


Abstract
An increasing amount of ligand binding data on G protein-coupled receptors (GPCRs) is not compatible with the prediction of the simple mass action law. This may be related to the propensity of most GPCRs, if not all, to oligomerize. Indeed, one of the consequences of receptor oligomerization could be a possible cross-talk between the protomers, which in turn could lead to negative or positive cooperative ligand binding. We prove here that this can be demonstrated experimentally. Saturation, dissociation, and competition binding experiments were performed on vasopressin and oxytocin receptors expressed in Chinese hamster ovary or COS-7 cells. Linear, concave, and convex Scatchard plots were then obtained, depending on the ligand used. Moreover, some competition curves exhibited an increase of the radiotracer binding for low concentrations of competitors, suggesting a cooperative binding process. These data demonstrate that various vasopressin analogs display either positive or negative cooperative binding. Because positive cooperative binding cannot be explained without considering receptor as multivalent, these binding data support the concept of GPCR dimerization process. The results, which are in good accordance with the predictions of previous mathematical models, suggest that binding experiments can be used to probe the existence of receptor dimers.
G protein-coupled receptors (GPCRs) have typically been considered functional monomeric proteins, their activation resulting from the stoichiometric binding of one ligand per receptor. For the last 2 decades, however, pharmacological data obtained on various GPCR models are not compatible with such a model. For instance, negative cooperative binding has been reported for various class A GPCRs such as muscarinic receptors (Christopoulos and Kenakin, 2002; van Koppen and Kaiser, 2003) or dopamine receptors (Armstrong and Strange, 2001), and more recently for glycoprotein hormone receptors (Urizar et al., 2005) and chemokine receptors (El-Asmar et al., 2005). In addition, very few studies have reported positive cooperative binding, for instance on muscarinic receptors (Mattera et al., 1985; Wreggett and Wells, 1995) and opioid κ-δ heterodimers (Jordan and Devi, 1999). Finally, apparent variations in the maximal binding capacity on a given preparation (Wreggett and Wells, 1995; Serradeil-Le Gal et al., 1996; Armstrong and Strange, 2001) and discrepancies in the ligand affinity estimations, both depending on the radioligand used, have been described.
A large amount of data have shown that many GPCRs, if not all, have the propensity to oligomerize (Terrillon and Bouvier, 2004). One expected consequence of receptor oligomerization is a possible cross-talk between the protomers, which can lead to negative and positive cooperative ligand binding. Various theoretical models have considered receptors as multivalent complexes (Mattera et al., 1985; Wreggett and Wells, 1995; Armstrong and Strange, 2001; Durroux, 2005; Franco et al., 2005). Few of them, including ours, have analyzed the impact of receptor cross-talk within a dimer on ligand binding (Durroux, 2005; Franco et al., 2005). Predictions show that ligand-induced negative or positive cooperativities result in modifications of both shapes and slopes of saturation and competition binding curves (Durroux, 2005). As a consequence, cross-talk between protomers can also explain apparent variations in the maximal binding capacity in saturation experiments performed with various ligands on a given preparation and discrepancies in the ligand affinity estimations according to the radioligand used (Durroux, 2005).
Whereas theoretical models have considered different possibilities of cooperative binding, it has not yet been experimentally demonstrated that, for a given receptor, different selective ligands can bind with various cooperative processes. To investigate such a hypothesis, we performed saturation, dissociation, and competition binding experiments on vasopressin and oxytocin receptors expressed in COS-7 and CHO cells with various selective ligands. We focused more specifically on four peptidic ligands: the natural agonists vasopressin (AVP) and oxytocin (OT) and two antagonists, the linear vasopressin antagonist HO-LVA (Barberis et al., 1995) and the cyclic antagonist OTA (Elands et al., 1988b; Breton et al., 2001). We carefully examined the data, which were obviously not compatible with the predictions of a ligand/receptor interaction model that follows the simple mass action law.
We therefore demonstrate that negative and positive cooperative binding can be observed with different ligands on vasopressin and oxytocin receptors, further validating mathematical models and demonstrating that receptors dimerize.
Materials and Methods
Drugs. [3H]AVP (60-80 Ci/mmol) and [3H]OT (33 Ci/mmol) were from PerkinElmer Life and Analytical Sciences (Courtaboeuf, France) Products. 4-OH-Phenylacetyl-d-Tyr(Me)2,Phe3,Gln4,Asn5,Arg6,Pro7,Arg8-NH92, (HO-LVA) (Barberis et al., 1995), d(CH2)5-[Tyr(Me)2,Thr4,Orn8,Tyr-NH92]vasotocin (OTA) (Elands et al., 1988b), d(CH2)5 [d-Thi2,Thr4,Orn8,Eda9←Tyr10]vasotocin (Manning et al., 2001), and [Phe2,Orn8]vasotocin were synthesized in the laboratory of author M. Manning. We performed radioiodinations of HO-LVA and OTA to give 125I-HO-LVA and 125I-OTA, respectively, as described previously (Elands et al., 1988a; Barberis et al., 1995). GTPγS was from Sigma-Aldrich (Saint-Quentin Fallavier, France).
Cell Culture. The CHO cell lines that stably express the human vasopressin V1a or V1b receptors or the human oxytocin receptor were maintained in culture in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum and 100 units/ml penicillin and streptomycin in an atmosphere of 95% air and 5% CO2 at 37°C.
COS-7 cells were transiently transfected by electroporation as described previously (Cotte et al., 2000). In brief, electroporation was performed with 1 μg of a pRK5-containing hemagglutinin (HA)-tagged or 6-histidine (6His)-tagged human V1a receptor and 9 μg of empty vector. Membrane preparations were made as follows.
Membrane Preparation. Culture dishes of CHO or COS-7 cells expressing the human vasopressin V1a,V1b, or oxytocin receptors were washed twice in phosphate-buffered saline without calcium and magnesium and ice-cold lysis buffer (15 mM Tris-HCl, 2 mM MgCl2, and 0.3 mM EDTA, pH 7.4) was added. Cells were scraped with a rubber policeman, homogenized with a Ultra-Turrax homogenizer (Janke-Kunkel IKA-Labortechnik, Staufen, Germany), and centrifuged at 100g for 5 min at 4°C. Supernatants were recovered and centrifuged at 44,000g for 30 min at 4°C. Pellets were resuspended in a suspension medium (50 mM Tris-HCl and 5 mM MgCl2, pH 7.4) and centrifuged at 44,000g for 30 min at 4°C. Pellets were resuspended in an appropriate volume of the same buffer. For each membrane preparation, protein content was evaluated, and membranes were then aliquoted and frozen in liquid nitrogen.
Homogeneous Time-Resolved Fluorescence-Fluorescence Resonance Energy Transfer Assays. Homogeneous time-resolved fluorescence-FRET experiments were performed as described previously (Maurel et al., 2004). These experiments involve the transfer of energy from a europium cryptate pyridine-bipyridine as the donor fluorophore to an Alexa Fluor 647 as the acceptor fluorophore. We used the monoclonal anti-6His antibody and the monoclonal anti-HA antibody (12CA5) labeled with the different fluorophores kindly provided by Eric Trinquet and Hervé Ansanay (Cis Bio International, Marcoule, France). After transfection, 10 × 104 COS-7 cells/well were dispatched into a black 96-well assay plate (Costar; Corning Life Sciences, Acton, MA) in 100 μl of Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum. Twenty-four hours later, COS-7 cells or membrane preparations (25 μg/well) expressing the indicated 6His- or HA-tagged human V1a receptor were incubated in 100 μl of Tris/Krebs buffer (20 mM Tris-HCl, pH 7.4, 118 mM NaCl, 5.6 mM glucose, 1.2 mM KH2PO4, 1.2 mM MgSO4, 4.7 mM KCl, and 1.8 mM CaCl2) supplemented with 0.1% bovine serum albumin and containing 1 nM europium cryptate pyridine-bipyridine and 1 nM Alexa Fluor 647-labeled antibodies. As a negative control, COS-7 cells or membranes were incubated with only the donor fluorophore-labeled antibody. After an 8-h incubation at 4°C, fluorescence emissions were measured both at 620 nm and at 665 nm on a RubyStar fluorometer (BMG Labtechnologies) equipped with a nitrogen laser as excitation source (337 nm). A 400-μs reading was measured after a 50-μs delay to eliminate the short-lived fluorescence background from the specific signal. The fluorescence intensities measured at 620 nm and at 665 nm correspond to the total europium cryptate emission and to the FRET signal, respectively. The specific signal was calculated using the following equation: ΔF = (R - Rneg)/(Rneg). R is the ratio (fluorescence 665 nm/fluorescence 620 nm) × 104 calculated in wells incubated with both donor- and acceptor-labeled antibodies, whereas Rneg is the same ratio for the negative control.
Binding Assay. For saturation experiments, affinities of 125I-HO-LVA and 125I-OTA were determined from saturation experiments. Membranes (1-3 μg/assay) were incubated with increasing concentrations of radioactive tracer (5-500 pM for 125I-HO-LVA and 20 pM to 2 nM for 125I-OTA) for 1 h at 30°C. For each concentration of tracer, nonspecific binding was determined by the addition of an excess of HO-LVA or OTA, respectively. Bound ligand fractions were separated from the free ligand either by filtration or centrifugation. When performed by filtration, we used Whatman GF-C filters (Whatman, Maidstone, UK) preincubated in polyethylenimine (5 μl/ml for 125I-HO-LVA) or bovine serum albumin (10 mg/ml for 125I-OTA). Filtration was performed on a Brandel apparatus (Brandel Inc., Gaithersburg, MD). Radioactivity on the filter was counted on a Packard Cobra gamma counter (PerkinElmer Life and Analytical Sciences). Each assay was performed in triplicate. When centrifugation was used, 1 ml of ice-cold medium containing 100 mM Tris-HCl, 10 mM MgCl2, and 1 mg/ml bovine serum albumin, pH 7.4 was added to the assay at the end of the incubation. Samples were centrifuged at 22,000g for 5 min and supernatants were discarded. Pellets were washed with 1 ml of the same medium, and centrifuged at 22,000g for another 5 min. Supernatants were removed, and radioactivity in the pellet was counted on a Cobra gamma counter as explained above.
Affinities of [3H]AVP and [3H]OT were determined from saturation and homologous competition experiments. CHO membranes (8-15 μg) were incubated with [3H]AVP or [3H]OT (1-2 nM) plus increasing concentrations of AVP or OT (1 pM to 1 μM). Nonspecific binding was determined by the addition of a large excess of AVP or OT (1 μM). Bound and free ligand fractions were separated by filtration or centrifugation as mentioned above. Each assay was performed in triplicate. Radioactivity was counted on a beta-counter (Packard).
For competition experiments, membranes were incubated with 125I-HO-LVA (50-80 pM) or [3H]AVP (1-2 nM), and increasing concentrations of unlabeled ligand ranging from 1 pM to 1 μM. Nonspecific binding was determined with an excess of HO-LVA (1 μM) or AVP (1 μM). Bound radioactivity was determined as described above.
All binding data were analyzed with the program Kell (Biosoft, Cambridge, UK). Fitting parameters for saturation experiments were determined using a nonlinear curve-fitting routine to the Hill equation B = Bmax [1 + (Kd/[L])nH]-1 where Bmax is the maximal binding, [L] is the concentration of labeled ligand, Kd is the equilibrium dissociation constant for the labeled ligand, and nH is the Hill coefficient (Segel, 1975). Because of a potential cross talk between protomers within a dimer, the Hill coefficient was allowed to vary.
For dissociation experiments, membranes were preincubated in a volume of 100 μl for 40 min at 30°C in the presence of [3H]AVP or [3H]OT (0.5-1.5 nM). Three milliliters of incubation medium (50 mM Tris, 5 mM MgCl2, and 1 bovine serum albumin mg/ml) with or without unlabeled AVP or OT (1 or 10 μM) were then added at different times. The addition of 3 ml of incubation medium induces a tracer dilution with a factor of 31. We checked that in such new equilibrium conditions, less than 10% of the binding sites were still able to be labeled by the tracer. The data are presented as the percentage of specific binding at time 0 of dissociation (B0). At each dissociation time, bound radioactivity was determined as described above. The dissociation rate, k-1, is determined by the slope of the curve when fitting the data with the equation Ln [B/B0] = k-1t, in which t is time, B is the specific binding at time t, B0 is the specific binding at time 0 of dissociation.
Results
Predictions from Mathematical Models. A few mathematical models have been proposed to describe ligand binding on dimeric receptors (Mattera et al., 1985; Wreggett and Wells, 1995; Christopoulos and Kenakin, 2002; Durroux, 2005; Franco et al., 2005). Consequences of a positive or a negative cooperative ligand binding between the binding sites of protomers within a dimer have been analyzed (Durroux, 2005). As illustrated in Fig. 1, the profiles of the saturation and competition curves depend on the existence and on the nature of the cooperativity between the two binding sites of a dimer.
The predicted saturation curves obtained with a radioligand L1* (middle, L1 = L2) exhibit Hill coefficient of 1 and a linear Scatchard curve in the absence of any cooperativity (Fig. 1a). By contrast, negative (Fig. 1b) and positive (Fig. 1c) cooperative bindings result in saturation curves with slope factors less or greater than 1, respectively and in concave and convex Scatchard plot, respectively.
In addition, predicted competition curves obtained with a radioligand L1* and a competitor L2 different of L1 display slopes of 1 in the absence of any cooperative binding (Fig. 1a, bottom). By contrast, negative and positive cooperativities result in curves with slope factors less or greater than 1, respectively. Moreover, the patterns of the competition curves can be complex; curves that exhibit two plateaus (Fig. 1b) or an increase in the binding of the tracer for low concentrations of competitors (Fig. 1c) can be observed for negative or positive cooperative binding, respectively. An unexpected consequence of the models is that the ligand affinity estimation can be different depending on the radiotracer used to perform competition experiments (Durroux, 2005). To validate the model, we performed binding experiments on membrane preparations from cells expressing vasopressin and the oxytocin receptors.

Outlines of the predictions of the mathematical model: Binding of ligands on dimeric receptor is considered in three different contexts: the ligands L1 and L2 bind to the binding sites without any cooperativity (a), with a negative (b) or a positive (c) cooperativity. Middle, L1 and L2 are identical and the conditions correspond to saturation experiments. The saturation and Scatchard curves (inset) exhibit Hill coefficient equal to 1 (a), lower than 1 (b), or greater than 1 (c). Bottom, L1 and L2 are assumed to be different, and the resulting competition curves are illustrated in the three cooperativity contexts. The absence of cooperativity results in curve with a slope factor equal to 1, whereas negative and positive cooperativities result in curves with slope factors lower or greater than 1, respectively. It is noteworthy that the intermediate plateau in b can be more or less marked according to the parameter used for the mathematical simulation. In the same way, the amplitude of the hump in c, which corresponds to an increase of the binding of the tracer, is variable according to the parameters used in the simulation.
FRET Experiments Prove the Existence of Vasopressin Receptor Dimers. To demonstrate the presence of dimers at the cell surface and in membrane preparations, we performed homogeneous time-resolved FRET experiments as described previously by Maurel et al. (2004). In this assay, vasopressin receptors were tagged with HA or 6His sequences, and FRET signals were measured on intact cells or on membrane preparations with anti-HA and/or anti-6His monoclonal antibodies labeled with the fluorescent probes europium cryptate-pyridine bipyridine or Alexa Fluor 647.
First, cells expressing HA-tagged vasopressin V1a receptors were incubated with europium cryptate-pyridine bipyridine and Alexa Fluor 647 labeled anti-HA monoclonal antibodies. A FRET signal was measured at 665 nm (the emission wavelength of Alexa Fluor 647) after excitation of europium cryptate-pyridine bipyridine at 337 nm (Fig. 2, white bars). This signal was significantly higher than that measured in mock-transfected cells, indicating that HA-tagged vasopressin V1a receptors homodimerized.
Second, we performed FRET experiments on membrane preparations. A FRET signal was recorded when membranes that coexpressed HA- and 6His-tagged V1a receptors [V1a (HA)/V1a (6His)] were incubated with Alexa Fluor 647 labeled anti-HA and europium cryptate-pyridine bipyridine labeled anti-6His monoclonal antibodies (Fig. 2, black bars). By contrast, no significant signal was recorded from mock membrane preparations or from a membrane preparation mix [mix V1a (HA) + V1a (6His)] which expressed either HA-tagged or 6His-tagged V1a receptors. The lack of FRET signal on the membrane mix was not due to a too-low expression level of receptors. Indeed, a specific FRET signal was detected when each membrane preparation that either expressed HA-tagged [V1a (HA)] or 6His-tagged [V1a (6His)] V1a receptors were incubated with europium cryptate-pyridine bipyridine and Alexa Fluor 647-labeled anti-tag monoclonal antibodies. Then, these data indicated that the FRET signal recorded on membranes that coexpressed HA- and 6Histagged V1a receptors [V1a (HA)/V1a (6His)] was specific and reflected receptor dimerization. Similar results have been obtained with oxytocin receptors (data not shown).
Saturation Assays Display Ligand Cooperative Binding on Vasopressin Receptor. We carried out saturation and homologous competition experiments with tritiated vasopressin or tritiated oxytocin from CHO cells expressing either V1a or V1b or oxytocin receptors. As illustrated in Fig. 3, all saturation experiments exhibited curves with a slope factor less than 1 and a concave Scatchard plot (Fig. 3, insets). The fits of the experimental curves with a Hill coefficient that was allowed to vary (i.e., when considering a potential cooperativity between the binding sites) are significantly better than those obtained when only one binding site type was considered (absence of cooperative binding). The Hill coefficient and the Kd values with [3H]AVP are 0.45 and 85 nM for the V1a receptor (Fig. 3a) and 0.27 and 85 nM for the V1b receptor (Fig. 3b). The fitting procedures gave a Hill coefficient of 0.38 and an affinity constant of 129 nM for the oxytocin receptor (Fig. 3c).

Time-resolved FRET experiments performed on COS-7 cells expressing tagged V1a receptors. FRET experiments on cell surface-targeted receptors (white bars): cells expressing HA-tagged V1a receptors or not (mock) were incubated with europium cryptate-pyridine bipyridine and Alexa Fluor 647 labeled anti-HA monoclonal antibodies as described under Materials and Methods. FRET experiments on total receptors expressed in COS-7 cell membranes (black bars): membranes expressing tagged V1a receptors were incubated with europium cryptate-pyridine bipyridine and Alexa Fluor 647 labeled anti-tag monoclonal antibodies as indicated in the figure. The condition V1a (HA)/V1a (6His) corresponds to membranes in which HA-tagged V1a and 6His-tagged V1a receptors are coexpressed; mix V1a (HA) + V1a (6His) corresponds to a mix of membranes that expressed either HA- or 6His-tagged V1a receptors. The conditions V1a (HA) and V1a (6His) correspond to membranes that expressed only HA- or 6His-tagged V1a receptor, respectively. The FRET signal was measured at 665 nm after excitation at 337 nm. The illustrated experiments are representative of at least three experiments. Values are means ± S.D. of triplicates. The FRET signal (ΔF%) was calculated as described under Materials and Methods.
Because these results can be an argument in favor of the existence of a negative cooperativity between binding sites or in favor of the coexistence of two receptors conformations with different affinities, we also investigated negative cooperative using dissociation kinetics experiments (De Meyts, 1976; Christopoulos et al., 1997; Urizar et al., 2005; Springael et al., 2006). The results illustrated in Fig. 4 show that the dissociation rates of [3H]AVP on the vasopressin V1b (Fig. 4a) and [3H]OT on the oxytocin receptor (Fig. 4b) are promoted by an excess of unlabeled AVP (1 μM) or OT (1 μM). The fits of the data on a semilogarithmic plot (Fig. 4, insets) provide the dissociation rates of [3H]AVP on the vasopressin V1b of 0.0024 min-1 and 0.0063 min-1 without or with an excess of unlabeled vasopressin, respectively (Fig. 4a, inset). Likewise, the dissociation rates of [3H]OT on the oxytocin receptor are 0.0168 min-1 and 0.0067 min-1 in the absence or the presence of an excess of unlabeled oxytocin (Fig. 4b, inset). Moreover, because AVP has been described as a partial agonist for oxytocin receptor (Chini et al., 1996), we also performed dissociation experiments with [3H]AVP (Fig. 4c) and showed that, as for OT, the dissociation rate is multiplied by 2.6 in the presence of an excess of unlabeled vasopressin (Fig. 4c, inset).
In the next step, we respectively carried out saturation experiments with iodinated antagonists and the vasopressin V1a receptor. Typical results obtained with two antagonists are illustrated in Fig. 5. Figure 5a illustrates a saturation curve obtained with 125I-HO-LVA, a vasopressin antagonist (Barberis et al., 1995) on membranes from CHO cells expressing V1a receptors, and the resulting Scatchard plot is almost linear (Fig. 5a, inset, continuous line) with a Hill coefficient of 0.97 and a dissociation constant, Kd, of 22pM. Fitting the data when considering a potential cooperative binding or not, did not significantly change the Kd and Hill coefficient values (Table 1).
Affinities and Hill coefficients determined after fitting the saturation and Scatchard curves when performing saturation experiments with 25I-HO-LVA, [3H]AVP, and 125I-OTA on membranes of CHO cells expressing human V1a receptors
Two data analyses were performed. The first one (“cooperative binding site” analysis) took into account a possible cooperativity between ligand binding sites; the second (independent binding site analysis) considered binding sites as independent and did not consider any potential cooperativity (the Hill coefficient nH is equal to 1). The right column indicates the number of experiments.
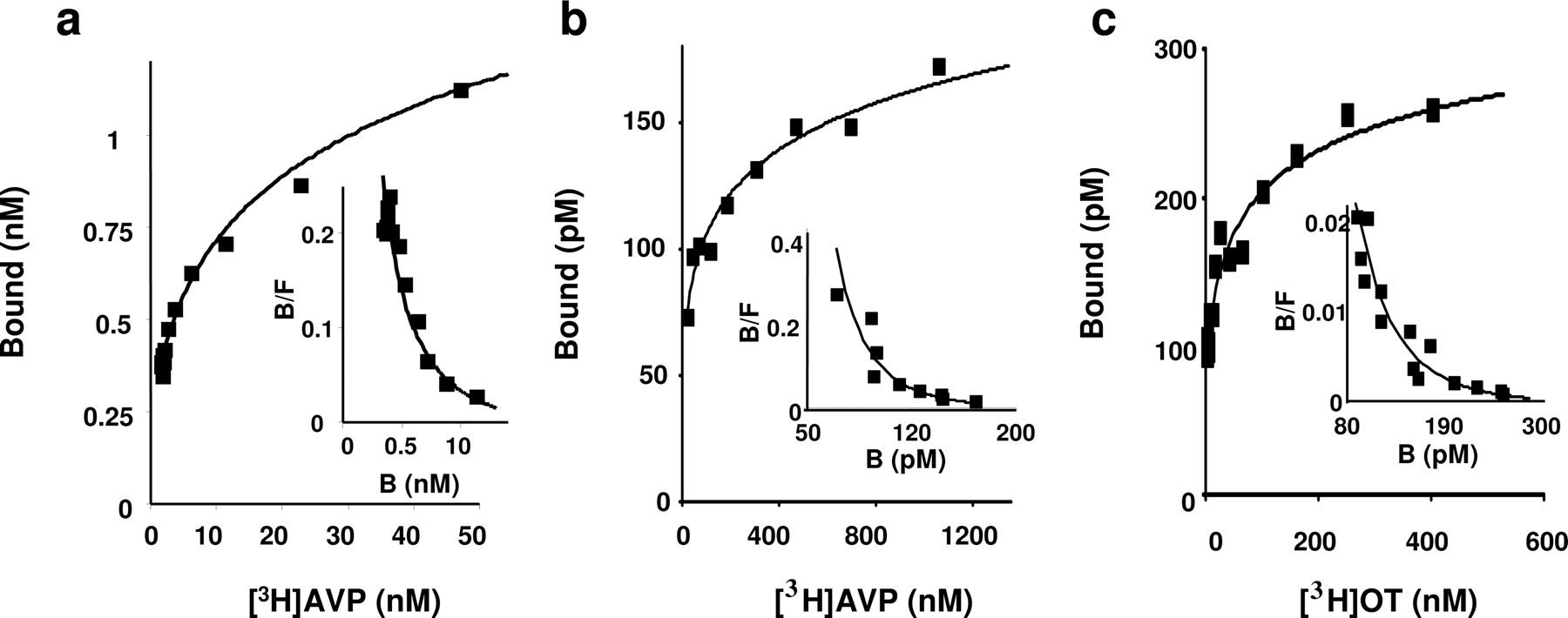
Saturation experiments performed with [3H]AVP and [3H]OT on CHO cells expressing vasopressin and oxytocin receptors. Saturation and homologous competition experiments with [3H]AVP (a and b, respectively), and homologous competition experiments with [3H]OT (c) were performed on membranes of CHO expressing V1a (a), V1b (b), or OT (c) (10-15 μg protein/assay). Each curve is representative of at least three independent experiments performed in triplicate samples. Values are means ± S.E. of triplicates. Data were fitted using the nonlinear curve-fitting routine of the computer software Kell (Biosoft) to the Hill equation: B = Bmax [1 + (Kd/[L])nH]-1, where Bmax is the maximal binding, [L] is the concentration of labeled ligand, Kd is the equilibrium dissociation constant of the labeled ligand, and nH is the Hill coefficient. Insets, Scatchard plots obtained from the corresponding saturation curves. The fitting parameters obtained for Kd and n are 85 nM and 0.45 (a) 85 nM and 0.27 (b), and 129 nM and 0.38 (c) for the V1a,V1b, and oxytocin receptors, respectively.
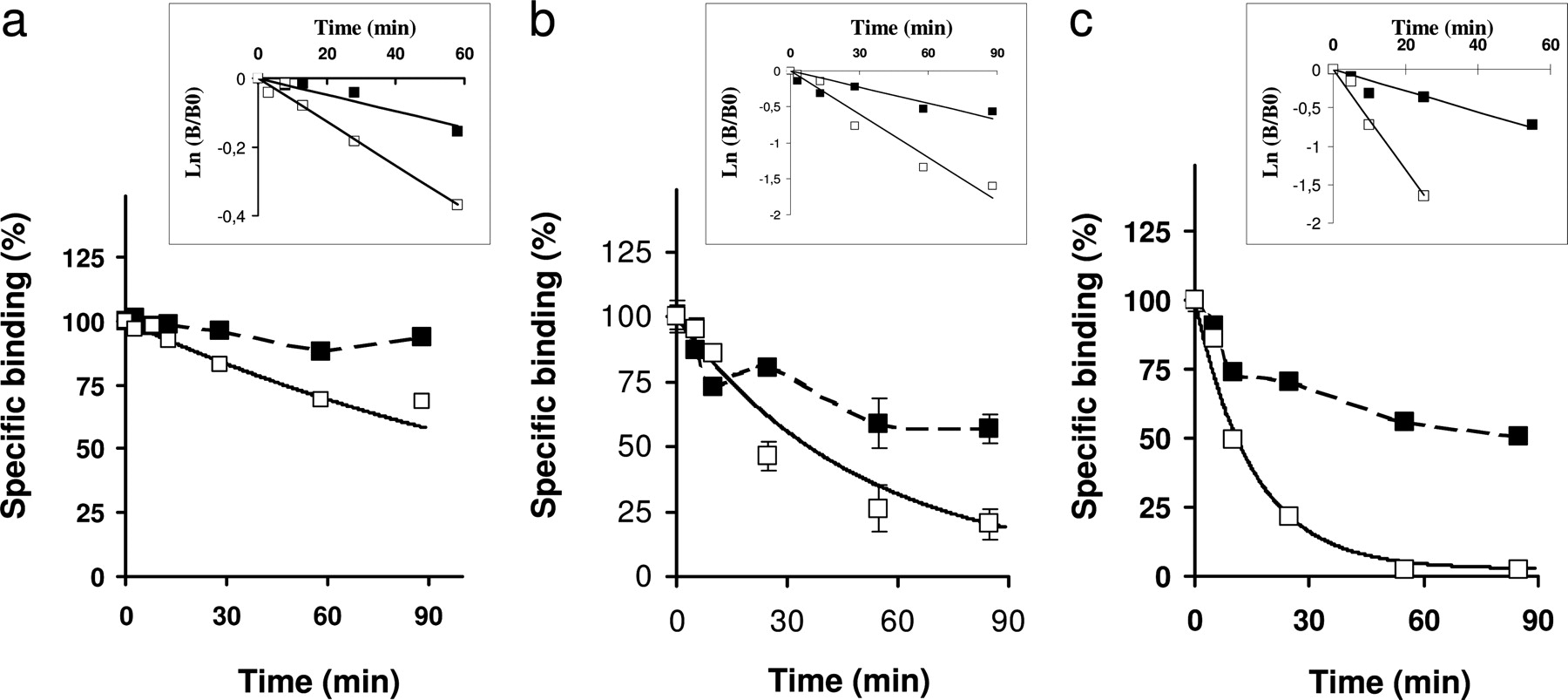
Dissociation of [3H]AVP and [3H]OT from vasopressin and oxytocin receptors expressed in CHO cells. Membranes were incubated for 40 min at 30°C with [3H]AVP (a and c) or [3H]OT (b) (0.5-1.5 nM). At any given time, 3 ml of incubation medium containing (or not) an excess of vasopressin (1 μM) (a and c) or oxytocin (1 μM) (b) were added. The bound fractions were determined by filtration. Data are presented as the percentage of specific binding at time 0 of dissociation. Each curve is representative of at least three independent experiments performed in triplicate samples. Values are means ± S.E. of triplicates. Insets, to determine dissociation rates, data are presented in a semilogarithmic plots. The dissociation rate, k-1, is given by the slope of the curve when fitting the data with the equation Ln [B/B0] = k-1t in which t is time, B is the specific binding at time t, and B0 is the specific binding at time 0 of dissociation. The values of the dissociation rates in the absence or the presence of an excess of unlabeled ligand are, respectively, 0.0024 min-1 and 0.0063 min-1 (a), 0.0168 min-1 and 0.067 min-1 (b), and 0.0076 min-1 and 0.0201 min-1 (c).
On the contrary, a saturation experiment performed with and 125I-OTA, a mixed vasopressin V1a/oxytocin receptor antagonist (Elands et al., 1988b; Breton et al., 2001) exhibited a curve with slope factors greater than 1 (Fig. 5b) and a convex Scatchard plot (Fig. 5b, inset). The best fit was obtained when considering cooperativity between the binding sites and gave Hill coefficient and a Kd values of 1.29 and 302 pM, respectively. Mean values for the Hill coefficient and the dissociation constant obtained from seven independent experiments are 1.2 ± 0.035 and 0.35 ± 0.13 nM, respectively (Table 1).
The Kd estimations for the binding of 125I-HO-LVA, [3H]AVP, and 125I-OTA obtained when considering the absence of cooperative binding (Table 1, “independent binding site” analysis) are in the same range than those reported previously (Mouillac et al., 1995a; Breton et al., 2001). It is noteworthy that the differences between the Kd estimations given by the two analyses are obviously negligible when the Hill coefficient is approximately 1 (Table 1), but can be large when the Hill coefficient is different from 1. Moreover the nature of the cooperative binding is ligand-dependent.
Competition Assays Exhibit Ligand Cooperative Binding on Vasopressin Receptor. We carried out competition experiments with various ligands on CHO cells that expressed V1a receptors, and we observed ligand binding that is not compatible with the classic model of ligand binding on a receptor (i.e., a single ligand binds to a monomeric receptor).
First, we performed competition experiments between 125I-HO-LVA and vasopressin on CHO cell membranes expressing human V1a receptors. Slope factor values are below 1 and are frequently between 0.7 and 0.8, suggesting a negative cooperativity between the binding sites. Typical experimental data are presented in Fig. 6a and are fitted with the following equation: specific binding = Bg + [(Bmax - Bg)/(1 + ([L]/IC50)nH)], in which Bg corresponds to the background binding, Bmax is the maximal binding, [L] is the ligand concentration, IC50 is the concentration of a competitor that induces a decrease of 50% of the specific binding of the labeled ligand, and nH is the Hill coefficient. The gray curve corrresponds to the best fit obtained with the equation in which n is set to 1, assuming that no cooperativity exits between the binding sites. The black curve has been obtained with the same equation but the Hill coefficient is allowed to vary. The best fit is obtained with a n equal to 0.75.

Saturation experiments performed with iodinated antagonists on CHO cells expressing vasopressin V1a receptors. Saturation experiments with 125I-HO-LVA (a) and 125I-OTA (b) were performed on membranes of CHO cells expressing the human vasopressin V1a receptor. Each curve is representative of at least three independent experiments performed in triplicate samples. Values are means ± S.E. of triplicates. Data were fitted using the nonlinear curve-fitting routine of the computer software Kell (Biosoft) to the Hill equation: B = Bmax [1 + (Kd/[L])nH]-1, where Bmax is the maximal binding, [L] is the concentration of labeled ligand, Kd is the equilibrium dissociation constant of the labeled ligand, and nH is the Hill coefficient. Insets, Scatchard plots obtained from the corresponding saturation curves. The fitting parameters obtained for Bmax, Kd, and nH are 0.0126 nM, 0.022 nM, and 0.97 (a) and 0.044 nM, 0.302 nM, and 1.29 (b), respectively.
Second, competition experiments were performed with 125I-HO-LVA as radioligand and an analog of OTA, d(CH2)8[d-Thi2,Thr4,Orn8 ,Eda9←Tyr10]vasotocin, which exhibits potent oxytocic antagonist effects (Manning et al., 2001, 2005). We observed an increase in the binding of 125I-HO-LVA for low concentrations of the competitor. Indeed, low concentrations of d(CH2)5 [d-Thi2,Thr4,Orn8,Eda9← Tyr10]vasotocin induced a 25% increase of 125I-HO-LVA binding (Fig. 6b). The mean amplitude of the increase calculated from three independent experiments was 15% ± 5. Although this increase was generally of small amplitude (i.e., it did not exceed 20% of the binding of the tracer in the absence of competitor), it was nevertheless frequently observed with [Phe2,Orn8]vasotocin, a vasopressin V1a agonist (Berde et al., 1964) and numerous other analogs on membranes of CHO cells expressing human V1a receptors.
Third, we observed that the estimation of the affinity of a ligand for a receptor can vary depending on the method of its evaluation. As reported in Table 2, when considering the classic model of ligand binding, vasopressin had a Kd value of 0.7 ± 0.2 nM when estimated by saturation and a Ki value of 3.4 ± 1.1 nM for the human vasopressin V1a receptor (Cotte et al., 2000) expressed in COS-7 cells, when measured by competition experiments with 125I-HO-LVA as radiolabeled ligand (Table 2). This discrepancy can be greater in some cases. For instance, when experiments are performed on the Q185A V1a mutant, which displays a loss of affinity for agonists and antagonists (Cotte et al., 2000), the Kd and Ki values are 46 ± 7 nM and 810 ± 148 nM, respectively (Table 2). Such discrepancies have also been observed for two other mutations on rat vasopressin V1a receptor, K128A and Q131A, which have been shown to be involved in the binding site of ligands. For the mutated K128A and Q131A vasopressin V1a receptors, vasopressin has Kd values of 1.2 and 11.4 nM, respectively, and Ki values of 266 ± 108 and 168 ± 87 nM, respectively, when estimated by displacement of 125I-HO-LVA (Barberis et al., 1993; Mouillac et al., 1995a) (Table 2). Such discrepancies in Ki estimations have also been reported with the vasopressin agonist [Phe2,Orn8]VT depending on whether the competition experiments were performed with [3H]AVP or 125I-HO-LVA (Barberis et al., 1993; Mouillac et al., 1995a).
Affinities of AVP determined by saturation experiments (Kd) or by competition experiments (Ki) with 125I-HO-LVA as radiotracer on membranes of COS-7 cells expressing vasopressin V1a receptor
Lack of Effects of GTPγS. We then verified whether cooperative binding could be due to the coupling of V1a receptor to G proteins. Two studies have reported that the cooperative binding on myocardial muscarinic receptor are GTP-dependent (Mattera et al., 1985; Wreggett and Wells, 1995). On membrane preparations of CHO cells expressing the V1a receptors, addition of GTPγS (200 μM), a nonhydrolyzable analog of GTP, did not significantly modified the profiles of saturation and Scatchard curves (Fig. 7).
Discussion
In this study, we provide evidences that different ligands bind to vasopressin and oxytocin receptors by various cooperative processes. We showed that 125I-HO-LVA binding results in a saturation plot with a Hill coefficient of 1, suggesting that it binds to one class of binding sites. By contrast, [3H]AVP binding exhibits a saturation curve with a Hill coefficient significantly lower than 1, suggesting the existence of various binding sites. The heterogeneity in the binding sites might correspond to conformations coupled or not to G protein, although we cannot observe a significant effect of GTPγS, a nonhydrolyzable GTP analog, on vasopressin binding.
Dissociation kinetic experiments, which are an established procedure by which to demonstrate negative cooperativity (De Meyts, 1976; Christopoulos et al., 1997; Urizar et al., 2005; Springael et al., 2006), indicated that an excess of vasopressin or oxytocin promotes the dissociation of [3H]AVP or [3H]OT bound to the vasopressin and oxytocin receptors. The increases in the dissociation rates are not large compared with those reported for the chemokine receptors (Springael et al., 2006). Two different hypotheses can be formulated to explain these results: 1) there is a very strong negative cooperativity between the binding sites within a homodimer. The second ligand molecule has then a very low affinity for the second binding site (greater than 1 μM) and an excess of ligand at 1 or 10 μM is not enough to induce an important increase in the dissociation rate; 2) the receptor exists under different conformations, and dimers in which there is negative cross-talk between binding sites constitute one of these conformations.

Competition experiments performed on CHO cells expressing V1a receptor. a, competition experiments between [125I]HO-LVA and AVP on membranes of CHO cells expressing human vasopressin V1a receptor. Membranes were incubated in the presence of 60 pM 125I-HO-LVA and increasing concentration of AVP (1 pM to 1 μM) for 1 h at 30°C. Bound radioactivity was separated from the free ligand fraction by filtration. Experimental data (▪) are fitted using the equation: Bound = Bg + ((Bmax - Bg)/(1+ ([L]/IC50)nH)) with nH set to 1 (gray curve) or when nH is allowed to vary (black curve). b, competition experiments between the antagonist 125I-HO-LVA and the agonist d(CH2)5[d-Thi2,Thr4,Orn8,Eda9←Tyr10]vasotocin, a structural analog of vasotocin on membranes of CHO cells expressing human V1a receptors. Membranes were incubated in the presence of 60 pM [125I] HO-LVA and increasing concentrations of d(CH2)5[d-Thi2,Thr4,Orn8,Eda9←Tyr10]vasotocin (1 pM to 1 μM) for 1 h at 30°C. Bound radioactivity was separated from the free ligand fraction by filtration. Each curve is representative of at least three independent experiments performed in triplicate samples. Values are means ± S.E of triplicates.
However, the hypothesis of the existence of a binding site heterogeneity is not compatible with the positive cooperative bindings revealed 1) by convex saturation curve with the antagonist 125I-OTA and 2) by bell-shaped competition curve with 125I-HO-LVA and d(CH2)5[d-Thi2,Thr4,Orn8, Eda9←Tyr10]vasotocin or [Phe2,Orn8]vasotocin. Similar positive cooperative bindings have only been reported for the muscarinic receptor (Mattera et al., 1985; Wreggett and Wells, 1995).
Cooperative ligand binding on GPCRs has been extensively reviewed (Christopoulos and Kenakin, 2002; Schwartz and Holst, 2006). By contrast to negative cooperativity, positive cooperativities imply that the receptor should behave as a multivalent complex (Mattera et al., 1985; Wreggett and Wells, 1995; Christopoulos and Kenakin, 2002). However, the binding of two 125I-OTA ligands in the same pocket or in neighboring pockets on a single vasopressin V1a monomer seems to be unlikely because computational modelization of the vasopressin V1a receptor, based on receptor mutagenesis approaches (Mouillac et al., 1995a; Mouillac et al., 1995b) and on photolabeling strategies (Phalipou et al., 1997, 1999; Breton et al., 2001), has predicted that only one vasopressin or its analogs are buried in a binding pocket defined by the seven transmembrane domains of the receptor (Mouillac et al., 1995a).
On the contrary, the binding of two ligands on the two protomers within a dimer is highly probable. Using the FRET strategy described previously (Liu et al., 2004; Maurel et al., 2004), we have shown that vasopressin receptors in our model are able to dimerize and that dimers are present at the cell surface. It confirms the results previously obtained (Terrillon et al., 2003). Moreover the binding data reported in the present article are in accordance with the predictions from the mathematical models (Durroux, 2005) and more specifically for what concerns positive cooperativity. 125I-OTA binds to the V1a receptor in a positive cooperative manner. As mentioned above, this can only be explained by considering the binding complex as multivalent. Binding of 125I-OTA on one protomer should promote the binding of a second ligand on the second protomer. Although 125I-HO-LVA is a peptidic vasopressin V1a antagonist as 125I-OTA, no significant positive or negative cooperativity has been observed for its binding, suggesting that the cooperative binding does not exclusively depend on the agonistic or antagonistic nature of the ligand.
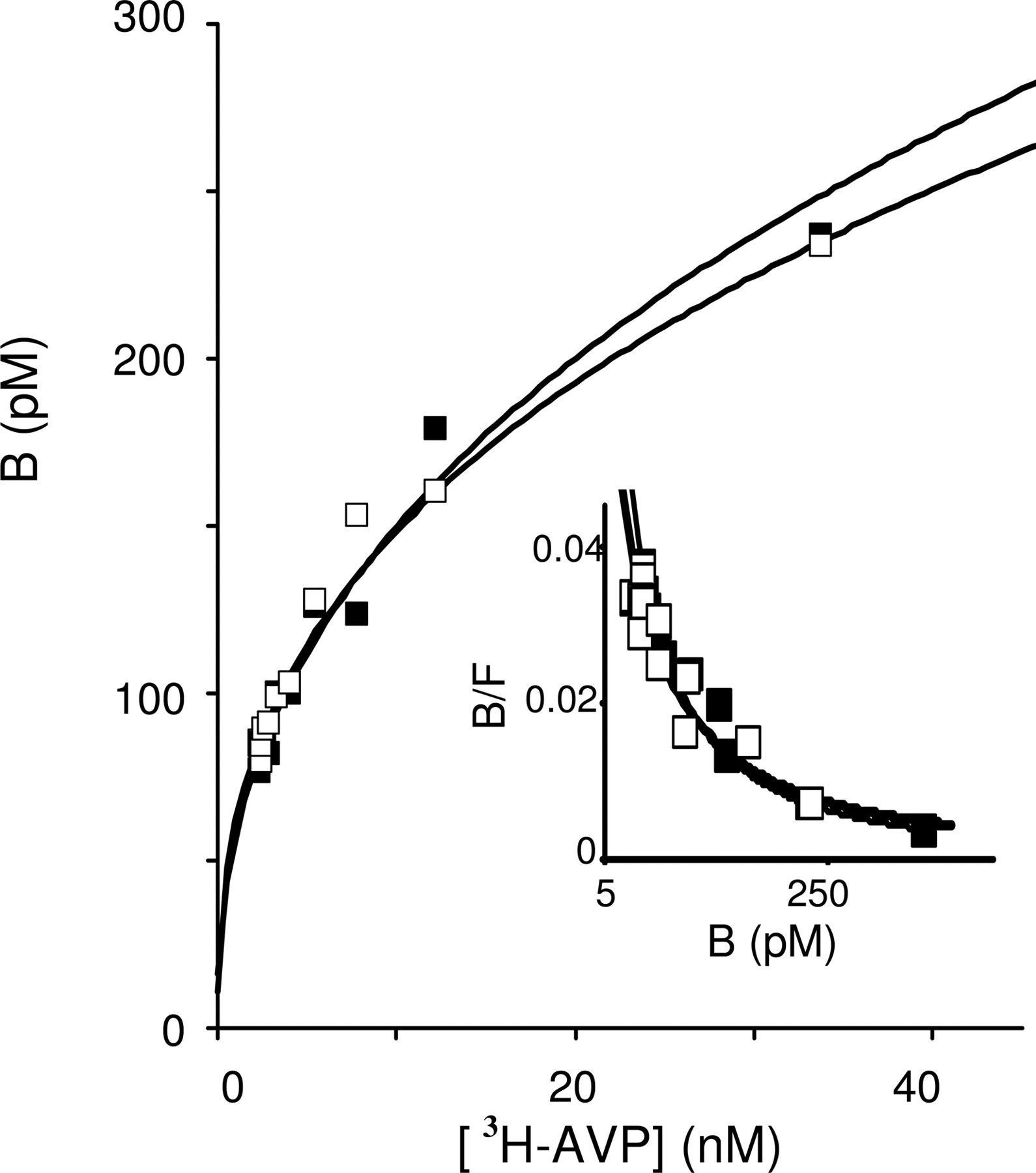
Absence of any effect of GTPγS on binding of [3H]AVP. Homologous competition experiments with [3H]AVP were performed on membranes of CHO cells expressing V1a receptors which exhibit cooperative binding. Membranes (8-15 μg per assay) were incubated with (▪) or without (□) GTPγS (100 μM). Each curve is representative of at least three independent experiments. Values are means ± S.E. of triplicates.
By contrast, the curvilinear saturation curve with [3H]AVP and [3H]OT can result from a negative cooperativity between the protomers within vasopressin and oxytocin receptor homodimers. It is noteworthy that this negative cooperativity can be at the origin of the variation of the maximal binding depending on the ligand used to perform the saturation experiments (Serradeil-Le Gal et al., 1996). Because the linear part of the saturation curve obtained with [3H]AVP is often considered to fit the Scatchard curve, the resulting maximal binding, which in fact corresponds to the saturation of the higher affinity binding site, is underestimated. However, the determination of the maximal binding when considering the whole curve remains difficult, because the precision of the last points of the curve is low.
In addition, we observed negative cooperative bindings with vasopressin and oxytocin, the two natural agonists, on V1b and OT receptors when expressed in COS-7 or CHO cell lines. Strong negative cooperative agonist bindings on glycoprotein hormone receptor (Urizar et al., 2005) and on chemokine receptors (El-Asmar et al., 2005) have been associated to the propensity of the receptors to dimerize. By contrast, a positive cooperative binding of U69593 (a κ agonist) and [d-Pen2,d-Pen5]-enkephalin (a selective δ agonist) on the opioid κ-δ heterodimer has been reported (Jordan and Devi, 1999).
It should also be pointed out that an nonobvious consequence of ligand binding on a dimeric receptor can be the loss of a competitive binding characteristics, and this might explain discrepancies in the affinities of ligands depending on the ligand used as tracer (Durroux, 2005). We observed such discrepancies in the measurement of the affinities of ligands on wild type V1a receptor; this is worsened on the mutated K128A and Q131A V1a receptors. How such mutations affect the competitive binding properties of receptors remains unclear. Because no computational dimeric receptor structure model has been proposed, it is then difficult to propose new hypotheses to describe cross-talk between protomers within a dimer and the role of particular residues. Likewise, a discrepancy between Kd and Ki has been reported for the binding of raclopride on the D2 receptor when experiments were done in the absence of sodium ions (Armstrong and Strange, 2001). Moreover, recently, Griffante and collaborators reported that the discrepancies in the affinity of SSR149415 for the OT receptor observed in two studies (Serradeil-Le Gal et al., 2002; Griffante et al., 2005) could be attributed to the radioligand chosen in the OT receptor binding assays. This might be of great importance for drug screening. It strongly suggests that the determination of the value of the ligand dissociation constant from IC50 (the concentration of competitor for which the binding of the tracer is 50% of its binding in the absence of competitor) is inaccurate when ligand bindings are not strictly competitive.
In conclusion, we showed that ligand binding exhibits cooperative process and that the nature of the cooperativities observed is ligand-dependent. We provided evidence that definitely demonstrates cooperative binding of vasopressin, oxytocin, and their analogs on the vasopressin and oxytocin receptors. These cooperativities can be only explained if receptors are considered multivalent receptor oligomers and are in very good accordance with predictions from the mathematical models that consider binding of ligands on dimer. The binding properties of GPCR can provide strong evidence of receptor dimerization, and their analysis can be a valuable tool to study dimers in native tissues.
Oligomerization of GPCR has been shown to play a role in receptor ontogeny and targeting, in pharmacological properties, in receptor internalization, and, more precisely, in the fate of the internalized receptor (for review, see Milligan, 2004; Terrillon and Bouvier, 2004). Herein, we showed that the binding of a first ligand can modulate the binding of a second one. Whether this step can constitute another level of regulation in signal transduction and then in physiological function remains to be established.
Acknowledgments
Thanks are due to Dr Gilles Guillon for helpful discussion.
Footnotes
-
This work was supported by the research grants from the Centre National de la Recherche Scientifique, from the Institut National de la Santé etdela Recherche Médicale, by grants from ACI Molécules Cibles et Thérapeutiques no. 240 and 355, by National Institutes of Health grant GM25280, and by the European Strep Program LSHB-CT-2003-503337.
-
ABBREVIATIONS: GPCR, G protein-coupled receptor; CHO, Chinese hamster ovary; AVP, arginine vasopressin; OT, oxytocin; HO-LVA, 4-OH-phenylacetyl-d-Tyr(Me)2,Phe3,Gln4,Asn5,Arg6,Pro7,Arg8-NH29; OTA, d(CH2)5[Tyr(Me)2,Thr4,Orn8,Tyr-NH29]vasotocin; GTPγS, guanosine 5′-O-(3-thio)triphosphate; HA, hemagglutinin; 6His, hexahistidine; FRET, fluorescence resonance energy transfer; U69593, (+)-(5α,7α,8β)-N-methyl-N-[7-(1-pyrrolidinyl)-1-oxaspiro[4.5]dec-8-yl]benzeneacetamide.
- Received April 13, 2006.
- Accepted August 22, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}