR/SNSR-4, and HEK293-SNSR-4 cells. As shown in Fig. 3A and as expected from the previously established selectivity of the compounds, saturating concentrations of the SNSR-4-selective agonists BAM2-22 and BAM8-22 activate IPn production in HEK293-SNSR-4 and HEK293-δOR/SNSR-4 but not in the HEK293-δOR cells. Saturating concentrations of the δOR-selective agonists Leu-enkephalin and BAM1-12 inhibit forskolin-induced cAMP accumulation in HEK293-δOR and HEK293-δOR/SNSR-4 but not in HEK293-SNSR-4 cells (Fig. 3B). Furthermore, selective δOR agonists Leu-enkephalin and BAM1-12 did not promote any IPn production in HEK293-δOR cells (Fig. 3A), and the SNSR-4 agonists BAM8-22 and BAM2-22 did not inhibit forskolin-stimulated cAMP accumulation in HEK293-SNSR-4 cells (Fig. 3B), confirming the selective coupling of the δOR to Gαi/o and of the SNSR-4 to Gαq. When considering cells expressing the δOR/SNSR-4 hetero-oligomer, no evidence of receptor trans-activation was observed, and each receptor acted as an independent signaling unit, coupling exclusively to its cognate G protein upon selective activation. Indeed, as shown in Fig. 3A, the δOR-selective agonists Leu-enkephalin and BAM1-12 did not activate the phospholipase C pathway in HEK293-δOR/SNSR-4 cells. Likewise, no inhibition of the adenylyl cyclase activity could be promoted by the SNSR-4-selective agonists BAM8-22 and BAM2-22 in these cells (Fig. 3B).

Signaling properties of δOR/SNSR-4 hetero-oligomeric complexes. A, HEK293-SNSR-4, HEK293-δOR, and HEK293-δOR/SNSR-4 cells were stimulated for 60 min at 37°C with 1 μM concentrations of the SNSR-4-specific agonists BAM2-22 or BAM8-22 or the δOR-specific agonists Leu-enk or BAM1-12. Phospholipase C activity was assessed by measuring the accumulation of [3H]IPn in cells prelabeled with [myo-3H]inositol and expressed as -fold over basal of non-stimulated cells. B, HEK293-SNSR-4, HEK293-δOR, and HEK293-δOR/SNSR-4 cells were stimulated for 30 min at 37°C with 20 μM forskolin and 1 μM concentrations of the SNSR-4-specific agonists BAM2-22 and BAM8-22 or the δOR-specific agonists Leu-enk and BAM1-12. Adenylyl cyclase activity was assessed by measuring the accumulation of [3H]cAMP in cells prelabeled with [3H]adenine and expressed as the percentage of the agonist-promoted inhibition of forskolin-induced cAMP accumulation. Results are expressed as the mean ± S.E. of three independent experiments carried out in triplicates.
Trans-Inhibition of SNSR-4 by the δOR Antagonist Naltrexone. Naltrexone is a well established OR antagonist (Portoghese, 1993) that has previously been shown not to affect SNSR-mediated signaling (Lembo et al., 2002; Kirchmayer et al., 2003). To determine whether naltrexone could affect signaling of the δOR/SNSR-4 hetero-oligomer, the effect of naltrexone was assessed on the BAM22-stimulated IPn production in HEK293-SNSR-4 and HEK293-δOR/SNSR-4 cells. In line with a previous report (Lembo et al., 2002) and consistent with the notion that naltrexone does not bind SNSR-4, the dose-response curve of BAM22-stimulated IPn production in HEK293-SNSR-4 cells was unaffected by a cotreatment with naltrexone (Fig. 4A, left). In contrast, the efficacy of BAM22 to stimulate IPn production was clearly reduced in HEK293-δOR/SNSR-4 cells when naltrexone was coadministered (Fig. 4A, right). Given that the activation of δOR in HEK293-δOR cells did not promote any detectable IPn production (Fig. 3A), these data indicate that naltrexone acted as an antagonist on the δOR/SNSR-4-promoted IPn production stimulated by BAM22. The fact that naltrexone did not completely block the SNSR-4-promoted IPn production suggests either that naltrexone only partially blocks the hetero-oligomer activation by SNSR agonists or that a significant fraction of SNSR homo-oligomer is present in HEK293-δOR/SNSR-4 cells. This later possibility is consistent with the finding that SNSR-4 seems to have greater propensity to form homo-than hetero-oligomers (Fig. 1). It is noteworthy that this trans-inhibition of the SNSR-4 signaling by naltrexone within the δOR/SNSR-4 hetero-oligomer was restricted to the mixed, bivalent agonist BAM22. Indeed, the naltrexone treatment had no effect on the IPn production elicited by the SNSR-4-selective agonists BAM2-22 or BAM8-22, in HEK293-δOR/SNSR-4 cells (Fig. 4B). These data suggest that coexpression of δOR and SNSR-4 promotes a BAM22-mediated SNSR-4 signaling (IPn production) that is sensitive to the δOR antagonist naltrexone, most likely as a result of hetero-oligomerization between the two receptors and the interaction of BAM22 with the two protomers. Because no selective SNSR-4 antagonist has been identified yet, the possible reciprocal antagonism of the δOR-mediated inhibition of adenylyl cyclase by blockade of the SNSR-4 receptor could not be tested.
Costimulation of the δOR and SNSR-4 Protomers Leads to the Selective Activation of the SNSR-4 Signaling Pathway. Even though no trans-activation of one receptor signaling pathway by the other receptor could be observed in cells coexpressing δOR and SNSR-4 (see above), we examined the possible influence of their simultaneous stimulation on signaling. In the case of δOR-expressing cells, the presence of SNSR-4-selective agonists BAM2-22 and BAM8-22 had no influence on the ability of the mix δOR/SNSR-4 agonist BAM22 and δOR agonists DPDPE, Leu-enkephalin, and BAM1-12 to inhibit forskolin-stimulated cAMP production (Fig. 5A). It is surprising that, in HEK293-δOR/SNSR-4 cells, the two selective SNSR-4 agonists almost completely blocked the adenylyl cyclase inhibition promoted by the δOR-selective agonists DPDPE, Leu-enkephalin, and BAM1-12, indicating that the SNSR-4 agonists trans-inhibited the δOR signaling pathway. In line with this apparent blockade of the δOR signaling pathway, after concomitant stimulation of the SNSR-4 receptor, we found that the mix δOR/SNSR-4 agonist BAM22 did not inhibit the forskolin-stimulated adenylyl cyclase in HEK293-δOR/SNSR-4 cells. This is in clear contrast with the ability of BAM22 to inhibit cAMP production in δOR-expressing cells (Fig. 5A). The observation that the treatment of cells with the dual SNSR-4/DOR ligand (BAM22) or with the SNSR-4 selective ligands (BAM2-22 and BAM8-22) almost completely blocked the δOR-mediated inhibition of forskolin-stimulated cAMP production (the remaining inhibition being less than 8% of the δOR inhibition observed in the absence of SNSR agonists) suggests that most δOR are engaged in hetero-oligomerization with SNSR-4.

Naltrexone-sensitivity of BAM22-promoted production of IPn in HEK293-δOR/SNSR-4 cells. A, HEK293-SNSR-4 and HEK293-δOR/SNSR-4 cells were stimulated for 60 min at 37°C with increasing concentrations of BAM22 in the absence (open symbols) or presence (filled symbols) of 10 μM naltrexone. B, HEK293-SNSR-4 and HEK293-δOR/SNSR-4 cells were stimulated with 1 μM BAM22, BAM2-22, or BAM8-22 in the presence or absence of 10 μM naltrexone. Phospholipase C activity was assessed as in Fig. 3 and expressed as -fold over basal of nonstimulated cells (A) or as the percentage of naltrexone-induced inhibition of agonist-promoted IPn production (B). Results are expressed as the mean ± S.E. of three independent experiments carried out in triplicates. Asterisk indicates a significant (p < 0.05) difference of the BAM22-promoted IPn accumulation in naltrexone-treated versus nontreated HEK293-δOR/SNSR-4 cells.

Coactivation of the δOR/SNSR-4 hetero-oligomer promotes phospholipase C activity but does not inhibit adenylyl cyclase activity. A, HEK293-δOR and HEK293-δOR/SNSR-4 cells were stimulated for 30 min at 37°C with 20 μM forskolin and 1 μM concentrations of the mixed, bivalent agonist BAM22 or the δOR-specific agonists DPDPE, Leu-enk, or BAM1-12, in the absence (white columns) or presence of 1 μM concentrations of the SNSR-4-specific agonists BAM8-22 (gray columns) or BAM2-22 (black columns). Adenylyl cyclase activity was assessed as in Fig. 3 and expressed as picomoles of cAMP produced per hour per well. Results are expressed as the mean ± S.E. of three independent experiments carried out in triplicates. B, HEK293-SNSR-4 and HEK293-δOR/SNSR-4 cells were stimulated for 60 min at 37°C with 1 μM concentrations of the mixed, bivalent agonist BAM22 or SNSR-4-selective agonist BAM8-22 in the absence (white columns) or presence of 10 μM concentrations of the δOR-specific agonists DPDPE (gray columns) or Leu-enk (black columns). Phospholipase C activity was assessed as in Fig. 3 and expressed as -fold over basal of nonstimulated cells. Results are expressed as the mean ± S.E. of three independent experiments carried out in triplicates. Asterisk in A indicates a significant (p < 0.05) difference between single and coactivated receptors in HEK293-δOR/SNSR-4 cells.
It is noteworthy that this trans-inhibition of the δOR signaling by SNSR-4 agonists was found to be unidirectional because simultaneous activation of both receptors did not affect the SNSR-4-mediated production of IPn in HEK293-δOR/SNSR-4 cells (Fig. 5B). Together, these findings suggest that simultaneous agonist binding to the two protomers of the δOR/SNSR-4 hetero-oligomer leads to the exclusive SNSR-4-promoted Gαq activation, to the expense of the δOR-activated Gαi/o.

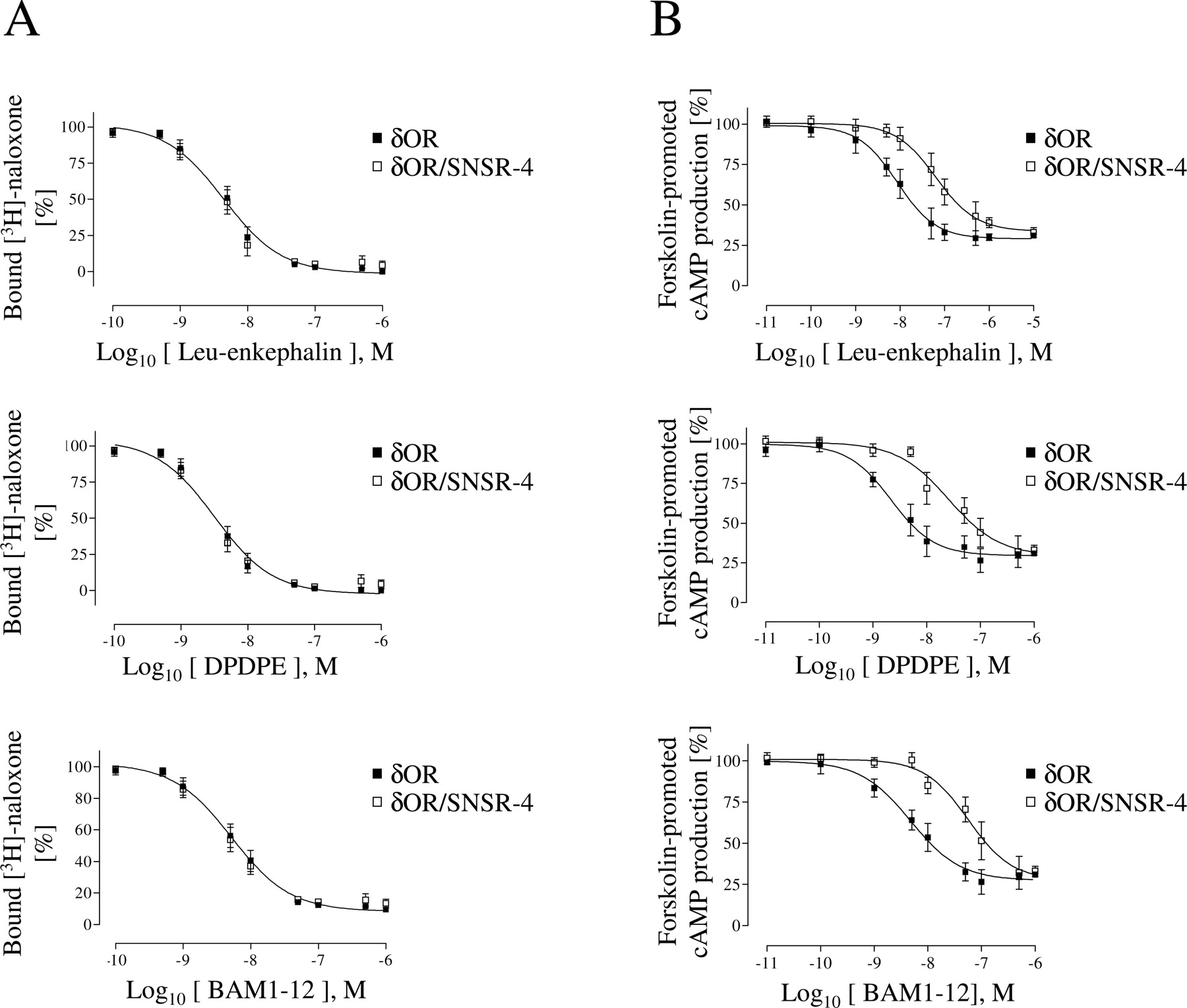
Binding and signaling properties of the δOR/SNSR-4 hetero-oligomer. A, competition binding studies with the δOR-specific antagonist [3H]naloxone (10 nM) were performed on attached cells as described under Materials and Methods, and Ki values were calculated. HEK293-δOR cells: Leu-enkephalin, 4.4 ± 0.3 nM; DPDPE, 3.2 ± 0.2 nM; and BAM1-12, 5.2 ± 0.4 nM. HEK293-δOR/SNSR-4 cells: Leu-enkephalin, 4.2 ± 0.4 nM; DPDPE, 3.4 ± 0.2 nM; and BAM1-12, 5.1 ± 0.3 nM. Binding curves represent the mean ± S.E. of three independent experiments performed in duplicates. B, HEK293-δOR and HEK293-δOR/SNSR-4 cells were stimulated for 30 min at 37°C with 20 μM forskolin and increasing concentrations of the δOR-specific agonists Leu-enkephalin, DPDPE, or BAM1-12. Adenylyl cyclase activity was assessed as in Fig. 3 and expressed as the percentage of the maximal accumulation induced by forskolin in the absence of any agonist. EC50 values are as follows. HEK293-δOR cells: Leu-enkephalin, 8.6 ± 0.4 nM; DPDPE, 2.2 ± 0.2 nM; and BAM1-12, 4.8 ± 0.4 nM. HEK293-δOR/SNSR-4 cells: Leu-enkephalin, 64.1 ± 1.4 nM; DPDPE, 25.5 ± 0.9 nM; and BAM1-12, 56.7 ± 2.3 nM. Dose-response curves represent the mean ± S.E. of three independent experiments performed in duplicates.
The δOR/SNSR-4 Hetero-Oligomer Has a Reduced Coupling Efficiency to Gi/o. The data described in the above-mentioned section suggests that the δOR/SNSR-4 hetero-oligomer has a reduced propensity to interact with Gαi/o in the presence of SNSR-4 agonists. The potency of an agonist to activate a given signaling pathway reflects both the affinity of the agonist to the receptor and the coupling efficiency of the agonist/receptor complex to its cognate G protein. Thus, we determined the binding affinities and potencies of the selective δOR agonists in both HEK293-δOR and HEK293-δOR/SNSR-4 cells. As shown in Fig. 6A, the propensity of Leu-enkephalin, DPDPE, and BAM1-12 to compete for [3H]naloxone binding was identical in δOR- and δOR/SNSR-4-expressing cells. In contrast, however, their potency to inhibit forskolin-stimulated cAMP production was shifted to the right by 7.5 to 12.1 times in HEK293-δOR/SNSR-4 compared with HEK293-δOR cells (Fig. 6B). This shift was not due to difference in δOR spare receptors between the two cell lines because they expressed very similar numbers of δOR binding sites (HEK293-δOR, 4.1 ± 0.7 pmol/mg versus HEK293-δOR/SNSR-4, 3.8 ± 0.8 pmol/mg), a finding that is not surprising given that the HEK293-δOR/SNSR-4 cell line was generated by stably integrating SNSR-4 into the HEK293-δOR cells. The two cell lines also displayed very similar levels of forskolin-stimulated cAMP accumulation (HEK293-δOR, 55.4 ± 5.8 versus HEK293-δOR/SNSR-4, 53.2 ± 6.7 pmol/h/well). The unaltered affinity in face of a reduced potency suggests firstly that the δOR in HEK293-δOR/SNSR-4 cells is less efficiently coupled to Gαi/o. The observation that such loss of coupling to Gαi/o was not accompanied by a reduction of the high-affinity state for agonist (believed to reflect the G protein coupling state) further suggests that coupling to another G protein (most likely Gαq) can compensate for Gαi/o in maintaining the high-affinity binding states for agonists to the δOR in the hetero-oligomer. In summary, these data provide additional evidence that coexpression of the SNSR-4 has a negative effect on the Gαi/o coupling of the δOR, even in the absence of SNSR-4 agonists.
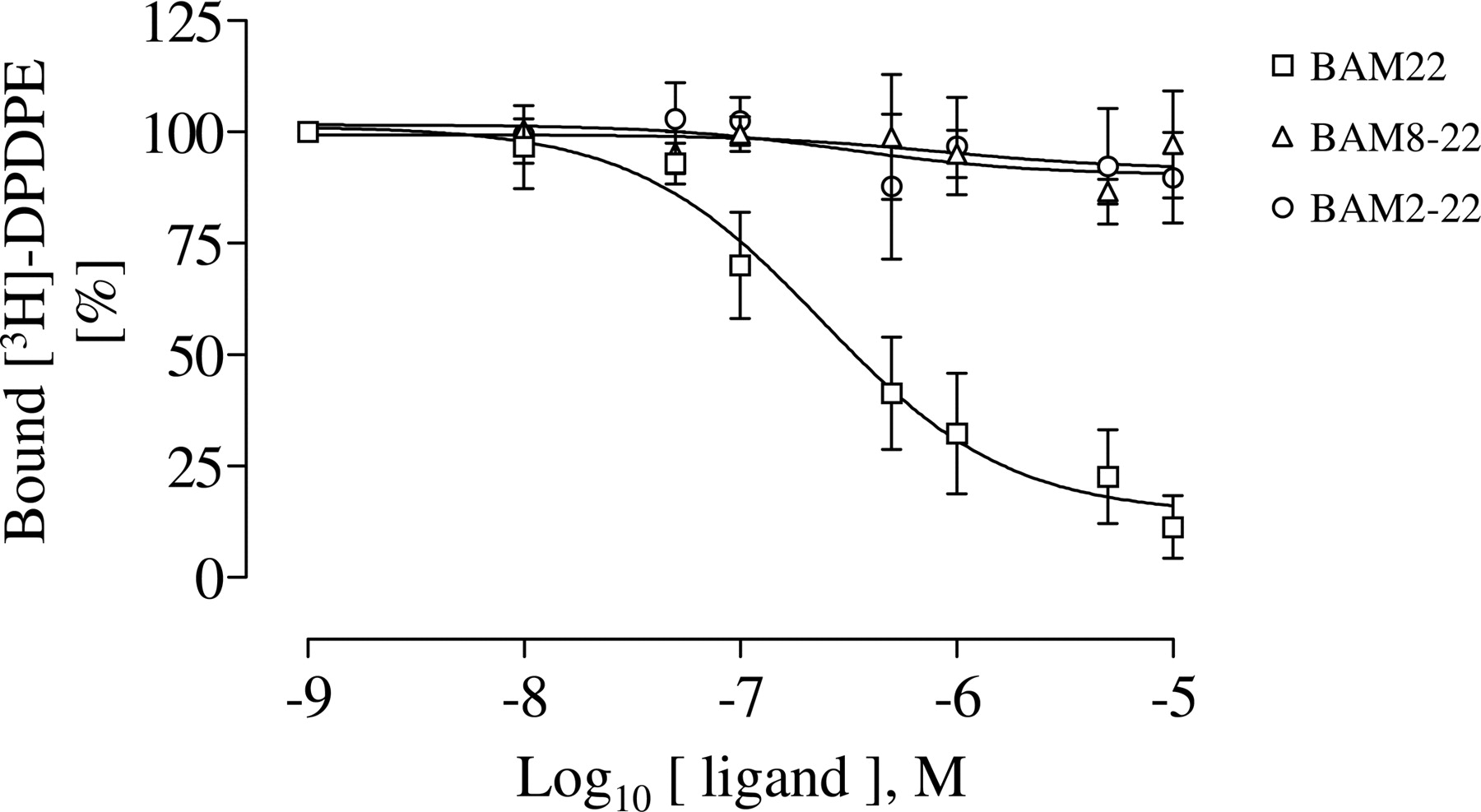
Selective SNSR-4 and δOR Agonists Can Bind Simultaneously to the δOR/SNSR-4 Hetero-Oligomer. The dramatically reduced efficacy of δOR agonists in the presence of SNSR-4 agonists observed for HEK293-δOR/SNSR-4 cells define SNSR-4 agonists as functional antagonists of δOR within the δOR/SNSR-4 hetero-oligomer. To determine whether this antagonism resulted from an inhibition of δOR agonist binding to the oligomer, the ability of the selective SNSR-4 agonists BAM2-22 and BAM8-22 to inhibit [3H]DPDPE binding to HEK293-δOR/SNSR-4 cells was assessed. As illustrated in Fig. 7, neither BAM2-22 nor BAM8-22 inhibited the binding of [3H]DPDPE. This contrasts with the potent inhibition of the [3H]DPDPE binding by the mixed δOR/SNSR-4 agonist BAM22 and indicates that both δOR and SNSR-4 protomers can bind their selective agonists simultaneously.
Protein Kinase C-Dependent Cross-Talk between SNSR-4 and δOR Cannot Account for the Decreased Efficacy of δOR-Selective Agonist after SNSR-4 Coactivation. The results presented so far are consistent with the notion that the inhibitory action of SNSR-4 agonists on the δOR signaling could result from the formation of δOR/SNSR-4 hetero-oligomers that have reduced ability to couple to Gαi/o. However, it could also result from cross-talk regulation. This is particularly relevant when considering that protein kinase C activation can promote δOR desensitization (Ueda et al., 1995; Xiang et al., 2001). Hence, the activation of the phospholipase C/protein kinase C signaling pathway by SNSR-4 in HEK293-δOR/SNSR-4 cells could promote δOR desensitization, resulting in a decreased efficacy for δOR agonists. To verify this possibility, we assessed the effect of the protein kinase C activator PMA on the δOR-mediated adenylyl cyclase inhibition in HEK293-δOR and HEK293-δOR/SNSR-4 cells. As illustrated in Fig. 8, PMA only marginally reduced the efficacy of BAM22 (HEK293-δOR) or BAM1-12 (HEK293-δOR and HEK293-δOR/SNSR-4) to inhibit forskolin-stimulated cAMP accumulation compared with the effect of the SNSR-4 agonists BAM2-22 and BAM8-22 in δOR/SNSR-4-expressing cells (Fig. 5A). Increasing PMA concentration did not promote any additional inhibition of the δOR mediated signaling (data not shown). To further assess whether protein kinase C-mediated desensitization of δOR could account for the inhibition of δOR signaling by SNSR-4 agonists, the effects of the protein kinase C inhibitor bisindolylmaleimide I were assessed in HEK293-δOR/SNSR-4 cells. Pretreatment of the cells with bisindolylmaleimide I only marginally restored the BAM22-promoted adenylyl cyclase inhibition, whereas it partially blocked the inhibitory action of BAM2-22 on the BAM1-12 promoted adenylyl cyclase inhibition (Fig. 8). Together, these data indicate that although protein kinase C-mediated δOR desensitization can contribute to the regulatory influence of SNSR-4 on δOR signaling, the extent of protein kinase C-mediated desensitization cannot account for the dramatic reduction in δOR/Gαi/o coupling efficacy resulting from the coactivation of SNSR-4 in HEK293-δOR/SNSR-4 cells.

SNSR-4-specific agonists do not inhibit [3H]DPDPE binding to HEK293-δOR/SNSR-4 cells. Competition binding studies using the δOR-specific agonist [3H]DPDPE (5 nM) and increasing concentrations of the mixed, bivalent agonist BAM22 or the SNSR-4-selective agonists BAM8-22 and BAM2-22 were performed on attached HEK293-δOR/SNSR-4 cells as described under Materials and Methods. Results represent the mean ± S.E. of three independent experiments performed in duplicates.

Effects of protein kinase C activity on δOR-promoted inhibition of adenylyl cyclase activity. HEK293-δOR or HEK293-δOR/SNSR-4 cells were stimulated for 30 min at 37°C with 20 μM forskolin and 1 μM concentrations of the various BAM peptides (white columns), costimulated with 50 μM concentrations of the protein kinase C-specific activator PMA (gray columns), or pretreated for 30 min at 37°C with 500 nM concentrations of the protein kinase C specific inhibitor bisindolylmaleimide I (BIM; black columns). Adenylyl cyclase activity was assessed as in Fig. 3 and expressed as picomoles of cAMP produced per hour per well. Results are expressed as the mean ± S.E. of three independent experiments carried out in triplicates. Asterisk indicates a significant (p < 0.05) difference of the agonist-promoted inhibition of forskolin-induced cAMP accumulation in cells treated with bisindolylmaleimide I or PMA versus nontreated cells.
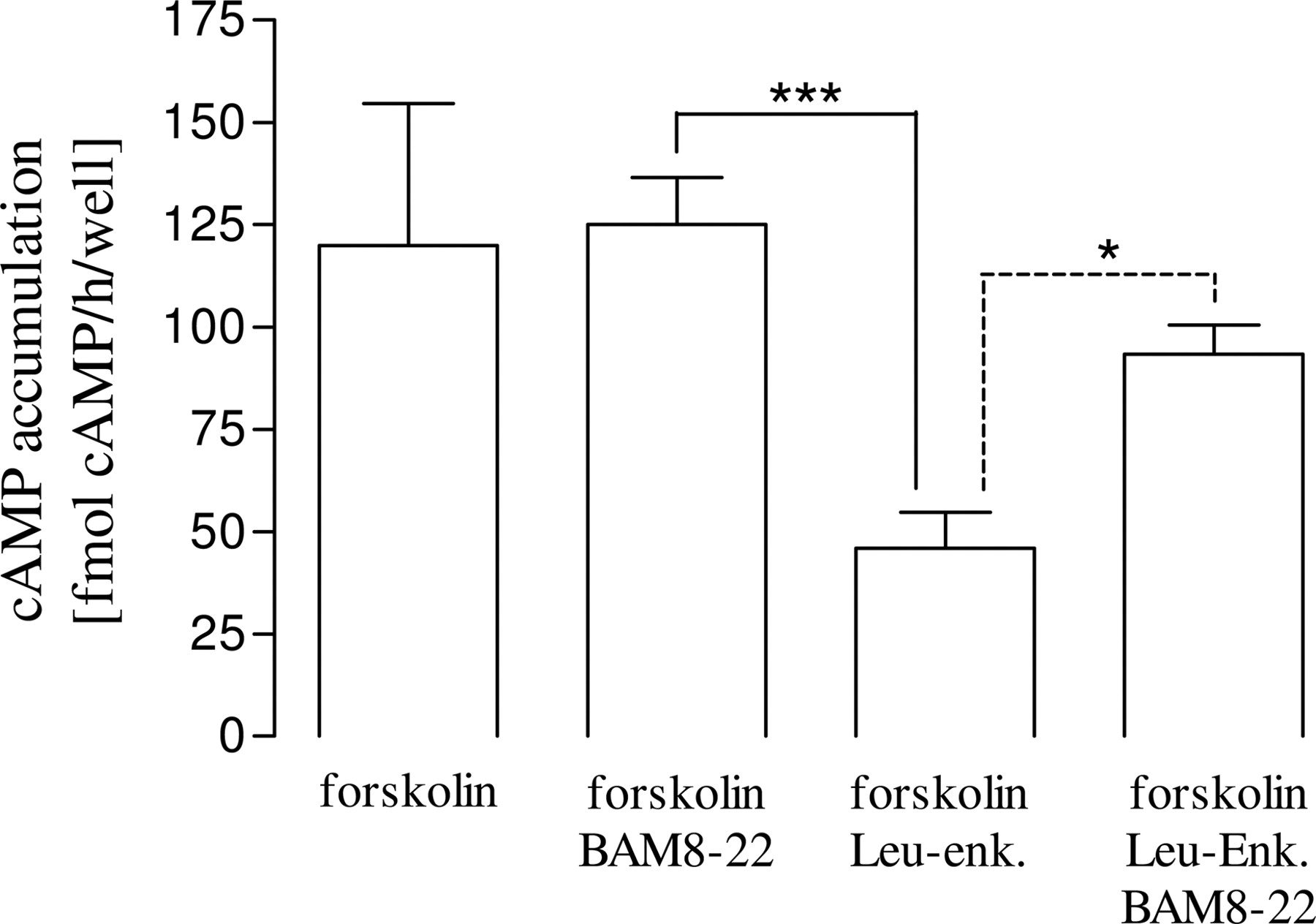
Signaling properties of OR and SNSR subtypes in rat dorsal root ganglion neurons. Neurons were stimulated for 30 min at 37°C with 20 μM forskolin and 1 μM concentrations of the SNSR-specific agonist BAM8-22 or the OR specific-agonist Leu-enk. Inhibition of adenylyl cyclase activity was assessed by measuring the amount of accumulated cAMP using a radioimmunoassay. Results are expressed as the mean ± S.E. of three independent experiments. Statistically significant differences between levels in cells treated with Leu-enk alone and those simultaneously treated with BAM8-22 are indicated (***, p < 0.001; *, p < 0.01).
BAM8-22 Reduces Leu-Enkephalin-Mediated Inhibition of Adenylyl Cyclase Activity in Cultured Neurons from Rat Dorsal Root Ganglia. In an effort to assess the possible physiological relevance of the δOR/SNSR-4 hetero-oligomer characterized in HEK293 cells, the influence of SNSR stimulation on Leu-enkephalin-promoted signaling was assessed in neurons derived from embryonic day 16 rat embryos dorsal root ganglia. As expected based on the G protein-coupling profile of OR and the SNSR subtypes, Leu-enkephalin significantly inhibited adenylyl cyclase activity in rat dorsal root ganglia, whereas BAM8-22 had no effect under the same conditions (Fig. 9). However, in line with our observation in HEK293-δOR/SNSR-4 cells shown in Fig. 5A, coadministration of BAM8-22 and Leu-enkephalin significantly decreased Leu-enkephalin signaling in cultured rat dorsal root ganglia (Fig. 9). These data provide evidence that concomitant activation of SNSRs expressed in cultured rat dorsal root ganglia inhibits Gαi/o signaling promoted by OR subtypes, suggesting the existence of functional interactions between SNSRs and OR that could result from hetero-oligomerization between these receptor. Given the presence of the three OR subtypes and potentially several SNSR subtypes in dorsal root ganglias, further work will be required to establish the identity of the hetero-oligomers potentially underlying this functional interaction in vivo.
Discussion
In this study, we provide evidence based on BRET titration experiments that the δOR and the SNSR-4 form hetero-oligomeric complexes when transiently coexpressed in HEK293 cells. The existence of such hetero-oligomers is further supported by the functional effects that stable coexpression of the two receptors had on each of the protomer properties. Although, protein kinase C-mediated cross-talk between the two receptors could contribute to some of the regulatory actions of the SNSR-4 on δOR signaling, most of the observed alterations cannot be attributed to such cross-talk and most likely results from the hetero-oligomerization between the receptors.
Resonance energy transfer approaches similar to the BRET assay are becoming tools of choice to monitor receptor-receptor interactions in living cells (Angers et al., 2000; Ayoub et al., 2002; Ramsay et al., 2002). The strength and limitations of the technique were extensively discussed in a previous report (Breit et al., 2004). In the present study, we report similar propensity of the δOR to homo-oligomerize and to form hetero-oligomers with the SNSR-4, suggesting that a significant amount of δOR/SNSR-4 hetero-oligomers arises when coexpressed. This is not unique to this receptor pair because similar high-affinity hetero-oligomerization has been reported for several closely related GPCR subtypes (Ayoub et al., 2002; Mercier et al., 2002; Ramsay et al., 2002; Terrillon et al., 2003; Breit et al., 2004).
Coexpression of two receptor subtypes often changed agonist-promoted receptor trafficking, probably as a result of direct receptor-receptor interactions (Jordan and Devi, 1999; AbdAlla et al., 2000; George et al., 2000; Rocheville et al., 2000b; Pfeiffer et al., 2001; Stanasila et al., 2003; Xu et al., 2003; Breit et al., 2004). In particular, coexpression of the endocytosis resistant κOR has been shown to inhibit δOR endocytosis (Jordan and Devi, 1999). In line with these previous studies, coexpression of the SNSR-4 significantly reduced BAM22-promoted δOR endocytosis. Similar to the κOR, SNSR-4 does not undergo agonist-promoted endocytosis (data not shown), suggesting that endocytosis-resistant receptors may dominate over the endocytosis-prone δOR. The lack of BAM22-promoted δOR endocytosis in the coexpressing cells could result from a cross-talk between the SNSR-4 and the δOR because of the BAM22-promoted activation of the SNSR-4/protein kinase C signaling pathway. This is, however, very unlikely because the agonist-promoted δOR endocytosis induced by the δOR-selective agonist Leu-enkephalin was also inhibited, indicating that the mere expression of SNSR-4 is sufficient to block δOR endocytosis. Inhibition of the δOR endocytosis independent of agonist-promoted SNSR-4 activity strongly suggests that direct receptor-receptor interactions are responsible for the reduced δOR endocytosis, pointing out to an important role of hetero-oligomerization in the regulation of agonist-promoted endocytosis of GPCRs.
The analysis of HEK293-δOR/SNSR-4 cells revealed changes in the sensitivity of the protomers to agonists and antagonists. For example, phospholipase C activity promoted by the nonselective agonist BAM22 was blocked by the δOR antagonist naltrexone, whereas the SNSR-4-selective agonists, BAM2-22 and BAM8-22 were not affected by the same treatment. This trans-inhibition of the SNSR-4-specific signaling pathway by a δOR antagonist provides strong evidence that, after coexpression of both receptors, a signaling unit arose that was clearly distinguishable from δOR (agonist-promoted IPn accumulation) and SNSR-4 (no sensitivity toward naltrexone). Given the fact that BAM22 is not able to penetrate the cell membrane, these results indicate that δOR/SNSR-4 hetero-oligomeric complexes are located at the cell surface and functionally coupled to the Gαq/phospholipase C signaling pathway. So far, trans-inhibition has only been described in cells coexpressing the dopamine-2 and the somatostatin-5 (Rocheville et al., 2000a) or the β-adrenergic and the angiotensin-II type 1 (Barki-Harrington et al., 2003) receptor subtypes. However, in contrast with these previous reports, the δOR antagonist naltrexone could not trans-inhibit the signaling promoted by the SNSR-4-selective agonists BAM2-22 or BAM8-22 but could only inhibit the phospholipase C activation promoted by the mixed δOR/SNSR-4 agonist BAM22, suggesting that the inhibition is linked to the potential of BAM22 to bind simultaneously to δOR and SNSR-4 through its N and C terminus, respectively. Such bivalent ligand binding mode has already been described for synthetic OR ligands that can bind to the δOR/κOR hetero-oligomer (Portoghese et al., 1986) and revealed unique properties of this receptor complex (Bhushan et al., 2004; Daniels et al., 2005). Assuming a comparable bivalent binding mode between BAM22 and each of the protomers within the δOR/SNSR-4 hetero-oligomer, naltrexone-induced competition of BAM22 from the δOR could in turn lead to the displacement of the agonist from the SNSR-4, thus inhibiting phospholipase C activation.
Coexpression of two receptors has previously been described to inactivate one of the two receptors. For example, coexpression of the angiotensin AT2 subtype blocked the agonist-promoted activation of the AT1 receptor (AbdAlla et al., 2001). A second report demonstrated that the presence of the somatostatin SST3 receptor inhibited the agonist-promoted activation of the SST2A (Pfeiffer et al., 2001). Of particular interest for the present study is the observation that coexpression of the κOR and δOR blocked the agonist-promoted activation of both receptors (Jordan and Devi, 1999). However, simultaneous activation of the δOR and κOR rescued the agonist-promoted receptor activity of both receptors, indicating positive synergistic effects after coactivation of these opioid receptor subtypes. In contrast, coactivation of the μOR/α-adrenergic-2A hetero-oligomer initiated less G coupling compared with single-activated protomers within the hetero-oligomer (Jordan et al., 2003). In the case of the δOR/SNSR-4 hetero-oligomer, both receptors maintained their signaling integrity (activation of Gαq and Gαi/o for SNSRs and δOR, respectively) when specifically activated. However, simultaneous activation of the two protomers by the mixed agonist BAM22 or by two receptor-specific agonists led to a selective activation of phospholipase C without any inhibition of the adenylyl cyclase pathway, indicating that the dual activation of the hetero-oligomer led to the activation of Gαq to the exclusion of Gαi/o. Such a protomer-exclusive signaling after receptor coactivation is unique for the δOR/SNSR-4 hetero-oligomer and indicates a dominant-negative effect of the SNSR-4 signaling on the δOR signaling. This intriguing regulation of hetero-oligomeric receptor activity could be due to three different mechanisms:
Competitive Antagonism. Agonist binding of one receptor protomer inhibits the agonist binding to the second protomer. In the case of the δOR/SNSR-4 hetero-oligomer, such competitive antagonism cannot account for the decreased efficacy of δOR agonists because [3H]DPDPE binding to the δOR was not inhibited by SNSR-4-specific agonists (Fig. 7). These findings further indicate that each of the protomer can bind agonist simultaneously within a hetero-oligomeric complex and that there is no apparent allosteric regulation between the agonistic binding sites of the two protomers.
Heterologous Cross-Talk. Activation of a signaling pathway by one receptor subtype desensitizes the second subtype. It has been reported that pretreatment of δOR expressing cells with PMA desensitized the agonist-promoted δOR activity and induced receptor internalization (Xiang et al., 2001). In our study, protein kinase C-dependent δOR desensitization (Fig. 8), but no internalization (data no shown) could be observed after PMA costimulation. However, the amplitude of the PMA induced-inhibition of δOR signaling (Fig. 8) was much smaller than that observed by the coactivation by either the bivalent ligand BAM22 or the concomitant activation with both δOR- and SNSR-selective agonists (Fig. 5A), making it unlikely that cross-talk desensitization mediated by protein kinase C could completely explain the trans-inhibition observed. Thus, although we cannot totally exclude that the protein kinase C-mediated cross-talk is partly involved in the inhibition of the Gαi/o coupling to the δOR after simultaneous coactivation of the SNSR-4, we suggest that other regulatory mechanisms also contribute to this inhibitory process.
G Protein Competition. The coupling of one receptor protomer to its cognate G protein inhibits the coupling of a second G protein to the hetero-oligomer. It has been proposed that rhodopsin and the leukotriene B4 receptor form oligomeric complexes that interact with only one trimeric G protein (Baneres and Parello, 2003; Filipek et al., 2004). A similar interaction of an oligomeric δOR/SNSR-4 complex with a single G protein would exclude simultaneous coupling of Gαi/o and Gαq to the hetero-oligomer. Assuming a preferential coupling of the hetero-oligomer to Gαq within this model, simultaneous activation of both receptors would lead to the Gαq coupling of the SNSR-4-protomer to the expense of the Gαi/o coupling. This process would lead to the substitution of Gαi/o by Gαq, resulting in the activation of the phospholipase C and the depression of the adenylyl cyclase pathway. In the case of a single δOR agonist, there would be no competition between Gαi/o and Gαq, explaining the high efficacy of δOR agonists to inhibit the cyclase after single activation. Consistent with this model, the δOR/SNSR-4 hetero-oligomer exhibits a lower ability to couple to Gαi/o compared with the δOR, even in the absence of SNSR-4 agonists, as indicated by the decreased potency of δOR agonists to activate the δOR/SNSR-4 hetero-oligomer despite their unchanged affinity to bind this complex (Fig. 6). Given the detectable amount of spontaneous SNSR-4 activity in δOR/SNSR-4 coexpressing cells (data not shown), the decreased affinity of the δOR/SNSR-4 hetero-oligomer in the absence of SNSR-4 agonist could be explained by the competition of Gαi/o and Gαq as a result of basal SNSR-4 activity.
The colocalization of δOR and SNSR-4 in dorsal root ganglia raises the intriguing possibility that hetero-oligomerization between these receptors may also occur in cells naturally expressing both receptors. Hetero-oligomerization between δOR and SNSR-4 in cells regulating pain perception would allow the δOR to mediate its antinociceptive effects when specifically activated by enkephalins. However, in a situation that requires the sensation of pain, coactivation of δOR and SNSR-4 would not only enhance the pain sensation by the pronociceptive effects of the SNSR-4 but also by the inhibition of the analgesic effects of the δOR. Such an agonist-dependent dominant-negative effect of the SNSR-4 on δOR would increase both the amplitude and the fine-tuning of pain sensation that may be advantageous in an evolutionary context. In this article, we provide data indicating that SNSR stimulation attenuates OR signaling in cultured rat dorsal root ganglia neurons. Although not proving the existence of δOR/SNSR-4 hetero-oligomers in native tissues, the recapitulation of the hetero-oligomer signaling property observed in HEK293 cells, in the dorsal root ganglia, supports the existence of such oligomers and warrants further investigation that would directly investigate these hetero-oligomers in dorsal root ganglias and their potential role controlling pain perception.
Acknowledgments
We are grateful to Drs. T. Groblewski and S. Ahmad (AstraZeneca Research Center) for the fruitful discussions and for providing various BAM peptides.
Footnotes
-
This work was supported by grants from the Canadian Institute for Health Research and from the Heart and Stroke Foundation of Canada. A.B. was supported by a postdoctoral fellowship from AstraZeneca Research Center, and K.G. and L.A.D. received National Institutes of Health grants NS026880 and DA008862, respectively. M.B. holds a Canada Research Chair in Signal Transduction and Molecular Pharmacology.
-
ABBREVIATIONS: SNSR, sensory neuron-specific G protein-coupled receptor; GPCR, G protein-coupled receptor; BAM, bovine adrenal medulla; OR, opioid receptor(s); BRET, bioluminescence resonance energy transfer; HEK, human embryonic kidney; DMEM, Dulbecco's modified Eagle's medium; IBMX, 3-isobutyl-1-methylxanthine; Leu-enk, Leu-enkephalin; DPDPE, [d-Pen2,d-Pen5]-enkephalin; GFP, green fluorescent protein; Rluc, Renilla reniformis luciferase; PCR, polymerase chain reaction; IPn, total fraction of phosphoinositols; BSA, bovine serum albumin; PMA, phorbol 12-myristate-13-acetate.
- Received January 24, 2006.
- Accepted May 8, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}