


Article Figures & Data
Figures
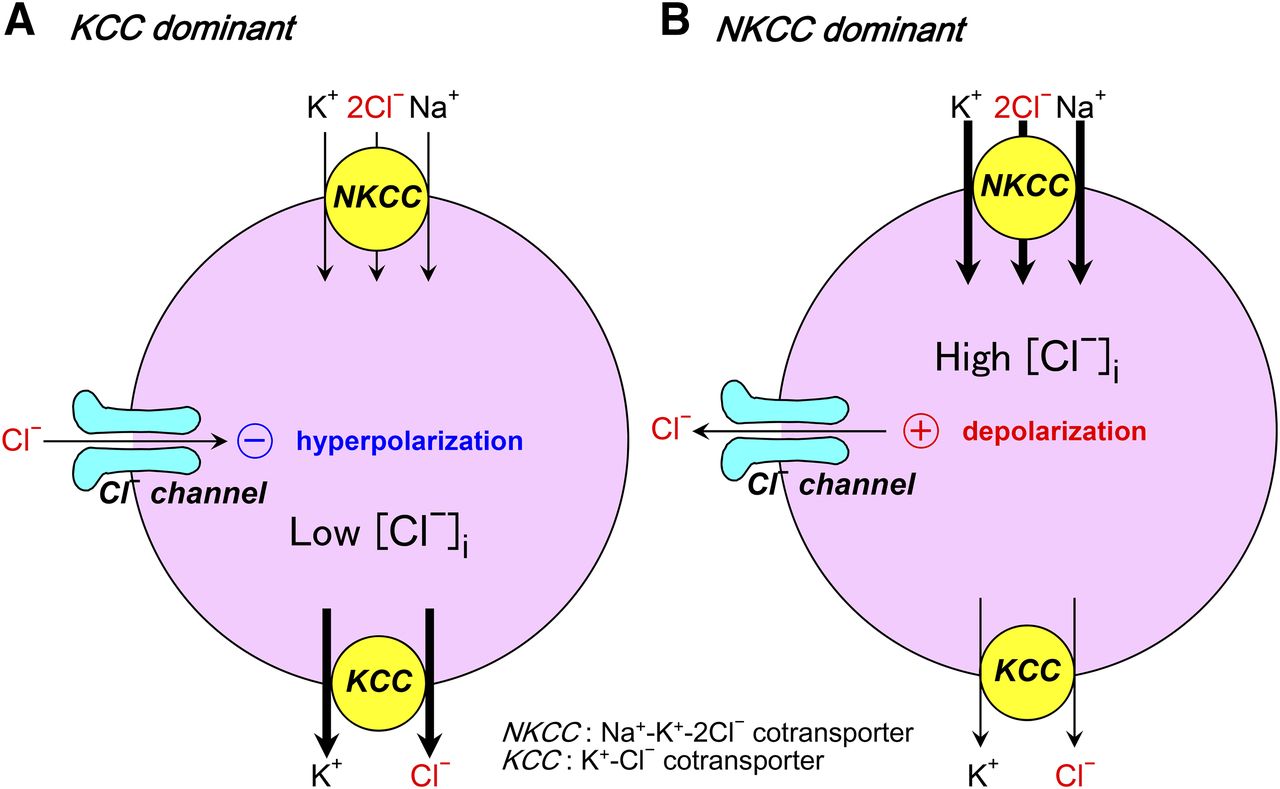
- Fig. 1.
Primary actions of opening of anion channels in either a KCC-dominant case (A) or an NKCC-dominant case (B) (see the text for details).
- Fig. 2.
VSOR and Maxi-Cl currents in human monocytic U937 cells. (A) Macroscopic whole-cell VSOR currents recorded from U937 cells after osmotic swelling (left) and the corresponding current-to-voltage relationship (right). Experimental conditions, other than the cell line used, are the same as reported previously (Sabirov et al., 2001; Kurbannazarova et al., 2011). (B) Single VSOR currents recorded from cell-attached patches on preswollen cells. Holding membrane potentials are indicated on the left of each trace. The right panel shows the unitary current-voltage relationship. The calculated slope conductance is 84.5 ± 5.8 and 14.7 ± 2.4 pS for outward and inward currents, respectively. Experimental conditions, other than the cell line used, are the same as reported previously (Ternovsky et al., 2004; Kurbannazarova et al., 2011) with 100 mM TEACl-pipette and 100 mM KCl-bath solutions. (C) Activation of Maxi-Cl currents in U937 cells. (Left) Voltage- and time-dependent inactivation of the steady-state membrane current as observed in excised macropatches in response to voltage pulses (500 milliseconds) from a holding potential at 0 to ±50 mV in 10-mV increments. The pipette and bath solutions were normal Ringer’s solution. (Right) Single-channel I-V relationships for maxi-anion channel events recorded. Each symbol represents the mean ± S.E. (vertical bar). The solid line is a linear fit with a slope conductance of 445.3 ± 9.6 pS. Experimental conditions, other than the cell line used, are the same as reported previously (Islam et al., 2012).
- Fig. 3.
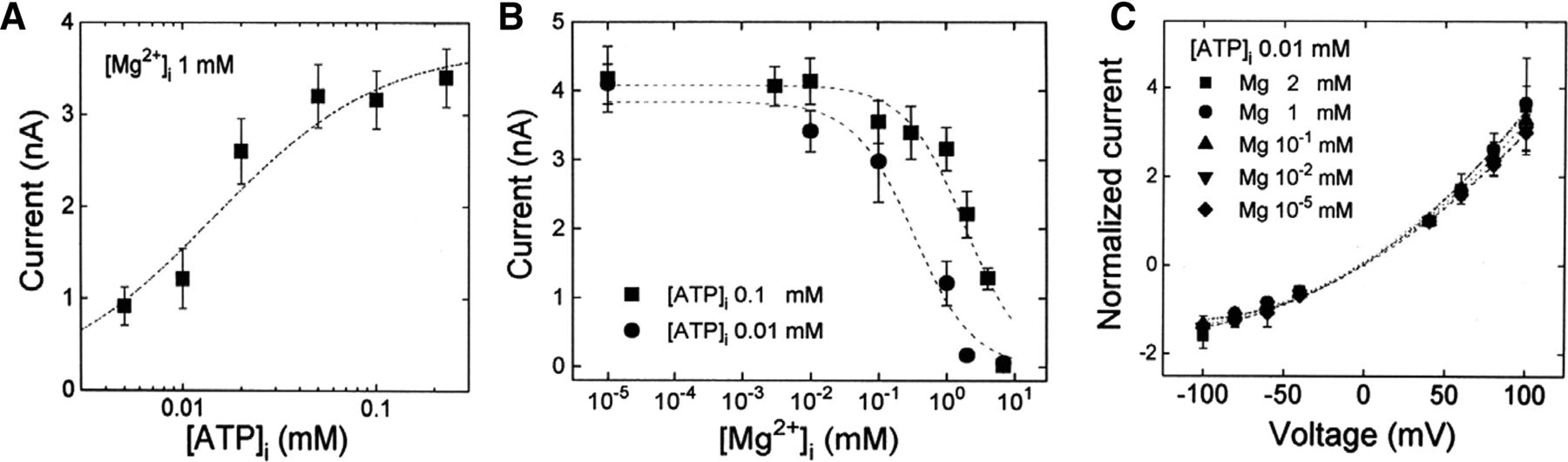
Cl− current amplitudes plotted against [ATP]i, [Mg2+]i, and voltages. Each symbol represents the mean current of four different Intestine 407 cells. Vertical bars represent the S.E. (A) [ATP]i dependence of the peak Cl− currents measured at +40 mV at a fixed [Mg2+]i (1 mM). Data points were fitted by the following equation: I = Imax([ATP]i/EC50)/{1 + ([ATP]i/EC50)}. (B) [Mg2+]i dependence of the peak Cl− currents measured at +40 mV at a fixed [ATP]i (0.1 or 0.01 mM). Data points were fitted by the following equation: I = Imax/{1 + ([Mg2+]i/IC50)}. (C) Current-voltage relationships. Normalized peak Cl− currents (IV/I+40-mV) were measured at different [Mg2+]i and fixed [ATP]i. Mg2+ did not change the reversal potential, indicating that the Mg2+ effect is independent of voltage and that Mg2+ cannot permeate the channel. Experimental conditions are the same as reported previously (Oiki et al., 1994).
- Fig. 4.
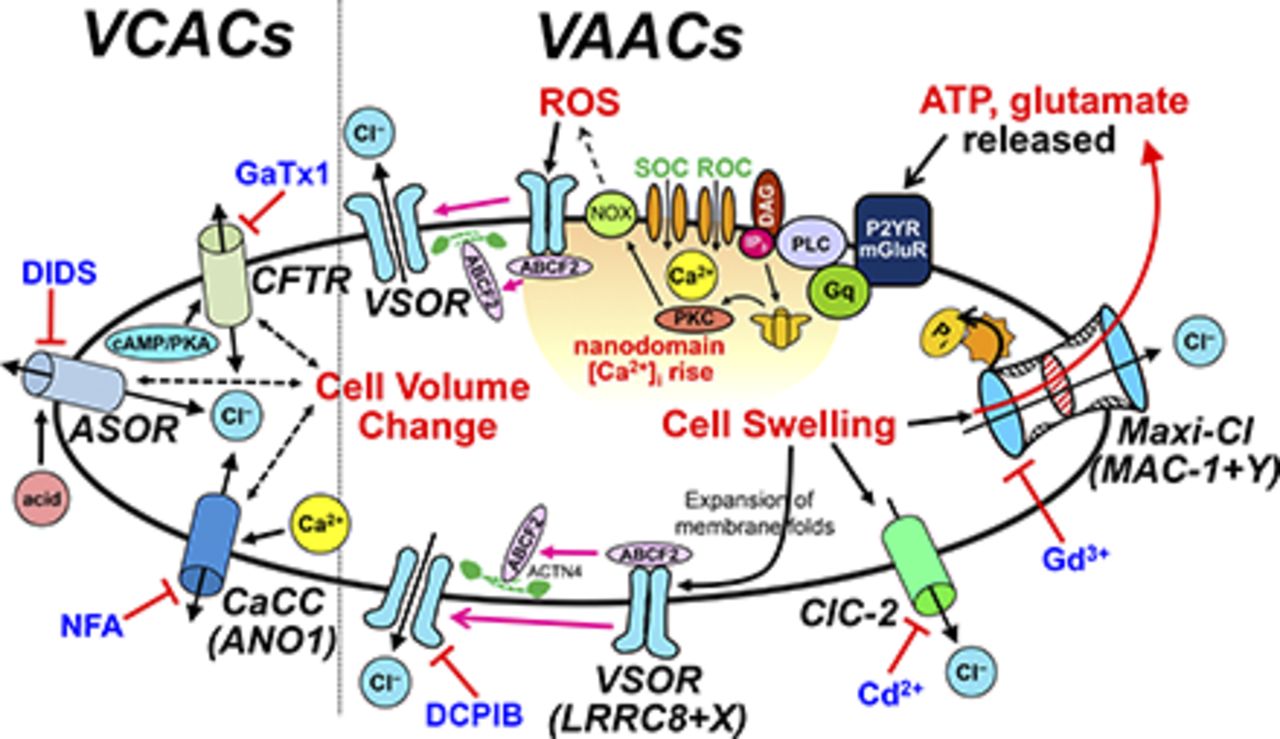
Dual mechanisms for VSOR activation. The bottom half represents the membrane unfolding-induced (type A) mechanism, which takes place upon cell swelling, and the top half represents the oxidation-induced (type B) mechanism, which takes place upon receptor stimulation (see the text for details and other abbreviations). BKR, bradykinin receptor; DAG, diacylglycerol; Gq, heterotrimeric G protein that activates phospholipase C; IP3, inositol trisphosphate; mGluR, metabotropic glutamate receptor; P2YR, P2Y purinergic receptor; PKC, protein kinase C; PLC, phospholipase C; ROC, receptor-operated channel; SOC, store-operated channel.
- Fig. 5.
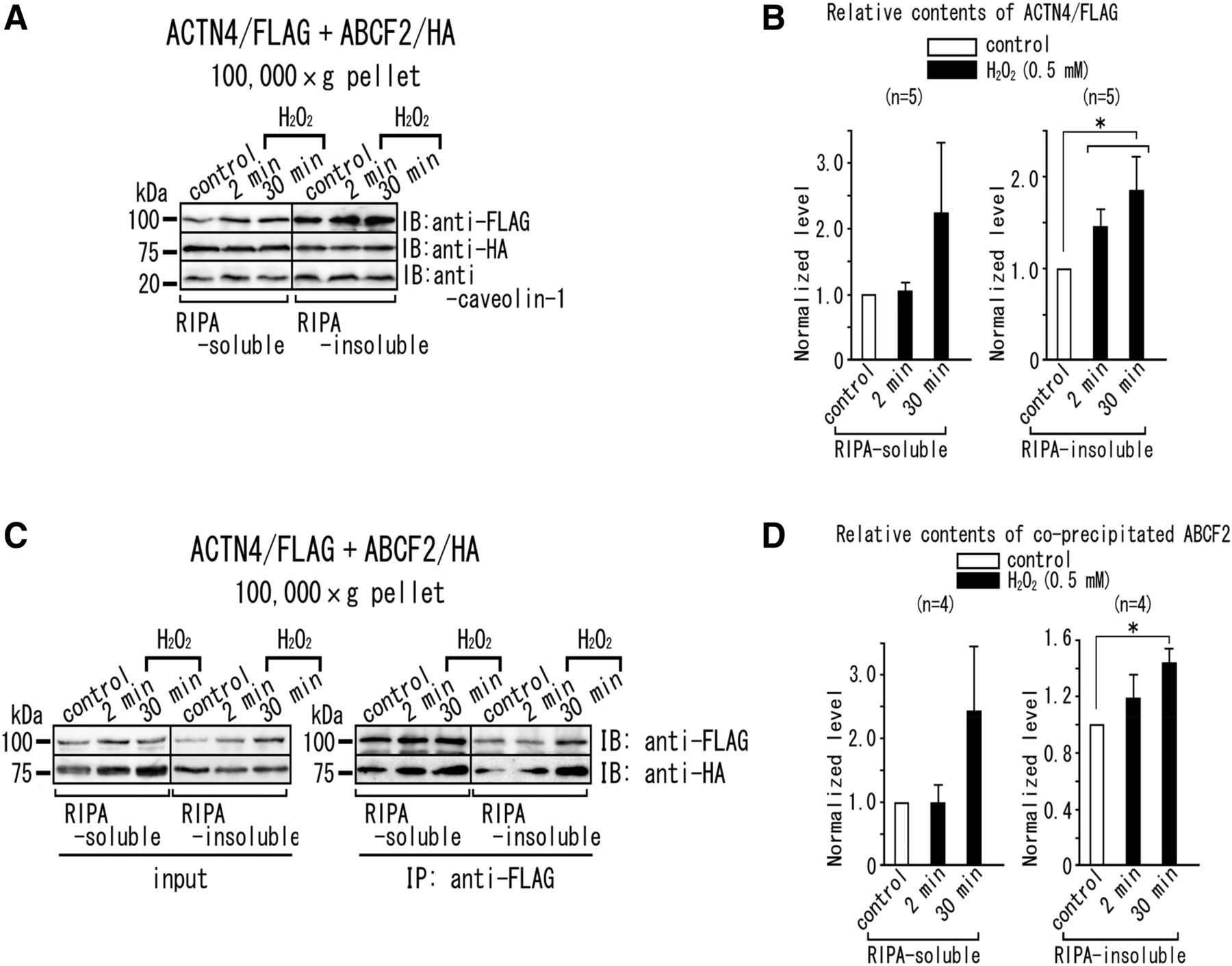
Effects of H2O2 stimulation on the subcellular localization of ACTN4 and molecular interaction between ACTN4 and ABCF2. (A and B) Accumulation of ACTN4, but not ABCF2, in the RIPA-insoluble fraction with 0.5 mM H2O2 stimulation. Five micrograms of RIPA-soluble and RIPA-insoluble fractions of the 100,000g pellet prepared from the cells transiently overexpressing ACTN4/FLAG and ABCF2/HA were subjected to SDS-PAGE and immunoblotting with anti-FLAG M2 mAb and anti-HA and anti–caveolin-1 antibodies. (A) Immunoblots. ACTN4/FLAG, ABCF2/HA, and caveolin-1 were detected. Molecular weight markers (in kilodaltons) are indicated on the left. (B) Densitometric analysis of the immunoblot data shown in (A). The ratio of the ACTN4/FLAG contents (normalized with caveolin-1 content) in the presence of 0.5 mM H2O2 to that in the absence of 0.5 mM H2O2 is plotted. *P < 0.05 (significant difference between the data from cells with or without H2O2 stimulation). (C and D) H2O2 stimulation enhances the interaction between ACTN4 and ABCF2 in the RIPA-insoluble fraction. (C) RIPA-soluble and RIPA-insoluble proteins from the 100,000g pellets were immunoprecipitated using anti-FLAG M2 mAb and were immunoblotted with anti-FLAG mAb and anti-HA antibody (right). Transfected ACTN4/FLAG and ABCF2/HA were shown in the input (left). (D) Densitometric analysis of the immunoblot data shown in (C). The ratio of the coprecipitated ABCF2/HA content (normalized with the precipitated ACTN4/FLAG content) with H2O2 stimulation (black columns) to that of control (white columns) is plotted. *P < 0.05 (significant difference between the data from cells with or without H2O2 stimulation). Experimental conditions are the same as reported previously (Ando-Akatsuka et al., 2012). HA, hemagglutinin; IB, immunoblotted; IP, immunoprecipitated; mAb, monoclonal antibody; RIPA, radioimmunoprecipitation assay buffer.
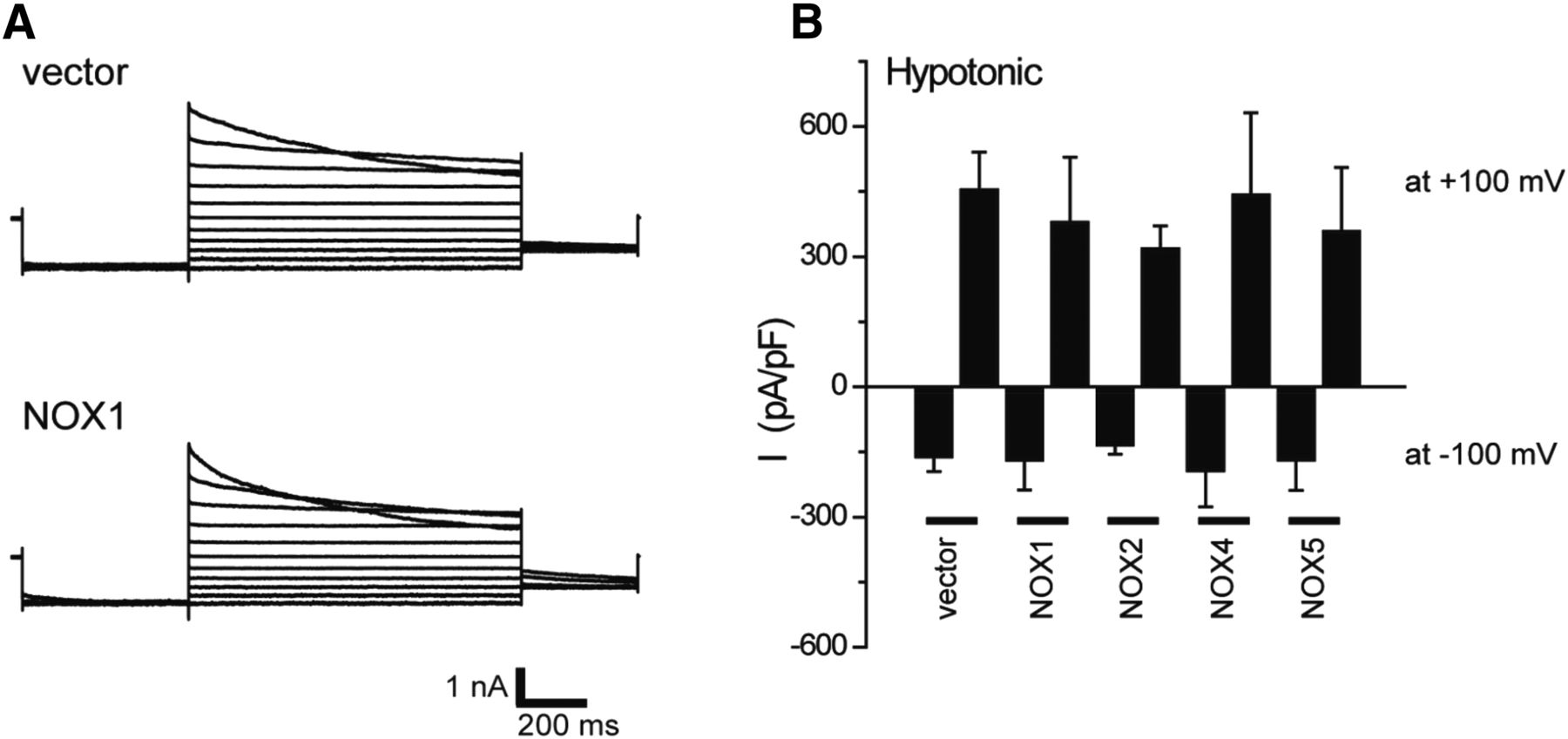
- Fig. 6.
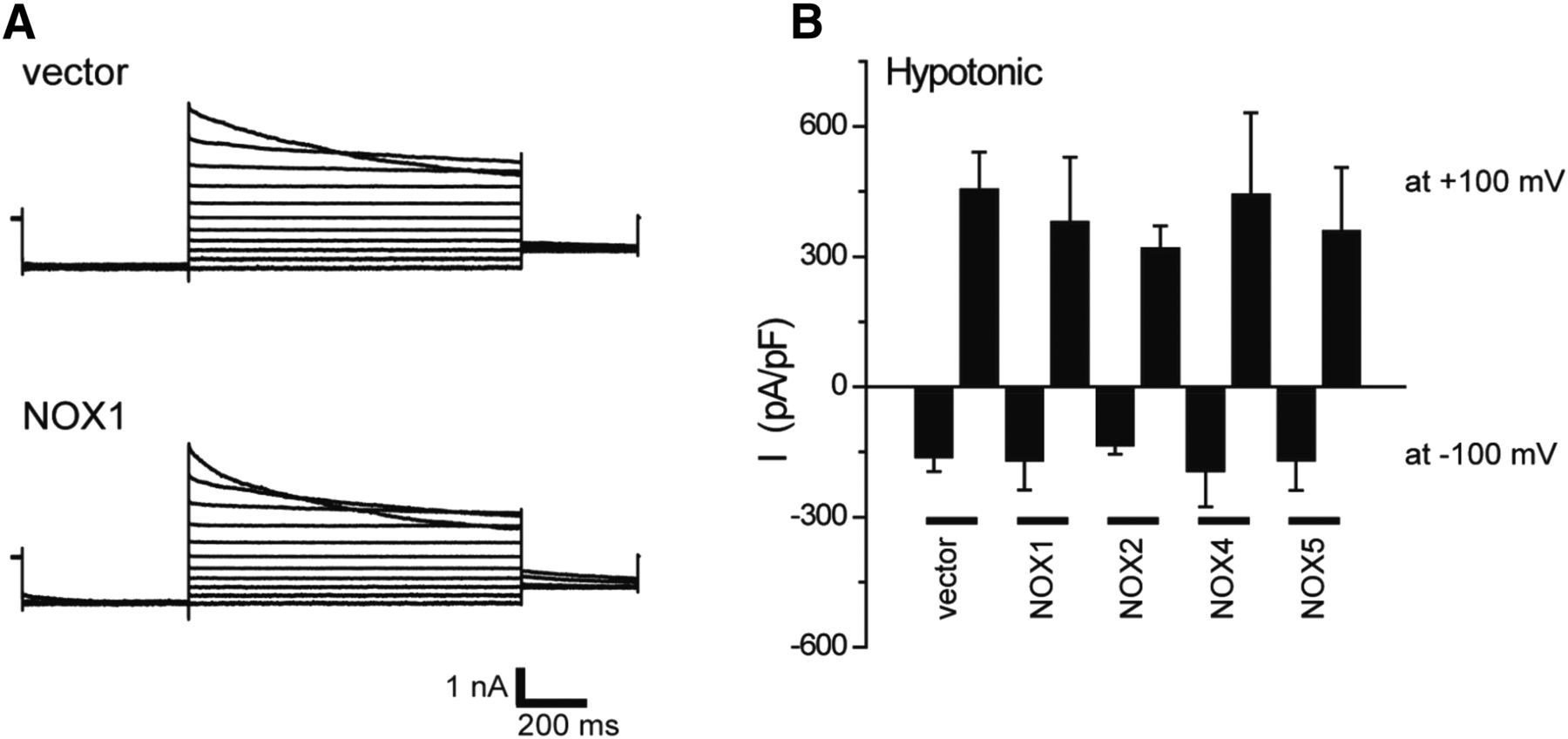
Effects of NOX expression on VSOR currents in HEK293T cells. (A) Representative VSOR whole-cell currents elicited by step pulses from −100 to +100 mV in 20-mV increments during application of hypotonic (83% osmolality) solution in vector-transfected cells (upper panel) and NOX1-transfected cells (lower panel). (B) Mean VSOR currents recorded at +100 and −100 mV in vector-, NOX1-, NOX2-, NOX4-, and NOX5-transfected cells (n = 5–14). Experimental conditions are the same as reported previously (Okada et al., 2009a; Ando-Akatsuka et al., 2002).
- Fig. 7.
Strategy adopted in our study for molecular identification of the core component of Maxi-Cl. By performing bleb membrane proteomics followed by gene silencing and disruption, mutagenesis, pharmacology, heterologous expression, and recombinant protein reconstitution, SLCO2A1 protein was identified as a core component of the ATP-conductive Maxi-Cl channel (see the text and Sabirov et al., 2017 for details). LC, liquid chromatography; MS/MS, tandem mass spectrometry.
- Fig. 8.
Dual modes of SLCO2A1 functions as a transporter, PGT (left), and an ATP-permeable maxi-anion channel, Maxi-Cl (right).
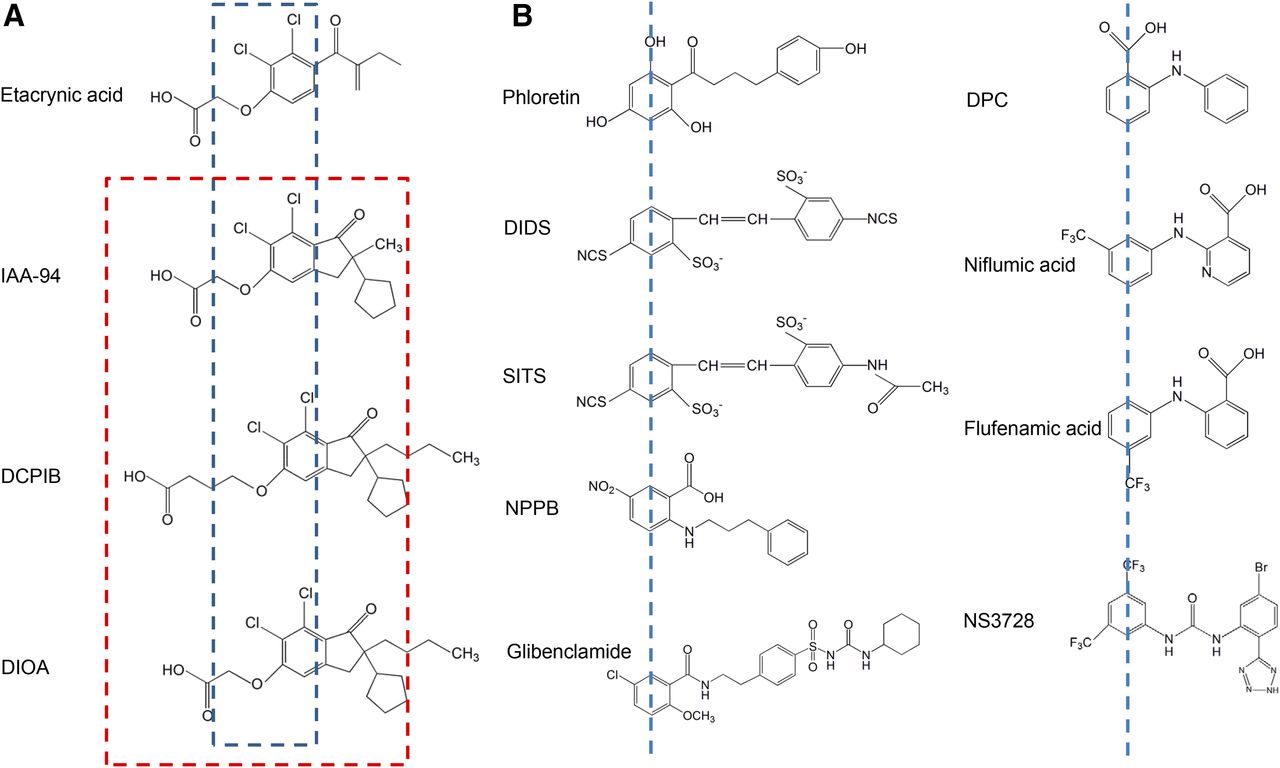
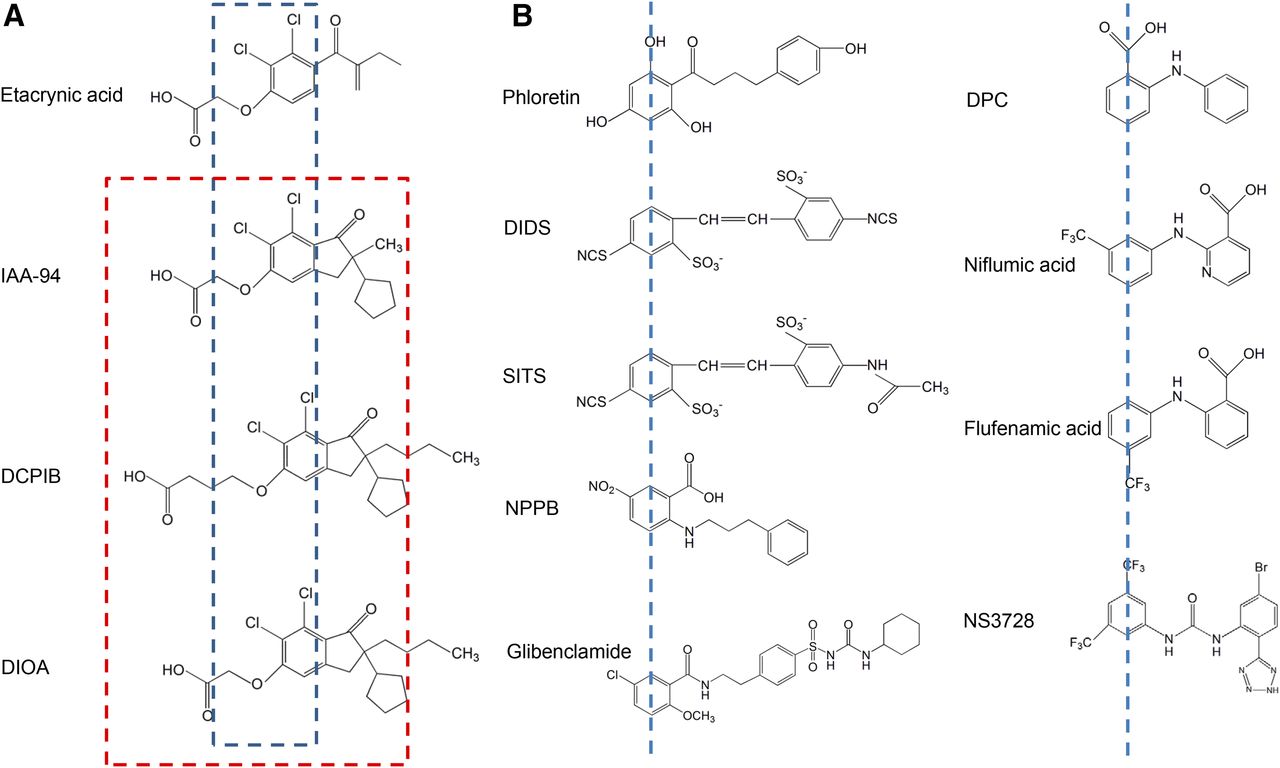
- Fig. 9.
Chemical structures of some VSOR blockers. (A) Etacrynic acid derivatives. The blue dashed line depicts the 2,3-dichlorophenoxy fragment, which is identical for all of these molecules. The red dashed line delineates a fragment (short-chain oxy-carbonic acid combined with chlorinated cyclopentyl-oxo-indanyl) common for VSOR blockers (see the text for details) but not for etacrynic acid. (B) VSOR blockers that have two aromatic rings connected with a chain of one to four atoms (carbon, nitrogen, or mixed). The blue dashed lines mark the first aromatic ring (see the text for details). IAA-94, indanyloxyacetic acid 94.
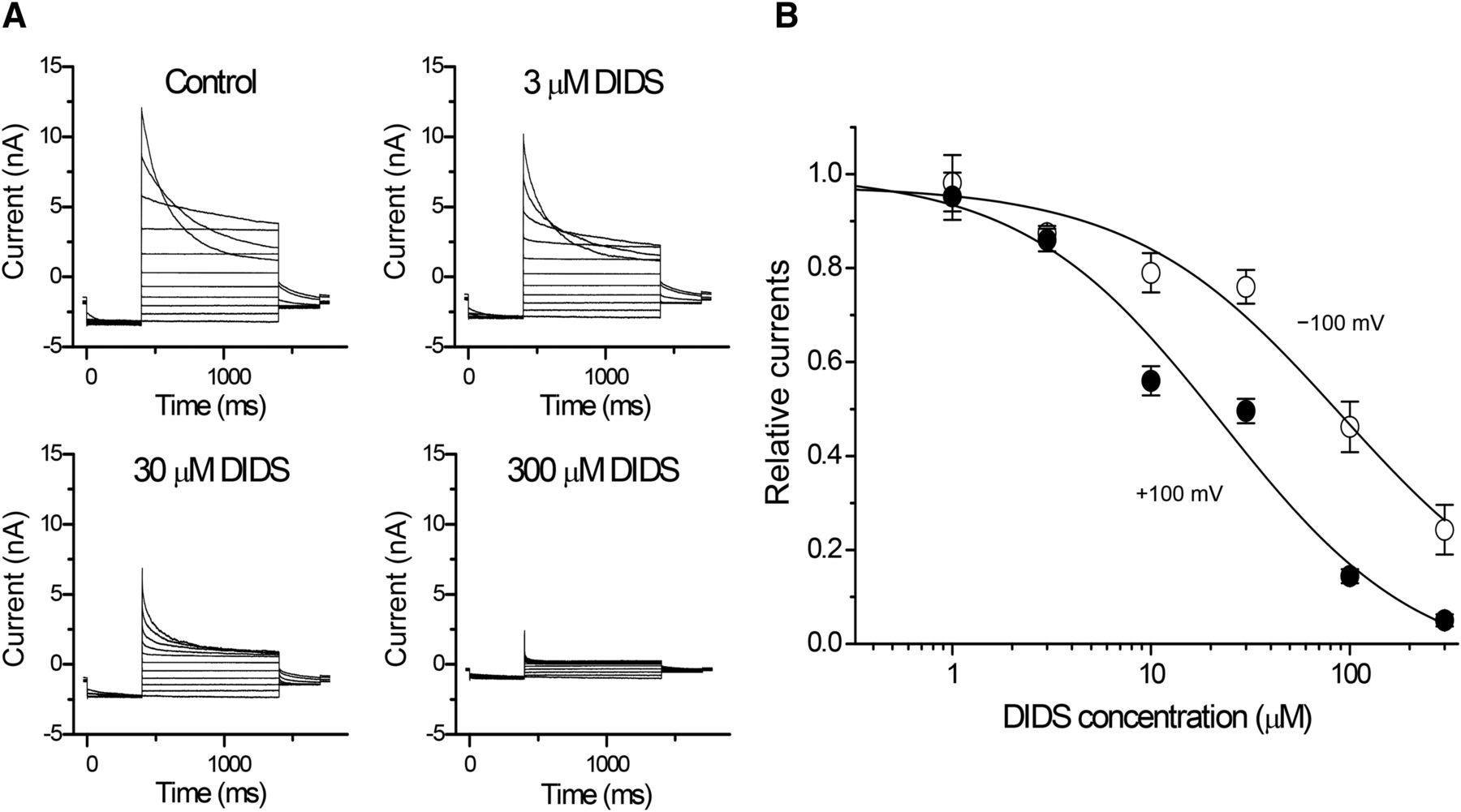
- Fig. 10.
Voltage-dependent inhibition by DIDS of volume-sensitive outwardly rectifying Cl− currents in human cervical carcinoma HeLa cells. (A) Representative current traces before and after application of DIDS. Step pulses were applied from −100 to +100 mV in 20-mV increments, with a prepulse of −100 mV and a postpulse of −60 mV. (B) Concentration dependence of DIDS on the currents recorded at +100 mV (filled circles) and −100 mV (open circles) (n = 3–6). The IC50 values at +100 and −100 mV were 22.7 and 88.5 µM, respectively. Experimental conditions are the same as reported previously (Shimizu et al., 2004).
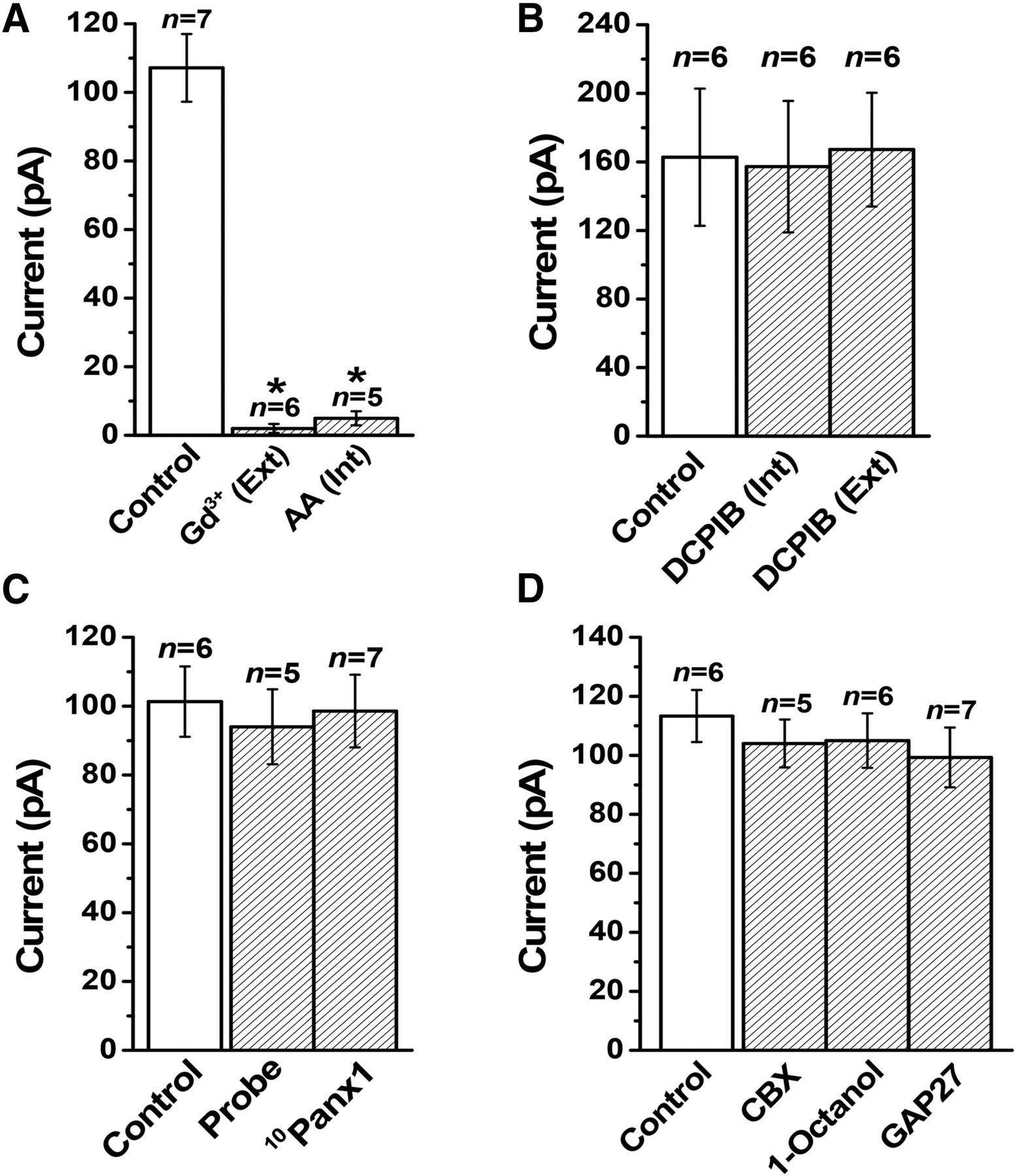
- Fig. 11.
Pharmacological profile of Maxi-Cl currents in patches excised from C127 cells. All of the columns represent the mean currents recorded at +25 mV. (A) Inhibitory effects of extracellular application of Gd3+ (50 μM) and intracellular application of arachidonic acid (20 μM). (B) Insensitivity to a VSOR blocker, DCPIB (10 μM), added from the intracellular or extracellular side. (C) Insensitivity to pannexin hemichannel antagonists, probenecid (1 mM) and 10Panx1 (8 μg/ml), added to the extracellular (pipette) solution. (D) Insensitivity to gap junction antagonists, CBX (100 μM), 1-octanol (2 mM), and GAP27 (4 μg/ml), added to the extracellular (pipette) solution. Experimental conditions, other than the cell line used, are the same as reported previously (Islam et al., 2012; Sabirov et al., 2017). AA, arachidonic acid; Probe, probenecid.
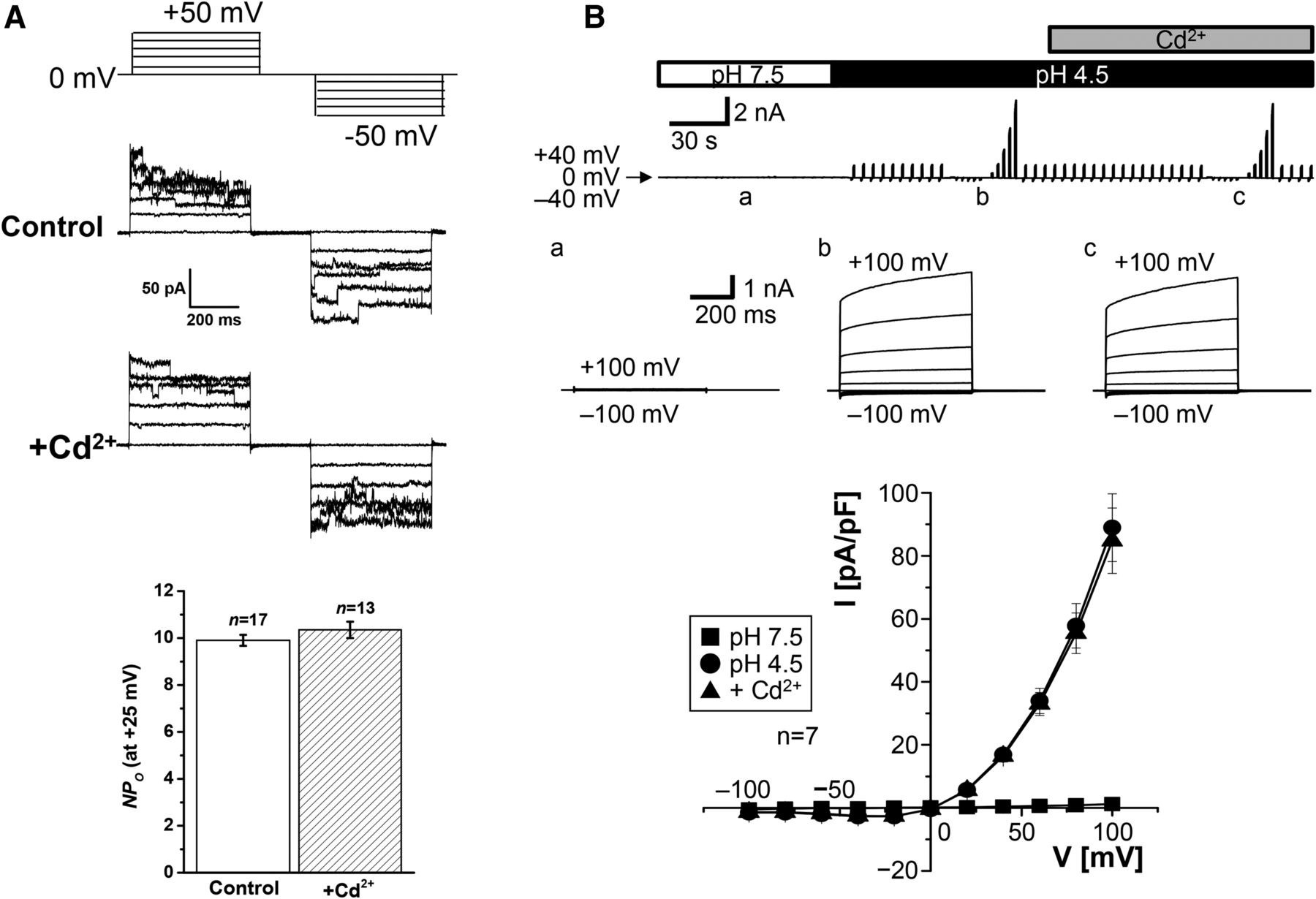
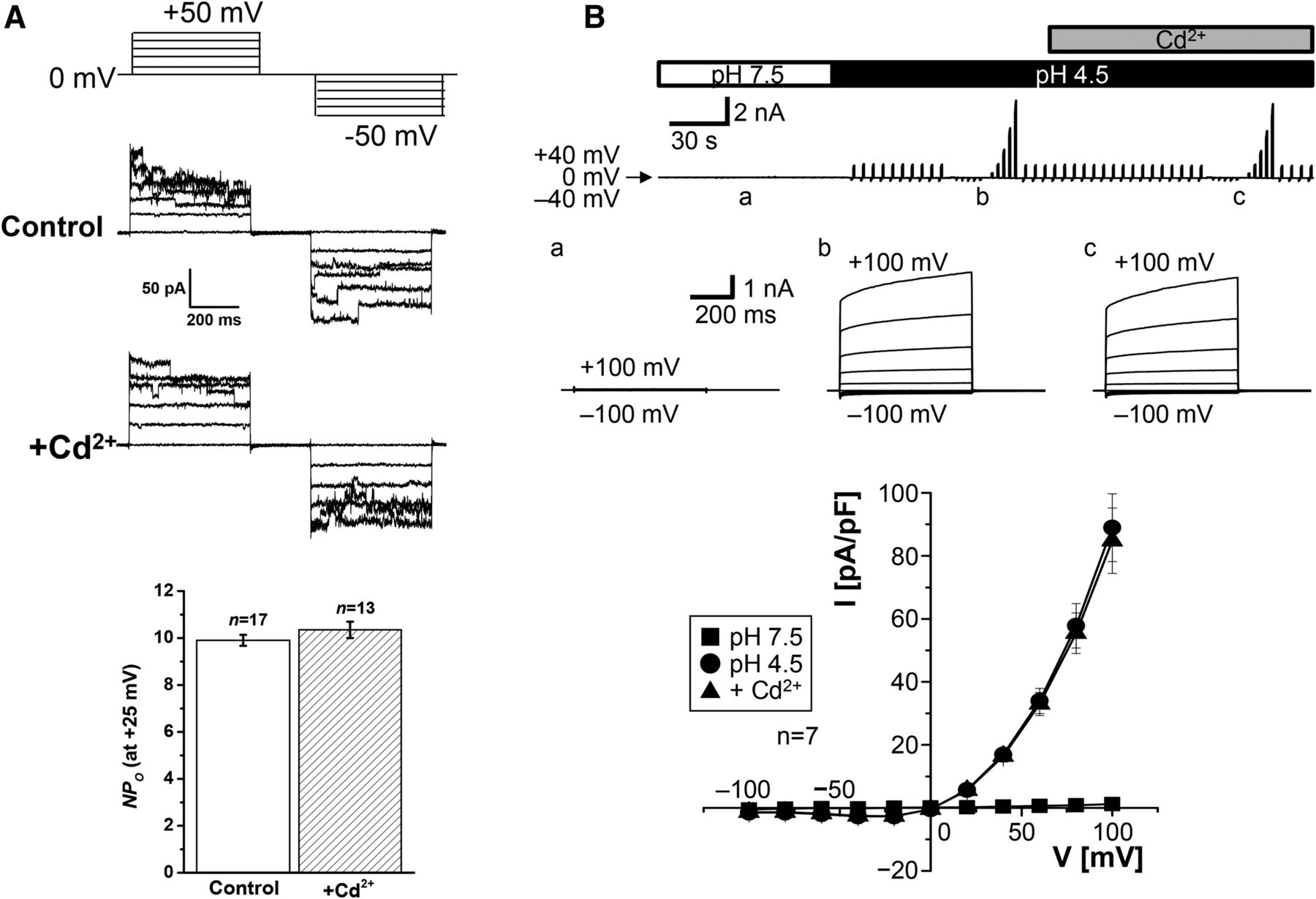
- Fig. 12.
Cd2+ insensitivity of Maxi-Cl (A) and ASOR (B) currents. (A) Representative Maxi-Cl currents recorded in inside-out patch membranes excised from C127 cells in the absence (control) and presence (+Cd2+) of 1 mM Cd2+ added to the extracellular (pipette) solution during application of step pulses from 0 to ±50 mV in 10-mV increments (protocol shown at the top). The bottom panel shows mean (± S.E.M.) NPo values measured at +25 mV in the absence and presence of Cd2+. There was no significant difference (at P > 0.05) between the two values. Experimental conditions are the same as reported previously (Islam et al., 2012; Sabirov et al., 2017). (B) Whole-cell ASOR currents recorded in HeLa cells in the absence and presence of 1 mM Cd2+ added to the bath solution. The upper panel shows whole-cell currents before (pH 7.5) and during (pH 4.5) exposure to acidic bath solution in the absence and presence of Cd2+. The currents were elicited by application of alternating pulses from 0 to ±40 mV or of step pulses (at a, b, and c) from −100 to +100 mV in 20-mV increments. The middle panel shows expanded traces of current responses (at a, b, and c) to the step pulses. The bottom panels show I-V relationships for the whole-cell current densities at pH 7.5 (squares) and pH 4.5 in the absence (circles) and presence (triangles) of Cd2+. There was no significant difference (at P > 0.05) between the circles and triangles at given voltages. Experimental conditions are the same as reported previously (Sato-Numata et al., 2016). NPo, number of open channels.









Tables
- TABLE 1
Mammalian anion channels: Classification and relation to cell volume and organic anion transport
Species Channel Name Abbreviation Molecule Ligand gated Glycine receptor GlyR GlyR GABAA receptor GABAAR GABAA GABAC receptor GABACR GABAC Voltage gated ClC-type chloride channel ClC-1 CLCN1 ClC-2 (one of VAAC and VRAC) CLCN2 ClC-K1 CLCNK1 ClC-K2 CLCNK2 cAMP activated Cystic fibrosis transmembrane conductance regulator CFTR (one of VCAC and VSOAC) CFTR Ca2+ activated CaCC (one of VCAC) TMEM16A/ANO1 TMEM16B/ANO2 TMEM16F/ANO6 Volume activated Volume-sensitive outwardly rectifying anion channel VSOR (one of VAAC, VRAC,a and VSOAC) LRRC8A+8C/8D/8E(+?c) Maxi-anion channel Maxi-Cl (one of VAAC, VRAC,a VSOAC, and MAC-1b) SLCO2A1(+?c) Acid activated Acid-sensitive outwardly rectifying anion channel ASOR (one of VCAC) ? ↵a The VRAC abbreviation used in this work (for volume-regulated anion channel) is sometimes also referred to as the volume-regulatory anion channel.
↵b Maxi-Cl is the major type of maxi-anion channel (MAC-1) (see the text for details).
↵c The question mark represents some missing molecular component which may exist to reproduce a full set of phenotypes (see the text for details).
Chemical ClC-2 VSOR Maxi-Cl CFTR CaCC ASOR Cd2+ + − − − NS − Gd3+ − −/+a +∼+++ − − − DCPIB − ++ − − − − DIDS (out) − ±∼++ VD + − ±∼+ VD ++∼+++ NPPB ± VD + + +++ VD + − NFA − ± − ± VD +∼++ + Glibenclamide NS ± (VDb) − +∼++ VD + ± Phloretin NS + − ± VD − + CFTRinh-172 +++ VD −/+∼++a NS +++ − NS CaCCinh-A01 NS − NS NS ++ NS T16Ainh-A01 NS −/++a NS NS ++∼+++ NS GaTx1 − NS NS +++ − NS GaTx2 +++ NS NS − − NS Methadone +++ NS NS − NS NS Insensitivity and sensitivity are denoted by minus and plus signs, respectively, as follows: −, insensitive; ±, sensitive at sub-millimolar (100 ≤ IC50 < 1000 μM) concentrations; +, sensitive at deca-micromolar (10 ≤ IC50 < 100 μM) concentrations; ++, sensitive at micromolar (1 ≤ IC50 < 10 μM) concentrations; and +++, sensitive at sub-micromolar (IC50 < 1 μM) concentrations. NS, not studied; VD, voltage-dependent block.
↵a Depending on cell type (see the text for details).
↵b Voltage-dependent block only by the charged form.
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Visual Overview
- Abstract
- I. Introduction
- II. Volume-Activated Anion Channels
- III. Volume-Correlated Anion Channels
- IV. Pharmacological Properties of and Distinctions between Volume-Activated Anion Channels and Volume-Correlated Anion Channels
- V. Conclusions and Future Directions
- Acknowledgments
- Authorship Contributions
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters