


Abstract
Mounting evidence suggests safety and efficacy of psychedelic compounds as potential novel therapeutics in psychiatry. Ketamine has been approved by the Food and Drug Administration in a new class of antidepressants, and 3,4-methylenedioxymethamphetamine (MDMA) is undergoing phase III clinical trials for post-traumatic stress disorder. Psilocybin and lysergic acid diethylamide (LSD) are being investigated in several phase II and phase I clinical trials. Hence, the concept of psychedelics as therapeutics may be incorporated into modern society. Here, we discuss the main known neurobiological therapeutic mechanisms of psychedelics, which are thought to be mediated by the effects of these compounds on the serotonergic (via 5-HT2A and 5-HT1A receptors) and glutamatergic [via N-methyl-d-aspartate (NMDA) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors] systems. We focus on 1) neuroplasticity mediated by the modulation of mammalian target of rapamycin–, brain-derived neurotrophic factor–, and early growth response–related pathways; 2) immunomodulation via effects on the hypothalamic-pituitary-adrenal axis, nuclear factor ĸB, and cytokines such as tumor necrosis factor-α and interleukin 1, 6, and 10 production and release; and 3) modulation of serotonergic, dopaminergic, glutamatergic, GABAergic, and norepinephrinergic receptors, transporters, and turnover systems. We discuss arising concerns and ways to assess potential neurobiological changes, dependence, and immunosuppression. Although larger cohorts are required to corroborate preliminary findings, the results obtained so far are promising and represent a critical opportunity for improvement of pharmacotherapies in psychiatry, an area that has seen limited therapeutic advancement in the last 20 years. Studies are underway that are trying to decouple the psychedelic effects from the therapeutic effects of these compounds.
Significance Statement Psychedelic compounds are emerging as potential novel therapeutics in psychiatry. However, understanding of molecular mechanisms mediating improvement remains limited. This paper reviews the available evidence concerning the effects of psychedelic compounds on pathways that modulate neuroplasticity, immunity, and neurotransmitter systems. This work aims to be a reference for psychiatrists who may soon be faced with the possibility of prescribing psychedelic compounds as medications, helping them assess which compound(s) and regimen could be most useful for decreasing specific psychiatric symptoms.
I. Introduction
A. Review Outline
In the last 30 years, the revamped interest in the application of psychedelic compounds in psychiatry has generated a huge body of work on the pharmacological and therapeutic action of these molecules. This has recently led to approval by the FDA of ketamine as a first in a new class of antidepressants (Kim et al., 2019). Other compounds are being tested in multicenter phase III and II clinical trials, such as 3,4-methylenedioxymethamphetamine (MDMA)-augmented psychotherapy for treatment-resistant post-traumatic stress disorder (PTSD) and psilocybin for treatment-resistant major depressive disorder (MDD) (ClinicalTrials.gov). Hence, the clinical implementation of these compounds has begun and may soon expand, indicating that the use of psychedelic compounds for therapeutic purposes may be incorporated into modern society (Rucker et al., 2018; Ona et al., 2019).
Psychiatrists are faced with a new class of therapeutic tools that may be licensed as medicines and can almost immediately improve psychiatric symptoms, but that can elicit profound changes in consciousness and perception. The experiences elicited have been described by participants in clinical trials as among some of the most transformative in their lives. Although this aspect will not be discussed in detail in this review, it raises the critical concept that adequate preparation should be provided to psychiatrists who will prescribe and administer these compounds in clinical settings. Importantly, the context in which the therapy takes place can influence the final outcomes, adding a layer of complexity (Carhart-Harris et al., 2018b). Specialized therapist training is already being provided by research centers currently researching psychedelics and may need to be scaled up should this type of treatment become more widely used (Phelps, 2017; Nutt and Carhart-Harris, 2020).
To elicit significant improvements in the psychiatric symptomatology, in most cases, this type of therapy requires administration of the compound only once or twice over a few weeks, which are preceded and followed by preparation and integration sessions with a trained therapist. A four-step treatment model is becoming standardized in randomized controlled trials (RCTs) and could be applied to the clinic. This model includes 1) the assessment of the patient’s mental and physical suitability for this type of therapy; 2) the preparation provided by a trained therapist to the multifaceted possibilities of the “psychological journey,” as well as indications on how to navigate potentially challenging experiences and get the most from the therapy; 3) the experience session itself, which involves the administration of the compound in comfortable settings, such as a lounge-like environment, the possibility of listening to music and wearing eyeshades, and the continuous presence of a trained therapist, which is available should the patient feel the need to interact verbally or physically; and 4) the integration session to discuss with a trained therapist how the experience might relate to the patient’s illness and to help the patient integrate the experience with his or her life. Ideally, further psychotherapeutic integration sessions should be available to process issues or insights that might arise and to provide guidance on how to cultivate lifestyle and cognitive adjustments (Nutt and Carhart-Harris, 2020). The integration part seems especially relevant given that antidepressant responses are enhanced by an enriched environment but can be counteracted by a stressful one (Alboni et al., 2016). Therefore, considering that patients often report significant changes in their value sets, world views, and meaning (Hartogsohn, 2018), which might clash with their pre-existing lifestyle generating substantial distress, it is essential that patients are adequately followed up to optimize treatment outcome and avoid undesirable side effects or relapsing into unhealthy habits/patterns (Richards, 2016; Sloshower, 2018; Watts and Luoma, 2020).
Most importantly, the clinical implementation of these compounds requires psychiatrists to be familiarized with the pharmacology of these compounds, the neurobiological mechanisms at the root of therapeutic improvement, and potential mechanisms and pharmacological interactions that could mediate the insurgence of more or less severe side effects. Here, we discuss the current understanding of the neurobiology of psychedelic compounds, focusing on compounds which have been, or could soon be, classified as novel psychiatric medications. We discuss the current understanding of the pharmacology underlying the effects of psychedelics on neuroplasticity, immunomodulation, and neurotransmission. When possible, we will draw parallels in terms of current psychiatric pharmacotherapies and how psychedelic compounds weigh up in comparison. The many published results available in the scientific literature show that notable efforts are being poured in this emerging field, and as a result, our understanding of the neurobiology of psychiatric drugs is deepening exponentially. To narrow the scope of this review, we decided to focus on compounds that are already approved as medications or are under scrutiny for their potential application in psychiatry. Other psychedelics may be touched upon if relevant for this review but will not be discussed in depth.
B. Psychiatric Disorders and the Need for Novel Pharmacotherapies
Psychiatric disorders are a major public health concern and a leading cause of economic burden worldwide, affecting about 350 million people (Wittchen et al., 2011; Whiteford et al., 2013; Global Burden of Disease Study 2013 Collaborators, 2015; Vigo et al., 2019). It is estimated that one in two individuals (50% of the population) in high-income countries will meet the diagnosis for at least one psychiatric disorder in his or her lifetime (Wittchen et al., 2011; Kessler et al., 2012). Although this represents a historical peak, in terms of monetary expenditure and years lived with disability, the incidence of these conditions is still dramatically increasing (Vos et al., 2012; Global Burden of Disease Study 2013 Collaborators, 2015). Psychiatric disorders are multifactorial disorders arising from genes × environment interaction (Wong et al., 2008; Koenen et al., 2009; Flint and Kendler, 2014; Kaufman, 2018). Genetic variabilities among populations (such as clusters of single nucleotide polymorphisms) have been identified that confer vulnerability or resilience to psychiatric disorders, spanning DNA regions involved in neuroplasticity, immune system modulation, and neurotransmission (Caspi et al., 2010; Cross-Disorder Group of the Psychiatric Genomics Consortium, 2013; Smoller, 2016; Wray et al., 2018).
Individuals diagnosed with a psychiatric disorder, which may be chronic or relapsing, have increased odds to develop comorbid systemic illnesses, which aggravate the psychiatric symptomatology in a feedback loop (Koenen et al., 2009; Rogers et al., 2016; Inserra et al., 2018; Sporinova et al., 2019). A further emerging biological contributor to the development of psychiatric disorders is epigenetics, the ensemble of functional chromatin modifications shaped by one’s life (Nestler et al., 2016; Kular and Kular, 2018). Contrasting a long-standing paradigm, epigenetic modifications are transgenerational (can be passed onto the offspring) and may well play a role in the current psychiatric epidemics (Tsankova et al., 2007; Yeshurun and Hannan, 2019). Despite tremendous efforts to uncover pathophysiological determinants, available treatments are only partly effective and are still far from ideal. These treatments seldom lead to clinical remission and present significant side effects, delayed therapeutic onset, and residual symptoms (Penn and Tracy, 2012; Duman et al., 2016). Therefore, identifying novel therapeutic strategies is of paramount importance to the current public health system, especially for psychiatric diseases in which no fully efficacious treatments are available, such as PTSD or autism spectrum disorder (ASD). Ideally, the next generation of psychiatric medications will have a fast onset of therapeutic action and few, acceptable, side effects (Duman et al., 2016; Harmer et al., 2017).
C. Psychedelic Compounds as Novel Therapeutics in Psychiatry: Overview and Comparison with Current Available Treatments
Preliminary evidence suggests that psychedelic compounds may hold potential as therapeutics for psychiatric disorders. The term “psychedelic” derives from the Greek words meaning “mind manifesting,” which refers to the ability of these compounds to bring hidden aspects of the subconscious mind into a conscious framework. Psychedelics induce profound changes in consciousness, perception, emotions, and self-awareness. These effects have been described as paradoxical given that, although acute psychotic-like symptoms are experienced, significant mid- to long-term improvements in psychological well-being are usually reported (Vollenweider and Kometer, 2010; Carhart-Harris et al., 2016b; De Gregorio et al., 2018). Preliminary evidence of safety, efficacy, and tolerability generated by clinical trials is available for 1) ketamine, psilocybin, and ayahuasca for recurrent and treatment-resistant MDD (Murrough et al., 2013a; Carhart-Harris et al., 2016a; Palhano-Fontes et al., 2019; Fava et al., 2020; Davis et al., 2020); 2) MDMA and LSD for treatment-resistant PTSD (Mithoefer et al., 2018, 2019; Schmid et al., 2020); 3) psilocybin for obsessive-compulsive disorder (OCD) (Leonard and Rapoport, 1987; Moreno et al., 2006), 4) alcohol abuse (Bogenschutz et al., 2015), and 5) smoking cessation (Johnson et al., 2014); 6) ayahuasca for suicidality (Zeifman et al., 2020); and 7) psilocybin and lysergic acid diethylamide (LSD) for anxiety, depression, pain, and distress associated with a life-threatening illness (Gasser et al., 2014; Griffiths et al., 2016).
First-line pharmacological treatments in unipolar depression, anxiety, PTSD, and OCD revolve around selective serotonin reuptake inhibitors (SSRIs) [such as fluoxetine, fluvoxamine, citalopram and paroxetine, which inhibit serotonin (5-HT) reuptake at the synapse] and serotonin-norepinephrine reuptake inhibitors (SNRIs) [such as clomipramine, venlafaxine, duloxetine, milnacipran, and levomilnacipran, which inhibit 5-HT and norepinephrine (NE) reuptake at the synapse]. Second-line treatments include the second-generation (atypical) antipsychotics acting on 5-HT2A, 5-HT1A, and D2 receptors (Gobbi et al., 2018). In some cases, tricyclic antidepressants (such as imipramine and amitriptyline, which also inhibit monoamine reuptake) are similarly, or more, effective for the treatment of some patients, especially in hospitalized patients with severe depression (Bauer et al., 2002; Bandelow et al., 2008).
Second-generation (or atypical) antipsychotics (such as clozapine, olanzapine, quetiapine, aripiprazole, risperidone) are used for the long-term treatment of psychoses (Kapur and Remington, 1996; Hasan et al., 2012, 2013) since they have less risk of producing extrapyramidal symptoms as compared with typical antipsychotics. Atypical antipsychotics have a mixed DA/5-HT2A/1A receptor pharmacological profile, including antagonist action at the level of 5-HT2A receptors (olanzapine and quetiapine) or partial agonism at the level of D2 receptor (aripiprazole). Clozapine, which is a potent 5-HT2A/2C receptor antagonist, is suggested as second-line treatment in these cases because of its superior efficacy in treatment-resistant schizophrenia [for revision of the mechanism of action, see Comai et al. (2012)]. Interestingly for this context, phencyclidine (PCP) and other 5-HT2A receptor agonist hallucinogens at high doses have been proposed as a model of animal psychosis that can be reversed by atypical antipsychotics with 5-HT2A and D2 receptor antagonism (Geyer and Ellenbroek, 2003).
Pharmacological overlap exists between the serotonergic action of psychedelic compounds, routinely prescribed psychiatric drugs such as SSRIs (about 80% of all prescribed antidepressants), and atypical (second-generation) antipsychotics, mostly stemming from the interaction with one or more 5-HT receptors or with the 5-HT transporter (SERT) (Celada et al., 2004). Psychedelics are agonists or partial agonists at presynaptic 5-HT1A receptors and postsynaptic 5-HT1A and 5-HT2A receptors; the onset of antidepressant effects with other antidepressants seems to require the hippocampal upregulation of presynaptic 5-HT1A receptor–mediated transmission, suggesting a potential common mechanism of therapeutic improvement (Haddjeri et al., 1998). Both the SSRIs clomipramine and LSD increase 5-HT synaptic availability and inhibit dorsal raphe nucleus (DRN)/5-HT cell firing (Freedman, 1961; Aghajanian et al., 1968; Adell and Artigas, 1991; De Gregorio et al., 2016b). Most antidepressants also downregulate 5-HT2A receptor expression, and although evidence for a downregulation of 5-HT2A receptors in response to psychedelics remains limited, such an effect cannot be excluded (Gray and Roth, 2001).
One study found a decrease in 5-HT binding sites in long-term ayahuasca users, suggesting that similarly to currently approved psychiatric drugs, psychedelic compounds affect the homeostasis of the 5-HT system via downregulating 5-HT2A receptor expression (Callaway et al., 1994). Animal studies corroborate this notion, confirming a downregulation of 5-HT2A receptor expression and functional signaling changes in response to the repeated administration of LSD, which is concomitant to the onset of behavioral tolerance (Gresch et al., 2005; Martin et al., 2014). On the one hand, atypical antipsychotics are potent 5-HT2A/2C receptors antagonists (and weak D2 receptor antagonists), and their antipsychotic potency positively correlates with their profile of 5-HT2A/2C and D2 receptor affinity (Richtand et al., 2007; Lord et al., 2017). This action is thought to counteract the aberrant dopaminergic neurotransmission in SCZ by directly or indirectly increasing 5-HT1A receptor–mediated neurotransmission and modulating the biosynthesis and release of dopamine, ultimately eliciting antipsychotic activity (Schmidt et al., 1995; Kehne et al., 1996; Rollema et al., 1997; Lieberman et al., 1998; Díaz-Mataix et al., 2005; Meltzer and Huang, 2008;; Meltzer and Massey, 2011). On the other hand, most psychedelics are 5-HT2A receptor agonists, and this action is thought to mediate their psychedelic, and hallucinogenic, effects (Kroeze and Roth, 1998; Meltzer, 1999; Celada et al., 2004; Nichols, 2016).
Although the mechanism of action of SSRIs and psychedelics appears somewhat similar—for example, SSRIs and LSD both decrease the 5-HT firing activity through the stimulation of 5-HT1A autoreceptor (De Gregorio, 2016b)—one discrepancy between the dynamics of therapeutic improvement elicited by SSRIs and psychedelics is the timing: psychedelics appear to induce almost immediate improvements in psychiatric symptoms, whereas routinely approved antidepressants require days to weeks (Blier and de Montigny, 1994). This can be explained by the fact that SSRIs elicit noticeable therapeutic improvements after the 5-HT1A autoreceptor is desensitized and the firing of 5-HT is restored, and this requires a significant time frame (days to weeks) to elicit pre- and postsynaptic adjustments that translate to therapeutic improvements, which are thought to be mediated by epigenetic processes (Tsankova et al., 2007; Baudry et al., 2019). Psychedelics, like LSD, also require a desensitization of the 5-HT1A receptor (unpublished data) to elicit anxiolytic effects, but this delay is shorter, and it is therefore possible that the therapeutic improvements are elicited through mechanisms other than 5-HT1A receptor desensitization, including direct action on cortical 5-HT2A and α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors (Marek and Aghajanian, 1996). Although further studies are required to answer this research question, none of the proposed possibilities can be categorically excluded at this stage.
The studies performed so far that investigate the effects of psychedelic compounds for a variety of psychiatric illnesses have yielded encouraging results, suggesting that these compounds might have a place in the treatment of several psychiatric disorders that represent a huge economic burden over the health system and society as a whole. However, one major barrier hampers preclinical and clinical research on these compounds, ultimately hindering the potential application of these substances in psychiatry and medicine in general: their classification as schedule 1 substances in the United States and their homologous classifications in other countries. According to their status, these compounds are drugs with high abuse potential and no medical application. Although such classification is necessary for substances that objectively display highly addictive or highly reward-stimulating profiles, the classification of these compounds as schedule 1 appears not to entirely reflect their pharmacological profiles, including low addictive liability (discussed throughout this review).
Steps that might reconcile this duality include but are not limited to 1) increased preclinical and phase I to III clinical trials demonstrating the clinical efficacy of this class of compounds for specific mental diseases and approval by drug regulatory governmental agencies (i.e., FDA, European Medicines Agency, Health Canada); 2) an evidence-based approach by legislators and funding bodies; 3) destigmatization of these compounds by society, which requires efforts from regulatory government and funding bodies, academic institutions, the health system, and mass media; and 4) priority funding dedicated to answering such questions through evidence-based scientific investigations. Given that no medicine is devoid of risks, such steps could help define more clearly the boundaries between the real risks versus the real benefits brought on by the application of these compounds in psychiatry and in medicine more generally speaking. Ultimately, if the results obtained will be positive, such an evidence-based approach could lead to the use of these compounds in medical practice. However, this requires an evidence-based debate to catalyze the acknowledgment of their therapeutic utility by regulatory bodies.
D. Classical or Serotonergic Psychedelics versus Nonclassical Psychedelics: Definition
The definition of psychedelics is still based on a classification from the 1960s, and it continues to be a matter of debate. Classic or serotoninergic psychedelic compounds are so called mainly because they interact with the serotonergic system and most of them derive from plants or are semisynthetics (Andén et al., 1968; Glennon et al., 1984; Vollenweider et al., 1998). Serotonergic psychedelics include 1) the semisynthetic ergoline LSD; 2) plant-derived tryptamines, such as psilocybin (the active ingredient found in “magic mushrooms”), N,N-dimethyltryptamine (DMT, the active ingredient in ayahuasca), 5-methoxy-N,N-dimethyltryptamine (5-MeO-DMT), and lysergic acid amide (the active ingredient in morning glories); and 3) phenethylamines, such as mescaline (the active ingredient in the peyote and San Pedro cacti), and phenethylamine-based synthetic designer drugs (Vollenweider et al., 2007; Maciulaitis et al., 2008; Passie et al., 2008; Vollenweider and Kometer, 2010; Dean et al., 2013; Steiner and Leistner, 2018; Dinis-Oliveira et al., 2019). In some cases, they share part of the chemical structure with the endogenous neurotransmitter serotonin (5-HT)—in particular, the indole scaffold. However, some of them, like mescaline, do not possess an indole but are still considered serotonergic psychedelics.
Serotonergic psychedelics are mostly found in nature (e.g., psilocybin, DMT, mescaline, and LSD, which is derived from naturally occurring ergotamine); however, LSD can also be created in a laboratory by modifying natural psychedelics (Hofmann, 1979). Newer analogs and derivatives of pharmacophore lysergamides, tryptamine, and phenethylamine structures are serotoninergic psychedelics even if considered “nonclassical” (Johnson et al., 2019). Despite their name, however, the pharmacology of serotonergic classic psychedelics is not limited to 5-HTRs but has several layers of complexity that involve several receptor families, homo- and heteroreceptor complexes, and biased intracellular cascades. The complete pharmacological profile of these compounds should therefore be considered when trying to dissect the molecular mechanisms underlying psychedelic-induced therapeutic improvements.
LSD is a semisynthetic ergosterol that can be derived by the naturally occurring ergot alkaloid lysergic acid, which is contained in the rye parasite Claviceps purpurea (Castagnoli and Mantle, 1966; Tsai et al., 1995; Tudzynski and Scheffer, 2004). Psilocybin (4-phosphoryloxy-N,N-dimethyltryptamine) is a substituted indolealkylamine (Hofmann et al., 1958). Ayahuasca is an ethnobotanical preparation used in the Amazon Basin for religious, spiritual, and healing purposes (McKenna and Towers, 1985; Frecska et al., 2016). The tea, which is brewed from plants indigenous to the rainforest, contains DMT and β-carboline alkaloids (harmine, harmaline, and tetrahydroharmine) (McKenna et al., 1984; Yritia et al., 2002; Pires et al., 2009). The β-carbolines harmol, harmine, harmaline, and tetrahydroharmine are obtained from Banisteriopsis caapi (ayahuasca, or “vine of the souls”) and function as monoamine oxidase inhibitors (MAOIs) to block the metabolism of DMT, rendering it orally active (Pähkla et al., 2000; Yritia et al., 2002; Gambelunghe et al., 2008; Carbonaro et al., 2015). These compounds are tricyclic indole alkaloids, which resemble tryptamines (Hamill et al., 2019). The presence of an endogenous counterpart in the pineal gland and retina, 6-methoxytetrahydro-β-carbolin (pinoline), has been reported (Langer et al., 1984; Leino, 1984). A close relative of DMT, 5-MeO-DMT, is found in several plants (such as Anadenanthera peregrine) and in certain species of toads (such as Incilius alvarius, which also produces bufotenine) (Glennon and Rosecrans, 1982; Shen et al., 2010).
E. Dissociative Anesthetics
Compounds that trigger psychedelic-like effects and a dissociative-like state, which partly resembles the anesthetic state, are termed dissociative anesthetics (Evoniuk et al., 1991; Yamakura et al., 2000; Zorumski et al., 2016). Dissociation refers to the disruption of usually integrated functions, such as consciousness, memory, emotions, and behavior (Carlson et al., 2018). Renewed interest has been placed on this class of compounds following the FDA approval of esketamine as a novel antidepressant (Kim et al., 2019; Krystal et al., 2019). The other most known, and more infamous, dissociative anesthetic is PCP (“angel dust”), from which ketamine was derived (Budd and Lindstrom, 1982; Bertron et al., 2018).
Ketamine is a phenylcyclohexylamine derivative consisting of two optical enantiomers, (S)-ketamine and (R)-ketamine (Adams et al., 1978, 1981; White et al., 1985; Leung and Baillie, 1989; Oye et al., 1992; Berman et al., 2000; Yeung et al., 2010; Hashimoto, 2014). The racemic mixture esketamine [containing the (S) and (R) isomers in equal parts], has been approved as a medication by the FDA for treatment-resistant MDD (Zhou and Zhao, 2000; Cristea and Naudet, 2019). Ketamine also possesses analgesic activity, which might stem from interactions with the serotonergic, opioidergic, and endocannabinoid systems, and several voltage-gated sodium channels (Zhou and Zhao, 2000; Schnoebel et al., 2005; Ferreira et al., 2018; Savic Vujovic et al., 2019).
F. Empathogens-Entactogens
A third class of compounds structurally related to psychedelic phenylethylamines has been termed empathogen-entactogen (“having the connotation of producing a touching within”) (Nichols, 1986; Vollenweider, 2001), which we will refer to as entactogens for the rest of this review. Compounds in this class include MDMA and 3,4-methylenedioxyamphetamine (MDA) (Riedlinger and Riedlinger, 1994). Although entactogens do not induce a full-blown distortion of perception and consciousness, as with the other psychedelics discussed here, their empathogenic properties are nonetheless relevant for this review. These effects have been broadly examined in psychotherapy augmentation in the context of PTSD, and potential effects on fear extinction and memory reconsolidation have been suggested (Feduccia and Mithoefer, 2018; Sessa et al., 2019; Bahji et al., 2020). MDMA has been considered by the FDA superior to current pharmacological therapies for PTSD and granted the breakthrough designation for treatment-resistant PTSD (Feduccia et al., 2019).
G. Full Dosing versus Microdosing
Most human studies so far have used “full psychedelic doses,” which elicit profoundly altered states of consciousness. The occurrence of such effects is perhaps part of the reason why psychedelic compounds are classified as schedule 1 drugs—that is, substances with high abuse potential and no recognized medical value. More recently, somewhat circumnavigating the hurdle of deeply altered states of consciousness, the concept of “microdosing” has emerged, which consists in taking a “dose of drug that is 1% of the pharmacologically active dose, up to a maximum of 100 µg” (Tewari and Mukherjee, 2010). This kind of dosage is well below the “psychedelic threshold,” eliciting minimal or no psychoactive effects, and it is mostly sought for cognitive and emotional enhancement (Hutten et al., 2019; Rifkin et al., 2020) or by patients who are dissatisfied with conventional treatments to improve mental health or cease prescription or illegal substance use (Hutten et al., 2019; Lea et al., 2020). Despite the lack of systematic studies investigating beneficial outcomes and potential long-term side effects of microdosing, a growing number of individuals are seeking this alternative therapeutic approach, sometimes believing it might be a universal remedy for all somatic and psychological ailments. However, the lack of supporting clinical research has prompted some researchers in the field to demystify these beliefs, highlighting that the notion that microdosing enhances cognition and “cures” psychiatric illness is not supported by scientific research in the field, which in fact has more questions than answers (Kuypers et al., 2019; Preller, 2019).
The most used substances for microdosing among the general population are LSD, followed by psilocybin (Rosenbaum et al., 2020). Microdosing has been described by microdosers as “embracing traditional middle-class values” (Webb et al., 2019), and this approach might be more easily managed by naïve patients compared with full doses and better received by psychiatrists who have no experience in dealing with psychedelic effects. It may even become a therapy for the elderly, and preliminary evidence suggests safety and tolerability of LSD microdosing in elderly populations (Family et al., 2020). Preliminary reports suggest that microdosing has low but appreciable consciousness-altering effects (Prochazkova et al., 2018; Bershad et al., 2019; Yanakieva et al., 2019; Hutten et al., 2020) and enhances mood and focus (Anderson et al., 2019; Webb et al., 2019; Lea et al., 2020; Hutten et al., 2020), cognitive performance (Prochazkova et al., 2018), and empathy (Johnstad, 2018) and decreases mind wandering (Polito and Stevenson, 2019). Microdosing might also help alleviate symptoms of existing medical conditions (Fadiman and Korb, 2019) potentially given the powerful anti-inflammatory effects of psychedelic 5-HT2A receptor agonists (Yu et al., 2008; Nau et al., 2013, 2015; Flanagan et al., 2019b). In fact, strong biological effects have been reported in response to very low doses of psychedelic compounds, such as the potent anti-inflammatory effects of the highly selective 5-HT2A receptor agonist 2,5-dimethoxy-4-iodoamphetamine (DOI), demonstrated extensively in both in vivo and in vitro paradigms (Nau et al., 2013, 2015; Flanagan and Nichols, 2018; Flanagan et al., 2019a,b). Although further studies are required, preliminary findings suggest that microdoses might elicit therapeutic improvements without eliciting profound changes in consciousness. Some have reported microdosing in preparation for full doses of other psychedelics (Johnstad, 2018). Other potential applications are the treatment of migraines, cluster headaches (Andersson et al., 2017), and pain conditions (Johnstad, 2018). Concerning LSD, some authors have suggested that doses <20 μg might be considered microdoses, doses between 21 and 30 μg might be considered minidoses, and doses >30 might be considered psychedelic doses (Holze et al., 2020). In preclinical studies, antidepressant-like effects and enhancement of fear-memory extinction were observed (Cameron et al., 2019), suggesting microdosing might be valuable for comorbid depression and PTSD.
Contrasting the increasingly popular belief that microdosing might represent a universal cure, preliminary preclinical findings support the notion that microdosing might have anxiogenic (not anxiolytic) and neurotic effects (Horsley et al., 2018; Anderson et al., 2019; Polito and Stevenson, 2019), induce confusion (Hutten et al., 2020) and potentially impact metabolism and hinder synaptic plasticity (Cameron et al., 2019), which unequivocally are undesirable effects in psychiatry. Although the concept of microdosing raises the intriguing possibility of decoupling the therapeutic benefits of psychedelic compounds from their psychedelic effects, whether this type of regimen 1) elicits the full spectrum of therapeutic improvement as full doses, 2) presents abuse potential, and 3) elicits acute and long-term side effects remains to be ascertained (the available evidence is discussed throughout the present work).
Preliminary data suggesting that about 1 in 5 patients experience physical discomfort and that about 1 in 10 experience anxiety when microdosing (Anderson et al., 2019) should be considered in future clinical trials, and studies should aim at elucidating the underlying mechanisms, attempting to pharmacologically counteract these side effects. The main off-target effects that are cause for concern so far revolve around the repeated stimulation of cardiac 5-HT2B receptor by microdosing serotonergic psychedelics, which might increase the likelihood of developing cardiovascular disease (discussed in the Abuse Potential and Potential Long-Term Side Effects of Psychedelic Compounds and Side Effects of Psychedelic Compounds sections). Other drugs with 5-HT2B receptor activity have been retracted from the market because of these effects (Connolly et al., 1997), prompting the need to assess potential deleterious effects of psychedelic microdosing on repeated cardiac 5-HT2B receptor stimulation.
Although we do not discredit the possibility that microdosing may potentially be a valuable therapeutic approach, there is not enough available evidence to support this possibility yet. Further larger RCTs are needed to determine the safety, efficacy, and tolerability of microdosing on large cohorts while systematically assessing potential acute and long-term side effects that might arise from this type of treatment. Undoubtedly, given the promising results obtained so far, the increasing number of individuals who self-medicate with psychedelics, and the safety concerns revolving around potential neurobiological and systemic side effects, priority governmental funding should be made available to fill this knowledge gap. Ongoing and future studies should aim at identifying the specific indication, doses, and duration, as well as short- and long-term side effects of each psychedelic compound for specific psychiatric or somatic conditions.
H. Historical Studies
Aside from pharmacological and behavioral studies in rodents (Uyeno and Benson, 1965; Bílková et al., 1971a,b), cats (Hobson, 1964; Kay and Martin, 1978), spiders, (Groh and Lemieux, 1968), snails (Abramson and Jarvik, 1955), and elephants (West et al., 1962; Siegel, 1984), hundreds of human studies involving tens of thousands of people were performed with psychedelic compounds between the 1950s and 1970s. The main areas of interest included 1) psychotherapy augmentation (Cohen and Eisner, 1959; Pahnke et al., 1970), 2) schizophrenia research (Osmond and Smythies, 1952; Cholden et al., 1955; Bolton, 1962), 3) trauma retrieval (Spencer, 1963), 4) social impairments (Cheek and Holstein, 1971; Krsiak et al., 1971; Vojtĕchovský et al., 1972), 5) alcoholism (O’Reilly and Funk, 1964; Smart and Storm, 1964), 6) neurosis (Denson and Sydiaha, 1970), and 7) cognitive enhancement (Jarvik et al., 1955; Goldberger, 1966). Case reports are available in which LSD treatment was reported to improve sexual perversion and pedophilic impulses (Ball and Armstrong, 1961) and ameliorate anxiety-induced compulsive habits, as well as to “cure” nonbinary sexuality and “relieve the fear of being homosexual” (Abramson, 1955; Ball and Armstrong, 1961). Although most of these studies do not satisfy contemporary scientific research standards, they nonetheless represent a wealth of valuable investigations and could help with the development of modern study design.
I. Ongoing Clinical Trials
Contemporary RCTs investigating safety and efficacy of psychedelics for the amelioration of mood disorders suggest that these compounds produce a number of desirable improvements in psychiatric populations and healthy volunteers, such as 1) rapid and sustained antidepressant and anxiolytic effects (Murrough et al., 2013b; Carhart-Harris et al., 2016a, 2018; Palhano-Fontes et al., 2019), 2) decreased suicidality and emotional distress (Hendricks et al., 2015; Argento et al., 2017), 3) positive personality changes (Bouso et al., 2018), 4) brain activity and connectivity changes (Carhart-Harris et al., 2012; Palhano-Fontes et al., 2015), and 5) long-term neuromorphologic changes, which correlate with positive psychological outcomes (Bouso et al., 2015).
After many years of preclinical and clinical research, esketamine has been approved by the FDA as a medication for treatment-resistant depression. As of August 2020, there are 260 clinical trials investigating ketamine for the treatment of several disease states that are listed as ongoing, recruiting, or not yet recruiting on ClinicalTrials.gov (for a list of clinical trials investigating ketamine application in psychiatry, see Supplemental Table 1) [also reviewed in Peyrovian et al. (2020))]. Ongoing and future clinical trials aim to assess the effects of ketamine on 1) suicidality; 2) treatment-resistant MDD; 3) bipolar disorder (BD); 4) ASD; 5) borderline personality disorder; 6) cluster headaches; 7) epilepsy; 8) asthma; 9) subarachnoid hemorrhage; 10) renal colic; 11) opioid, alcohol, cocaine, marijuana, and cigarette use; and 12) as a pre- or perioperative preventative antidepressant in populations with depression. Nevertheless, several studies are investigating the analgesic properties of ketamine for critically ill patients with cancer, pediatric fracture reduction, traumatic injuries, and major surgeries. Other RCTs involving ketamine are investigating neurobiological and biochemical biomarkers of successful ketamine antidepressant treatment, which may be used as treatment-efficacy predictors in treatment-resistant populations (reviewed in Supplemental Table 1).
The focus of MDMA clinical research has mostly revolved around the empathogenic effects of this substance, which seems useful as an augmenting agent in psychotherapy (Mithoefer et al., 2016). In controlled clinical settings, MDMA induces a relaxed euphoric state, feelings of emotional openness, enhanced empathy, and disinhibition, creating a cathartic moment in which trauma can be reaccessed, re-elaborated, and integrated (Liechti et al., 2001; Holze et al., 2020). Although phase III RCTs are investigating the safety of large-scale employment of MDMA-augmented psychotherapy for PTSD survivors, other trials are investigating the safety and efficacy of the drug for the treatment of ASD and alcohol abuse disorder and the brain circuits mediating the observed clinical improvements (see Table 1 for a list of clinical trials investigating the application of MDMA in psychiatry).
Clinical trials available on ClinicalTrials.gov investigating the use of MDMA as a potential therapeutic approach for PTSD and other psychiatric disorders
For completed trials, the references of published work were added if available either from the ClinicalTrials.gov web site or PubMed upon searching the NCT identifier. If information was available on the main findings of the study, it was added. If the trial was not completed or it was completed but no information was available in the form of a published original manuscript, we reported the main research question(s) the study aims to address.
Investigations into the potential therapeutic effects of psilocybin have yielded promising results for treatment-resistant depression (Carhart-Harris et al., 2016a, 2017, 2018), tobacco addiction (Garcia-Romeu et al., 2014; Johnson et al., 2014), and OCD (Moreno et al., 2006), as well as anxiety and distress associated with a life-threatening illness (Griffiths et al., 2016; Ross et al., 2016). Ongoing RCTs are investigating the utility of psilocybin for MDD, treatment-resistant MDD, alcohol use disorder, smoking cessation, cocaine addiction, anorexia nervosa, cognitive impairment and Alzheimer disease, OCD, chronic cluster headache, migraine headache, and post-traumatic headache (see Table 2 for a list of clinical trials investigating the application of psilocybin in psychiatry).
Clinical trials available on ClinicalTrials.gov investigating the use of psilocybin as a potential therapeutic approach for MDD and other psychiatric disorders
For completed trials, the references of the published work were added if available either from the ClinicalTrials.gov web site or PubMed upon searching the NCT identifier. If information was available on the main findings of the study, it was added. If the trial was not completed or it was completed but no information was available in the form of a published original manuscript, we reported the main research question(s) the study aims to address.
LSD has been investigated in RCTs for its anxiolytic properties (Gasser et al., 2014; Gasser et al., 2015). LSD seems to help patients with a potentially life-threatening illness elaborate and accept the emotions connected to the potentially terminal nature of their illness. The main improvements observed include decreased anxiety and depression, increased acceptance, and decreased fear toward their potential imminent death (Gasser et al., 2015). Ongoing RCTs are assessing the efficacy of LSD for the treatment of MDD and anxiety disorders and cluster headache. Studies on LSD microdosing are also underway (see Table 3 for a list of clinical trials investigating the application of LSD in psychiatry).
Clinical trials available on ClinicalTrials.gov investigating the use of LSD as a potential therapeutic approach for MDD, anxiety, and other psychiatric disorders
For completed trials, the references of published work were added if available either from the ClinicalTrials.gov web site or PubMed upon searching the NCT identifier. If information was available on the main findings of the study, it was added. If the trial was not completed or it was completed but no information was available in the form of a published original manuscript, we reported the main research question(s) the study aims to address.
Although ayahuasca has been shown to rapidly relieve depression symptoms in treatment-refractory depression in controlled clinical settings (Osório et al., 2015; Palhano-Fontes et al., 2019), the use of DMT and 5-MeO-DMT in naturalistic settings is receiving considerable attention given the rapid improvements elicited in several mental health domains accompanied by long-lasting decreases in drug and alcohol use and abuse (Davis et al., 2019; Uthaug et al., 2019, 2020; Garcia-Romeu et al., 2020). In vitro evidence supports the notion that 5-MeO-DMT might be beneficial for drug and alcohol addiction. For example, 5-MeO-DMT downregulates metabotropic glutamate receptor (mGluR) 5 in human cerebral organoids, and this gene is involved in alcohol- and drug-induced rewards (Dakic et al., 2017) (see Table 4 for a list of clinical trials investigating the application of ayahuasca and DMT in psychiatry).
Clinical trials available on ClinicalTrials.gov investigating the use of ayahuasca and DMT for treatment-resistant depression and in healthy individuals
For completed trials, the references of published work were added if available either from the ClinicalTrials.gov web site or PubMed upon searching the NCT identifier. If information was available on the main findings of the study, it was added. If the trial was not completed or it was completed but no information was available in the form of a published original manuscript, we reported the main research question(s) the study aims to address.
II. Psychedelic Compounds as Neuroplastic Agents
A. Neuroplasticity Impairments in Psychiatric Disorders
Neuroplasticity refers to the ability of the nervous system to respond to internal and external stimuli via a remodulation of its physical structure and functional connections (Pittenger and Duman, 2008; Cramer et al., 2011). Neuroplasticity occurs on several levels, starting from signal transduction, to gene expression, to synaptic, neuronal, network, and whole-brain adaptations (Duman, 2002; Pittenger and Duman, 2008; Abel et al., 2013). Ultimately, if neuroplasticity leads to a gain of function, it is useful for adaptation, shaping behaviors which help the system adapt to a changing environment (Cramer et al., 2011). However, a dysregulation of neuroplasticity can lead to detrimental adaptive changes and the development of psychiatric illness (Kuipers et al., 2003). Genetic variability in DNA regions that span areas involved in neuroplasticity represent resilience or vulnerability factors in developing psychiatric disorders (Belsky et al., 2009). Not surprisingly, psychiatric drugs elicit therapeutic improvements on behavior at least partially via a remodulation of neuroplasticity-related processes, events which ultimately tend to restore neuroplasticity and behavior (Duman et al., 1994; D’Sa and Duman, 2002).
Psychiatric disorders share neuroplasticity impairments, such as the reduction in hippocampal (MacQueen et al., 2003) and cortical volumes (Bremner et al., 2000; Schmaal et al., 2017) observed in patients with depression and the progressive prefrontal atrophy observed in patients with PTSD who had worsening symptoms (Cardenas et al., 2011). Although the trend appears to be an overall reduction in brain volume in psychiatric disorders, condition-specific symptoms are associated with discrete neuromorphologic changes. For example, greater self-reported anxiety correlates with greater inferior temporal cortex volume decrease in PTSD, contrasting with the slight global volume increase in patients with MDD, or higher scores of self-reported depression, which are correlated with a slight overall volumetric decrease in PTSD together with a slight increase in cuneus and precuneus volume (Kroes et al., 2011). The expression of neurotrophic factors, such as brain-derived neurotrophic factor (BDNF), which modulates neurogenesis, is decreased in psychiatric disorders (Malberg et al., 2000; Molendijk et al., 2014).
Neurogenesis is an essential step to achieve antidepressant outcomes in animal models and humans, and its blockade leads to the development of depressive-like behavior, making it a critical mediator of stress resilience and susceptibility (Ampuero et al., 2010; Kraus et al., 2017). Accordingly, postmortem studies found a reduction in hippocampal dentate gyrus granule cells in medication-free patients with depression compared with treated patients and controls (Gururajan et al., 2016). Neurogenesis-enhancing approaches also seem useful for PTSD paradigms by boosting hippocampal neurogenesis and promoting fear-memory extinction, ultimately decreasing PTSD-like behavior (Ishikawa et al., 2016, 2019). Similar to psychiatric disorders, neurodegenerative disorders are accompanied by decreased neurogenesis (Hollands et al., 2017), although high comorbidity exists among psychiatric and neurologic conditions (Silberstein, 2001; Nuti et al., 2004; Nuyen et al., 2006). This suggests that the potential therapeutic reach of psychedelic compounds might extend to neurodegenerative states and potentially elderly populations with neurodegenerative disorders (Family et al., 2020). Future clinical trials should investigate the efficacy and potential side effects profile of a similar therapeutic approach in elderly populations with neurodegenerative conditions.
BDNF represents a prototypical neurotrophin in the brain that interfaces stress susceptibility, psychiatric disorders, and antidepressant response (Shimizu et al., 2003a; Martinowich et al., 2007; Martinowich and Lu, 2008). For example, decreased BDNF levels are consistently observed across psychiatric disorders such as MDD (Kim et al., 2007), PTSD (Dell’Osso et al., 2009; Angelucci et al., 2014), and BD (Palomino et al., 2006; Reinhart et al., 2015), and low BDNF levels correlate with suicidal behavior (Kim et al., 2007; Lee et al., 2007). Psychotic depression (Lee et al., 2007), but not SCZ, is associated with higher BDNF levels compared with controls (Shimizu et al., 2003b). Not surprisingly, pharmacological antidepressant therapies, sleep, and physical exercise increase BDNF levels (Shimizu et al., 2003a; Lu et al., 2014), and these effects might be mediated at least partially by 5-HT2A signalling in the hippocampus and the neocortex (Vaidya et al., 1997). In terms of predictive biomarkers of treatment response, higher baseline BDNF levels correlate with greater depression improvement after SSRIs (Wolkowitz et al., 2011). Similarly, PTSD is characterized by low BDNF levels (Angelucci et al., 2014), and genetic variability within the BDNF gene confers susceptibility or resilience to PTSD (Hemmings et al., 2013; Zhang et al., 2014). It has also been suggested that BDNF may be a common denominator in PTSD-BD comorbidity (Kauer-Sant’Anna et al., 2007; Rakofsky et al., 2012). Preclinical models indicate that individual susceptibility to PTSD might result from sustained transcriptional downregulation of BDNF (Kozlovsky et al., 2007), which might be mediated by epigenetic processes such as DNA methylation and histone acetylation (Roth et al., 2011) and an increased expression of the BDNF receptor, the neurotrophic receptor tyrosine kinase 2 (NTRK2, also known as tropomyosin related kinase B [TrkB]), in brain areas susceptible to stress, such as the cornu ammonis (CA) 1 region of the hippocampus (Kozlovsky et al., 2007).
B. Effects of Psychedelic Compounds on Neuronal and Synaptic Plasticity
BDNF enhances AMPA receptor delivery to the synapse (Caldeira et al., 2007), and this mechanism might be involved in the psychedelic-induced, BDNF-mediated enhancement of neural plasticity elicited by psychedelic compounds (Ly et al., 2018). This possibility is supported by the fact that AMPA receptor potentiation is beneficial in the treatment of psychiatric disorders (Zarate and Manji, 2008) via enhancing neuroplastic pathways (Seo et al., 2020). Importantly, given that AMPA receptors mediate the formation of neural networks during development (O’Neill et al., 2004), this effect might be involved in the reported “resetting” feelings experienced by individuals who ingest psychedelics (discussed in the Going Beyond Receptors: Neuronal Circuits Activated by Psychedelic Drugs section) (Carhart-Harris et al., 2017). Stimulation of neurotrophic pathways, leading to increased neurogenesis, cognitive flexibility, and ultimately changes in behavior, has long represented a desirable characteristic in psychiatric drugs, and the neuroplastic and cognition-enhancing effects elicited by psychedelics could be exploited to achieve such outcomes.
Psychedelics elicit profound changes in neuronal and synaptic plasticity through mammalian target of rapamycin (mTOR) and BDNF-NTRK2 signaling (see Fig. 1 and Table 5 for details) (Li et al., 2010; Dakic et al., 2017; Ly et al., 2018). The neuroplasticity-enhancing effects of serotonergic psychedelics resemble those of the dissociative anesthetic ketamine, which boost neural and synaptic plasticity, at least partially, via potentiating BDNF and mTOR signaling (Li et al., 2010; Ly et al., 2018). Given these similarities, it seems likely that the stimulation of neurotrophic pathways by psychedelics is involved in the psychiatric improvements observed in clinical trials (Berman et al., 2000; Carhart-Harris et al., 2017; Mithoefer et al., 2018; Palhano-Fontes et al., 2019).

Neuroplastic effects of psychedelic compounds relevant to psychiatric disorders and comorbidities. The main outcomes elicited by psychedelic compounds on neuroplastic and neurogenesis-related pathways are reported in the green box (purple brain). The resulting outcomes on psychiatric symptoms are reported in the blue box (orange brain). For each compound, or group of compounds with similar pharmacology, the main classes of transcription factors and signaling pathways activated that are thought to mediate the effects of the compound on synaptic and neuronal plasticity are reported. In the bottom part of the figure, the main receptors involved in signal transduction for each compound or group of compounds with similar pharmacology and the resulting neurotransmitter released are reported. AR, androgen receptor; ALT, alternative lengthening of telomeres; ANIA3, activity and neurotransmitter-induced early gene 3; CAMK2, calcium/calmodulin-dependent protein kinase 2; CEBPB, CCAAT/enhancer-binding protein beta; COMT, catechol O-methyltransferase; CREB, cAMP response element-binding protein; MKP1, mitogen-activated protein kinase phosphatase; NOR1, neuron-derived orphan receptor-1; IKB, inhibitor of kB kinase.
Main receptor interactions and downstream effects of psychedelic compounds on pathways involved in synaptic and neuronal plasticity, neuroimmunomodulation, and modulation of neurotransmitter systems of relevance to psychiatry
For each compound (column 1), the receptors involved in signal transduction are reported with decreasing Ki (column 2). The transcription factors, enzymes, hormones, and cytokines up/downregulated by each compound are reported in terms of acute and chronic responses (where data are available, column 3). The main outcomes on modulation of neurotransmitter systems are reported in column 4. Systemic and psychological effects elicited are reported for each compound (column 5). Referenced articles are reported in column 6.
1. Ketamine
Subanesthetic doses of ketamine elicit rapid, robust, and sustained antidepressant effects (Berman et al., 2000; Price et al., 2009; Murrough et al., 2013a) at least partially via activating mTOR (Li et al., 2010), a key intracellular signaling pathway altered in MDD and other psychiatric disorders, such as ASD and SCZ (Li et al., 2010; Jernigan et al., 2011; Costa-Mattioli and Monteggia, 2013). In turn, this leads to the BDNF-mediated stimulation of dendritic, synaptic, and neuronal plasticity, which may well translate to network plasticity (Murrough et al., 2013b; Choi et al., 2017; Ly et al., 2018). Ketamine treatment increases BDNF levels in the prefrontal cortex (PFC) and CA3 and dentate gyrus regions of the hippocampus, and the latter region is intimately involved with adult neurogenesis (Dong et al., 2017). Moreover, ketamine treatment during stress exposure prevents the dendritic spine density loss observed in the CA3 region and dentate gyrus of the hippocampus of controls subjects (Dong et al., 2017). Similarly, ketamine increases spine density and synaptic plasticity markers such as synapsin 1, postsynaptic density protein 95, and mGluR1 in rats exposed to stress (Sarkar and Kabbaj, 2016).
Ketamine also interacts with σ1 receptor (S1R, discussed in the immunomodulation section) and S2R, key signal transducers of neuroplastic and neurotrophic pathways that at least partially mediate the therapeutic improvements elicited by currently approved antidepressant medications (Yagasaki et al., 2006; Dhir and Kulkarni, 2007; Fishback et al., 2010; Fujimoto et al., 2012; Krystal et al., 2019). For this reason, S1R and S2R may be involved in ketamine’s antidepressant action. (R)-Ketamine possess higher affinity for S1R than its enantiomer (S)-ketamine, and S1R antagonism does not decrease the antidepressant effects of ketamine (Robson et al., 2012). However, S1R antagonism does decrease the nerve growth factor (NGF)-mediated neuroplastic effects of ketamine. These results demonstrate that the ketamine-initiated, S1R-mediated modulation of NGF might be involved in the antidepressant effects elicited by ketamine (Fujimoto et al., 2012; Yagasaki et al., 2006; Krystal et al., 2019). Lastly, low-dose ketamine increases hippocampal AMPA/NMDA receptor density ratio in a preclinical MDD model (Tizabi et al., 2012). Therefore, decreased AMPA/NMDA receptor density ratio could be involved in the pathogenesis of MDD (Aleksandrova et al., 2017), and ketamine might alter this ratio to ultimately enhance synaptogenesis via upregulating mTOR/BDNF signaling (Tizabi et al., 2012; Aleksandrova et al., 2017).
2. N,N-Dimethyltryptamine, 5-Methoxy-N,N-dimethyltryptamine, and Ayahuasca
Neuroplastic effects have been observed in response to ayahuasca. For example, a single dose of ayahuasca increases circulating BDNF levels in healthy controls and patients with treatment-resistant depression (de Almeida et al., 2019). Preclinical findings support that neuroplasticity-enhancing effects might mediate therapeutic improvements in response to psychedelics. For instance, repeated administration of ayahuasca increases hippocampal BDNF levels in female rats (Colaço et al., 2020). Moreover, a single dose of 5-MeO-DMT increases neurogenesis in the dentate gyrus of the adult mouse (Lima da Cruz et al., 2018). This is of particular relevance given that antidepressant-free patients with depression have decreased BDNF levels (Diniz et al., 2010), and antidepressants increase BDNF (Aydemir et al., 2005), thereby enhancing hippocampal neurogenesis (Anacker et al., 2011). Similarly, other nonpsychedelic S1R agonists suppress neurodegeneration and attenuate disease progression in preclinical models via boosting neuronal proliferation and maturation (Ono et al., 2014), increasing hippocampal BDNF, and activating antioxidant pathways (Kikuchi-Utsumi and Nakaki, 2008; Pal et al., 2012).
S1R modulation might represent a synergistic mechanism to 5-HT2A receptor modulation by psychedelics, which mediates the upregulation of neurotrophic factors. In fact, S1Rs are enriched in brain areas involved with cognition, stress, and psychiatric disorders, and their stimulation induces adaptive neuroplasticity (Hindmarch and Hashimoto, 2010); psychedelic and nonpsychedelic S1R agonists ameliorate psychiatric symptoms (Hindmarch and Hashimoto, 2010; Davis et al., 2019; Palhano-Fontes et al., 2019). The other active compounds contained in ayahuasca are the β-carbolines harmol, harmine, harmaline, and tetrahydroharmine (found in B. caapi), which inhibit the metabolism of DMT to render it orally active. These compounds also elicit neuroplastic effects. In vitro studies found that these molecules induce differentiation, proliferation, and migration of neural precursors (Morales-Garcia et al., 2017). This strengthens the notion that ayahuasca alkaloids might be useful for neurodegenerative states. Contrasting preclinical evidence suggests that repeated DMT might hinder synaptic plasticity (a sex-specific effect in females) (Cameron et al., 2019). Although preliminary, these results highlight the need for further preclinical studies with greater statistical power to identify potential neuroplasticity-related side effects.
3. Lysergic Acid Diethylamide
It was recently reported that microdoses of LSD increase circulating BDNF levels in healthy volunteers, suggesting that LSD might have neurotrophic effects in humans (Hutten et al., 2020). Another study investigated whether LSD produces acute gene expression changes in peripheral blood mononuclear cells 1.5 and 24 hours after administration, with no detected changes in 5-HT2A receptor or EGR-1, -2, and -3 transcript levels (Dolder et al., 2017a). In rodents, LSD upregulates CCAAT/enhancer-binding protein-β in the prefrontal cortex (Nichols and Sanders-Bush, 2004). This transcription factor affects synaptic scaling, an essential “housekeeping” neuronal process that modulates synaptic function (Turrigiano, 2008). Consistent with neuroplastic and neuroprotective outcomes, LSD increases cFOS, Egr-1, and Egr-2 in murine primary neuronal cultures (González-Maeso et al., 2003, 2007; Li et al., 2005). Moreover, LSD increases dehydroepiandrosterone (DHEA), the most abundant neurosteroid in the central nervous system (Strajhar et al., 2016). Similar to DMT (Fontanilla et al., 2009), DHEA is an S1R agonist, and signaling of DHEA and pharmacological compounds at S1R stimulates synaptic activity and neurogenesis and ameliorates drug-induced cognitive impairments (Meunier and Maurice, 2004; Moriguchi et al., 2013). This indirect action of LSD on S1R might be involved in the clinical improvements elicited by LSD (Moriguchi et al., 2013; Szabo et al., 2014; Schmidt et al., 2016). S1R forms heterodimers with D2 receptors, and such interaction may also boost neurogenesis (Beggiato et al., 2017). Lastly, DOI, a close relative of LSD, rapidly increases spine density growth in cortical cells, activating a synaptogenic pathway mediated by kalirin 7, while increasing Egr-1 in the mouse somatosensory cortex (Jones et al., 2009). Given that the EGR family of transcription factors is involved in synaptic plasticity, neurogenesis, and the pathologic processes underlying psychiatric symptoms, the effects of psychedelics on EGR-mediated transcription should be further elucidated (González-Maeso et al., 2003, 2007; Clark et al., 2010; Duclot and Kabbaj, 2017).
4. Psilocybin
A recent study reported that psilocybin modulates neurotrophic-related gene expression in the PFC and hippocampus, with a preferential effect on the PFC, but inducing changes which are also appreciable in the hippocampus (Jefsen et al., 2020). More specifically, in the PFC psilocybin increased the expression of CEBPB, c-Fos, dual specificity protein phosphatase 1 (Dusp1), transcription factor jun-β, NF-kappa-β inhibitor-α (Iκβα), nuclear receptor subfamily 4 group A member 1, while the serine/threonine-protein kinase 1 (Sgk1), protein fosB, protein S100-A10, and postsynaptic density protein 95 were increased only response to some doses of psilocybin. Dual specificity protein kinase (CLK1) was dose dependently decreased in response to psilocybin (Jefsen et al., 2020). In the hippocampus, Dusp1, Iκβα, and Sgk1 transcripts were similarly increased by acute psilocybin and Clk1 expression was strongly decreased (Jefsen et al., 2020). Psilocybin affects hippocampal neurogenesis in a biphasic fashion (Catlow et al., 2013). Although at lower doses (0.1 mg/kg) a nonstatistically significant trend was observed toward increased neurogenesis, higher doses (1–5 mg/kg) decreased neurogenesis 2 weeks after treatment (Catlow et al., 2013). These neuroplastic changes were associated with a facilitation of fear extinction at low doses, hypothetically mediated by a psilocybin-induced, 5-HT–mediated DA enhancement, an effect known to facilitate fear-extinction learning (Borowski and Kokkinidis, 1998; Catlow et al., 2013).
5. 3,4-Methylenedioxymethamphetamine
The debate on the neurotoxicity of MDMA creates a long-standing divide (Mithoefer et al., 2003; Parrott, 2013, 2014; Doblin et al., 2014; Pantoni and Anagnostaras, 2019; Ricaurte et al., 2002 [retracted]). Most of the available early preclinical research is focused on the neurotoxic effects of MDMA, which may explain cognitive impairments and psychiatric sequelae in MDMA abusers (Parrott, 2001, 2013). To simulate binge abuse and the resulting neurotoxic effects, relatively high doses (often in a chronic-administration design) were administered in these studies, with considerable neurotoxicity (Pantoni and Anagnostaras, 2019). Indeed, human studies indicate damaging effects on SERT homeostasis in heavy MDMA users (Baumann et al., 2007; Müller et al., 2019). However, although these studies represent a valid paradigm for MDMA abuse, they do not seem to adequately model the sparing use of relatively low doses employed in MDMA-augmented psychotherapy (Amoroso, 2019; Pantoni and Anagnostaras, 2019), which elicits notable improvements in treatment-refractory PTSD symptoms (Mithoefer et al., 2011, 2013; Mithoefer et al., 2018; Feduccia et al., 2019; Bahji et al., 2020).
Importantly, metabolites from MDMA metabolism seem to be responsible for the neurotoxic effects of MDMA given that direct intracerebroventricular administration of MDMA does not elicit neurotoxicity (Green et al., 2003). Accordingly, intrastriatal administration of 2,4,5-trihydroxymethamphetamine significantly depletes both 5-HT and DA, intracortical administration decreases 5-HT, and intracerebroventricular administration moderately depletes striatal DA without affecting 5-HT levels (Johnson et al., 1992; Zhao et al., 1992). Other metabolites, such as 2-hydroxy-4,5(methylenedioxy)methamphetamine (6-HO-MDMA), appear to be nontoxic given that intrastriatal and intracerebroventricular administration does not affect 5-HT or DA levels (Zhao et al., 1992). Such metabolites are quinone-thioethers, orto-quinones, and the glutathione conjugates 5-(glutathion-Syl)-α-methyldopamine (5-GSyl-α-MeDA) and 2,5-bis-(glutathion-S-yl)-α-methyldopamine (2,5-bis-(glutathione-S-yl)-α-MeDA) (Miller et al., 1996; Bai et al., 1999; Monks et al., 2001; Green et al., 2003).
The existing lack of clear knowledge on potential neurotoxic effects of MDMA could be filled by employing realistic preclinical models that simulate clinically relevant pharmacodynamics (Pantoni and Anagnostaras, 2019; Vollenweider et al., 1999a). If further studies will determine that neurotoxic effects are elicited by MDMA administration in humans at clinically relevant doses, strategies could be implemented in an effort to protect against these effects (Tourino et al., 2010).
III. Psychedelic Compounds as Immunomodulatory and Anti-Inflammatory Agents
A. Inflammation
Inflammation entails a strong but short-lived cascade of events that are mobilized in response to stressful stimuli with the ultimate goal of dealing with the stressor and returning the system to homeostasis. Although a spatiotemporally fine-tuned inflammatory response is key to physical and psychological defense and for tissue repair and remodeling, it presents collateral damage potential if the process is too violent or if it does not reach resolution within a reasonable spatiotemporal frame (Kotas and Medzhitov, 2015; Hotamisligil, 2017). Activation of peripheral inflammatory pathways leads to the activation of central inflammatory cascades directly via 1) circulation of inflammatory mediators in central nervous system lymphatic vessels, 2) active transport and/or compromised blood-brain barrier, and 3) crossing at circumventricular organs (Maier and Watkins, 2003; Banks, 2005; Dantzer et al., 2008; Louveau et al., 2015) and indirectly via stimulating the de novo production of cytokines in the brain (Hanamsagar et al., 2012; Weber et al., 2015; Inserra et al., 2019).
B. Inflammation-Induced Psychiatric Symptoms
Increased proinflammatory signaling in the brain results in the development of “depressive-like” symptoms, a behavioral repertoire encompassing anxiety, low motivation, fatigue, loss of interest, inability to seek and experience pleasure, exaggerated pain responses, lack of concentration, and sleep pattern alterations, manifestations that closely resemble clinical MDD symptomatology (Dantzer et al., 2008). Supporting an involvement of the immune system in the development of psychiatric symptoms, therapies involving the administration of inflammatory molecules (such as interferon-α) and polymorphisms in inflammation-related genes increase the susceptibility to psychosocial stress via affecting immune signaling (Wong et al., 2008; Lotrich, 2009). On the other hand, patients experiencing depressive, PTSD, or BD symptoms present dysregulated reactivity of immune cells accompanied by increased levels of central proinflammatory mediators such as IL1β, IL6, TNF-α, C-reactive protein, and the translocator protein (TSPO), some of which correlate with symptom severity (Rohleder et al., 2004; Dowlati et al., 2010; Setiawan et al., 2015; Lindqvist et al., 2017; Zou et al., 2018) [meta-analyses: (Modabbernia et al., 2013; Passos et al., 2015; Baumeister et al., 2016; Goldsmith et al., 2016; Leighton et al., 2018)]. Psychiatric patients also present abnormal functioning of the hypothalamic-pituitary-adrenal (HPA) axis, one of the most important stress-responsive systems, which increases inflammation upon activation by psychosocial and physical stressors (Melhem et al., 2016; Keller et al., 2017; Dunlop and Wong, 2019).
Patients diagnosed with MDD and PTSD present a systemic low-grade chronic inflammatory state driven by a shift of immune responses toward T-helper (Th) 1 and decreased T-regulatory cell activity (Myint et al., 2005; Gola et al., 2013; Alcocer-Gómez et al., 2014). Such a shift signifies an increase in cell-mediated immune responses and a decrease of humoral immune responses, events which create a proinflammatory milieu that may lead to, or be a consequence of, psychiatric disorders (Myint et al., 2005; Dantzer et al., 2008; Gola et al., 2013; Alcocer-Gómez et al., 2014). Given that in psychiatric disorders there is an imbalance in this equilibrium that is shifted toward Th1 (proinflammatory) responses, therapeutic approaches with the ability to shift this balance in favor of an enhancement of Th2 (immunomodulatory and anti-inflammatory) responses have long been sought and may help ameliorate the psychiatric symptomatology (Pulendran, 2004; Myint et al., 2005; Gola et al., 2013; Alcocer-Gómez et al., 2014). This dysregulated immune milieu is accompanied by pathologic fluctuations in gut microbiome composition that further fuel inflammation, increasing the likelihood of developing comorbid systemic illnesses (Rogers et al., 2016; Inserra et al., 2018; Cheung et al., 2019). The changes elicited by psychedelics over immune function represent a marked shift of inflammatory responses from Th1 to Th2 responses, which overall denotes a shift from proinflammatory to anti-inflammatory and immunomodulatory effects (Romagnani, 1997; Maldonado-López and Moser, 2001; Pulendran, 2004).
C. Anti-Inflammatory Therapies in the Treatment of Psychiatric Disorders
Anti-inflammatory treatments are being tested as adjunctive therapies in psychiatry, but with mixed results. For example, minocycline, a second-generation, semisynthetic tetracycline with anti-inflammatory properties, seems to be a promising adjunctive in patients with MDD, SCZ, and BD and dysregulated inflammation, potentially via its acute effects on inflammatory mediators and long-term effects over gut microbiome composition (Chaudhry et al., 2012; Wong et al., 2016; Dean et al., 2017). Clinical trials partly confirmed the antidepressant effects of minocycline, suggesting it might be a useful first-line or adjunct therapy in psychiatric patients with high baseline inflammatory markers (Raison et al., 2006; Miyaoka et al., 2012; Raison and Miller, 2013; Dean et al., 2017; Rosenblat and McIntyre, 2018). Not surprisingly, classic antidepressant and anti-inflammatory therapies similarly reduce signals of immune activation (such as IL1β, IL2, IL6, and TNF-α), and higher TNF-α levels predict an increased likelihood of treatment resistance (Köhler et al., 2014; Strawbridge et al., 2015). Also relevant for this review, nonpsychedelic S1R agonism inhibits Th1 responses by suppressing interferon-γ (IFN-γ), granulocyte-macrophage colony-stimulating factor and TNF-α and increasing the anti-inflammatory IL10 (Carayon et al., 1995; Zhu et al., 2003). However, clinical studies with anti-inflammatory agents are still under investigation, since a few produced negative results in depression [including the TNF-α inhibitor infliximab (Raison et al., 2013)] or mixed results (Köhler-Forsberg et al., 2019).
D. Immunomodulatory and Anti-Inflammatory Pathways Activated by Psychedelics
Psychedelic compounds activate immunomodulatory and anti-inflammatory programs mediated at least partially by 5-HT2A receptor agonism (see Fig. 2) (Flanagan and Nichols, 2018). Although serotonin-mediated 5-HT2A receptor activation is canonically considered a proinflammatory signal (Shajib and Khan, 2015), psychedelic-induced 5-HT2A receptor activation largely results in the recruitment of anti-inflammatory and immunoregulatory pathways (Nau et al., 2013, 2015; Flanagan and Nichols, 2018; Flanagan et al., 2019b). Such selectivity of anti-inflammatory over proinflammatory effects likely stems from biased signaling cascades (discussed later in the review) mediated by psychedelic-specific conformational stabilization of the 5-HT2A receptor. This leads to the recruitment of psychedelic-specific anti-inflammatory signal transducers, which involve the inhibition of TNF-α and NF-ĸB–propagated proinflammatory signaling (House et al., 1994; Nau et al., 2013; Flanagan et al., 2019b), outcomes which may prove useful for the treatment of autoimmune conditions (Thompson and Szabo, 2020). Ultimately, this effect elicits psychedelic-specific inflammatory gene expression fingerprints. Given that nearly all immune cells express 5-HT receptors (such as macrophages, monocytes, eosinophils, dendritic cells, and natural killer cells) (Ahern, 2011; Baganz and Blakely, 2013), it is not surprising that psychedelics profoundly affect inflammation and immunity, and it is seems highly likely that these effects are involved in the anxiolytic and antidepressant effects elicited by psychedelics. Importantly, the anti-inflammatory and immunomodulatory effects of psychedelics suggest that they might be useful in “inflammaging” (conditions caused or exacerbated by age-induced chronic inflammation) (Franceschi et al., 2018; Aday et al., 2020; Family et al., 2020).

Anti-inflammatory and immunomodulatory effects of psychedelic compounds relevant to psychiatric disorders and comorbidities. The main outcomes elicited by psychedelic compounds on inflammation and immunity-related pathways are reported in the blue box (purple brain). The resulting outcomes on psychiatric symptoms are reported in the purple box (orange brain). For each compound, or group of compounds with similar pharmacology, the main classes of cytokines, chemokines, hormones, transcription factors, and signaling pathways activated, which are thought to mediate the effects of the compound on inflammation and immunity-related pathways, are reported. In the bottom part of the figure, the main receptors involved in signal transduction for each compound or group of compounds with similar pharmacology and the resulting neurotransmitter released are reported. ADRA, alpha adrenergic receptor; ADRB, beta adrenergic receptor; ALPHA2, alpha 2 adrenergic receptor; CCL5, C-C motif chemokine 5; CORT, cortisol; CX3CL1, fractalkine; CXCL10, C-X-C motif chemokine 10; GLU, glutamate; HRH, histamine receptor; I1, imidazoline receptor 1; IKBA, NF-kappa-B inhibitor alpha; ICAM1, intercellular adhesion molecule 1; M3, muscarinic receptor subtype 3; M5, muscarinic receptor subtype 5; SIGMAR, sigma receptor; TAAR, trace amine-associated receptor; TLR, toll-like receptor; TSH, thyroid stimulating hormone; VCAM1, vascular cell adhesion protein 1.
1. N,N-Dimethyltryptamine, 5-Methoxy-N,N-dimethyltryptamine, and Ayahuasca
Strong anti-inflammatory effects mediated by psychedelic-induced S1R activation have been described (Szabo et al., 2014, 2016). These effects closely resemble the anti-inflammatory action of some SSRIs, SNRIs, and tricyclic antidepressants, (Kenis and Maes, 2002; Köhler et al., 2014; Strawbridge et al., 2015) and the antidepressant outcome of some anti-inflammatory therapies (Kappelmann et al., 2018). For example, DMT and its analog 5-MeO-DMT decrease IL1β, IL6, IL8, and TNF-α while increasing IL10 in immune-challenged, human monocyte–derived dendritic cells (Szabo et al., 2014). Similarly, in hypoxic stress–challenged neuronal and microglial cultures, DMT administration enhances cell survival and hypoxic stress–related neuroprotective signaling (Szabo et al., 2016). Accordingly, a single inhalation of 5-MeO-DMT decreases the levels of circulating IL6 (Uthaug et al., 2020). Such fast onset of anti-inflammatory effects is of particular interest and could be harnessed in psychiatry and other areas of emergency medicine—for example, in the treatment of systemic inflammatory response syndrome, also called the “cytokine storm” (Mastronardi et al., 2007; Tisoncik et al., 2012; Szabo et al., 2014, 2016; Mastronardi et al., 2015; Inserra et al., 2019; Mehta et al., 2020; Uthaug et al., 2020). Given that in psychiatric disorders there is an imbalance in immune system equilibria, which are shifted toward proinflammatory responses, therapeutic approaches with the ability to shift this balance in favor of an enhancement of immunomodulatory and anti-inflammatory responses have long been sought in psychiatry (Pulendran, 2004; Myint et al., 2005; Gola et al., 2013; Alcocer-Gómez et al., 2014). Whether psychedelics produce a decrease of proinflammatory cytokines in microglial cells similar to SSRIs and SNRIs, which decrease TNF-α and nitric oxide production and microglial activation, is an intriguing possibility that has been partially demonstrated and that warrants further investigation (Tynan et al., 2012; Szabo et al., 2016).
A study investigating proteomics changes in brain organoids in response to 5-MeO-DMT reported a downregulation of NF-ĸB and nuclear factor of activated T cells via toll-like signaling and Gq-coupled receptors (Dakic et al., 2017). Importantly, those molecules represent critical checkpoints of immune homeostasis interfacing inflammatory responses, mood, and psychiatric disorders (Keller et al., 2017; Zorn et al., 2017). Similarly, ayahuasca and DMT modulate cortisol release (Strassman and Qualls, 1994; Dos Santos et al., 2012; Galvao et al., 2018), and ayahuasca normalizes the blunted awakening cortisol responses observed in patients with treatment-resistant depression (Galvao et al., 2018) while decreasing the circulating levels of C-reactive protein (CRP) in these patients (Galvão-Coelho et al., 2020). Such an HPA axis modulation is quite remarkable given the well characterized dysregulation in psychiatric disorders and the long-sought possibility of modulating the HPA axis to ameliorate psychiatric symptoms (Carvalho and Pariante, 2008; Pariante and Lightman, 2008; Heim et al., 2008; Holsen et al., 2013; Keller et al., 2017).
2. Psilocybin
Psilocybin was shown to acutely increase the circulating levels of the stress-responsive adrenocorticotropic hormone (ACTH) and cortisol, as well as prolactin and thyroid-stimulating hormone (Hasler et al., 2004). The increase in the levels of these hormones was not correlated with stress-induced symptoms, such as anxiety, leading the authors to speculate that the observed temporary increase was likely due to a transient 5-HT2A receptor–mediated activation of the HPA axis, resulting in an increase in the release of ACTH and circulating cortisol (Van de Kar et al., 2001; Hasler et al., 2004).
3. Lysergic Acid Diethylamide
LSD possesses anti-inflammatory activity in vitro. Specifically, LSD downregulates IL2, IL4, and IL6 and upregulates mitogen-activated protein kinase phosphatase-1 (House et al., 1994). Moreover, LSD administration increases circulating cortisol, cortisol, cortisone, corticosterone, prolactin, oxytocin, and epinephrine in humans (Schmid et al., 2015a; Strajhar et al., 2016). No reports are available so far on potential long-term immunomodulatory outcomes of LSD. A recent study investigated the effects of LSD on retinal cells and suggested that LSD might be toxic to retinal cells via releasing proinflammatory cytokines (Hu et al., 2018). However, retinal cells do not adequately model central and peripheral neural networks, and whether the concentrations of LSD employed in this study are reached in retinal cells in vivo remains to be determined. Therefore, these results should be interpreted with caution. Nevertheless, more studies are required to investigate whether LSD administration might be damaging to retinal cells in vivo.
4. 2,5-Dimethoxy-4-iodoamphetamine
DOI has been widely studied for its selectivity as a 5-HT2A/2C receptor agonist and for its powerful anti-inflammatory effects, which can be elicited with very low doses of this compound. DOI binding to the 5-HT2A receptor triggers anti-inflammatory signaling, which is different from 5-HT binding to the same receptor, which elicits proinflammatory signaling (Flanagan and Nichols, 2018). DOI powerfully downregulates inflammation in vitro and in vivo via decreasing TNF-α, IL1β and IL6, intracellular adhesion molecule-1, vascular cell adhesion molecule-1, monocyte chemoattractant protein 1 (MCP1), and fractalkine, ameliorating parameters of immune-induced gut pathology (Yu et al., 2008; Nau et al., 2013; Flanagan et al., 2019). Importantly, DOI blocks NF-ĸB activity and nuclear translocation while decreasing iNOS activity (Miller and Gonzalez, 1998; Yu et al., 2008; Nau et al., 2013). DOI was also shown to be protective in a mouse model of asthma via inhibiting IL3, IL5, IL6, IL13, MCP1, and granulocyte-macrophage colony-stimulating factor (Nau et al., 2015). A recent study suggests that the anti-inflammatory effects of DOI could also be harnessed for the treatment of systemic inflammatory conditions, such as high-fat diet–induced cardiovascular dysfunction (Flanagan et al., 2019b). In this study, the authors observed that DOI treatment during diet-induced cardiovascular inflammation in apolipoprotein-E knockout mice lowers total and low-density lipoprotein cholesterol. Moreover, this treatment decreases IL6, TNF-α, and vascular cell adhesion molecule-1 in the aortic tissue. Furthermore, DOI decreases the circulating levels of the C-X-C motif chemokine 10 (Flanagan et al., 2019b). Together, these findings suggest that DOI and other serotonergic psychedelics might prove valuable for systemic inflammatory diseases, which are often comorbid with psychiatric symptoms.
5. 3,4-Methylenedioxymethamphetamine
MDMA also possesses immunomodulatory properties that influence both the innate and adaptive arms of the immune system (Boyle and Connor, 2010; Mithoefer et al., 2018). Acute administration in humans increases cortisol and prolactin levels (Gouzoulis-Mayfrank et al., 1999) as well as the inflammatory mediators hydroxyeicosatetraenoic acid, dihydroxyeicosatetraenoic acid, and octadecadienoic acid (Boxler et al., 2018) while decreasing circulating CD4+ helper T cells and increasing natural killer cells, potentially as a result of increased HPA axis activation (Pacifici et al., 1999, 2000, 2007; Young, 2004). Increases in the production of immunosuppressive Th2 cytokines, such as IL4 and IL10, and a decrease of Th1 cytokines, such as IL2 and IFN-γ, were also reported (Pacifici et al., 2001). Interestingly, repeated exposure to MDMA decreases the number of PV+ GABA neurons in the hippocampal dentate gyrus, and this effect is attenuated by pretreatment with the anti-inflammatory ketoprofen, suggesting that it might be mediated by cyclooxygenase-mediated pathways (Anneken et al., 2013).
Given that PTSD is underlined by a low-grade chronic inflammatory state and a shift of cytokine production from Th2 to Th1 coupled to chronic lymphocyte activation, the immunomodulatory and anti-inflammatory effects of MDMA might relate to PTSD symptom reduction after MDMA-assisted psychotherapy (Wilson et al., 1999; Boyle and Connor, 2010; Yuan et al., 2016; Mithoefer et al., 2018). MDMA also decreases the production of the proinflammatory cytokines IL1β, IL6, TNF-α, and IFN-γ while stimulating the production of the anti-inflammatory IL10 and oxytocin (OT) (Boyle and Connor, 2010; Yuan et al., 2016). Similarly, MDMA inhibits lipopolysaccharide-induced TNF-α secretion (Connor et al., 2000). These lines of evidence suggest that MDMA might decrease the immune reactivity, and therefore the damaging potential, of immune cells in patients with PTSD (which are hyper-reactive in this disorder), and such an effect might be involved in reduction of clinical symptoms (Wilson et al., 1999; Mithoefer et al., 2018; Bahji et al., 2020).
6. Ketamine
Ketamine displays a biphasic action on immune responses. Acutely, it upregulates IL1β, IL6, and TNF-α. Chronically, it decreases TNF-α (Blandino-Rosano et al., 2017). Mice exposed to highly immunogenic stressors, such as the bacterial toxin lipopolysaccharide, were shown to have higher survival rates when administered ketamine and decreased levels of the proinflammatory TNF-α and IFN-γ, which might be responsible for their improved survival rates (Takahashi et al., 2010). Given the direct role of inflammation in psychiatric symptoms, the anti-inflammatory action of ketamine could relate to its beneficial and fast-onset antidepressant properties (Dantzer et al., 2008; Hotamisligil, 2017). In fact, TNF-α is depressogenic, and TNF-α antagonism is antidepressant (Raison et al., 2013). The short-term proinflammatory effects elicited by ketamine might activate necroptotic pathways, which are required for brain regeneration, a process that may (hypothetically) mediate fear extinction and memory reconsolidation (Feduccia and Mithoefer, 2018; Lloyd et al., 2019).
Together, this body of evidence suggests that the anti-inflammatory and immunomodulatory action of psychedelics might be involved in the antidepressant and anxiolytic effects observed in psychiatric populations via pushing the system toward a homeostatic state and, overall, ameliorating psychiatric symptoms. Future studies should explore whether these immunologic outcomes observed in vitro and in preclinical models translate to human physiology. Safety concerns arising from immunosuppression, even though partial, need to be addressed in immunocompetent and immunodeficient models (Boyle and Connor, 2010).
E. σ1 Receptor
Aside from their well known interaction with the 5-HT system, most of the compounds discussed here, such as DMT, 5-MeO-DMT, DOI, ketamine, and MDMA, all bind S1R, a still-mysterious receptor whose origin remains puzzling (Fontanilla et al., 2009; Frecska et al., 2013; Szabo et al., 2014; Kourrich, 2017; Kim and Pasternak, 2018). This interaction contrasts the 5-HT2A receptor–centric approach to psychedelic activity and suggests that the action of these compounds at S1R might be more relevant than previously thought (Brammer et al., 2006; Fontanilla et al., 2009; Ray, 2010; Nguyen et al., 2014; Szabo et al., 2014). S1R is highly expressed in limbic areas of the human brain and in several central and peripheral immunocompetent cells, such as monocyte-derived dendritic cells and microglia, which are involved in innate and adaptive immune responses (Ishikawa et al., 2007; Fujimoto et al., 2012; Frecska et al., 2013; Szabo et al., 2014, 2016). Interestingly, ibogaine is the only psychedelic to have its highest affinity at this receptor (Ray, 2010). Recently, a study found for the first time evidence of endogenously produced DMT in the frontal cortex of rat in normal conditions to similar extents to other monoamines, which might help explain the presence of a DMT receptor in the human brain (Dean et al., 2019). Consistent with the putative role of S1R as a prosurvival and neuroprotective receptor, and with the putative role of DMT as a stress-related neurotransmitter, the levels of endogenous DMT were reported to increase after cardiac arrest in some cortical areas (Szabo and Frecska, 2016; Szabo et al., 2016; Dean et al., 2019).
S1R is a protein that can be 1) membrane-bound at the cell membrane, where it interacts with several G protein–coupled receptors (GPCRs) and voltage-gated ion channels, thereby modulating signal transduction and neurotransmission; 2) bound to the mitochondria-associated endoplasmic reticulum membrane, where it acts as a molecular chaperone and modulates Ca2+ signaling; or 3) bound to chromatin remodeling complexes at the nuclear envelope, where it modulates gene expression (Hayashi and Su, 2007; Kourrich et al., 2013; Balasuriya et al., 2014; Tsai et al., 2015; Beggiato et al., 2017; Feltmann et al., 2018; Ortíz-Rentería et al., 2018). Activation of S1R elicits antiamnesic effects in animal models (Maurice et al., 1998) and enhances synaptic stability and plasticity in vitro (Ryskamp et al., 2019). A modulation of junctional cholinergic and glutamatergic neurotransmission via heteroreceptor-mediated biased signaling in brain areas involved in fear extinction and memory reconsolidation might be involved in mediating these effects (Earley et al., 1991; Mash and Zabetian, 1992; Maurice et al., 1998; Antonini et al., 2009; Woodcock et al., 2015; Beggiato et al., 2017; Feltmann et al., 2018; Inserra, 2018). The other main interesting feature of S1R is its modulatory action over chromatin remodeling complexes, which lead to a rearrangement of gene expression (Demmerle et al., 2012; Tsai et al., 2015). Therefore, the positive outcomes on trauma-related fear-memory extinction and memory reconsolidation elicited by compounds such as ayahuasca and MDMA might result from the activation of this receptor (Feduccia and Mithoefer, 2018; Inserra, 2018; Hake et al., 2019).
S1R dysregulation is implicated in neurodegenerative and psychiatric disorders, as well as in systemic illnesses [reviewed in Tsai et al. (2009); Kourrich et al. (2012); Szabo and Frecska (2016)], suggesting that therapeutic S1R stimulation could hold promise in the treatment of those disorders (Maurice and Su, 2009; Su et al., 2016). Indeed, S1R modulation elicits prohomeostatic, neuroprotective, and immunomodulatory outcomes, prompting that this might be the case (Nguyen et al., 2015). For example, 5-MeO-DMT and DMT increase neurogenesis in vitro (Morales García et al., 2017) and in vivo (Morales-Garcia et al., 2020), and ayahuasca acutely increase circulating BDNF levels in humans (de Almeida et al., 2019). Future studies should assess the utility of these compounds in preclinical models of neuroinflammatory and neurodegenerative disorders.
IV. Psychedelic Compounds as Modulators of Neurotransmitter Systems
A. Neurotransmitter Dysfunctions in Psychiatric Disorders
Psychiatric disorders present serotonergic, norepinephrinergic, dopaminergic, glutamatergic, and GABAergic dysregulation, which is rescued, at least partially, by psychiatric drugs (Grace, 2016; Murrough et al., 2017; Savitz, 2020). Psychedelic compounds have profound modulatory outcomes on these neurotransmitter systems, which are of great relevance to psychiatry both in terms of understanding psychiatric pathophysiology and in terms of drug discovery and development opportunities (see Fig. 3).

Effects of psychedelic compounds on the serotonergic, dopaminergic, glutamatergic, and GABAergic neurotransmitter systems relevant to psychiatric disorders and comorbidities. For each compound, or group of compounds with similar pharmacology, the main known modulatory effects over the serotonergic, dopaminergic, glutamatergic, and GABAergic neurotransmitter systems in different areas of the brain are reported.
B. Serotonin
Serotonin modulates an ample spectrum of physiological and pathological behaviors (Lesch et al., 1996; Domínguez-López et al., 2012). A common feature of psychiatric disorders is a chronic 5-HT depletion state, mediated at least partially by upregulated indoleamine 2,3-dioxygenase (IDO), the rate-limiting enzyme of tryptophan catabolism (Messaoud et al., 2019; Savitz, 2020). IDO represents a key node in the neuroimmune crosstalk, which decreases 5-HT bioavailability and increases the toxic metabolic compound kynurenine, among others, exacerbating psychiatric symptoms (Messaoud et al., 2019; Savitz, 2020). Compounds that increase synaptic 5-HT availability have represented the core of psychiatric pharmacotherapy so far, and psychedelic compounds elicit profound effects over the serotonergic system, which could translate to long-term increased synaptic 5-HT availability.
1. Lysergic Acid Diethylamide
LSD is a 5-HT2A receptor partial agonist (Marek and Aghajanian, 1996; Rickli et al., 2015; De Gregorio et al., 2016a). Similarly, LSD is an agonist at the 5-HT1A receptor (Norman et al., 1985; Reissig et al., 2005; Rickli et al., 2016). Chronic administration of the widely used antidepressant and anxiolytic SSRI medications desensitizes the postsynaptic 5-HT1A receptor, increasing serotonergic firing and ultimately eliciting antidepressant effects (Blier et al., 1993; Artigas et al., 1996; Haddjeri et al., 1998). The effects of LSD are strikingly similar to the mechanism of action of SSRIs in terms of 5-HT1A receptor dynamics. In fact, acutely, LSD decreases the firing and burst activity of 5-HT/DRN neurons via 5-HT1A receptor activation, and this effect can be blocked by 5-HT1A and 5-HT2A receptor antagonism (De Gregorio et al., 2016b). On the contrary, repeated LSD administration increases firing and burst activity in the DRN via a mechanism mediated by 5-HT1A, D2, and TAAR receptors, resembling classic antidepressants (De Gregorio et al., 2016b). LSD is also an agonist at the 5-HT2B receptor and a partial agonist at the 5-HT2C receptor (Fiorella et al., 1995; Egan et al., 1998; Rickli et al., 2016; Wacker et al., 2017). Interactions at the 5-HT2C receptor are thought to be responsible for the lack of addictive properties of psychedelic compounds (Canal and Murnane, 2017) and to partially mediate their anxiolytic and antidepressant effects (Millan, 2005; Nichols and Nichols, 2008). Agonist action has been described at the 5-HT1B, 5-HT1D, 5-HT1E, 5-HT6, and 5-HT7 receptors (Hoyer, 1988; Erlander et al., 1993; Lovenberg et al., 1993; Blaho et al., 1997; Passie et al., 2008; Wacker et al., 2017). Evidence for LSD interaction at SERT remains controversial. In fact, although an in vitro study reported no interaction with SERT (Rickli et al., 2015), in vivo investigations reported decreased LSD effects in Sert null mice (Krall et al., 2008; Kyzar et al., 2016). Given that 1) suicidal patients with depression and PTSD have increased incidence of the low-expressing SERT polymorphisms (Austin et al., 2002; Oquendo et al., 2007), 2) long-term psychedelic users have increased SERT binding (Callaway et al., 1994), 3) intact SERT signaling is necessary for fear extinction (Young et al., 2017), and 4) there is lower incidence of psychological distress and suicidality among psychedelics users (Hendricks et al., 2015; Argento et al., 2017), it cannot be excluded that SERT modulation is a common pharmacological denominator mediating the therapeutic effects of psychedelics in psychiatric disorders. Aside from serotonergic interaction, the pharmacological and behavioral effects of LSD, and likely of other psychedelics, require activation of the 5-HT2A-mGluR2 complex in glutamatergic pyramidal neurons (discussed below in the Biased Signaling, Biased Phosphoproteomics, and Psychedelic Compounds section) (Moreno et al., 2011). Recently, LSD was reported to increase the social adaptation to opinions similar to one’s own through stimulation of 5-HT2A receptors, which increase neuronal activity in the mPFC in response to social feedback processing (Duerler et al., 2020).
2. 3,4-Methylenedioxymethamphetamine
Upon ingestion, MDMA is rapidly absorbed in the intestinal tract (Kalant, 2001; Green et al., 2003). Peak plasmatic concentrations are reached after about 2 hours (Kalant, 2001). MDMA is metabolized by several pathways, such as N-demethylation, O-dealkylation, conjugation, and O-sulfation, into 14 metabolites [reviewed in Green et al. (2003))]. Initially, N-demethylation gives rise to MDA, demethylenation gives rise to 3,4-dihydroxymethamphetamine and N-methyl-α-methyldopamine, and ring hydroxylation gives rise to 6-HO-MDMA (Green et al., 2003). Given that both MDMA and its main metabolite, MDA, elicit psychoactive effects, and given that both interact (although to different extents) with the 5-HT system, it is difficult to separate the effects of one from the other. Therefore, the behavioral and neurobiological effects observed should be considered in light of this duality (Green et al., 2003).
MDMA is a 5-HT2A receptor agonist, and this effect is thought to be responsible for the MDMA-induced mesolimbic DA release (Teitler et al., 1990; Orejarena et al., 2011). Acute MDMA administration induces a transient, MDA-mediated, dose-related increase in extracellular 5-HT in the mPFC, striatum, NAc, and hippocampus (Gough et al., 1991; Gudelsky and Nash, 1996; Kankaanpää et al., 1998; O’Loinsigh et al., 2001; Mechan et al., 2002), and repeated MDMA decreases 5-HT concentration in the striatum (Nash and Yamamoto, 1992). MDMA is a weak 5-HT1A receptor agonist, and MDMA administration results in a postsynaptic upregulation of this receptor in the cortex and hypothalamus that is noticeable 1 week after acute administration (Aguirre et al., 1998; Battaglia et al., 1988). Indirect 5-HT1B receptor activation is involved in MDMA-induced hyperlocomotion when the substance is administered at low doses, whereas 5-HT2A receptor activation is involved at high doses (Rempel et al., 1993; McCreary et al., 1999; Vollenweider et al., 2002). The acute MDMA-elicited 5-HT increase is disrupted by the administration of a 5-HT reuptake inhibitor, suggesting an inhibitory effect of MDMA on SERT (Gudelsky and Nash, 1996; Liechti et al., 2000; Mechan et al., 2002). Indeed, MDMA increases the available concentration of 5-HT via inhibiting SERT, reversing the direction of the membrane transporter and ultimately resulting in the accumulation of 5-HT in the synaptic cleft (Battaglia et al., 1988; Teitler et al., 1990; Koch and Galloway, 1997; Cole and Sumnall, 2003; Green et al., 2003; Verrico et al., 2007). MDMA use has been associated with decreased SERT density, and abstinence time and SERT density correlate positively, suggesting that these alterations are reversible to extents to be determined (Müller et al., 2019). The increase in available 5-HT after MDMA administration is only partially responsible for the enhanced release of DA given that the SSRI fluoxetine does not completely block the increase in 5-HT (Koch and Galloway, 1997). Preclinical studies corroborate the crucial involvement of SERT in the effects of MDMA given that MDMA has no psychostimulant effects in SERT−/− mice (Bengel et al., 1998). Remarkably, the effects of MDMA on SERT within the NAc are sufficient to elicit the prosocial effects of MDMA (Heifets et al., 2019).
The modulation of SERT by MDMA has implications in psychiatry given that SERT abundance has a dramatic impact on synaptic 5-HT and behavior. For example, genetic SERT variants and environmental exposure eliciting epigenetic modifications (Caspi et al., 2003) downregulate SERT expression, leading to increased psychiatric susceptibility (Karg et al., 2011; Lee et al., 2005). Accordingly, patients with MDD and BD, as well as suicide completers, have decreased SERT expression (Austin et al., 2002; Oquendo et al., 2007). On the other side of the spectrum, high-expressing SERT polymorphisms are associated with obsessive-compulsive disorder (Hu et al., 2006). Therefore, MDMA-induced SERT modulation could be exploited therapeutically—for example, in the treatment of ASD (Danforth et al., 2016, 2018). Future studies should assess acute and long-term MDMA-induced SERT transcriptional dynamics to clarify the effects of trauma, MDMA, and psychotherapy on SERT modulation.
MDMA has an inhibitory effect on the production of 5-HT via decreasing tryptophan hydroxylase (the rate-limiting enzyme of 5-HT biosynthesis) activity within 15 minutes from administration (Johnson et al., 1992). This effect might be mediated by the peripheral generation of metabolites given that it is not appreciable in vitro (Schmidt et al., 1987; Che et al., 1995; Colado et al., 1999). The decrease in tryptophan hydroxylase has been observed for up to 2 weeks after a single acute administration of MDMA (Schmidt and Taylor, 1987). A further effect on the 5-HT system is exerted via the inhibitory effect of MDMA on monoamine oxidase A and monoamine oxidase B, which are also involved in the extracellular monoaminergic increase after MDMA administration (Leonardi and Azmitia, 1994). Repeated MDA administration was shown to cause long-lasting depletion of cortical, hippocampal, and striatal 5-HT (Miller et al., 1997).
3. N,N-Dimethyltryptamine, 5-Methoxy-N,N-dimethyltryptamine, and Ayahuasca
The psychedelic tryptamine DMT, contained in the shrub Psychotria viridis (among other plants), is a 5-HT2A/2C receptor agonist, although its action at 5-HT2C receptors appears less relevant than its effects at 5-HT2A receptors (Smith et al., 1998; Carbonaro et al., 2015). DMT stimulates 5-HT release and inhibits its reuptake via interacting with SERT, in which it shows a high binding-to-uptake ratio, meaning that DMT interacts with different substrate and inhibitor SERT sites and that it might be actively taken up and stored by cells (Cozzi et al., 2009; Blough et al., 2014; Rickli et al., 2016). DMT is also both an inhibitor and substrate of the vesicular monoamine transporter 2 (VMAT2), which is involved in the reuptake of monoamines. Much like the effects at SERT, DMT also shows a high binding-to-uptake ratio, suggesting that VMAT2 might be another carrier involved in the uptake of DMT by cells (Cozzi et al., 2009). DMT also interacts with the 5-HT1A, 5-HT1D, 5-HT1E, 5-HT2B, 5-HT5A, 5-HT6, and 5-HT7 receptors (Deliganis et al., 1991; Smith et al., 1998; Bunzow et al., 2001; Ray, 2010). The 5-HT1A receptor antagonism potentiates the hallucinogenic effects of DMT, suggesting that 5-HT1A receptor blockade can enhance 5-HT2A receptor–mediated hallucinogenic effects (Strassman, 1996). Like other serotonergic psychedelics, DMT inhibits 5-HT firing in the DRN (Aghajanian et al., 1970).
The β-carbolines harmol, harmine, harmaline, and tetrahydroharmine are obtained from B. caapi (ayahuasca, or “vine of the souls”) and function as MAOIs to block the metabolism of DMT, rendering it orally active (Pähkla et al., 2000; Yritia et al., 2002; Gambelunghe et al., 2008; Carbonaro et al., 2015). These compounds are tricyclic indole alkaloids that resemble tryptamines (Hamill et al., 2019). The presence of an endogenous counterpart in the pineal gland and retina, 6-methoxytetrahydro-β-carbolin (pinoline), has been reported (Langer et al., 1984; Leino, 1984). Harmine and harmaline act as reversible monoamine oxidase A inhibitors, whereas tetrahydroharmine binds 5-HT2A and 5-HT2C receptors but not the presynaptic 5-HT1A receptor as previously hypothesized (Udenfriend et al., 1958; Callaway et al., 1999). This effect results in an increase in available extracellular 5-HT and might be involved in the fast-onset antidepressant effects of ayahuasca in treatment-resistant MDD (Palhano-Fontes et al., 2019; Jiménez-Garrido et al., 2020) and potentially in the long-term neuromorphologic changes observed in long-term ayahuasca users (Bouso et al., 2015).
Although data on the acute monoaminergic effects of DMT and ayahuasca in humans are scarce, preclinical studies suggest potent and region-specific modulation of neurotransmitter release. For example, a study quantifying monoaminergic changes in the rat brain found that ayahuasca increases 5-HT in the hippocampus at high doses and in the amygdala at all doses tested. Hippocampal 5-HT turnover was significantly decreased only at high doses. Amygdalar 5-HT turnover was decreased at all doses tested (de Castro-Neto et al., 2013). Similarly, a study investigating the effects of repeated ayahuasca administration on whole-brain neurotransmitter levels found an increase in whole-brain 5-HT levels in females receiving repeated ayahuasca at the highest does tested without significant changes in 5-HT turnover (Colaço et al., 2020). A study investigating potential anticancer activity of tryptamine and DMT found that these compounds inhibit IDO in vitro, potentiating antitumor activity (Tourino et al., 2013). Although this study was not psychiatry-oriented, the fact that compounds that modulate the kynurenine pathway to decrease IDO activity are useful in psychiatry to decrease IDO emphasizes the relevance of this finding for psychiatry (Savitz, 2020). Consistent with the presence of naturally occurring MAOIs in ayahuasca, neurotransmitter turnover has been reported to be decreased upon ayahuasca administration in rodents (de Castro-Neto et al., 2013). Clinical imaging and biochemical studies should aim at closing the research gap on the monoaminergic effects of DMT and ayahuasca in humans to better elucidate its mechanism of action and maximize its therapeutic applications.
DMT is a selective SERT releaser (Blough et al., 2014; Rickli et al., 2016), whereas 5-MeO-DMT inhibits synaptosomal 5-HT reuptake (Berge et al., 1983). A study observed platelet SERT upregulation in humans after repeated ayahuasca ingestion, potentially as a result of repeated MAOI ingestion and/or repeated exposure to high levels of DMT, which affect SERT pharmacodynamics (Callaway et al., 1994; Blough et al., 2014). Tentatively, SERT polymorphisms could be investigated as predictors of positive/negative outcomes and experience intensity given that low- and high-expressing SERT genotypes have different psychopharmacological profiles (Pezawas et al., 2005; Möller et al., 2019). A close relative of DMT, 5-MeO-DMT, is found in several plants (such as A. peregrine) and in certain species of toads (such as I. alvarius, which also produces bufotenine) (Glennon and Rosecrans, 1982; Shen et al., 2010). 5-MeO-DMT acts as a 5-HT2A, 5-HT2C, 5-HT1A, S1R, and TAAR1 receptor agonist (Krebs-Thomson et al., 2006; Ray, 2010; Szabo et al., 2014). 5-MeO-DMT is a weak 5-HT uptake inhibitor (Blough et al., 2014). The affinity of 5-MeO-DMT for 5-HT1A receptor is lower compared with that of DMT (McKenna et al., 1990).
4. Ketamine
Acute ketamine does not affect DRN/5-HT firing (El Iskandrani et al., 2015) and the release of 5-HT in the DRN (López-Gil et al., 2019), but it does enhance the stimulated release of 5-HT in the DRN (Tso et al., 2004). Moreover, ketamine enhances the release of 5-HT in the mPFC (López-Gil et al., 2019), strongly activating the 5-HT/PFC system through an AMPA receptor–independent mechanism (Ago et al., 2019). Supporting an enhancement of 5-HT release, 5-hydroxy-indoleacetic acid levels are increased after both acute and repeated ketamine administration (Lindefors et al., 1997). The antidepressant effects of ketamine appear to be elicited via a 5-HT–dependent mechanism given that 5-HT depletion in mice blocks its antidepressant effects (Gigliucci et al., 2013). Similarly to SSRIs, ketamine reduces 5-HT reuptake, although the exact mechanism underlying this effect have not been fully elucidated yet (Martin et al., 1982; Nishimura et al., 1998; Can et al., 2016; Yamamoto et al., 2013). Another known serotonergic interaction occurs at 5-HT1B receptors (an action mediated by AMPA receptors) (Yamanaka et al., 2014), and this effect has been suggested to be involved in ketamine’s antidepressant effects (du Jardin et al., 2018). Antagonist action has been reported at 5-HT3 receptors at higher than clinically relevant concentrations (Appadu and Lambert, 1996). The analgesic action of ketamine is thought to be mediated by 5-HT2B and 5-HT2C receptor activation, among others (Crisp et al., 1991). Increased 5-HT and NE efflux to the mPFC seems to be the essential step to produce ketamine’s antidepressant effects, and such a process is thought to be mediated by ketamine-mediated glutamate release in the mPFC, which in turn excites DRN neurons both directly, via AMPA receptors on serotonergic terminals, and indirectly, via DRN efferents (Amargós-Bosch et al., 2006; Pham et al., 2017; López-Gil et al., 2019).
5. Psilocybin
Similarly to LSD, psilocybin decreases DRN/5-HT firing (Aghajanian and Hailgler, 1975). Serotonergic and dopaminergic activity are mutually regulated. Indeed, the selective lesion of DA neurons decreases the spontaneous firing activity of DRN/5-HT neurons by 60%, whereas the selective lesion of 5-HT neurons enhances the firing activity of VTA/DA neurons by 36%, indicating an inhibitory effect of the 5-HT input on DA neurons and an excitatory effect of DA on 5-HT neurons (Guiard et al., 2008; Dremencov et al., 2009). The 5-HT–mediated regulation of striatal DA neurotransmission via serotonergic afferents is involved in modulating reward, and serotonergic antidepressants improve mood, cognition, and hedonic behavior partially via 5-HT/DA interactions (Shuto et al., 2020). Employing radioligand binding assays, psilocybin has been demonstrated to display a high affinity at human serotonin receptors 5-HT1A, 5-HT2A, and 5-HT2B (Chadeayne et al., 2020).
5-HT2A receptors located on DA neurons within the striatum and NAc are thought to be involved in the modulation of psychedelic-induced, 5-HT–mediated dopaminergic effects (Vollenweider et al., 1999b). Psilocybin is thought to exert its psychedelic effects mostly through 5-HT2A receptor agonism, occupying up to 72% of all human brain 5-HT2A receptors at clinically relevant doses (Vollenweider et al., 1998; Winter et al., 2007; Quednow et al., 2012; Kometer et al., 2013; Madsen et al., 2019). Psilocybin also interacts with the 5-HT1A receptor in vivo without producing behavioral effects and 5-HT1D, 5-HT1E, 5-HT2B, 5-HT2C, 5-HT5, 5-HT6, and 5-HT7 receptors in vitro, although it is not known whether these interactions produce a clinically relevant effect (McKenna et al., 1990; Passie et al., 2002; Winter et al., 2007). Some authors have suggested that the 5-HT1A receptor mediates the deficits in attentional performance induced by psilocybin (Carter et al., 2005). Although the modulatory action of psilocybin on mood states and emotional face recognition are mediated by the 5-HT2A receptor, the psilocybin-induced bias toward the processing of positive emotions is mediated by other receptors, possibly the 5-HT2B, 5-HT2C, or 5-HT1A receptor (Kometer et al., 2012). Also, psilocybin decreases 5-HT reuptake by inhibiting SERT (Rickli et al., 2016).
C. Dopamine
Although psychedelics are mainly associated with their action over the serotonergic system, important interactions (direct and/or indirect) exist with the dopaminergic system that are thought to mediate the psychotic-like effects of psychedelics and that might mediate at least some of the therapeutic (and also the side effects) of psychedelic compounds. Our laboratory has shown, for example, that although at low doses, LSD acts mostly on 5-HT firing activity, at doses higher than 30 µg/kg, it also decrease the DA firing activity, effects that could explain why LSD can produce psychotic symptoms at higher doses (De Gregorio et al., 2016a). Several psychiatric symptoms are associated with DA effects. For example, anhedonia, one of the main dimensions of depression, associates with dysfunction of the DA system (Belujon and Grace, 2017). Similarly, in PTSD, genetic variability of DA receptors and stress-induced perturbations of DAT play a role in the development and severity of symptoms (Dragan and Oniszczenko, 2009; Drury et al., 2009). DAT dynamics are also altered in attention deficit hyperactivity disorder (Rowe et al., 1998).
1. Ketamine
The direct involvement of DA receptors in the pharmacology of ketamine is contradictory. In fact, although earlier studies reported high affinity and partial agonistic action at D2 receptors (Breier et al., 1998; Vollenweider et al., 2000; Kapur and Seeman, 2002; Seeman and Kapur, 2003) and ketamine-induced blockade of striatal DAT (Tsukada et al., 2001), later reports failed to replicate these findings, potentially because of the high doses necessary to initiate DA interactions (Aalto et al., 2002; Kegeles et al., 2002; Can et al., 2016). Nevertheless, ketamine is thought to restore DA activity and synaptic plasticity, at least partially, via D1 activation in the NAc (Belujon and Grace, 2014). Given that patients with MDD present NAc dysfunction with impaired dopamine-mediated reward circuits, a protective effect of ketamine on the DA system might relate to its therapeutic effects in patients with treatment-resistant depression (Feder et al., 2014; Kim et al., 2019; Krystal et al., 2019; Pizzagalli et al., 2009; Phillips et al., 2019). Acute ketamine induces a 5-fold increase in DA release in the mPFC and enhances striatal DA release, whereas repeated (7 days) ketamine increases basal dopamine to almost double that of preketamine values and attenuates ketamine-induced DA release in the mPFC (Lindefors et al., 1997; Breier et al., 1998; El Iskandrani et al., 2015; Can et al., 2016). Moreover, ketamine antagonizes DA reuptake in the striatum (Tso et al., 2004). Although ketamine does not affect firing activity in the VTA (El Iskandrani et al., 2015), it restores the stress-induced dopaminergic dysfunction in the VTA (Rincón-Cortés and Grace, 2017). Nevertheless, ketamine restores the amphetamine-induced decrease of VTA activity (Belujon et al., 2016).
2. Lysergic Acid Diethylamide
LSD is a DA D1 (Schindler et al., 2012), D2 (Seeman et al., 2005), and D4 (Marona-Lewicka et al., 2009) receptor agonist. LSD modulates DA neurotransmission in a biphasic, dose-dependent fashion. Although at low doses no effects are observed in VTA/DA activity, the latter is decreased at higher doses via a multireceptorial mechanism involving D2, 5-HT1A, and TAAR1 receptors (Marona-Lewicka et al., 2005; De Gregorio et al., 2016b). Confirming a dopaminergic involvement in LSD pharmacology, in vitro studies confirmed that LSD binds the human and murine D1 and D2 receptors (Pieri et al., 1974; Watts et al., 1995; Marona-Lewicka et al., 2009; Rickli et al., 2015). Moreover, the D4 receptor is involved in the discriminative stimulus effects in rats of LSD (Marona-Lewicka et al., 2009).
3. 3,4-Methylenedioxymethamphetamine
MDMA stimulates dopaminergic neurotransmission via 1) reversing the direction of the DA transporter; 2) a 5-HT2A receptor–induced DA release, which results in an overall increase in synaptic DA availability; and 3) an inhibition of DA nigrostriatal and mesolimbocortical reuptake in a time-, dose-, and region-dependent manner (Yamamoto and Spanos, 1988; Gudelsky et al., 1994; Bankson and Cunningham, 2001; Mayerhofer et al., 2001; Verrico et al., 2007). Acute MDMA induces DA release, and repeated administration decreases DA concentration in the striatum (Nash and Yamamoto, 1992). At high doses (i.e., 10 mg/kg), the effects of MDMA on DA release are more marked in the caudate nucleus (∼400%) compared with the NAc (∼200%), whereas at lower doses (i.e., 2.5 and 5 mg/kg), this increase is almost equivalent in the two regions and is not so remarkable (∼20%) (Yamamoto and Spanos, 1988). Interaction of MDMA with the DA transporter is relatively low, and it is thought to be partially responsible for the increase in extracellular DA after MDMA administration (Hagino et al., 2011). Decreased DAT binding in the striatum has been reported in rodents 1 week after acute MDMA (Biezonski et al., 2013). In humans, a decrease in striatal DAT was observed only in recreational MDMA users who also used amphetamines (Reneman et al., 2002a). Lastly, MDMA possesses weak D1 and D2 receptor affinity (Battaglia et al., 1988), and these effects are involved in MDMA-induced hyperlocomotion (Risbrough et al., 2006), while D2 receptors are involved in the dopaminergic toxicity induced by high doses of MDMA (Granado et al., 2011).
The MDMA-induced increase in available 5-HT is partially responsible for the increased release of DA given that fluoxetine (SSRI) only partially blocks the MDMA-induced increase in extracellular DA (Koch and Galloway, 1997). These dopaminergic effects elicited by MDMA are the basis for concern around potential abuse liability and addiction. However, at clinically relevant doses (i.e., 1.5 mg/kg), the increase in DA release is minimal, suggesting that abuse concerns are not so relevant at clinical doses (Yamamoto and Spanos, 1988; Pantoni and Anagnostaras, 2019). As discussed earlier, the hepatic metabolism of MDMA to MDA and other catechols with substantial DA activity is likely involved in the potential neurotoxic effects of MDMA (Green et al., 2003).
4. N,N-Dimethyltryptamine, 5-Methoxy-N,N-dimethyltryptamine, and Ayahuasca
Acute Ayahuasca administration increases amygdalar DA levels and decreases its turnover (de Castro-Neto et al., 2013). A recent study reported that repeated ayahuasca administration increases the concentration of the DA metabolites 3,4-dihydroxyphenylacetic acid in whole-brain homogenates in rodents (Colaço et al., 2020). Earlier studies reported that DMT increases striatal (Smith, 1977) and whole-brain DA synthesis (Waldmeier and Maitre, 1977), although it lacks direct dopaminergic activity (Hungen et al., 1975). DMT was also reported to increase the levels of the extraneuronal DA metabolite 3-methoxytyramine (Waldmeier et al., 1976) acutely and after 1 month of treatment. This suggests that repeated DMT might accelerate DA turnover, potentially as a mechanism to compensate the increased DA synthesis (Waldmeier et al., 1976; Smith, 1977; Colaço et al., 2020). One study reported decreased levels of forebrain DA in response to DMT (Haubrich and Wang, 1977). Given that S1R agonists modulate DA neurotransmission, important effects of ayahuasca, DMT, and 5-MeO-DMT over the dopaminergic system are plausible, which might be involved in the amelioration of suicidality and addiction behaviors by modulating reward-related circuitries. For example, S1R ligands can interact with the DA transporter, preventing DA efflux and nullifying drug-induced reward (Sambo et al., 2017, 2018). Therefore, future studies should investigate whether these compounds could address reward circuitry dysfunction in addiction (Sambo et al., 2017).
Together, a dose-dependent involvement of the DA system is recognized in the action of psychedelics and entactogens. Given the role of the DA system in mediating reward responses and developing addiction, future studies are required to clarify whether 1) acute and repeated psychedelic administration leads to changes in homeostasis of reward-related pathways 2) and, if so, at which doses; 3) whether this represents a concrete danger for abuse; and 4) whether adjunctive therapies can pharmacologically counteract these effects without blocking the desirable effects.
D. Glutamate
Glutamate is involved in excitatory neurotransmission, synaptogenesis, synaptic plasticity, memory, mood, and cognition (Murrough et al., 2017). Stress responses are partly glutamate-dependent (Nasca et al., 2015; Moghaddam and Javitt, 2011), and enhanced glutamatergic neurotransmission during stress is beneficial for returning to homeostasis. Glutamate can act via NMDA receptors, AMPA receptors, or metabotropic receptors. Aside from its physiological role in stress responses (Sanacora et al., 2008), dysfunction of the glutamatergic ionotropic receptor NMDA can be triggered by repeated stress and is appreciable in populations with depression (Marsden, 2011). NMDA receptor modulation is part of both the mechanism of action and outcome of antidepressant therapies (Trullas and Skolnick, 1990; Layer et al., 1995; Baumann, 1996; Sanacora et al., 2008; Bennett, 2010; Autry et al., 2011).
Glutamate release plays a key role in mediating the acute effects of psychedelics, and it is likely involved in the creation of a long-lasting state of synaptic and neural plasticity in corticolimbic circuits, which might represent the substrate for clinical improvement. The glutamatergic system drives the frontal hypermetabolic state induced by psychedelics, which also correlates to therapeutic efficacy and enhanced neurotrophic activity (Aghajanian and Marek, 1999; Scruggs et al., 2003; Muschamp et al., 2004; Lambe and Aghajanian, 2006; Carhart-Harris et al., 2016c).
NMDA receptor is involved in neuroplasticity, and corticolimbic NMDA receptor hypofunctioning is thought to be involved in the pathogenesis of SCZ (Jentsch and Roth, 1999; Olney et al., 1999). AMPA receptors are also involved with stress, depression, antidepressant treatment, and neurogenesis (Alt et al., 2005, 2006a,b; Chourbaji et al., 2008; Kallarackal et al., 2013).
1. Lysergic Acid Diethylamide
LSD elevates prefrontal-limbic glutamate via stimulating postsynaptic 5-HT2A receptors on pyramidal cells in deep cortical layers (Aghajanian and Marek, 1997, 1999; Scruggs et al., 2003; Muschamp et al., 2004), and this effect is reversed by 5-HT2A receptor antagonists (Vollenweider et al., 1998), AMPA receptor antagonists (Benneyworth et al., 2007), selective antagonists of the NR2B subunit of NMDA receptors (Zhang and Marek, 2008), and positive mGluR2 allosteric modulators (Lambe and Aghajanian, 2006). Electrophysiological studies show that antagonism of the NMDA receptor subunit NR2B suppresses the glutamate release induced by LSD, suggesting an essential role for glutamate in the downstream effects of LSD (Lambe and Aghajanian, 2006). Accordingly, neurodegeneration, aging, and inflammation associate with layer 5 synaptic and dendritic dysfunction (de Brabander et al., 1998; Fogarty et al., 2015). LSD also needs a functional mGluR2 or mGluR3 given that mGluR2/3 antagonists block LSD-induced gene transcriptional changes (Moreno et al., 2013). Therefore, it seems plausible that psychedelics might have protective effects on the neurodegenerative aspect of MDD and other psychiatric disorders via their glutamatergic-enhancing effects in corticolimbic areas.
2. Psilocybin
Multimodal neuroimaging studies after psilocybin administration reported a region-specific hypermetabolic state, especially in the frontolateral and frontomedial cortices. Given that these areas signal primarily via glutamatergic projections and are rich in 5-HT2A-mGluR2 heteroreceptors, it is possible that glutamate might mediate these effects (González-Maeso et al., 2008; Carhart-Harris et al., 2016c). Interestingly, in a preclinical study, although administration of psilocybin or ketamine alone did not ameliorate depressive-like behavior, the concomitant administration of these compounds elicited antidepressant-like effects comparable to those of fluoxetine (Martin-Ruiz et al., 2001). This finding suggests that subthreshold doses of multiple psychedelic compounds might have synergistic antidepressant effects, potentially via activating pathways that converge on the modulation of corticolimbic serotonergic and glutamatergic systems (Martin-Ruiz et al., 2001).
3. Ketamine
The main effects of ketamine and its metabolite norketamine were first attributed to NMDA receptor antagonism (Lodge et al., 1982; Anis et al., 1983; MacDonald et al., 1987; Franks and Lieb, 1994; Ebert et al., 1997). Subsequent work found that mTOR activation is a step required for ketamine to elicit antidepressant effects and to enhance synaptogenesis in the PFC (Li et al., 2010). It seems likely that these effects are not mutually exclusive and that the enhancement of AMPA receptors over NMDA receptor throughput in cortical circuits, leading to increased plasticity and neurogenesis, might be responsible for the onset of antidepressant effects (Maeng et al., 2008). The antidepressant effects of ketamine are elicited via NMDA receptor antagonism, which results in an increase of AMPA receptor activation compared with NMDA receptor activation (Maeng et al., 2008; Sanacora et al., 2008; Zarate and Manji, 2008; Autry et al., 2011). Ketamine rapidly increases extracellular cortical and striatal glutamate (Stone et al., 2012; López-Gil et al., 2019), enhancing cortical excitability (Cornwell et al., 2012), and this action correlates with improvement of psychiatric symptoms (Stone et al., 2012; Abdallah et al., 2015; Lisek et al., 2017). Moreover, ketamine increases glutamate release in the DRN (López-Gil et al., 2019). The metabolism of ketamine to hydroxynorketamine may be another key necessary step to elicit antidepressant effects, which are mediated by early and sustained activation of AMPA receptor, independently from NMDA receptor (Maeng et al., 2008). The therapeutic effects of ketamine may also be driven, at least partially, by a downregulation of excitatory amino acid transporter (EAAT) 2, which delays glutamate reuptake (Lisek et al., 2017). Ketamine administration 1) potentiates firing but decreases burst activity in glutamatergic neurons in the PFC (Jackson et al., 2004), 2) decreases firing activity in PFC GABAergic interneurons (ultimately diminishing the inhibitory influence on the cortex) (Homayoun and Moghaddam, 2007), 3) enhances striatal dopamine release (Breier et al., 1998), and 4) modulates reticular thalamus (RT) activity (Troyano-Rodriguez et al., 2014). These effects resemble the schizophrenic brain and can be partly restored by antipsychotic treatment (Kargieman et al., 2007). AMPA receptor blockade prevents the behavioral effects of ketamine and the increase in firing and bursting in the locus coeruleus and VTA activity elicited by ketamine (Moghaddam et al., 1997; El Iskandrani et al., 2015). Moreover, AMPA receptor potentiation of CA3 pyramidal neurons was also observed after ketamine (El Iskandrani et al., 2015). These findings suggest that an enhancement of catecholaminergic neurotransmission via AMPA receptor–mediated neurotransmission is required for ketamine to elicit antidepressant effects (Maeng et al., 2008; Sanacora et al., 2008; El Iskandrani et al., 2015). Importantly, ketamine blocks NMDA receptor–mediated burst firing in the lateral habenula, the “antireward” center, disinhibiting monoaminergic reward centers and rapidly relieving depression (Yang et al., 2018). A novel mechanism has recently been suggested that might at least partially explain the psychotomimetic and/or the antidepressant effects of ketamine: although physiologically α2-adrenoceptors and GABAB receptors selectively inhibit AMPA and NMDA receptors, respectively, ketamine reduces the levels and activity of the neuromodulatory regulator of G protein signaling 4, leading to selectivity loss and broad glutamatergic inhibition (Lur et al., 2019).
4. 3,4-Methylenedioxymethamphetamine
Acute MDMA increases glutamate release in the anteromedial striatum and dorsal hippocampus (Nash and Yamamoto, 1992; Anneken et al., 2013), whereas repeated MDMA does not affect glutamate efflux in the striatum (Nash and Yamamoto, 1992). This effect might be mediated by the stimulation of 5-HT2A/2C receptors on non-neuronal cells given that pretreatment with the 5-HT2A/2C receptor antagonist ketanserin prevents the increase in hippocampal glutamate release (Anneken and Gudelsky, 2012). Microiontophoretic investigations found that MDMA inhibits neuronal firing in the NAc similarly to 5-HT and DA, and this effect might be due to the MDMA-induced increase in 5-HT and DA release (White et al., 1994, 1995).
Repeated administration of MDMA during adolescence leads to important neuroadaptive changes in glutamatergic-related gene expression in corticolimbic structures (Kindlundh-Hogberg et al., 2008). For example, GluR2, mGluR1, mGluR5, NR1, NR2A, NR2B, EAAT1, and EAAT2 were increased in the adult cortex, whereas GluR3, NR2A, and NR2B receptor subunits were increased in the caudate putamen after repeated adolescent MDMA (Kindlundh-Hogberg et al., 2008). Moreover, transcription levels of GluR1 were reduced in the hippocampus, whereas GluR1, GluR3, mGluR1, and mGluR3 were increased in the hypothalamus (Kindlundh-Hogberg et al., 2008). These effects might be involved in the neurotoxic effects of MDMA binge use and repeated use in individuals of young age and potentially in the MDMA-induced, 5-HT–mediated loss of parvalbumin interneurons in the hippocampal dentate gyrus (Collins et al., 2015). NMDA receptors take part in the acquisition of conditioned rewarding effects of MDMA (García-Pardo et al., 2015), whereas NMDA and AMPA receptor antagonists block these, as well as striatal DA efflux and GABAergic toxicity in the hippocampus, suggesting potential in the treatment of MDMA and methamphetamine abuse (Finnegan and Taraska, 1996; Johnson and Kotermanski, 2006; Huff et al., 2016; García-Pardo et al., 2018, 2019).
5. N,N-Dimethyltryptamine, Ayahuasca, 5-Methoxy-N,N-dimethyltryptamine
Acute ayahuasca administration (up to 800 mg/kg) did not affect glutamate levels in the hippocampus or amygdala in rats (de Castro-Neto et al., 2013). mGluR5 is downregulated by 5-MeO-DMT in brain organoids, and this receptor is involved in addiction; therefore, the lower incidence of alcohol and substance use disorder among people who ingest psychedelics multiple times during their lifetime could be mediated by the influence of this receptor (Bird et al., 2008; Stoker et al., 2012; Dakic et al., 2017). Future studies should investigate the glutamatergic effects elicited by repeated administration of ayahuasca, DMT, and 5-MeO-DMT, which might mediate the activation of antidepressant and antiaddiction corticolimbic circuits (Bouso et al., 2013; Riga et al., 2014; Osório et al., 2015; Sanches et al., 2016; Morales-Garcia et al., 2017; Inserra, 2018; da Silva et al., 2019; de Almeida et al., 2019; Palhano-Fontes et al., 2019; Zeifman et al., 2019; Murphy-Beiner and Soar, 2020).
E. GABA
GABA is the main inhibitory neurotransmitter in the adult brain, and GABA-orchestrated circuits play key roles in physiological processes such as pain modulation (Francois et al., 2017) and in the serotonergic modulation of cortical networks (Cathala et al., 2019). GABAergic dysfunction is appreciable across psychiatric disorders (Petty, 1995; Vaiva et al., 2006). For example, NMDA receptor hypofunction in corticolimbic GABAergic interneurons, leading to corticolimbic pyramidal neuron disinhibition, is a putative pathologic mechanism of schizophrenia (Olney et al., 1999; Belforte et al., 2010; Coyle, 2012). In response to stress exposure, a downregulation of GABAB receptors has been described, which is reversed in antidepressant responders (Lloyd et al., 1985; Martin et al., 1989). In depression, altered cortical GABAergic neurotransmission disrupts canonical prefrontal information processing (Choudary et al., 2005). Accordingly, patients with depression have decreased levels of cerebrospinal GABA (Kasa et al., 1982), and antidepressants re-establish cortical GABA equilibria (Sanacora et al., 2002). Although most studies investigating the neurotransmitter effects of psychedelics focused largely on the serotonergic, glutamatergic, and dopaminergic systems, the GABAergic system is also involved in the acute and potentially the long-term effects of psychedelic compounds.
1. Ketamine
The main body of work investigating the effects of psychedelics on the GABAergic system probably comes from studying the effects of ketamine on GABAergic corticolimbic interneurons (Behrens et al., 2007; Quirk et al., 2009; Belforte et al., 2010). The effects of ketamine in modulating GABAergic populations within the CSTC circuit have been described (Behrens et al., 2007; Höflich et al., 2015; Rivolta et al., 2015; Jeevakumar and Kroener, 2016), including a disruption of RT activity (Liu et al., 2016; Mahdavi et al., 2020). The antidepressant effects of ketamine require the modulation of GABAA and GABAB receptors in discrete networks of cortical PV+ GABAergic interneurons (Quirk et al., 2009; Rosa et al., 2016; Yang et al., 2016; Wang et al., 2017a; Gerhard et al., 2020) and a downregulation of GABA reuptake (Mantz et al., 1995; Flood and Krasowski, 2000). Further strengthening the case for a modulatory effect of ketamine on the GABAergic system, ketamine administration in young animals leads to alterations in adult GABA circuits (Jeevakumar and Kroener, 2016). Both corticothalamic (Dawson et al., 2013; Anderson et al., 2017) and thalamocortical (Höflich et al., 2015) neurotransmission is modulated by ketamine via NMDA receptor blockade on GABAergic neurons in the RT, which leads to a disinhibition of DA neurons and increased DA release (Liu et al., 2016). Such effects on the RT are of particular interest given that NMDA receptor antagonism in the RT disrupts T-type calcium channel–mediated bursting in GABAergic neurons, leading to decreases in burst activity (Zhang et al., 2009) that are reminiscent of SCZ (Ferrarelli and Tononi, 2011; Hunt et al., 2017). Given that the RT (Min, 2010) and, more broadly, the CSTC circuit (Herrera et al., 2016) are 1) implicated in the generation of consciousness (Ward, 2011), 2) dysfunctional in psychiatric conditions (Swerdlow and Koob, 1987; Brown et al., 2017a; Posner et al., 2017), and 3) targeted by psychedelics (Scruggs et al., 2000; Preller et al., 2019), it is plausible that a modulatory action of this circuit is at the therapeutic core of psychedelics in psychiatric disorders (discussed in the Going Beyond Receptors: Neuronal Circuits Activated by Psychedelic Drugs section).
2. Lysergic Acid Diethylamide
LSD and DOI are partial agonists at 5-HT2A receptors in cortical interneurons, and this effect is involved in the inhibition of specific cortical pyramidal networks (Marek and Aghajanian, 1996). DOI was shown to activate cortical GABAergic interneurons and to dose-dependently increase cortical GABA levels (Abi-Saab et al., 1999; Wischhof and Koch, 2012). DOI directly depolarizes interneurons (Marek and Aghajanian, 1994), and this effect might be mediated by direct presynaptic 5-HTR binding by DOI or by psychedelic-induced, 5-HT–mediated presynaptic stimulation (Willins et al., 1997; Jakab and Goldman-Rakic, 1998). Moreover, DOI upregulates PV expression in cFOS+ GABAergic interneurons in the mPFC and somatosensory cortex (Martin and Nichols, 2016).
3. 3,4-Methylenedioxymethamphetamine
MDMA-induced DA release is mediated at least partially by GABAergic interactions. For example, MDMA enhances GABA efflux in the VTA, which dampens the MDMA-mediated DA release in the NAc shell (Bankson and Yamamoto, 2004) and decreases GABA efflux in the substantia nigra (Yamamoto et al., 1995). MDMA also modulates GABA-related gene expression in a specific spatiotemporal fashion. For example, MDMA increased GABA transporter (GAT) 1 and GAT4, but not GAT2 transcription, in the PFC and midbrain (Peng and Simantov, 2003). These results suggest a pivotal effect of MDMA on GABAergic gene expression (Simantov and Peng, 2004). Accordingly, a binge-like MDMA administration decreased PV+ GABAergic interneurons in the dorsal hippocampus (Anneken et al., 2013). Interestingly, repeated exposure to MDMA decreases the number of PV+ GABA neurons in the dentate gyrus, and this effect is attenuated by pretreatment with the anti-inflammatory ketoprofen, suggesting that the cyclooxygenase-mediated pathway might be involved (Anneken et al., 2013).
4. Psilocybin
5-HT2A receptors located on GABAergic interneurons within the striatum and NAc might be involved in the modulation of psychedelic-induced, 5-HT–mediated dopaminergic effects (Vollenweider et al., 1999b). Given that psilocybin, and psychedelics in general, decrease neuronal activity in discrete brain areas, such as the default mode network (DMN) (Carhart-Harris et al., 2012, 2016c; Palhano-Fontes et al., 2015; Tagliazucchi et al., 2016), these effects could be mediated by the serotonergic action of psilocybin or psilocybin-induced 5-HT release on GABAergic interneurons in these areas, enhancing local inhibition, as is the case for DOI (Cumming-Hood et al., 1993; Marek and Aghajanian, 1996) and LSD (Marek and Aghajanian, 1996).
5. N,N-Dimethyltryptamine, 5-Methoxy-N,N-dimethyltryptamine, Ayahuasca
Ayahuasca increases GABA levels in the hippocampus and in the amygdala only at high doses in preclinical models (de Castro-Neto et al., 2013). This mechanism could be involved in the retrieval of repressed traumatic memories experienced by some individuals who ingest ayahuasca given that the GABAergic network synchronization in the hippocampus can create the prerequisites for the synaptic changes (Paulsen and Moser, 1998; Inserra, 2018) required for memory reconsolidation (Tronson and Taylor, 2007). Such effects could be also exploited for the treatment of substance abuse (Liester and Prickett, 2012; Thomas et al., 2013) given the high comorbidity rates with PTSD (Brown and Wolfe, 1994).
F. Norepinephrine
Although not often discussed in relation to psychedelics, dissociative anesthetics, and entheogens, the neuromodulatory NE system might play an important role in both the acute effects of these compounds and in the long-term therapeutic improvements observed (Aston-Jones and Cohen, 2005). NE is the main neurotransmitter of the autonomic nervous system, which regulates crucial functions such as cognition, emotions, stress responses (including fight or flight), learning, memory, behavioral flexibility, sleep/wake cycle, heart rate, digestion, respiration, and sexual arousal (Aston-Jones and Bloom, 1981; Aston-Jones et al., 1999; Berridge and Waterhouse, 2003; Sara, 2009; Kreibig, 2010; Howells et al., 2012). NE plays a fundamental role in the stress response. In situations of stress, CRF is secreted by the paraventricular nucleus of the hypothalamus, and it activates NE neurons in the locus coeruleus (LC), which in turn stimulate further CRF production, leading to HPA axis activation and, potentially, HPA dysfunction (Valentino et al., 1983; Dunn and Swiergiel, 2008). Interestingly, the female LC/NE system appears to be more sensitive to CRF overexpression (and therefore more susceptible to stress) because of the estrogen-enhanced NE release in target brain areas (Bangasser et al., 2013, 2016).
The LC is the brain area that contains the majority of NE neurons and plays an important role in determining stimuli salience and optimizing performance via modulating the activity of frontal areas such as the orbitofrontal cortex and ACC (Aston-Jones and Bloom, 1981; Aston-Jones and Cohen, 2005; Sara, 2009). NE binds adrenergic receptors, which are both autoreceptors and heteroreceptors: α and β, both of which have at least two subtypes (De Blasi, 1990; Fraser and Venter, 1990; Strosberg, 1993, 1995). The α2 receptors are modulated by chronic, unescapable stress (Taneja et al., 2011), and drugs that modulate noradrenergic receptors are used for the treatment of PTSD (Southwick et al., 1999). Dysfunction of the NE system is appreciable in stress-induced psychiatric disorders (Klimek et al., 1997; Kosten et al., 1987). Some studies found that CSF NE levels are increased in patients with depression (Wong et al., 2000), whereas others suggested that NE turnover might be increased in depression given that patients with depression have increased levels of the NE metabolite 3-methoxy,4-hydroxyphenylethylene glycol (Koslow et al., 1983). The blockage of the reuptake of the NE by the tricyclics or the serotonin-norepinephrine reuptake inhibitors has antidepressant effects (Gobbi and Blier, 2005).
1. Ketamine
The dissociative anesthetics ketamine and PCP have been shown to increase NE release and to inhibit its reuptake. Ketamine increases NE efflux in the mPFC in vivo (Kubota et al., 1999; López-Gil et al., 2019 and NE efflux in the ventral bed nucleus of the stria terminalis in vitro (Tso et al., 2004). No changes in the DRN efflux of NE have been reported (López-Gil et al., 2019). Early studies suggested that ketamine potentiates the contraction-inducing effects of NE on vascular adrenergic neurons (Nedergaard, 1973). α-Adrenergic, but not β-adrenergic, blockade attenuated most of the cardiorespiratory responses to ketamine, suggesting an involvement of α-adrenergic neurotransmission in the effects of ketamine (Traber et al., 1970, 1971). More recent studies reported that ketamine interacts both with peripheral α1- and β2-adrenoceptor binding sites, eliciting vasoconstriction and vasodilation, respectively (Bevan et al., 1997). Supporting an involvement of α2-adrenoceptors in the mechanism of action of ketamine, α2-adrenoceptor antagonism attenuates the cardiostimulatory and psychotomimetic effects of ketamine anesthesia (Doak and Duke, 1993; Tanaka and Nishikawa, 1994; Levanen et al., 1995). Further, ketamine attenuates the cardiodepressant effects of the α2-adrenoceptor agonist dexmedetomidine (Char et al., 2013).
Ketamine and PCP were shown to inhibit NE reuptake in NE neurons of the rat cortex (Taube et al., 1975; Mandela and Ordway, 2006). However, although the inhibitory activity of PCP was high (to extents similar to those of cocaine), the inhibitory activity of ketamine was much lower (Taube et al., 1975). Similarly, in the bed nucleus of the stria terminalis, ketamine decreased the rate of NE reuptake (Tso et al., 2004). Other studies reported that ketamine inhibits NE reuptake in the heart tissue (Miletich et al., 1973; Aronson and Hanno, 1978; Salt et al., 1979) and that the antagonistic effects of ketamine at NET are responsible for the arrhythmias generated by the drug at high doses (Koehntop et al., 1977). However, different timing of ketamine exposure appears to have different effects over NE reuptake. For example, acute (30 minutes) exposure of bovine adrenal medullary cells to ketamine decreases NE uptake (Hara et al., 1998), whereas exposure for longer than 3 hours increases it (Hara et al., 2000). Importantly, 24-hour incubation with ketamine increases NET mRNA (Hara et al., 2000, 2002). The exact mechanisms through which ketamine affects NET remain unknown. It has been hypothesized that the inhibitory effects of ketamine on NE reuptake might be mediated by a direct action at the desipramine binding site of NET (Hara et al., 1998) or by an indirect effect of NMDA receptors over NET (Mandela and Ordway, 2006).
2. 3,4-Methylenedioxymethamphetamine
MDMA is an α2-adrenergic receptor agonist (Battaglia et al., 1988; Bexis and Docherty, 2005, 2009; Hysek et al., 2012a, 2013). Lower affinity has been described for β-adrenoceptors (Battaglia et al., 1988). Agonism at β3-adrenergic receptor, together with agonism at α1-adrenergic receptor, is thought to be involved in hyperthermia-induced rhabdomyolysis given that concomitant antagonism at β3- and α1-adrenergic receptor attenuates hyperthermia-induced rhabdomyolysis (Sprague et al., 2004). The empathogen MDMA increases NE release (Rothman et al., 2001; Verrico et al., 2007; Mithoefer et al., 2018). Moreover, MDMA interacts with NET, inhibiting NE reuptake (Rothman et al., 2001; Verrico et al., 2007). Accordingly, the NET inhibitors reboxetine and desipramine, respectively, decrease the biological (such as plasmatic NE release and cardiostimulant action) and subjective (such as subjective drug high, psychostimulation, and emotional excitation) effects of MDMA in humans (Hysek et al., 2011) and the acute MDMA-induced goal-oriented task impairments observed in other primates (Verrico et al., 2008). Further suggesting an adrenergic involvement in the biological and cardiostimulatory effects of MDMA, the β-adrenergic antagonist propranolol, but not the α-adrenergic antagonist prazosin, suppresses the MDMA-induced glycogenolysis and consequent glucose release in the striatum (Pachmerhiwala et al., 2010), whereas the nonselective β-blocker pindolol and the α1- and β-adrenergic antagonist carvedilol attenuate the cardiostimulatory effects of MDMA (Hysek et al., 2010, 2012a,b).
A study investigating a potential causal role of genetic variants at the NET gene, which could explain biological and subjective variability in the effects of MDMA, found a weak correlation between NET polymorphisms and the cardiostimulatory effects of MDMA (Vizeli et al., 2018). Specifically, 1) individuals carrying the GG genotype of the NET polymorphism rs1861647 presented greater cardiostimulant effects compared with subjects with one or no G alleles, 2) subjects carrying a C allele of the NET polymorphism rs2242446 presented greater MDMA-induced heart rate elevations compared with the TT genotype, and 3) G allele carriers of the polymorphism rs36029 displayed decreased elevations of arterial pressure compared with the AA genotype (Vizeli et al., 2018). Lastly, NE is involved in the MDMA-induced prolongation of the latency and light-induced miosis (pupil constriction) (Hysek and Liechti, 2012).
3. Lysergic Acid Diethylamide
LSD is an α2-adrenoceptor agonist (Marona-Lewicka and Nichols, 1995). Microiontophoretically applied LSD was reported to slightly increase the firing rate of some, but not all, LC/NE neurons, whereas intravenous LSD induces a delayed partial suppression of some others (Svensson et al., 1975; Rogawski and Aghajanian, 1979). Interestingly, aside from its direct effects on firing rate, LSD increases the reactivity of LC/NE neurons to stimuli that normally do not activate these neurons during anesthesia, such as fur stroking or air puffs, and it potentiates the response to stimuli that normally excite LC/NE neurons, such as skin pinching (Aghajanian, 1980). Further, LSD was reported to decrease the NE-induced cAMP increase in vivo in the rat hypothalamus and brainstem and in vitro in the hippocampus and to attenuate the vascular and cardiac responses mediated by α- and β-adrenoceptor activation, suggesting that LSD possesses adrenergic activity via α- and β-adrenoceptor interactions (Palmer and Burks, 1971; Hungen et al., 1975; Tang and Seeman, 1980). Several earlier preclinical studies reported no effects of LSD administration over brain NE levels or turnover (Katz and Kopin, 1969; Leonard and Tonge, 1969; Peters, 1974; Stolk et al., 1974; McCall and Aghajanian, 1980), whereas others reported a slight (∼20%) decrease in nonstressed and stressed animals (Barchas and Freedman, 1963; Diaz et al., 1968; McGrath and Olverman, 1978). In humans, LSD was reported not to alter the urinary excretion of NE (Hollister and Moore, 1967). Acutely, LSD was shown to facilitate neuronal excitation in the rat facial motor nucleus, potentiating the response to microiontophoretically applied NE (McCall and Aghajanian, 1980). Moreover, NE-depleted rats were reported to have an attenuated behavioral phenotype in response to LSD, suggesting that NE projections from the LC may indeed be relevant for the behavioral effects of LSD (Geyer et al., 1985). One study investigating the effects of repeated LSD administration (14 days) reported a decrease of brainstem NE levels of 20% compared with controls (Peters, 1974). LSD was reported not to inhibit NE reuptake (Dengler et al., 1961).
4. Psilocybin
Psilocybin is an α2-adrenoceptor agonist (Marona-Lewicka and Nichols, 1995). Psilocybin decreases available NE levels by up to 25% in the rat brain in the 4 hours after administration (Stolk et al., 1974). Interestingly, NE synthesis in the brain was transiently increased after psilocybin administration, whereas normetanephrine, a product of NE turnover, was increased by over 2-fold 1 hour after psilocybin administration, suggesting that aside from affecting the levels of extrasynaptic NE, psilocybin might affect NE turnover (Stolk et al., 1974). Earlier in vitro studies reported that psilocybin increases NE reuptake (Herblin and O’Brien, 1968), whereas recent studies suggest that psilocybin binds NET, likely inhibiting NE reuptake (Rickli et al., 2016).
5. N,N-Dimethyltryptamine, 5-Methoxy-N,N-dimethyltryptamine, Ayahuasca
Similar to LSD, mescaline, and psilocin, DMT was reported to potentiate the effects of 5-HT and NE in the facial nucleus in rats (McCall and Aghajanian, 1980). One early report found that DMT slightly increases NE synthesis in the rat forebrain (Waldmeier and Maitre, 1977), whereas another found no effect (Smith, 1977). Neither report found evidence of effects on NE turnover (Smith, 1977; Waldmeier and Maitre, 1977). Ayahuasca administration increases NE levels in the amygdala but not the hippocampus of rats (de Castro-Neto et al., 2013). Moreover, the level of 4-hydroxy-3-methoxy mandelic acid, a product of NE metabolism, was decreased in the amygdala at all doses tested, and in the hippocampus, it was decreased only at the highest doses tested (de Castro-Neto et al., 2013). On the contrary, another study found no changes in the levels of the NE metabolite 3-methoxy-4-hydrohyphenylglycol after repeated ayahuasca administration (Colaço et al., 2020). In humans, ayahuasca administration was reported to increase the urinary excretion of normetanephrine (Riba et al., 2003). Given that harmaline binds adrenoceptors, it is possible that the effects observed in response to ayahuasca administration over NEergic neurotransmission might arise from such interaction (Miralles et al., 2005). Depletion of NE was shown to block and reverse the analgesic effects of 5-MeO-DMT, suggesting that 5-MeO-DMT might elicit analgesic effects via modulating the NEergic system (Archer et al., 1985; Alhaider et al., 1993). Lastly, 5-MeO-DMT has been suggested to suppress NE reuptake and to increase its turnover (Fuxe et al., 1972; Nagai et al., 2007).
G. Oxytocin
Although beyond the scope of this work, OT is a neurotransmitter hormone involved in sociability and anxiety that is dysregulated across psychiatric disorders (Panksepp, 1992; Costa et al., 2009; Cochran et al., 2013). Oxytocin is thought to mediate at least partially the empathogenic, antidepressant, and anxiolytic effects elicited by psychedelic compounds and SSRIs (Uvnäs-Moberg et al., 1999; Schindler et al., 2018) via modulating corticolimbic circuits (Lahoud and Maroun, 2013; Sobota et al., 2015; Triana-Del Rio et al., 2019). Transient OT increase and prosocial effects were reported in response to LSD (Schmid et al., 2015a; Duerler et al., 2020) and MDMA (Dumont et al., 2009; Kirkpatrick et al., 2014a,b) in healthy subjects through 5-HT1A/2A receptor stimulation (Thompson et al., 2007). Such enhancing effects over the oxytocinergic system also observed with MDMA and DOI in preclinical models (Van de Kar et al., 2001; Thompson et al., 2007) suggest that these compounds might be useful for the treatment of ASD (Dolder et al., 2016; Parker et al., 2017). In ASD, lower baseline OT levels predict greater sociability improvements after OT administration (Parker et al., 2017), and OT receptor gene variants predict the subjective effects of MDMA (Bershad et al., 2016). Therefore, given that MDMA, LSD, and DOI all increase OT levels, patients with ASD (especially those with low baseline OT levels), might benefit from the OT-enhancing and empathogenic outcomes of these compounds (Van de Kar et al., 2001; Schmid et al., 2015a). Indeed, clinical studies are investigating the utility of MDMA, ketamine, and LSD on ASD (see Supplemental Table 1; Tables 1⇑⇑–4). Moreover, given that OT augmentation in psychotherapeutic settings is beneficial for depression (MacDonald et al., 2013) and PTSD (Koch et al., 2014), future studies should investigate the utility of psychedelic-augmented psychotherapy for these conditions, as it is already the case of MDMA for treatment-resistant PTSD (Mithoefer et al., 2018). Interestingly, it was reported that MDMA leads to an OT-dependent reopening of long-term depression in the NAc, and this represents a critical mechanism for social reward learning (Nardou et al., 2019). It is thus possible that the creation of a temporary neuroplastic window in which trauma can be reprogrammed, and neural circuits reorganized accordingly, underlies the robust improvements in PTSD symptoms observed after MDMA-augmented psychotherapy (Mithoefer et al., 2018, 2019). As hypothesized for other psychedelics, this might be mediated by chromatin rearrangements, leading to changes in gene expression and ultimately behavior (Nardou et al., 2019; Inserra, 2018). Lastly, given that OT is useful in the treatment of migraine headaches (Phillips et al., 2006), the oxytocinergic properties of psychedelic compounds could be at the base of preliminary evidence suggesting that psychedelics might be useful for headache migraines and cluster headaches (Andersson et al., 2017).
V. Biased Signaling, Biased Phosphoproteomics, and Psychedelic Compounds
A. Role of Biased Signaling in Pharmacology and Psychedelic-Induced Biased Signaling
Several lines of evidence suggest that ligands can stabilize different receptor active states, leading to interaction with different cellular signaling proteins to form a range of different complexes, thus leading to the selective activation of some signaling pathways over others. This phenomenon is called “biased signaling” or “functional selectivity” (Luttrell et al., 1999; Urban et al., 2007; Reiter et al., 2012; Kenakin, 2019). GPCRs (such as serotonergic and glutamatergic metabotropic receptors, which are closely involved in the action of psychedelic compounds) can signal through different intracellular cascades and interact with other GPCRs to form homo- and heteroreceptor complexes. This leads to spatial changes in extracellular and intracellular receptor conformations, which magnify the versatility and specificity of GPCRs via 1) altering the repertoire of potential ligands and their affinity; 2) altering the recruitment of intracellular effector proteins, or β-arrestins; and ultimately 3) modifying the downstream transcriptional effects of ligand interaction (Luttrell et al., 1999; Angers et al., 2002; Lopez-Gimenez et al., 2007; Wootten et al., 2018; Dunn et al., 2019).
From a therapeutic point of view, it is important to identify molecules able to selectively activate a specific therapeutic pathway, avoiding side effects (Kenakin and Christopoulos, 2013; Kenakin, 2015). For example, opioids like morphine are commonly used to treat pain; however, prolonged treatment with these compounds leads to the development of tolerance and related opioid use disorder (Gomes et al., 2020), including respiratory depressant effects. Opioid ligands such as morphine produce their analgesic and respiratory depressant effects through activation of the μ-opioid receptor (MOR), a GPCR receptor that signals through activation of Gi/Go proteins and via arrestin signaling (Williams et al., 2013). In particular, several early studies showed that the opioid respiratory depression was mediated by the β-arrestin 2 signal (Violin et al., 2014), thus promoting the development of MOR ligands that do not recruit β-arrestin 2 to the receptor, with the assumption that this would avoid side effects that are β-arrestin–dependent. Recent in vitro studies evaluated novel and promising MOR-biased ligands and assessed their pharmacological profile versus existing opioid compounds. It was found that these molecules (including PZM21 and SR-17018) display improved safety profiles that were not due to a reduction in β-arrestin–mediated signaling but instead to low intrinsic efficacy (Hill et al., 2018; Gillis et al., 2020). However, despite these encouraging results, the potential clinical use of these compounds remains uncertain. Indeed, in vitro research showed that oliceridine, a promising MOR-biased agonist, elicits robust G protein signaling with potency and efficacy similar to that of morphine but with less β-arrestin 2 recruitment and receptor internalization (DeWire et al., 2013). Initially, the FDA declined the approval of oliceridine in 2018 because of concerns that the analgesic benefits of the drug did not exceed its risks (including respiratory depression) (Azzam et al., 2019). However, the FDA eventually approved the compound in August 2020, claiming that the safety profile of oliceridine is similar to that of other opioids (U.S. Food and Drug Administration, 2020). On the other hand, TRV027, a biased ligand of the angiotensin II type 1 receptor able to prevent the negative effects of typical angiotensin II blockers (including vasoconstriction and reduced renal perfusion), was tested in patients with acute heart failure in a randomized, double-blind, placebo-controlled dose-ranging trial. Unfortunately, the TRV027 was revealed not to improve the clinical status at 30-day follow-up compared with placebo, confirming the notion that the potential clinical application of biased ligands remains unclear (Pang et al., 2017; Sugihara and Burnett, 2017).
Although some evidence is available for psychedelic-specific receptor conformational changes, receptor internalization bias, heteroreceptor-mediated signal transduction, biased gene expression fingerprints, and psychedelic-specific phosphoproteomics changes, the full spectrum of psychedelic-induced signal transduction and signal plasticity remains to be characterized. A recent crystallography study investigated the effects of LSD binding to 5-HT2A and 5-HT2B receptors (Wacker et al., 2017). The authors described conformational receptor changes upon LSD binding (different from those induced by the nonhallucinogenic compound ergoline), leading to the “trapping” of LSD by 5-HT2A and 5-HT2B receptors in their binding pocket, thus contributing to the slow dissociation rate, long half-life, and likely the long-lasting psychoactive effects of this compound (Wacker et al., 2017). The resulting conformational changes of the 5-HT2A receptor lead to strong functional selectivity for β-arrestin signaling over Gq signaling (Wacker et al., 2013, 2017). An in vitro study conducted in mouse embryonic fibroblast derived from wild-type and β-arrestin knockout mice demonstrated that serotonin produces 5-HT2A receptor internalization via β-arrestin coupled to protein kinases 1 and 2 (Erk1/2) phosphorylation activation. On the contrary, when the selective 5-HT2A receptor agonist DOI was tested, the authors found that β-arrestin is not essential for DOI-induced 5-HT2A receptor desensitization and Erk1/2 activation (Schmid et al., 2008; Perez-Aguilar et al., 2014). Similarly, LSD was shown to have signaling bias at 5-HT2C receptors in vitro compared with canonical 5-HT signaling (Backstrom et al., 1999). In this study, the authors reported that although 5-HT, LSD, and DOI all promote phosphoinositide hydrolysis and protein kinase C membrane translocation upon binding, LSD does not promote a detectable calcium release (Backstrom et al., 1999). Moreover, LSD binding to the alternative (fully) edited 5-HT2C receptor isoform results in a decreased ability to activate phosphoinositide hydrolysis signaling compared with 5-HT (Backstrom et al., 1999). Intriguingly, a recent article reported the development of a stable HEK 293 T-cell–based bioassay to monitor β-arrestin 2 recruitment to the 5-HT2A receptor (Pottie et al., 2020). This in vitro bioassay system was used for the functional characterization of 30 phenylalkylamine psychedelics, including 12 phenethylamines (2C-X), 7 phenylisopropylamines (psychedelic amphetamines), and 11 N-benzyl derivatives, thus allowing the comprehensive assessment of the structure-activity relationship within and between the different subclasses. For details, see Pottie et al. (2020).
B. Psychedelics and Homo- and Heteroreceptor Complexes
Aside from biased signal specificity at individual receptors, signal specificity is also enhanced by the existence of homo- and heteroreceptor complexes that are formed when multiple GPCRs are in close proximity on the plasma membrane and physically interact through their transmembrane domains (Angers et al., 2002; González-Maeso et al., 2008). Similar to what happens with biased signaling, conformational changes of the individual GPCRs that form the complex alter the repertoire of potential ligands, as well as the affinity for intracellular signal transduction proteins, ultimately eliciting specific patterns of gene expression (Borroto-Escuela et al., 2017a).
A crucial receptor-receptor interaction has been described between 5-HT2A and mGluR2 receptors that 1) converges signals from glutamatergic and serotonergic neurotransmission, 2) is increased in schizophrenia and decreased by antipsychotics, and 3) is necessary to induce psychedelic effects in preclinical models (Moreno et al., 2011; Gewirtz and Marek, 2000; Moreno et al., 2016). Indeed, chronic treatment with mGlu2/3 inhibitors decreases the 5-HT2A receptor–dependent hallucinogenic effects of LSD (Moreno et al., 2013). The mechanism for the functional interaction between 5-HT2A and mGluR2 receptor signaling is not completely understood. Despite the presence in the literature of important evidence demonstrating the heterodimerization between these two receptors (González-Maeso et al., 2008; Moreno et al., 2011), this conclusion remains controversial (Delille et al., 2013). Indeed, Delille et al. reviewed the evidence for a heterodimer 5HT2A-mGluR2 complex showed by González-Maeso et al. (2008). Delille et al. (2013) note that the formation of a heterodimer complex in a recombinant system is not necessarily an evidence and justification for its occurrence in vivo. Moreover, the authors pointed out that several lines of evidence suggest that mGlu2 receptors are expressed at a presynaptic level, whereas the 5-HT2A receptor is mostly expressed postsynaptically. Furthermore, an allosteric interaction of 5-HT2A and mGlu2 receptors has not been independently replicated in any other laboratory. They conclude that until ultrastructural studies can be finalized, the physiological importance of a 5-HT2A-mGlu2 heterodimer remains questionable and controversial (Nichols, 2016; Murat et al., 2019).
The interaction of psychedelic compounds with these heteroreceptor complexes regulates cortical sensory gating and triggers psychedelic-specific patterns of gene expression, which differ from those activated by nonpsychedelic 5-HT2A receptor agonists (González-Maeso et al., 2003). Gene expression changes triggered by to the nonhallucinogenic 5-HT2A receptor agonist lisuride differ profoundly from those elicited by the hallucinogenic LSD and DOI. For instance, LSD and DOI, but not lisuride, upregulate the expression of Egr-1 and -2 in the somatosensory cortex (Davis et al., 1991; González-Maeso et al., 2003, 2007). A deeper layer of psychedelic-induced signal specificity arises from the fact that different psychedelic compounds induce compound-unique transcriptional responses. For example, LSD and psilocybin, but not DOI, upregulate solute carrier family 2, facilitated glucose transporter member 4 (Glut4) and GPCR26 (González-Maeso et al., 2003, 2007) [for an overview of these results, see Supplemental Table 3 in González-Maeso et al. (2007)]. Another heteroreceptor complex of interest, identified in the dorsal and ventral striatum and involved in psychedelic-induced signal transduction, is the 5-HT2A-D2L heteroreceptor complex, in which D2L indicates the long D2 receptor splice variant (Albizu et al., 2011; Borroto-Escuela et al., 2014).
This complex is of particular interest given that psychedelics mimic some of the positive symptoms of schizophrenia and antipsychotic drugs modulate D2 receptor function (Davis et al., 1991; Carlsson and Carlsson, 2008; Grace, 2016; Jauhar et al., 2019). When psychedelics bind the 5-HT2A receptor of a 5-HT2A-D2L complex, an allosteric facilitatory interaction takes place between 5-HT2A and D2L receptors that favors D2 receptor signaling to canonical Gi/o-mediated 5-HT2A receptor signaling (Borroto-Escuela et al., 2014). This effect is triggered by the 5-HT2A receptor binding of some psychedelic (LSD and DOI) compounds but not by others (such as 4-Bromo-3,6-dimethoxybenzocyclobuten-1-yl)methylamine hydrobromide, also called TCB-2) (Borroto-Escuela et al., 2014). Although the existence of a 5-HT2A-D3 heteroreceptor complex has not been reported, it is plausible that these two receptors interact given that a D3 receptor antagonist is sufficient to block the stereotypical psychedelic-induced head-twitch response in mice (Perreault et al., 2014). Another possibility is that a 5-HT2A-D2-D3 heteroreceptor complex exists given that both 5-HT2A-D2 and D2-D3 complexes have been described (Lukasiewicz et al., 2010; Perreault et al., 2014). Dopamine receptors also form other dopamine-dopamine receptor complexes, such as D1-D2 and D2-D5, and heterodimers with other receptors, such as D2-NMDA, D2-TAAR1, and D2-cannabinoid 1 receptor [reviewed in Guevara-Lora et al. (2016)]. Importantly, 5-HT1A-D2 heteroreceptor complexes have also been reported, and antipsychotics were shown to favor this signaling pathway (Łukasiewicz et al., 2016). Further studies should investigate whether psychedelics interact with, and signal through, these or other homo- and heteroreceptor complexes.
The existence of 5-HT1A-5-HT2A isoreceptor complexes has been described in the pyramidal cell layer of the dorsal hippocampus and in the ACC (Borroto-Escuela et al., 2017b). The spatial arrangement of these complexes is stress-responsive given that stress decreases their colocalization, potentially via enhancing internalization (Borroto-Escuela et al., 2017b). In these complexes, the binding of specific 5-HT1A receptor agonists was decreased by the steric hindrance of agonist-bound 5-HT2A receptor, whereas this effect was nullified by the 5-HT2A receptor antagonist ketanserin (Borroto-Escuela et al., 2017b). Interestingly, the 5-HT1A receptor is mainly inhibitory and signals through Gi/o, whereas the 5-HT2A receptor is excitatory and signals through Gq/11; the intracellular consequences of ligand binding to 5-HT1A-5-HT2A isoreceptor complexes remain to be fully characterized (Hamon and Blier, 2013). Given that 5-HT1A and 5-HT2A receptor unbalances increase the likelihood of developing psychiatric disorders and given that antidepressant responses rely strongly on the molecular dynamic interactions of these receptors, the 5-HT1A-5-HT2A isoreceptor complex is of enormous interest in psychiatry. This complex represents a potential driving mechanism in psychiatry disorders and a candidate therapeutic target for drug discovery (Bonson et al., 1996; Celada et al., 2004; Aghajanian, 2009; Yohn et al., 2017).
Other 5-HT isoreceptor complexes have been characterized, such as the heterodimers 5-HT2A-5-HT2C and 5-HT1A-5-HT7 and the homodimers 5-HT1A-5-HT1A and 5-HT7-5-HT7, although their involvement in psychedelic signaling remains to be elucidated (Renner et al., 2012; Felsing et al., 2018). Given that LSD displays affinity for both 5-HT1A and 5-HT7 receptors, it is possible that LSD might have affinity for the 5-HT1A-5-HT7 complex (Crider et al., 2003; Renner et al., 2012). Lastly, glutamate heteroreceptor complexes mGluR1-mGluR5 and mGluR2-mGluR4 exist in glutamatergic synapses (Pandya et al., 2016). Whether psychedelics bind to these complexes remains to be determined. Together, a deeper understanding of psychedelic-induced signal specificity, ligand sites that determine affinity, dissociation rates, and functional potency could be exploited to enhance the therapeutic efficacy (and potentially decrease psychoactivity) of psychedelic compounds in psychiatry, similar to the efforts in identifying more efficacious antipsychotic derivatives (Shah et al., 2019).
C. Psychedelic-Induced Biased Phosphoproteomics
Phosphoproteomic refers to the post-translational phosphorylation of proteins, a process that affects their function and is orchestrated by kinases and phosphatases, which modulate a wide range of cellular processes (Riley and Coon, 2016). Aside from psychedelic-biased effects on signal transduction and gene expression, psychedelic-specific phosphoproteomic changes have been reported. For example, DOI modulates the phosphoproteome in a specific fashion compared with the nonhallucinogenic 5-HT2A receptor agonist lisuride (Karaki et al., 2014). The authors of this study found that 16 sites were differentially phosphorylated by DOI, including a hyperphosphorylated intracellular residue relevant for 5-HT2A receptor desensitization and internalization (Ser280) (Karaki et al., 2014). Such biased phosphorylation dynamics may underlie the decreased 5-HT2A receptor desensitization and internalization elicited by psychedelic (as opposed to nonpsychedelic) 5-HT2A receptor agonists (Raote et al., 2013; Karaki et al., 2014). Although in that study it was not possible to unequivocally determine the molecular events leading to the specific phosphoproteomics changes observed, one potential explanation could be that the psychedelic-stabilized 5-HT2A receptor conformation might modulate protein kinase C access, leading to a psychedelic-specific phosphorylation fingerprint (Karaki et al., 2014). Interestingly, it was also reported that mGluR2 undergoes specific ligand-induced phosphorylation at Ser843 only in the presence of 5-HT2A receptor, which could represent a key molecular interaction mediating the psychedelic crosstalk between these two receptors and might be involved in psychedelic-initiated therapeutic improvements (Murat et al., 2019). Whether psychedelics affect 5-HT2A receptor–mediated mGluR2 phosphorylation remains to be ascertained. Further in vitro and in vivo studies should assess psychedelic-induced phosphoproteomics changes in a compound-, regimen-, and brain region–specific fashion while assessing whether these changes can be detected peripherally as peripheral biomarkers of psychedelic-induced brain phosphoproteomics changes.
Although our understanding of psychedelic-specific intracellular cascades is increasing, it seems plausible that the full pharmacological profile of psychedelic compounds is far more complex than previously thought, requiring the activation of 1) multiple homo- and heteroreceptors complexes, 2) biased signaling cascades, and 3) biased post-translational protein modifications leading to 4) specific gene transcription programs, ultimately eliciting beneficial effects on mood and behavior (Celada et al., 2004; Reissig et al., 2005; Savitz et al., 2009; Wesnes et al., 2016). Independent from whether psychedelics will be approved as medicines, the knowledge generated is creating a unique opportunity for the development of novel, more efficacious designer drugs with structural analogy to psychedelic compounds (McLean et al., 2006; Zanos et al., 2017; Kenakin, 2019). Such a paradigm is already being explored for the design of a ketamine- and opioid-like analgesic with reduced side effects and could be employed in designing antidepressant and anxiolytic molecules with enhanced efficacy and reduced side effects (Manglik et al., 2016; Conibear and Kelly, 2019; Harvey et al., 2019). If skillfully harnessed, the knowledge generated could drastically improve our pharmacotherapeutic toolbox in psychiatry.
VI. Going Beyond Receptors: Neuronal Circuits Activated by Psychedelic Drugs
Until now, research on the effects of psychedelics has mostly focused on the single effects of a compound on specific neurotransmitters and neuroplastic or neuroinflammatory mediators. However, the complex psychological effects triggered by psychedelics, such as increased empathy, overcoming of trauma in PTSD, and increased capacity of insight in psychotherapy, suggest that psychedelics can activate specific neuronal circuits in the brain. Recently, more emphasis has been placed on the importance of assessing the effects of psychedelics within a more comprehensive framework. This can be achieved, for example, by employing multiple imaging techniques to assess neural activity and directed and undirected functional connectivity while also appreciating the effects on neurotransmitter concentration levels (Barnett et al., 2020; Kringelbach et al., 2020). This type of approach could help illustrate a range of fast-onset psychological effects of psychedelics that are likely not due to long-term neuroplasticity or epigenetics.
Several neural circuits have been suggested that might drive the therapeutic improvements elicited by psychedelics. Although some evidence is available, more research is needed to understand how each circuit is connected to specific effects evoked by psychedelics. Rather than mutually exclusive, it is likely that a combination of the circuits so far proposed coupled with purely immunomodulatory and neurotrophic actions and subjective meaningful experiences concur to elicit the observed clinical improvements. The activation of 5-HT2A receptors and related GPCRs appear to play a major role in the behavioral effects of serotonergic hallucinogens. The effects of 5-HT2A receptor activation on layer 5 and 6 pyramidal neurons and the resulting electrical and chemical excitatory output may be responsible for mediating a range of therapeutic improvements linked to limbic circuitry, which involves connectivity between the PFC, striatum, thalamus, claustrum, amygdala, and hippocampus (Willins et al., 1997; Riba et al., 2006; Béïque et al., 2007; Anand et al., 2009; Surget et al., 2009; Carhart-Harris et al., 2016c; Peters et al., 2016; Lewis et al., 2017; Marek, 2018; Posner et al., 2014).
A. Effects on the Default Mode Network Hub Functioning, and the “Reset” Model
The DMN refers to a circuit involving several brain regions, such as the ventromedial and dorsal PFC, the PCC, and discrete regions of the hippocampal formation (Buckner et al., 2008). The DMN is mostly active in resting state and when attention is not directed toward an external stimulus. DMN activity has been associated with introspection, autobiographical memory retrieval, internal mentation, the separation of one’s self from others and the surrounding world, and “time travel” (such as remembering the past and envisioning the future) (Andrews-Hanna, 2012; Buckner et al., 2008; Østby et al., 2012; Li et al., 2014). The DMN is also thought to be involved in the construction of “ego,” or “self” (Lebedev et al., 2015). The DMN is overactive in several psychiatric disorders, including anxiety (Coutinho et al., 2016), depression (Hamilton et al., 2011), and obsessive-compulsive disorder (Beucke et al., 2014). Such overactivity is understood to shape feelings of rumination, worthlessness, and negative self-judgement (Buckner et al., 2008; Sheline et al., 2009).
Psychedelics such as psilocybin (Carhart-Harris et al., 2017), ayahuasca (Palhano-Fontes et al., 2015), and LSD (Muthukumaraswamy et al., 2013; Carhart-Harris et al., 2016c; Tagliazucchi et al., 2016; Müller et al., 2018) decrease DMN activity. For example, LSD and psilocybin decrease connectivity between the PCC and the parahippocampus, and this correlates with self-reported scores of “ego dissolution” and visual imagery (Lebedev et al., 2015; Carhart-Harris et al., 2016c; Kaelen et al., 2016). Similarly, disrupting PCC connectivity via means of electrical stimulation in men induces dream-like hallucinations and detachment from the surrounding world, effects which resemble the psychedelic state (Herbet et al., 2014). It has been suggested that a “reset” mechanism might take place in which the acute disintegration of canonical information processing within DMN substrates allows for a subsequent resumption and reintegration of normal brain function (Carhart-Harris et al., 2017). One study has investigated the long-lasting effects of psilocybin-assisted mindfulness training, observing that higher scores of self-reported “oceanic boundlessness” during psilocybin acute effects were positively correlated with postretreat decoupling of functional connectivity between the mPFC and ACC during meditation. This finding stresses the role of synaptic plasticity as a potential mechanism of long-term psychological improvements after psychedelic therapy (Smigielski et al., 2019).
B. The Cortico-Striato-Thalamo-Cortical Model
The activation of presynaptic 5-HT2A receptors in deep-layer cortical neurons is thought to mediate the psychedelic effects of LSD and other serotonergic psychedelics (Titeler et al., 1988; Vandermaelen and Aghajanian, 1983; Jakab and Goldman-Rakic, 1998; Peyron et al., 1998; Vollenweider and Kometer, 2010; Duerler et al., 2020; Puig et al., 2003). It has been suggested that this interaction might be the driver of the cortico-striato-thalamo-cortical (CSTC) feedback loop modulation observed in imaging studies (Höflich et al., 2015; Müller et al., 2017; Preller et al., 2018, 2019). Cortical layer 6 contains corticothalamic glutamatergic neurons, which excite thalamocortical relay neurons while disynaptically inhibiting them via exciting GABAergic neurons in the RT (Jones, 2001; Zhang and Jones, 2004; Riga et al., 2018). GABAergic neurons in the RT represent a main inhibitory input to the thalamus and, by means of depolarizing GABAA receptor potential–induced bursts, are involved in the generation of brain electrical oscillations and sleep spindles (Bazhenov et al., 1999; Steriade, 2003). RT dysfunction is involved in several psychiatric disorders, including SCZ, ASD, and BD (Wells et al., 2016; Ferrarelli and Tononi, 2017; Steullet et al., 2018). Neurons in the RT provide finely tuned spatiotemporal control of thalamocortical relay cells, thereby gating thalamocortical information flow (Jones, 2001; Wang et al., 2010). This pathway, which has been hypothesized to generate consciousness (Alkire et al., 2008; Min, 2010; Ward, 2011; Herrera et al., 2016), might represent one of the main neurobiological substrates generating the wide range of consciousness-altering effects of psychedelic compounds.
In other words, psychedelic compounds might “open the gate” of consciousness (Scruggs et al., 2000; Marek et al., 2001; Geyer and Vollenweider, 2008; Müller et al., 2017; Preller et al., 2019) via allowing the thalamocortical transfer of information that might otherwise be blocked by circuits of selective attention, including the RT (McAlonan et al., 2000, 2006). A potential mechanism that might mediate such effects is the presence of serotonergic projections from the DRN (Rodriguez et al., 2011) and norepinephrinergic projections from the locus coeruleus (Asanuma, 1992), which by releasing monoamines, keep RT neurons in a depolarized state, facilitating the generation of T-type calcium channel–mediated bursting (Bosch-Bouju et al., 2013). Given that LSD decreases serotonergic firing in the DRN (Aghajanian and Vandermaelen, 1982; De Gregorio et al., 2016b), it is possible that the LSD-induced decrease of serotonergic input from the DRN leads to a hyperpolarization of RT neurons that express 5-HTRs (Goitia et al., 2016), decreasing bursting activity and ultimately decreasing the inhibitory influence of the RT on thalamocortical relay cells and thereby “opening the gate.” The resulting increased thalamocortical connectivity (5-HT2A receptor–independent) is associated with decreased cortico-striato-thalamo connectivity (5-HT2A receptor–dependent) (Geyer and Vollenweider, 2008; Müller et al., 2017; Preller et al., 2019). The latter mechanism might be responsible for the decreased global thalamic activity after the administration of psychedelics (Carhart-Harris et al., 2012; Lewis et al., 2017; Preller et al., 2019), whereas the former could be responsible for the increased thalamic functional connectivity observed in human imaging studies (Tagliazucchi et al., 2016; Müller et al., 2017; Vlisides et al., 2017; Preller et al., 2019; Barnett et al., 2020).
SSRIs similarly increase thalamic functional connectivity, pointing out another common mechanism in the action of SSRIs and psychedelics (Schaefer et al., 2014). Given that psychedelics enhance thalamocortical functional connectivity, and given that thalamocortical activation is required for memory retrieval (Staudigl et al., 2012; Müller et al., 2017; Preller et al., 2018, 2019) and the formation of new memories (Sweeney-Reed et al., 2014), it cannot be excluded that modulation of this circuit by psychedelics might be involved in the therapeutic improvements observed in populations with PTSD who report an enhancement of fear extinction and memory reconsolidation after the administration of psychedelics in clinical settings.
C. The Claustrum
Other areas that deserve more systematic investigations include the claustrum (Smythies et al., 2012) and the thalamoclaustral (Vertes et al., 2006; Vertes and Hoover, 2008) and cortico-claustro-cortical circuits (Olson and Graybiel, 1980). The claustrum is an area rich in 5-HT2A receptors (Watts et al., 1994), which are internalized upon psychedelic binding (Martin and Nichols, 2016). The claustrum has extensive inhibitory functional connectivity with cortical areas related to the top-down control of cognitive function and emotions, such as the prelimbic prefrontal cortex and the anterior cingulate cortex (Jackson et al., 2018; White et al., 2018; Krimmel et al., 2019). Indeed, DOI administration induces cFOS expression in a subpopulation of claustral neurons that express 5-HT2A receptors (Martin and Nichols, 2016). Network theoretical analyses revealed that the claustrum is a primary contributor to global brain network architecture and that significant connectivity exists between the claustrum, the frontal lobe, and cingulate regions (Torgerson et al., 2015; Wang et al., 2017b; Chia et al., 2020). Therefore, the claustrum could be one of the areas where psychedelics activate neural information for altered states of consciousness and awareness, possibly via integrating and rendering coherent multiple frequencies of neural oscillations (Crick and Koch, 2005; Goll et al., 2015).
Indeed, a recent study reported that acute psilocybin significantly decreases claustral activity and connectivity with brain areas involved in emotions, memory, and attention (Barrett et al., 2020b). Specifically, psilocybin decreased the amplitude of low-frequency fluctuations and the variance of blood oxygenation level–dependent signal in the left and right claustrum. Moreover, acute psilocybin administration decreased functional connectivity of the right claustrum with the DMN, increased right-claustrum connectivity with the frontoparietal task control network (FPTC), and decreased left-claustrum connectivity with the FPTC (Barrett et al., 2020b). Interestingly, DMN integrity was associated with right-claustrum connectivity with the DMN, whereas FPTC integrity and modularity were associated with right-claustrum and left-claustrum connectivity with the FPTC (Barrett et al., 2020b).
The organization of the claustrum resembles that of the thalamus (Olson and Graybiel, 1980), suggesting that it might serve similar functions in terms of information processing and gating. Similarly to the thalamus, which is able to suppress distractors to attend to salient stimuli, the claustrum was shown to become activated in situations of cognitive conflict, which require changes in cognitive strategy, and may therefore be a brain area involved in cognitive plasticity and adaptability (Krimmel et al., 2019). Moreover, similarly to the thalamocortical circuit, which can inhibit the cortex, creating a state of unconsciousness, the claustrum might be involved in sleep via feedforward inhibition of the cortex (Jackson et al., 2018; Norimoto et al., 2020). Given that the final output between excitation and inhibition of the cortex is involved in a range of behaviors, such as reward, anxiety, and sociability (Jackson et al., 2018), and given that psychedelics affect claustral activity and connectivity (Barrett et al., 2020b), the modulation of claustral activity and connectivity might represent a valuable novel therapeutic strategy for the treatment of addiction, psychiatric disorders, and disorders characterized by sociability impairments, such as ASD.
Together, the modulatory effects of psychedelics on the DMN, CSTC, and claustral circuits seem to be closely related to experiencing 1) “mystical-type experiences” (Griffiths et al., 2011, 2018; Barrett and Griffiths, 2018), often considered among the 10 most meaningful experiences of one’s life (Schmid and Liechti, 2018); 2) “emotional breakthrough” episodes (Roseman et al., 2019), which have been suggested to act as a “reset” switch; 3) psychedelic-induced enhancement of psychological flexibility (Davis et al., 2020); 4) creative thinking (Barrett et al., 2020b; Girn et al., 2020); and 5) increased acceptance (Girn et al., 2020). These states might mediate the anxiolytic and antidepressant effects of psychedelic compounds (Carhart-Harris et al., 2017; Murphy-Beiner and Soar, 2020).
VII. Effects of Psychedelics on Sleep
Only a few studies investigated the effects of psychedelics on sleep. For example, daytime ayahuasca administration to healthy volunteers decreased the percentage and duration of rapid eye movement (REM) sleep the night of administration while decreasing the number of periods and increasing the duration of non–rapid eye movement sleep and the number of sleep cycles (Barbanoj et al., 2008). Moreover, individuals receiving ayahuasca showed a trend toward increased REM sleep onset latency and a trend toward increased total sleep time, especially in stage 2 (Barbanoj et al., 2008). Nevertheless, spectral analysis revealed that ayahuasca increased power in the high frequency range (limited to the 15–20 Hz band) mainly in stage 2 while enhancing slow-wave sleep (Barbanoj et al., 2008). These effects were not accompanied by changes in subjective measures of sleep quality (Barbanoj et al., 2008).
Similarly, LSD administered just before bedtime was shown to increase the duration of either the first or second REM periods (results consistent for each patient on different nights) up to 245% compared with controls in some patients, with a rebound effect in the rest of the REM periods of the night, which were fewer and briefer (Green, 1965; Muzio et al., 1966; Torda, 1968). Much like patients receiving ayahuasca, REM sleep onset was delayed by LSD administration (Green, 1965). In another subpopulation of patients, LSD induced arousal episodes that significantly disrupted sleep architecture and sleep quality (Muzio et al., 1966). Interestingly, the eye movements during REM sleep were slower and fewer after LSD (Muzio et al., 1966). In this study, sleep architecture was restored the following night (Muzio et al., 1966). The authors of this study reported the existence of a subjective LSD dose range within which a high probability of prolonging REM sleep exists; however, exceeding this dose results in an arousal-like effect (Muzio et al., 1966). Preclinical studies similarly observed an increase in wakefulness, a dose-dependent increase in REM sleep onset latency, a large decrease in REM sleep, and a smaller decrease in non–rapid eye movement sleep (Depoortere and Loew, 1971; Stern et al., 1972; Kay and Martin, 1978). LSD administration during sleep was reported to increase hippocampal theta activity during REM sleep in response to LSD administration during sleep (Bílková et al., 1971a,b). Aside from acute administration studies, few studies have investigated the long-term effects of psychedelics on sleep. Cats that received LSD once displayed an increase in low-voltage-frequency sleep in the days after LSD administration, suggesting that similar subacute effects might take place in humans, which might be correlated with therapeutic improvement (Hobson, 1964).
Indirect evidence that dissociative anesthetics might affect sleep is available from studies investigating the effects of ketamine and PCP on the RT. Both ketamine and PCP were shown to decrease the activity of RT GABAergic neurons, shifting the firing mode from burst to tonic, thus eliciting an arousal-like effect (Troyano-Rodriguez et al., 2014; Mahdavi et al., 2020). Future studies should similarly investigate the effects of serotonergic psychedelics over the RT. Together, given that REM sleep is involved with associative thinking, creativity, memory consolidation, fear extinction, and neuroplasticity (Landmann et al., 2015; Ravassard et al., 2015), and given that psychedelics affect sleep, it cannot be excluded that at least some of the therapeutic benefits of psychedelics might be mediated by their effects over sleep (Froese et al., 2018).
VIII. Long-Term Neurobiological and Psychological Effects of Psychedelic Compounds
Although there is a paucity of studies investigating the long-term outcomes of psychedelic use in the general population, the available evidence suggests no increase in psychopathology and a decreased incidence of psychiatric conditions in subjects that have ingested psychedelics multiple times during their lifetime (Halpern et al., 2005; Bouso et al., 2015; Hendricks et al., 2015; Johansen and Krebs, 2015; Argento et al., 2017; Ly et al., 2018). Potentially, the most remarkable effect of repeated lifetime exposure to psychedelic compounds is a decrease in suicidal ideation and suicidal behavior (Hendricks et al., 2015; Argento et al., 2017; Zeifman et al., 2019, 2020). Aside from the improvement in depression, anxiety, and compulsion domains discussed so far, the enhancement of several behavioral, social, and cognitive domains have been reported after psychedelic exposure. These include reduced psychological distress (Hendricks et al., 2015), decreased opioid dependence (Pisano et al., 2017; Argento et al., 2019; Garcia-Romeu et al., 2020), decreased criminal behavior (Hendricks et al., 2018), enhanced creative divergent thinking and problem-solving skills (Kuypers et al., 2016; Sweat et al., 2016), proenvironmental behavior via increased nature relatedness (Forstmann and Sagioglou, 2017), and a decreased incidence of extreme political views (Nour et al., 2017; Lyons and Carhart-Harris, 2018).
A. N,N-Dimethyltryptamine, 5-Methoxy-N,N-dimethyltryptamine, and Ayahuasca
The most studied psychedelic compound in terms of long-term effects arising from the repeated ingestion of a serotonergic psychedelic is probably ayahuasca. This stems from the fact that many people in the Amazon Basin, and more recently worldwide, use ayahuasca in religious ceremonies (Labate and Feeney, 2012). This phenomenon allows for the study of a population of individuals who ingest a psychedelic hundreds if not thousands of times in their lifetime and the investigation of long-term neurobiological changes that might arise from chronic use. In terms of acute psychological changes arising from acute ayahuasca administration, rapid improvements in depression scores have been reported in treatment-resistant depression (Palhano-Fontes et al., 2019) and recurrent depression (Osório et al., 2015; Sanches et al., 2016). Another study reported a statistical trend toward decreased suicidality in patients from these trials (Zeifman et al., 2019). Four weekly sessions of ayahuasca have been described to increase “acceptance capacities” scores in the nonjudging subscale of the Five Facet Mindfulness Questionnaire (Soler et al., 2018). Recently, it was reported that a 12-day ayahuasca retreat in traditional settings significantly reduces neuroticism scores in an inversely proportional fashion to the intensity and quality of the mystical experiences associated, and these changes are still appreciable at 6-month follow-up (Netzband et al., 2020). Agreeableness was also increased in this cohort, corroborating recent research on the acute prosocial effects elicited by LSD in humans (Duerler et al., 2020; Netzband et al., 2020). Interestingly, adolescents who regularly ingest ayahuasca scored lower for psychiatric symptoms such as anxiety, body dysmorphism, and attentional problems compared with controls, suggesting no psychiatric sequelae arising from regular consumption of ayahuasca during adolescence (Da Silveira et al., 2005). However, how acute or repeated ayahuasca intake during childhood and/or adolescence might affect individuals in adulthood—positively or negatively—remains to be determined. Future longitudinal or retrospective studies should aim to answer this research question.
Long-term neurobiological changes such as increased ACC and decreased PCC thickness have been observed in ayahuasca users (Bouso et al., 2015). PCC thickness was positively correlated with greater consumption (Bouso et al., 2015). These changes are consistent with the acute effects of ayahuasca on the DMN (Palhano-Fontes et al., 2015), which might induce plasticity in these brain regions. Individuals who regularly consume ayahuasca score lower in psychometric tests for depression and confusion and higher for personality trait agreeableness, openness, and life quality, and these changes are accompanied by recent and lifetime decreased alcohol use and abuse (Barbosa et al., 2016; Garcia-Romeu et al., 2019). No neuropsychological impairment or psychopathology was associated with these changes. Interestingly, lower drug abuse rates and increased spirituality were observed in ayahuasca users (Bouso et al., 2015). Another study found that long-term psychedelic users performed better in neuropsychological tasks, possibly suggesting a state of cognitive enhancement (Bouso et al., 2012). A preclinical study found that ayahuasca consumption during pregnancy and lactation results in decreased anxiety and decreased the interest for social interactions in the offspring, suggesting potential neuromodulatory effects in the fetus (de Oliveira et al., 2011). Inhalation of 5-MeO-DMT has been reported to occasion mystical experiences associated with almost immediate improvements in depression and anxiety scores with enduring positive effects (Davis et al., 2019) and decreased drug use (Garcia-Romeu et al., 2020; Uthaug et al., 2020). Lastly, a study investigating platelet SERT found an upregulation in long-term ayahuasca drinkers, suggesting that long-term psychedelic use might induce plasticity mechanisms in the serotonergic system, potentially as a result of the long-term potentiation effect of ayahuasca over serotonergic neurotransmission, 5-HT levels, and SERT (Callaway et al., 1994).
B. Lysergic Acid Diethylamide
A study investigating the long-term effects (1 and 12 months) of a single LSD administration reported improvements in mood, life attitude, and well-being/life satisfaction and no changes in negative attitude, antisocial effects, or negative behaviors resulting from LSD (Schmid and Liechti, 2018). In preclinical studies, a single high dose of LSD does not affect cortical 5-HT1A and 5-HT2A receptors, whereas repeated administration selectively downregulates the cortical 5-HT2A receptor but not the receptors 5-HT1A, 5-HT1B, β-adrenergic receptor, α-adrenergic receptor 1 and 2, D2 receptor, and SERT (Buckholtz et al., 1990; Gresch et al., 2005). These changes were reversible 4 days after the last administration (Buckholtz et al., 1990). Interestingly, patients with depression have increased 5-HT2A receptor expression in the mPFC (Meyer et al., 2003; Shelton et al., 2009), and antidepressant compounds desensitize the cortical 5-HT2A receptor (Gómez-Gil et al., 2004; Yamauchi et al., 2006) and decrease its membrane localization (Meyer et al., 2003). Such influences of LSD on 5-HT2A receptor dynamics, which might be shared by other psychedelic compounds, are likely involved in the anxiolytic and antidepressant mechanism of action of LSD (Gasser et al., 2014, 2015). In preclinical studies, one dose of LSD elicited antidepressant-like effects that were noticeable after 5 weeks in a rodent model of depression (Hibicke et al., 2020). Repeated LSD administration elicited antidepressant-like effects in a preclinical model of antidepressant response without affecting controls, suggesting that although the improvements elicited by LSD might be noticeable in disease states, they might not be so remarkable in homeostasis (Buchborn et al., 2014). Evidence of LSD-induced deleterious neurobiological changes is available from preclinical models of SCZ in which chronic administration of high doses of LSD induces changes in gene expression, such as increased D2 receptor and nuclear orphan receptor and decreased 5-HT2C receptor, resembling schizophrenic brains (Marona-Lewicka et al., 2011). No evidence of neuronal damage arising from the administration of clinically relevant doses of LSD is available.
C. 3,4-Methylenedioxymethamphetamine
MDMA potentially has a higher risk of abuse and long-term adverse neurobiological outcomes when used chronically. Binge abuse of this substance (such as the repeated self-administration of increasing doses due to acute tolerance) can lead to hyperthermia, cardiac arrhythmias, seizures, intracranial hemorrhage, hepatotoxicity, renal failure, and rhabdomyolysis (Hegadoren et al., 1999; Green et al., 2003; Hall and Henry, 2006). Long-term MDMA abuse can have neurotoxic effects on 5-HT and DA neurons and can therefore increase the likelihood of developing psychiatric disorders while decreasing the efficacy of classic psychiatric medications (Montoya et al., 2002; Thomasius et al., 2005; Durkin et al., 2008). However, at clinical doses, the toxicity arising from MDMA administration seems low (Pantoni and Anagnostaras, 2019) (also see the MDMA section of Psychedelic Compounds as Neuroplastic Agents in this review for more details). Importantly, repeated administration of MDMA during adolescence leads to extensive neuroadaptive changes in glutamatergic-related gene expression in corticolimbic structures (Kindlundh-Hogberg et al., 2008). For example, GluR2, mGluR1, mGluR5, NR1, NR2A, NR2B, EAAT1, and EAAT2 were increased in the cortex, whereas GluR3, NR2A, and NR2B receptor subunits were increased in the caudate putamen (Kindlundh-Hogberg et al., 2008). GluR1 transcripts levels were reduced in the hippocampus, whereas GluR1, GluR3, mGluR1, and mGluR3 were increased in the hypothalamus (Kindlundh-Hogberg et al., 2008). These findings suggest important neurobiological adaptation within the glutamatergic system, which might mediate the deleterious effects of MDMA when abused at a young age in recreational settings (Kindlundh-Hogberg et al., 2008). Future studies should elucidate whether a similar remodulation of gene expression takes place at clinically relevant doses (Pantoni and Anagnostaras, 2019) and, if so, whether it is possible to detect these changes peripherally in clinical trials participants as an index of potential neurotoxic effects. As suggested above, the possibility of administering a neuroprotective drug concomitantly to MDMA in clinical settings to alleviate potential neurotoxic effects should be investigated (Tourino et al., 2010).
D. Psilocybin
Psilocybin has been widely studied in terms of mystical and peak experiences, which seem to mediate and predict at least partially clinical improvements after administration (Griffiths et al., 2008). No main negative psychopathological outcomes seem to arise after the administration of psilocybin in clinical settings (Studerus et al., 2011; Davis et al., 2020). Similarly to LSD, psilocybin elicits antidepressant and state-dependent anxiolytic effects in preclinical models, possibly because of the serotonergic and noradrenergic effects, which are detectable 5 weeks after administration (Mahmoudi et al., 2018; Hibicke et al., 2020). Clinical studies corroborate preclinical findings. In fact, psilocybin induces sustained antidepressant and anxiolytic effects, which are detectable up to 6 months after administration (Grob et al., 2011; Carhart-Harris et al., 2016a, 2018a; Griffiths et al., 2016; Ross et al., 2016). Interestingly, 1) the quality of the subjective experience during dosing sessions, 2) increased functional connectivity between the ventromedial prefrontal cortex and bilateral inferior lateral parietal cortex, and 3) decreased functional connectivity between the parahippocampal formation and the PFC predict clinical improvement (Carhart-Harris et al., 2017). Patients with treatment-resistant MDD displayed increased brain-measured emotional responses after psilocybin, suggesting that neuroadaptive processes take place in response to psilocybin treatment, which might help patients reconnect with their emotions (Roseman et al., 2018). Indeed, psilocybin induces brain network and emotional plasticity which are noticeable one week after a single administration but that tend to rebound to baseline 1 month after treatment, while the global increase in functional connectivity is still appreciable after 1 month (Barrett et al., 2020a). Interestingly, psilocybin administration was also correlated with decreased authoritarian views and increased relatedness to nature (Lyons and Carhart-Harris, 2018). Concerning alcohol and substance abuse, long-term improvements have been observed after psilocybin, which are indicative of potential usefulness of this approach for the abuse of alcohol, tobacco, and other substances (Garcia-Romeu et al., 2014; Johnson et al., 2014, 2017; Bogenschutz et al., 2015; Nielson et al., 2018). Studies are underway that aim to assess whether psilocybin treatment is feasible, tolerable, and efficacious for the treatment of eating disorders (ClinicalTrials.gov identifier: NCT04052568).
E. Ketamine
As discussed throughout this review, ketamine elicits rapid, sustained, and long-lasting antidepressant effects in adolescents and adults diagnosed with treatment-refractory MDD (Zarate et al., 2006; Price et al., 2009; aan het Rot et al., 2010; Mathew et al., 2010; Murrough et al., 2013b, 2015; Ionescu et al., 2014; Lapidus et al., 2014; Wan et al., 2015; Cusin et al., 2017; Kiraly et al., 2017; Cullen et al., 2018; Evans et al., 2018a,b; Vidal et al., 2018; Nugent et al., 2019). Importantly, rapid and sustained improvements in suicidality have also been reported (Grunebaum et al., 2018; Sinyor et al., 2018; Domany et al., 2020), suggesting that ketamine might be useful in patients with suicidal ideation and behavior. The antidepressant effects of ketamine are comparable to those of electroconvulsive therapy but have faster onset (Basso et al., 2020). Importantly, greater adverse events can be encountered when ketamine is used instead of other anesthetics for electroconvulsive therapy (Yen et al., 2015; Carspecken et al., 2018). Ketamine also elicits rapid and sustained improvements in PTSD symptoms (Feder et al., 2014; Albott et al., 2018). Given that ketamine may be used in depressed woman who undergo cesarean section to decrease the likelihood of postnatal depression, potential deleterious effects of ketamine on the newborn are being assessed (ClinicalTrials.gov identifier: NCT04285684, NCT03927378, NCT03336541). Ketamine also seems promising for the treatment of alcohol and substance abuse. Decreased cue-induced craving and risk of relapse have been observed after ketamine administration (Dakwar et al., 2014, 2017, 2019, 2020). Suggesting long-term deleterious effects of ketamine abuse on sociability, decreased levels of circulating oxytocin have been described in ketamine-dependent individuals in the early phases of abstinence that were still present 2 weeks after (Huang et al., 2018). Future studies should assess whether this is true when employing clinically relevant regimens. Lastly, given that the antidepressant effects of ketamine require opioid system activation, further studies should investigate potential deleterious effects on the opioidergic system (Williams et al., 2018).
IX. Side Effects of Psychedelic Compounds
Overall, psychedelics administered in controlled clinical settings appear nonaddictive and present mild, transient side effects. There are no reports of deaths after the ingestion of medical-grade psychedelic compounds in controlled settings (Nichols, 2016; Nichols and Grob, 2018). However, there are several concerns that need to be considered in the drug development of psychedelics. First, it seems that psychedelic drugs are strongly influenced by the context and environment (setting), and negative acute or long-term effects such as dysphoric or maladaptive/dysfunctional responses can be experienced by individuals who ingest psychedelics, which might warrant the intervention of a trained therapist (Strassman, 1984; Krebs and Johansen, 2013; Nichols, 2016). The following section aims to discuss potential mild to moderate physical and psychological side effects that can be encountered after the administration of psychedelic compounds in controlled clinical settings and mild, severe, and potentially life-threatening side effects that can be encountered after the ingestion of illicit psychedelic compounds in uncontrolled settings.
A. Acute Side Effects Encountered in Clinical Trials
1. Acute Physical Side Effects Encountered in Clinical Trials
Mild to moderate acute physical side effects have been reported by individuals receiving human-grade psychedelic compounds in controlled clinical settings. The most common include 1) headaches; 2) nausea and vomiting (especially when ayahuasca or psilocybin is administered); and increased 5) blood pressure, 6) heart rate, and 7) body temperature (Vollenweider et al., 1999b; Carhart-Harris et al., 2012; Johnson et al., 2012; Murrough et al., 2013b; Feder et al., 2014; Palhano-Fontes et al., 2015, 2019; Schmid et al., 2015a; Griffiths et al., 2016; Nichols, 2016; Mithoefer et al., 2018). The cardiovascular effects elicited by psychedelic compounds are likely mediated by their serotonergic action and should not be overlooked, especially in patients with pre-existing cardiac conditions or a family history of cardiovascular disease.
2. Acute Psychological Side Effects Encountered in Clinical Trials
So far, psychedelics have been considered physiologically and psychologically safe when administered acutely in controlled clinical settings. In healthy patients, the administration of medical-grade psychedelics in controlled clinical settings induces a transient psychosis-like state, which can be accompanied by hallucinations, sense of unity and transcendence, mystical experiences, feelings of bliss and boundlessness, dissociation, derealization, revelations, and the re-experiencing of traumatic memories, which wear off as the compound is metabolized (Nichols, 2016; De Gregorio et al., 2018). Such acute psychological effects might be mediated by the activation of the serotonergic system and HPA axis, effects shared by almost all psychedelics (Vollenweider et al., 1999b; Carhart-Harris et al., 2012; Johnson et al., 2012; Murrough et al., 2013b; Feder et al., 2014; Palhano-Fontes et al., 2015, 2019; Schmid et al., 2015a; Griffiths et al., 2016; Nichols, 2016; Mithoefer et al., 2018).
Given that the safety concerns largely revolve around the psychological rather than physiological effects of psychedelics, interpersonal support should be available during clinical administration to address psychological distress that might arise. Crucially, since psychedelics can facilitate trauma access, psychological guidance to facilitate integration should be available before, during, and after administration (Nutt and Carhart-Harris, 2020). This could maximize the positive outcomes while minimizing the negative outcomes. For example, integration support might create the prerequisites for fear extinction and memory reconsolidation, thus erasing the fear memory (Feduccia and Mithoefer, 2018; Inserra, 2018; Nutt and Carhart-Harris, 2020). However, absence of guidance after trauma access may also lead to the reinstauration of trauma and significant distress, highlighting the importance of adequate follow-up.
B. Acute Side Effects Encountered in Uncontrolled Settings
Psychedelic compounds are considered psychologically and physically safe overall when administered in controlled clinical settings. However, the use of these compounds in uncontrolled clinical settings is potentially less safe because of the absence of trained practitioners and the lack of emergency psychiatric and physical treatments readily available should the need arise. Although no fatalities have been reported after the administration of medical-grade psychedelic compounds in controlled clinical settings, fatalities have been reported after the ingestion of multiple psychoactive substances, including psychedelics, or the ingestion of synthetic phenethylamines such as 4-Iodo-2, 5-dimethoxy-N-(2-methoxybenzyl) phenethylamine (25i-NBOMe) (Nichols, 2016; Nichols and Grob, 2018).
Serious psychological side effects that can be encountered in uncontrolled settings after the ingestion of LSD, ayahuasca, DMT, and psilocybin include 1) the experiencing of fear to extents that can be temporarily debilitating and can lead to suicidal or homicidal thought, ideation, or behavior; 2) dangerous and disordered behavior triggered by panic; 3) the manifestation or exacerbation of latent psychiatric conditions such as schizophrenia, bipolar disorders, and anxiety; and 4) the instauration of long-lasting perceptual disturbances (Nichols, 2016; Dos Santos et al., 2017; dos Santos et al., 2017). For example, patients with generalized anxiety disorder might undergo particularly challenging and potentially frightening experiences during the administration of the psychedelic brew ayahuasca, which might require pharmacological psychiatric treatment (dos Santos et al., 2017).
Patients diagnosed with acute LSD- and PCP-induced psychoses have been successfully treated with the antipsychotic haloperidol (D2 receptor antagonist, 5 mg, i.m.), and chlorpromazine (D1/2/3/5 and 5-HT2/6/7 receptor antagonist, 50 mg, i.m.). Haloperidol appears more efficacious in reducing anxiety, conceptual disorganization, and visual hallucinations, whereas chlorpromazine appears more efficacious for somatic and psychological tension (Giannini et al., 1984–1985). However, hallucinogen-induced psychosis is also treated with 5-HT antagonist antipsychotics like olanzapine in emergency settings (Dr. Gobbi, personal communication), even if official guidelines and controlled studies for the use of D2 receptor antagonist and/or 5-HT2 receptor antagonist to treat hallucinogen-induced psychosis are not yet available.
1. Risk of Switch to Mania in Patients with Bipolar Disorder
Another potential side effect that could be encountered after the administration of psychedelic compounds is the risk of switch to mania in patients with BD (Szmulewicz et al., 2015). At least two such case reports are available in the literature. One is of a patient with BD with a current depressive episode who switched to a manic episode after a 4-day ayahuasca ritual (Szmulewicz et al., 2015). The patient had previously experienced hypomanic episodes, with the most recent occurring 10 days prior to ritual. The man was admitted to hospital and treated with the antipsychotic risperidone and the benzodiazepine clonazepam for 1 month, after which he became asymptomatic and was discharged (Szmulewicz et al., 2015). Another case report is of a male BD type 1 treatment-refractory psychiatrist who had attempted to self-medicate with smoked DMT (up to 1 g daily), phenelzine (a MAOI inhibitor), and clonazepam and who was admitted to hospital after developing a hypomanic psychotic episode with highly disturbed and agitated behavior 2 to 3 days after interrupting his self-medication schedule (Brown et al., 2017c). These reports suggest the need to assess the likelihood that patients with BD might switch to mania if receiving psychedelic therapy and to identify the most suited antimanic pharmacological approaches to treat such patients should the switch take place.
2. Risk of Psychosis
One of the most significant potential psychological side effects after the ingestion of psychedelic compounds in uncontrolled settings is the onset of a psychotic episode, which can last for several hours or days after the drug has worn off. The most vulnerable patients to the onset of long-lasting psychoses appear to be those who have previously received a diagnosis of SCZ, BD, or personality disorder or patients more inclined to mistrustfulness, fearfulness, and susceptibility on projection as defense (Anastasopoulos and Photiades, 1962; De Gregorio et al., 2016a, 2018; Dos Santos et al., 2017). Other risk factors include mixing other psychoactive substances, such as cannabis. For example, it was reported that an individual with a history of cannabis consumption and hypomania started experiencing psychotic symptoms associated to his hypomanic condition after the inhalation of a cannabis/DMT mix (Umut et al., 2011). Psychotic episodes have also been reported in individuals affiliated with religious groups who ingest ayahuasca for ritualistic and religious purposes, although the incidence appears to overlap that of the general population (dos Santos et al., 2017).
3. Acute Severe Physical Side Effects Encountered in Uncontrolled Settings
From a physiological point of view, psychedelic compounds appear to have a considerable margin of safety. For example, even after the accidental ingestion of very high doses (up to 550 doses) of LSD, no major adverse physiological side effects have been reported in a recent case report of three individuals (Haden and Woods, 2020). On the contrary, positive mental health changes were reported in two out of three patients, such as a 20-year-long reduction of mania with psychotic features in one subject and a reduction in physical pain and morphine addiction in another (Haden and Woods, 2020).
Although rare, conspicuous physiological adverse reactions such as vasoconstriction, coronary artery spasms, and rhabdomyolysis have been reported after the consumption of high doses of psychedelic compounds in unsupervised settings. This may be attributable to the serotonergic action of psychedelics, which affects cardiovascular function and can lead to serotonin syndrome and cardiovascular complications (Berrens et al., 2010; Cogen et al., 1978; Nichols, 2016).
4. Serotonin Syndrome
Another concern surrounding the use of psychedelic compounds in psychiatry is the onset of serotonin syndrome, a potentially life-threatening condition due to excessive stress on central and peripheral postsynaptic serotonin receptors that results in autonomic and neuromuscular aberrations that can prove fatal (Martin, 1996; Parrott, 2002; Boyer and Shannon, 2005; Scotton et al., 2019). Serotonin syndrome is characterized by changes in mental status including mild agitation; hypervigilance; delirium; autonomic hyperactivity including tachycardia, hyperthermia, hypertension, shivering, and hyperactive bowels; and neuromuscular abnormalities such as hyperreflexia and muscular rigidity (Martin, 1996; Boyer and Shannon, 2005; Scotton et al., 2019). If hyperthermia is not treated, it can lead to more severe side effects, such as rhabdomyolysis, metabolic acidosis, seizures, renal failure, intravascular coagulopathy, and death (Martin, 1996; Boyer and Shannon, 2005; Scotton et al., 2019).
Several approved serotonergic psychiatric drugs, including SSRIs, have been associated with serotonin syndrome, both at clinical doses and after accidental or voluntary overdose (Graudins et al., 1998; Kinzie and Meltzer-Brody, 2005; Paruchuri et al., 2006). A major concern revolves around the ingestion of serotonergic psychedelics by patients who medicate with SSRIs, a combination which can result in serotonin syndrome due to synergistic effects of SSRIs and serotonergic psychedelics or other compounds with MAOI activity such as Peganum harmala (Syrian rue) (Bakim et al., 2012). Concerning psychedelics, dissociative anesthetics, and empathogens, serotonin syndrome has been reported after the ingestion of LSD, MDMA, 5-methoxydiisopropyltryptamine (“foxy methoxy”), dextromethorphan, and P. harmala and by the combined use of pharmaceutical amphetamines and MDMA, mostly in patients abusing one or more of these substances, and in patients receiving MAOI or SSRI/SNRI therapies (Silbergeld and Hruska, 1979; Parrott, 2002; Ener et al., 2003; Boyer and Shannon, 2005; Arora and Kannikeswaran, 2010; Bakim et al., 2012; Davies et al., 2014; Tao et al., 2017; Singh et al., 2019).
Although no cases have been reported of serotonin syndrome after the administration of medical-grade psychedelic compounds in clinical settings, this side effect should be taken into consideration. Importantly, the use of SSRIs and SNRIs needs to be carefully taken into consideration, and if required, it should be suspended under medical supervision prior to psychedelic therapy (Bakim et al., 2012). Pharmacogenomic variability might also increase the likelihood of serotonin syndrome, as it was shown for carriers of polymorphisms at the T102C site of the 5-HT2A receptor (Cooper et al., 2014). This raises the concept that pharmacogenomic screening tests could be implemented for the identification of patients who are suitable to receive psychedelic therapy and those more likely to undergo severe side effects.
5. 3,4-Methylenedioxymethamphetamine Neurotoxicity
Several studies have investigated whether MDMA is neurotoxic on neurotransmitter systems (Schmidt, 1987; Gudelsky et al., 1994; Armstrong and Noguchi, 2004; Granado et al., 2011; Shokry et al., 2019) given that this compound is also used to induce dopaminergic toxicity in mouse models (Blesa and Przedborski, 2014). Catechols derived from MDMA metabolism appear to be responsible for the neurotoxic effects of MDMA given that direct intracerebroventricular administration of MDMA does not elicit 5-HT neurotoxicity (Green et al., 2003). For example, intrastriatal administration of 2,4,5-trihydroxymethamphetamine significantly depletes both 5-HT and DA, intracortical administration decreases 5-HT, and intracerebroventricular administration moderately depletes striatal DA without affecting 5-HT levels (Johnson et al., 1992; Zhao et al., 1992). Other metabolites, such as 6-HO-MDMA, appear to be nontoxic given that intrastriatal and intracerebroventricular administration does not affect 5-HT or DA levels (Zhao et al., 1992). Further metabolites that might contribute to the neurotoxic properties of MDMA are quinone-thioethers, orto-quinones, and the glutathione conjugates 5-GSyl-α-MeDA and 2,5-bis-(glutathion-S-yl)-α-methyldopamine (2,5-bis-(glutathione-S-yl)-α-MeDA) (Monks et al., 2001; Green et al., 2003). Acute intracerebroventricular injections of 5-GSyl-α-MeDA were reported to acutely increase DA and 5-HT levels and to increase long-term DA (but not 5-HT) turnover (Miller et al., 1996). Multiple intrastriatal and intracortical administrations of this metabolite significantly deplete striatal and cortical (but not hippocampal) 5-HT (Bai et al., 1999). 2,5-Bis-(glutathion-S-yl)-α-MeDA administration was shown to reduce cortical 5-HT and hippocampal 5-HT and 5-hydroxy-indoleacetic acid, to modestly reduce striatal 5-HT concentrations, and to produce 5-HT neurotoxicity, which was limited to the terminal areas, without affecting DA concentrations or those of its metabolites (Miller et al., 1997). Similarly, intrastriatal and intracortical administration of 2,5-bis-(glutathion-S-yl)-α-MeDA significantly depletes striatal, cortical, and hippocampal 5-HT (Bai et al., 1999). However, in many toxicity studies, the administration is often repeated, and higher doses are employed (i.e., 5–20 mg/kg) compared with clinical trials (Pantoni and Anagnostaras, 2019). However, when taking into consideration clinical or recreational doses (i.e., 1–2 mg/kg), no cognitive impairments or long-term neurotoxicity is observable (Pantoni and Anagnostaras, 2019).
C. Abuse Potential and Potential Long-Term Side Effects of Psychedelic Compounds
1. Codependence and Psychosis
Several psychoses induced by psychedelics are linked to the codependence with other substances, especially cannabis and alcohol. Even if psychosis induced by multiple substances is a frequent occurrence in the psychiatric emergency departments, there are only a few studies and case reports available (Warren et al., 2013; Paterson et al., 2015; Dos Santos et al., 2017).
2. Abuse Potential
The abuse potential profile of psychedelic compounds is weak, and so is the recurrence of significant psychiatric sequelae from the administration of psychedelic compounds in controlled clinical settings (Nichols, 2016). Studies in animals require more deep investigations (see below). A rare case of physical dependence from, and tolerance to, LSD has recently been reported in the literature (Modak et al., 2019). One recent study investigated the abuse liability of low- and high-dose LSD, psilocybin, and mescaline and repeated LSD administration via the intracranial self-stimulation paradigm in rats, which is widely adopted to determine the abuse potential of a putative drug (Sakloth et al., 2019). Interestingly, the acute administration of LSD, psilocybin, and mescaline fails to produce reliable evidence of abuse (Sakloth et al., 2019). Moreover, the authors found that repeated LSD (at the highest doses tested, but not at low doses) tends to attenuate the rewarding effects of the psychostimulant methamphetamine and the depressant κ-opioid receptor agonist U69,593, although only findings for the latter were statistically significant (Sakloth et al., 2019). In another study, no preference for the self-administration of DMT or psilocybin was observed in comparison with saline (aside from one subject showing transient higher rates of DMT and psilocybin self-administration), and none self-administered DOI (Fantegrossi et al., 2004). On the other hand, in nonhuman primates, minimal rates of LSD self-injection were observed after a daily access procedure, but a more substantial self-administration was observed after an intermittent access procedure (Goodwin, 2016). Therefore, although it seems that these compounds present low abuse liability, more adequately powered studies are required to confirm these findings, especially those using different doses and different regimens of psychedelics.
3. Hallucinogen-Persisting Perception Disorder
Hallucinogen-persisting perception disorder is the reoccurrence of drug-like effects long after the substance has worn out (Orsolini et al., 2017; Halpern et al., 2018; Knuijver et al., 2018; Skryabin et al., 2018). Although the incidence of this condition is low and seldom impairs life quality significantly, it has been reported in patients with alcoholism (Batzer et al., 1999), military personnel (Stanton and Bardoni, 1972), and individuals with a long-term history of psychedelic use/abuse proportionally to the number of times the substance was ingested but sometimes after one administration (Batzer et al., 1999; Orsolini et al., 2017; Anderson et al., 2018; Halpern et al., 2018; Knuijver et al., 2018; Martinotti et al., 2018; Kurtom et al., 2019; Skryabin et al., 2018; Goldman et al., 2007). Successful treatment of hallucinogen-persisting perception disorder has been reported with high-potency serotonergic benzodiazepines such as clonazepam (Lerner et al., 2003).
D. Potential Long-Term Physical Side Effects in Uncontrolled Settings
1. Cardiovascular Side Effects
Other potential severe side effects that might arise after microdosing concern the direct repeated stimulation of cardiac 5-HT2B receptors by psychedelics, as well as the indirect 5-HT2B receptors stimulation due to increased circulating 5-HT levels (Connolly et al., 1997; Fishman, 1999). Cardiac 5-HT2B receptor overstimulation can lead to cardiovascular dysfunction after significant histologic and functional changes to the heart valves (Gustafsson et al., 2005; Elangbam et al., 2008; Hutcheson et al., 2011). For example, valvular fibrosis leading to valvular thickening and valvular regurgitation were observed in patients receiving anorectic (appetite-depressing) drugs with 5-HT2A/2B receptor action, such as the fenfluramine-phentermine combination (Connolly et al., 1997; Curfman, 1997; Hutcheson et al., 2011). Animal models of repeated 5-HT administration confirmed those findings and added that these effects are accompanied by increased cardiac 5-HT2B receptor and decreased SERT expression (Gustafsson et al., 2005; Elangbam et al., 2008; Hutcheson et al., 2011). Similarly, the ergot alkaloids ergotamine and methysergide, which are used for the treatment of migraine and cluster headaches, were found to damage valvular function to extents that may require valve replacement due to pathologic fibrosis and thickening (Graham et al., 1966; Graham, 1967; Mason et al., 1977; Redfield et al., 1992). Pulmonary hypertension is another risk that should be assessed in response to the repeated administration of serotonergic psychedelics. This severe side effect was observed after the repeated administration of fenfluramine and dexfenfluramine, a postsynaptic 5-HT releaser and presynaptic SERT inhibitor, respectively, and aminorex, which is an NE/catecholaminergic releaser (Gurtner, 1985; Fishman, 1999; Gaine et al., 2000).
Severe cardiovascular side effects (cardiopulmonary arrest) were reported in one individual after frequent ingestion of Psilocybe semilanceata mushrooms for 1 month (Borowiak et al., 1998). Although this represents an extremely rare case report, future efforts should be directed toward identifying potential cardiac effects that might result from the repeated administration of psychedelic compounds, even in microdoses. Further efforts should be devoted to identifying biological biomarkers and psychological and neuroimaging predictors of psychedelic-induced subjective side effects. Further efforts should be devoted to identifying biological biomarkers and psychological predictors of psychedelic-induced subjective effects (such as rostral ACC thickness), as well as dissociation, negative experiences, and psychoses. This could help identify patients more likely to encounter challenging experiences (such as those with high neuroticism, emotional instability, and family history of psychosis, bipolar disorders, and childhood trauma) and implement adequate strategies to maximize therapeutic improvement while minimizing potential biological and psychological short- and long-term side effects (Barrett et al., 2017; Thal et al., 2019; Lewis et al., 2020).
X. Recommendations for Future Research
Clinically administered psychedelics elicit rapid, robust, and sustained antidepressant and anxiolytic effects with no major apparent negative outcomes. Psychedelics modulate pathways involved in the improvement and remission of psychiatric disorders. Some pathways overlap those activated by currently available psychiatric drugs, whereas others are psychedelic-specific and result from biased signaling. From a psychiatric perspective, the most relevant transcriptional programs induced span neuroplasticity, neuroimmunomodulation, and serotonergic, glutamatergic, dopaminergic, and GABAergic systems modulation. Future studies should dissect transcriptional outcomes in a compound- and dosage-specific fashion. Acute and long-term positive and negative effects on neurotransmission should be investigated. Long-term neurobiological side effects need to be systematically assessed after acute and repeated administration of psychedelics. Although the profound effects of psychedelics on immune processes appear to be mostly modulatory and protective, host immunocompetency needs to be thoroughly assessed, especially should repeated administration be needed to achieve remission.
Given that microdosing is gaining momentum as a potential alternative therapeutic strategy for medical, psychiatric, and creativity-enhancing purposes, larger RCTs could investigate whether microdosing can elicit clinical improvement and/or remission of psychiatric symptoms. Whether long-term microdosing regimens would be necessary to reach remission, studies should investigate the sequelae, if any, of this posology. Future studies should aim at identifying psychological and biological predictors of positive (and negative) outcomes to screen patients who are more likely to encounter challenging experiences, such as those of young age, of high emotional excitability, and who do not feel ready (Studerus et al., 2012; Haijen et al., 2018). Pharmacogenomics (Evans and McLeod, 2003; Whirl-Carrillo et al., 2012) and pharmacoepigenomics (Peedicayil, 2019) could be applied in this endeavor to address biological variability underlying different responses to psychedelics, as previously done for other psychiatric compounds (Yamanouchi et al., 2003; Abdolmaleky et al., 2008). For example, a recent study found that carriers of the D1 receptor polymorphism rs11133767 (two T alleles) had disproportionally engaged in many more illicit drug use episodes compared with the carriers of the C allele (Vizeli and Liechti, 2019). Similar studies could identify polymorphisms that increase the likelihood of encountering challenging experiences or of abusing psychedelics.
Whether certain individuals are identified who have an increased likelihood of undergoing challenging experiences, greater psychophysical preparation should be required, as well as enhanced psychological support before, during, and after the treatment. If psychedelics will be approved as therapeutics, whether certain individuals should be excluded from accessing this type of treatment will be determined in phase III clinical trials. So far, in clinical studies, this has been the case given that schizophrenic and/or bipolar individuals may have an increased likelihood of drug-induced psychosis or drug-induced manic or depressive episodes. Another main challenge of research is to design a novel selective 5-HT2A receptor agonist with enhanced therapeutic potential and with low affinity for D2 and TAAR1 receptors that could be responsible for the propsychotic and drug-rewarding side effects.
Psychedelics might also prove to be useful for other neuroinflammatory and systemic conditions that may or may not be comorbid with psychiatric disorders. For example, preclinical efficacy has been shown in inflammatory conditions such as high-fat diet–induced obesity and diabetes (Flanagan et al., 2019b), gut inflammatory challenges (Nau et al., 2013, 2015), and asthma (Flanagan et al., 2019a). Possible utility in other psychiatric conditions, such as eating disorders, has been hypothesized and should be further explored (Foldi et al., 2020).
Ultimately, the knowledge generated by ongoing clinical studies may boost the acceptance of this approach by legislative, funding, and academic bodies, leading to the implementation of the necessary policy shifts to allow this therapy to reach the bedside. Lastly, compounds with structural analogy might represent an alternative to natural and semisynthetic psychedelics in an attempt to decouple the therapeutic from psychedelic effects of these compounds (McLean et al., 2006; Zanos et al., 2017; Harvey et al., 2019; Kenakin, 2019). For example, 2-([2-(4-cyano-2,5-dimethoxyphenyl)ethylamino]methyl)phenol, a novel and synthesized derivate of N-benzyl–substituted phenethylamine hallucinogens, displays strong binding affinity and selectivity for the 5HT2A receptor (Hansen et al., 2014; Jensen et al., 2020). In vivo studies revealed that this compound was able to induce behavioral effects (e.g., inducing head-twitch response) with less extent compared with the 5-HT2A/2c receptor agonist DOI (Fantegrossi et al., 2015), thus suggesting a different behavioral profile of this compound despite the high selectivity for the 5HT2A receptor. Moreover, 1-acetyl-LSD (ALD-52), a derivative of LSD, has been demonstrated to produce psychedelic effects in humans, as suggested by older studies (Rothlin, 1957; Malitz et al., 1960). Recently, several other 1-acyl–substituted LSD derivatives, including 1-propanoyl-LSD (1P-LSD) and 1-butanoyl-LSD (1B-LSD), have appeared as recreational drugs (Brandt et al., 2017). A recent study investigated the effects of these two compounds in mice, revealing that, although LSD acts as an agonist at 5-HT2A receptors, ALD-52, 1P-LSD, and 1B-LSD display weak affinity even though they induce head twitches (a preclinical index of psychedelic effect) in mice with relatively high potency (Halberstadt et al., 2020). In addition, LSD was detected in the plasma of rats after administration of ALD-52 and 1P-LSD, thus suggesting that these compounds are deacylated in vivo. Therefore, ALD-52, 1P-LSD, and 1B-LSD can be considered as prodrugs for LSD. For details, please see Halberstadt et al. (2020). Future studies should investigate the translational potential of these analogs and their effects in humans.
XI. Conclusion
Together, given the available evidence to date pointing toward safety, tolerability, and efficacy of psychedelic compounds as potential novel therapeutics in psychiatry, we conclude that psychedelics are challenging the current paradigm in psychiatry. If 1) larger RCTs will corroborate these preliminary findings and 2) the acute and long-term side effect profiles will be similar, or better, than those of existing psychiatric drugs, several psychedelic compounds may be soon developed in clinical trials and eventually be approved by pharmaceutical regulatory governmental authorities.
Acknowledgments
We thank Dr. Franco Recchia and Photo Grafic (www.photo-grafic.it) for help with the preparation of the figures.
Authorship Contributions
Participated in research design: Inserra, De Gregorio, Gobbi.
Wrote or contributed to the writing of the manuscript: Inserra, De Gregorio, Gobbi.
Footnotes
This work was supported by the Canadian Institute for Health Research (CIHR) [Grant # 436986] and by the Réseau Québécois sur le Suicide, les Troubles de l’Humeur et les Troubles Associés (RQSHA) [Grant # 268065]. D.D.G. is recipient of Fond Recherche Québec-Santé (FRQS) and Canadian Institutes for Health Research (CIHR) postdoctoral fellowships.
G.G. is a consultant for Diamond Therapeutic Inc. The other authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
↵
 This article has supplemental material available at pharmrev.aspetjournals.org.
This article has supplemental material available at pharmrev.aspetjournals.org.

Abbreviations
- ACC
- anterior cingulate cortex
- ACTH
- adrenocorticotropic hormone
- AD
- Alzheimer’s disease
- ADRA
- α adrenergic receptor
- ADRB
- β adrenergic receptor
- AIDS
- acquired immunodeficiency syndrome
- ALD-52
- 1-acetyl-LSD
- ALT
- alternative lengthening of telomeres
- AMPA
- α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
- AN
- anorexia nervosa
- ANIA3
- activity and neurotransmitter-induced early gene 3
- AR
- androgen receptor
- ASD
- autism spectrum disorder
- AUD
- alcohol use disorder
- BD
- bipolar disorder
- BDNF
- brain-derived neurotrophic factor
- 1B-LSD
- 1-butanoyl-LSD
- CA
- cornu ammonis
- CACNA1A
- voltage-dependent P/Q-type calcium channel subunit alpha-1A
- CAMK2
- calcium/calmodulin-dependent protein kinase 2
- CCL5
- C-C motif chemokine 5
- CEBPB
- CCAAT/enhancer-binding protein beta
- cFOS
- proto-oncogene c-Fos
- CHRM
- muscarinic acetylcholine receptor M2
- COMT
- catechol O-methyltransferase
- CORT
- cortisol
- CREB
- cAMP response element-binding protein
- CRF
- corticotropin-releasing factor
- CSTC
- cortico-striato-thalamo-cortical
- CX3CL1
- fractalkine CXCL10 C-X-C motif chemokine 10
- DA
- dopamine
- DAT
- dopamine transporter
- DHEA
- dehydroepiandrosterone
- D2L
- dopamine 2 receptor (long splice variant)
- DMN
- default mode network
- DMT
- N,N-dimethyltryptamine
- DOI
- 2,5-dimethoxy-4-iodoamphetamine
- DRN
- dorsal raphe nucleus
- EAAT
- excitatory amino acid transporter
- EEG
- electroencephalogram
- EGR
- early growth response
- EPAC1
- rap guanine nucleotide exchange factor 3
- ERK
- extracellular signal–regulated kinase
- FDA
- Food and Drug Administration
- fMRI
- functional magnetic resonance imaging
- FPTC
- frontoparietal task control network
- GAT
- GABA transporter
- GABR
- gamma-aminobutyric acid type B receptor
- GH
- growth hormone
- GLU
- glutamate
- GPCR
- G protein–coupled receptor
- 5-GSyl-α-MeDA
- 5-(glutathion-Syl)-α-methyldopamine
- 6-HO-MDMA
- 2-hydroxy-4,5(methylenedioxy)methamphetamine
- HPA
- hypothalamic-pituitary-adrenal axis
- HRH
- histamine receptor
- 5-HT
- serotonin
- I1
- imidazoline receptor 1
- ICAM
- intercellular adhesion molecule
- IDO
- indoleamine 2,3-dioxygenase
- IFN-γ
- interferon-γ
- IKB
- inhibitor of kB kinase
- IL
- interleukin
- iNOS
- inducible nitric oxide synthase
- IP3R
- inositol trisphosphate receptor
- KROX20
- early growth response protein 2
- LC
- locus coeruleus
- LSD
- lysergic acid diethylamide
- M3
- muscarinic receptor subtype 3
- M5
- muscarinic receptor subtype 5
- MAOI
- monoamine oxidase inhibitor
- MAPK1
- mitogen-activated protein kinase 1
- MCI
- mild cognitive impairment
- MCP1
- monocyte chemoattractant protein 1
- MDA
- 3,4-methylenedioxyamphetamine
- MDD
- major depressive disorder
- MDMA
- 3,4-methylenedioxymethamphetamine
- 5-MeO-DMT
- 5-methoxy-N,N-dimethyltryptamine
- mGluR
- metabotropic glutamate receptor
- MKP1
- mitogen-activated protein kinase phosphatase
- MOR
- μ-opioid receptor
- mPFC
- medial prefrontal cortex
- mTOR
- mammalian target of rapamycin
- NAc
- nucleus accumbens
- NCT
- ClinicalTrials.gov identifier
- NE
- norepinephrine
- NET
- norepinephrine transporter
- NFAT
- nuclear factor of activated T-cells
- NF-ĸB
- nuclear factor ĸB
- NGF
- nerve growth factor
- NMDA
- N-methyl-d-aspartate
- NOR1
- neuron-derived orphan receptor-1
- NR
- N-methyl d-aspartate receptor
- NTRK2
- neurotrophic receptor tyrosine kinase 2
- OCD
- obsessive-compulsive disorder
- OPRD1
- delta-type opioid receptor
- OPRK1
- kappa-type opioid receptor
- OPRM1
- mu-type opioid receptor
- OT
- oxytocin
- OUD
- opioid use disorder
- PCC
- posterior cingulate cortex
- PCP
- phencyclidine
- PFC
- prefrontal cortex
- PKA
- protein kinase A
- PKC
- protein kinase C
- PLC
- phospholipase C
- 1P-LSD
- 1-propanoyl-LSD
- PSD
- postsynaptic density
- PTSD
- post-traumatic stress disorder
- PV
- parvalbumin
- RCT
- randomized controlled trial
- REM
- rapid eye movement
- RT
- reticular thalamus
- SCZ
- schizophrenia
- SERT
- serotonin transporter
- SLC6A13
- sodium- and chloride-dependent GABA transporter 2
- SLC6A2
- sodium-dependent noradrenaline transporter
- SNRI
- serotonin-norepinephrine reuptake inhibitor
- S1R
- σ1 receptor
- S2R
- σ2 receptor
- SGK
- serine/threonine-protein kinase Sgk1
- SSRI
- selective serotonin reuptake inhibitor
- TAAR
- trace amine-associated receptor
- Th
- T-helper
- TLR
- toll-like receptor
- TNF-α
- tumor necrosis factor-α
- TRD
- treatment resistant depression
- TSH
- thyroid stimulating hormone
- TrkB
- tyrosine receptor kinase B
- VCAM
- vascular cell adhesion protein
- VMAT2
- vesicular monoamine transporter 2
- VTA
- ventral tegmental area
- 5-HTR
- serotonin receptor
- 5-HTTLPR
- serotonin transporter-linked polymorphic region
- Copyright © 2020 by The Author(s)
This is an open access article distributed under the CC BY-NC Attribution 4.0 International license.
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
In this issue





{kind=link}
{kind=link}
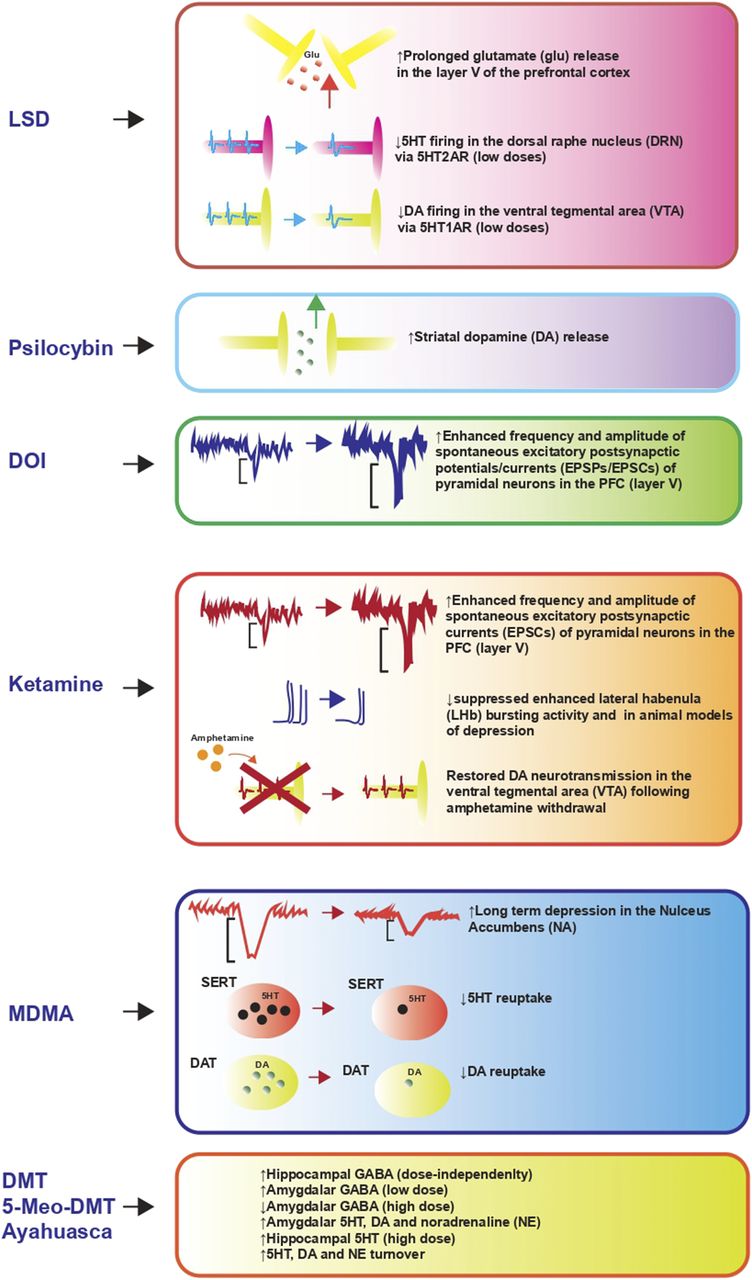{kind=link}
Jump to section
- Article
- Abstract
- I. Introduction
- II. Psychedelic Compounds as Neuroplastic Agents
- III. Psychedelic Compounds as Immunomodulatory and Anti-Inflammatory Agents
- IV. Psychedelic Compounds as Modulators of Neurotransmitter Systems
- V. Biased Signaling, Biased Phosphoproteomics, and Psychedelic Compounds
- VI. Going Beyond Receptors: Neuronal Circuits Activated by Psychedelic Drugs
- VII. Effects of Psychedelics on Sleep
- VIII. Long-Term Neurobiological and Psychological Effects of Psychedelic Compounds
- IX. Side Effects of Psychedelic Compounds
- X. Recommendations for Future Research
- XI. Conclusion
- Acknowledgments
- Authorship Contributions
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters