


Article Figures & Data
Figures
- Fig. 1
Overview of commonly used organotypic and microphysiological human liver models. (A) In the sandwich culture paradigm, conventional hepatocyte monolayers are overlaid with an additional layer of ECM. (B) Extravidin peroxidase staining shows canalicular expression of BSEP, BCRP, and MRP2 after 5 days of sandwich culture. BCRP and BSEP expression remained relatively stable over 8 days in different donors. (C) In spheroid systems using ultra–low attachment surfaces, hepatocytes (purple) with or without nonparenchymal liver cells (blue) aggregate to form spheroidal aggregates. (D) PHH spheroids retain the proteomic signature of the human liver, whereas cells from the same donors in 2D culture rapidly deteriorate. Furthermore, spheroids exhibit a stable metabolic configuration as shown by untargeted Orbitrap high-resolution mass spectrometry. (E) Schematic depiction of a chip design for microphysiological liver models in which hepatocytes (purple) and nonparenchymal liver cells (blue) are cocultured on a membrane that separates two adjacent flow channels. Note that these systems are methodologically diverse, and only one exemplary layout is shown for clarity. (F) Relative expression of markers related to liver function (left panel) and metabolic activity (right panel) remained mostly stable over 28 days in a microphysiological liver-on-a-chip. (G) In MPCCs, liver cells (purple) are spatially constrained in collagen-coated islands surrounded by stabilizing mouse fibroblasts (green). (H) In MPCCs using only hepatocytes, albumin production, urea synthesis, and CYP3A4, expression remains stable or increases during 2 weeks of culture. Coculture with human stellate cells (MPCC-HSC) causes a decrease in functional parameters. Figure modified with permission from (Bell et al., 2016; Lauschke et al., 2016a; Li et al., 2009; Maschmeyer et al., 2015; Davidson et al., 2017).
- Fig. 2
Organotypic liver models can faithfully predict human drug hepatotoxicity. (A) Representative image of a screen of 300 drugs and chemicals in which hepatotoxicity is assessed using high content imaging of markers for ROS, mitochondrial membrane potential (MMP) and intracellular glutathione (GSH) levels. Cells were treated for 24 hours at 100-fold therapeutic cmax. Nimesulide caused a significant increase in ROS, whereas telithromycin, nefazodone, and perhexiline caused a decrease in MMP and GSH. (B) In MPCCs based on hepatocyte-like cells derived from induced pluripotent stem cells, treatment with the four hepatotoxins acetaminophen, diclofenac, tolcapone, and troglitazone dose-dependently reduced albumin and urea secretion after 8 days repeated exposure, indicative of hepatocyte toxicity. In contrast, no decrease was detected for the nontoxic compounds aspirin, dextromethorphan, propranolol, and rosiglitazone. (C) Summary result of the hepatotoxicity test of 123 drugs (70 hepatotoxic, 53 nonhepatotoxic) in primary human hepatocyte spheroids. The model detected dose-dependent toxicity and, at 20-fold cmax concentration, successfully detected 69% of all hepatotoxic compounds without any false positive hits. Error bars indicate S.D. **, ***, and **** indicate P < 0.01, P < 0.001, and P < 0.0001, respectively. (D) Hepatotoxicity of acetaminophen and methotrexate (MTX) was evaluated using a perfused liver chip system. Increased ROS levels (magenta) were detected at acetaminophen concentrations of 10 mM, whereas MTX-induced toxicity manifested as intracellular lipid droplets (yellow) and αSMA (green). Figure modified with permission from (Jang et al., 2019a; Berger et al., 2015; Vorrink et al., 2018; Xu et al., 2008).
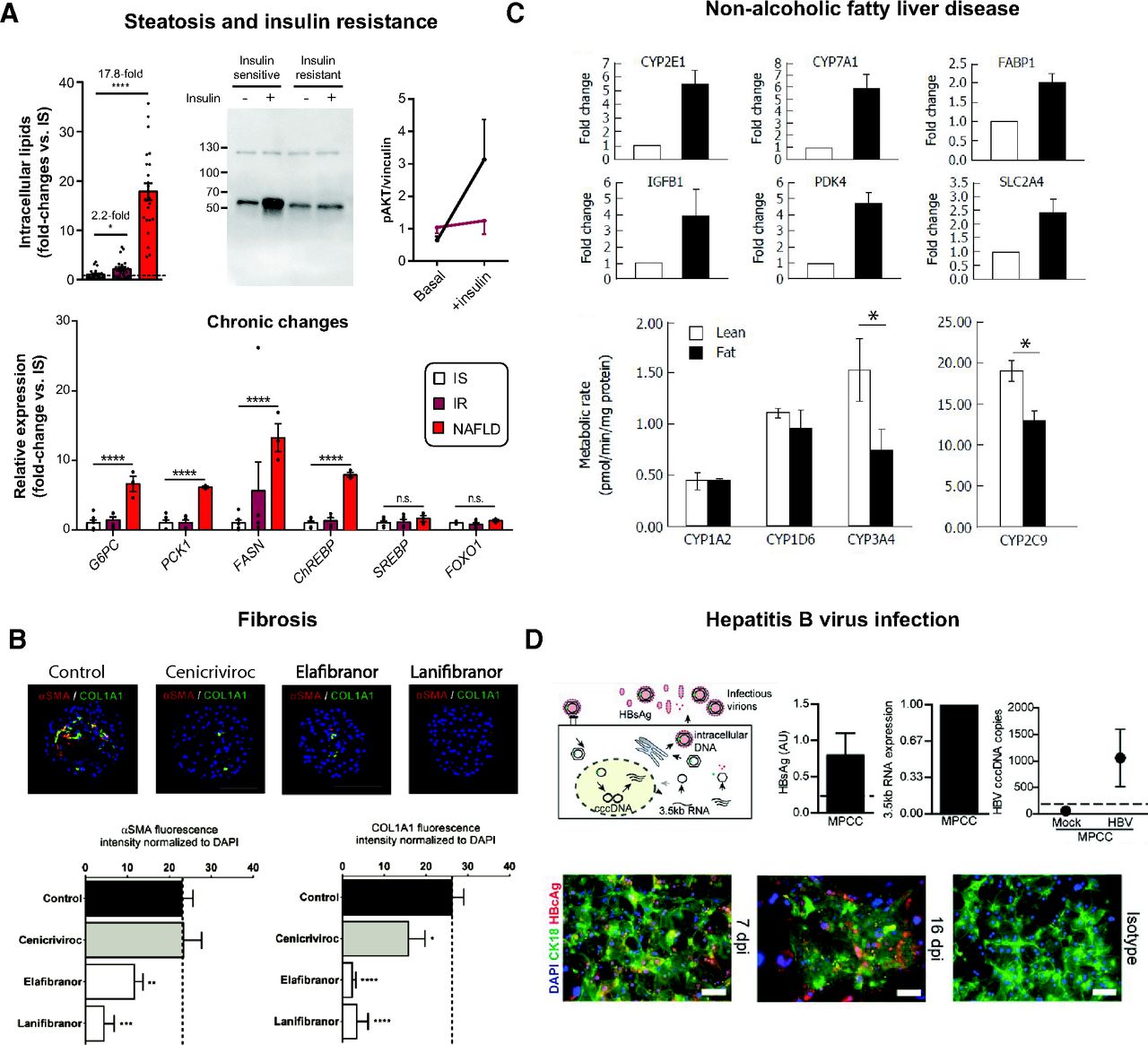
- Fig. 3
Organotypic culture systems for liver disease modeling. (A) Steatosis and insulin resistance can be induced in PHH spheroids by exposure to high insulin (IR) and supplementation with free fatty acids and fructose (“NAFLD”) for 7–14 days. Steatosis was measured as intracellular triglyceride levels, whereas insulin resistance was indicated as elevated expression of key genes involved in gluconeogenesis and de novo lipogenesis and reduced AKT phosphorylation upon insulin stimulation. IS, insulin-sensitive group. Error bars indicate S.E.M. * and **** indicate P < 0.05 and P < 0.0001, respectively. (B) The anti-NASH drug candidates cenicriviroc, elafibranor, and lanifibranor reduce free fatty acid–induced collagen deposition (COL1A1) and αSMA in PHH spheroids cocultured with liver nonparenchymal cells. (C) Exposure to high-fat media (black columns) increased expression of genes associated with fatty liver in a microfluidic liver culture system and reduced activity of CYP3A4 and CYP2C9. Error bars indicate S.D. * indicates P < 0.05. (D) PHH in micropatterned cocultures can be infected with HBV as evidenced by various viral life-circle readouts at 16 days postinfection (dpi). HBV surface antigen (HBsAg) was plotted as mean ± S.E.M. (n = 3). HBV covalently closed circular DNA copies were plotted as average of duplicates ± range. HBV core protein (HBVc) expression was indicated by immunostaining together with isotype-matched control. Figure modified with permission from (Hurrell et al., 2020; Kemas et al., 2021; Kostrzewski et al., 2017; Shlomai et al., 2014).
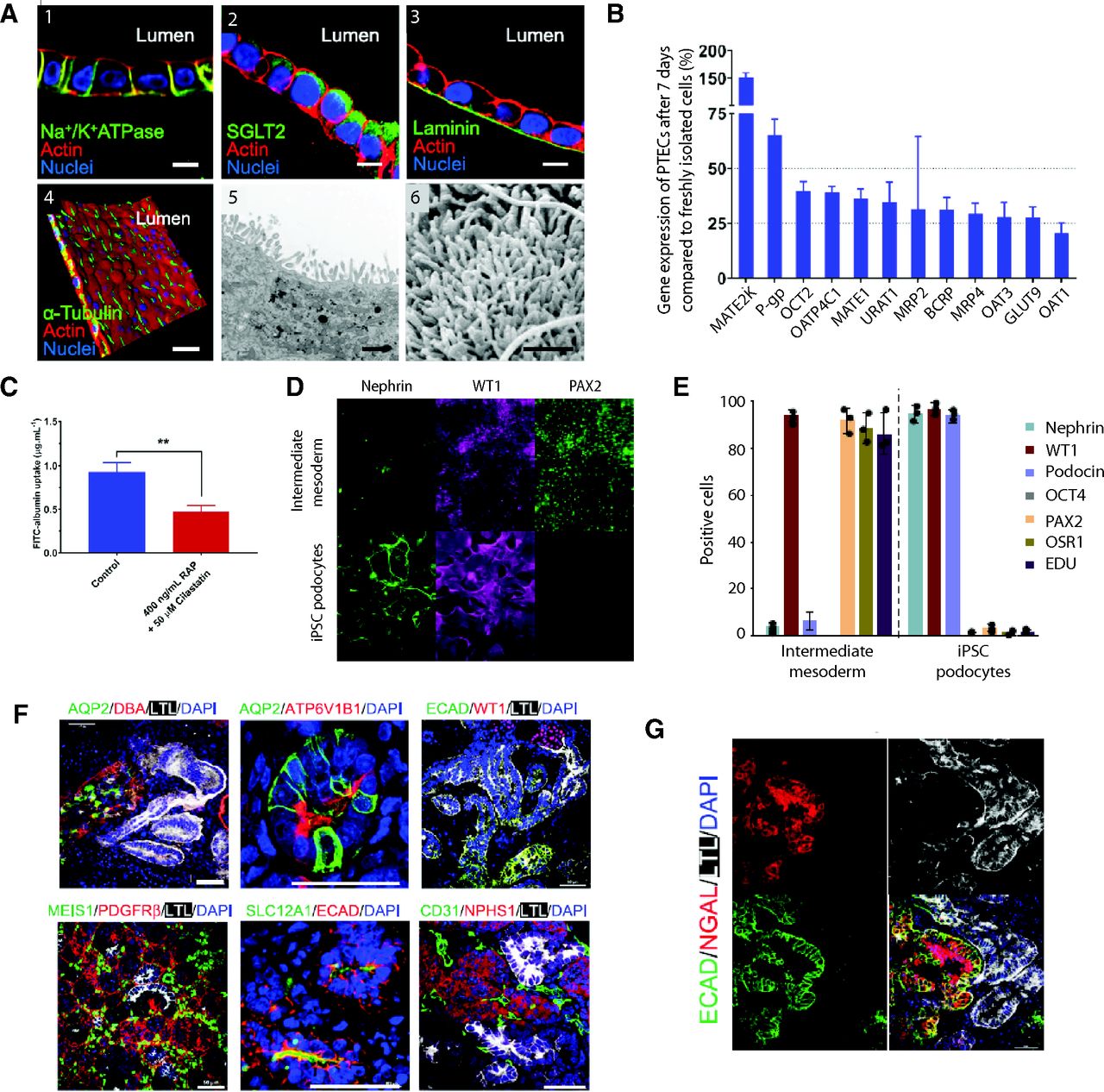
- Fig. 4
Overview of models of the human nephron. (A) Fluorescence images of immortalized human PTECs cocultured with glomerular microvascular endothelial cells showing basolateral expression of Na+/K+ ATPase (1) and apical expression of the glucose transporter SGLT2 (2) and deposition of a laminin-rich basement membrane (3). Furthermore, cells featured α-tubulin positive primary cilia (4). Transmission electron microscopy (5) and scanning electron microscopy (6) images showing dense carpets of microvilli. Scale bars for 1–4 = 10 µm; scale bars for 5–6 = 1 µm. (B) Expression of major renal transporters remains stable in primary human PTEC transwell cultures for at least 7 days. (C) Uptake of FITC-labeled albumin is sensitive to receptor-associated protein (RAP) and cilastatin, inhibitors of megalin/cubulin endocytosis, showing functionality of these endocytic receptors. Error bars indicate S.E.M. ** indicates P < 0.01. (D) Human iPSC-derived podocytes express nephrin and WT1, key markers of a mature phenotype, and lose expression of the intermediate mesoderm marker PAX2. (E) Quantification of changes in protein expression. Differentiated cells are positive for the podocyte markers nephrin, WT1, and podocin, whereas they are negative for the pluripotency marker OCT4 and the progenitor cell markers PAX2 and OSR1. Furthermore, iPSC-derived podocytes are nonproliferative as indicated by a lack of EdU incorporation. (F) Organoids differentiated using vasopressin and aldosterone show expression of the collecting duct markers (AQP2, DBA, and ATP6V1B1) as well as of proximal tubules (LTL), distal tubules (ECAD and SLC12A1), podocytes (WT1 and NPHS1), endothelial cells (CD31), and stroma (MEIS1 and PDGFRβ). Scale bars = 50 µm. (G) Response of collecting duct organoids to cisplatin showed upregulation of the proximal tubule injury marker HAVCR1 in LTL-positive proximal tubule cells and distal tubule injury marker NGAL in E-cadherin–positive distal tubules. Figure modified with permission from (Bajaj et al., 2020; Lin et al., 2019; Uchimura et al., 2020).
- Fig. 5
Human intestinal 3D models for studies of drug permeability and disease modeling. (A) Schematic depiction of a static transwell culture. Caco-2 cells are cultured as a monolayer on a porous membrane that separates the apical from the basolateral chamber. Measurement of TEER can be measured to evaluate barrier integrity. (B) Cross-sectional view of an integrated perfusion system for oral drug absorption. The drug of interest is first exposed to artificial gastric juice (pH 2) before being neutralized and mixed with artificial intestinal juice containing bile acids in the duodenum. Subsequently, the solution was perfused past a Caco-2 transwell culture to mimic intestinal permeability. Compound molecules that permeated into the basal channel were exposed HepG2 cells to mimic first-pass metabolism before subsequently entering the target compartment, comprising in this case MCF7 breast cancer cells. (C) Schematic depiction of an intestinal microchip comprising a stretchable porous membrane that separates two independently perfusable microchannels, which are lined by vacuum chambers that allow the repeated application of mechanical strain mimicking intestinal peristalsis. (D) Shear stress due to microperfusion (µF) and the application of cyclic mechanical strain (μF+St) increase cell height and polarization of Caco-2 monolayers while maintaining confluency as judged by localization of the tight junction protein occluding. Scale bars = 20 μm. (E) Intestinal organoids are comprised of villus domains containing enterocytes, enteroendocrine cells, and Goblet cells and alternating crypts enriched in LGR5-positive stem cells and Paneth cells. (F) Brightfield images (top row) and schematics (bottom row) of intestinal organoid injections using a customized robotic setup. For visualization purposes fluorescent DsRED-expressing E. coli are injected. Asterisk marks the needle tip. (G) Dissociated human intestinal organoids are seeded on the chip shown in (C and D). Immunofluorescence imaging shows expression of lysozyme (Lyz, Paneth cells), mucin 2 (Muc2, Goblet cells), chromogranin A (ChgA, enteroendocrine cells), and villin (cell apex). Figure modified with permission from (Imura et al., 2012; Kasendra et al., 2018; Kim et al., 2012; Robinson et al., 2019; Roeselers et al., 2013; Williamson et al., 2018).
- Fig. 6
Approaches for generating cerebral organoids for disease modeling. Easily accessible somatic cells from patients and controls can be reprogrammed into iPSCs that can be manipulated by genetic engineering to create isogenic lines. iPSCs can then be used to create either whole brain organoids through undirected differentiation or to region-specific organoids through patterned or directed differentiation. Region-specific organoids can moreover be fused to create so-called assembloids to recapitulate connectivity.
- Fig. 7
Organotypic platforms to mimic cardiac function and toxicity. (A) Schematic depiction of the most important endpoints in the evaluation of cardiomyocyte function. The arrival of an AP triggers intracellular calcium transients (Ca), which in turn entail contraction and the generation of contractile force (F). (B) Workflow showing the key principles of engineering laminar tissues and their integration within MEAs for electrophysiological studies or their formulation into contractility assays with embedded soft force gauges. Inlets on the right shows effect of terfinadine (Terf) on rate-corrected field potential duration (DcFPD; top) and an example optical micrograph of a deflecting cantilever with the corresponding electrical readout (bottom). Scale bar = 1 mm. (C) Key principles of the 3D EHT technology. Cardiomyocytes (CMs) encapsulated in a hydrogel are stretched and subjected to electrical stimulation, resulting in tissue compaction and maturation. Insert displays cross-section of a highly mature iPSC-derived EHT with wheat germ agglutinin in green and cardiac troponin T in red. Scale bars = 500 µm (i) and 10 µm (ii and iii). Figure modified with permission from (Kujala et al., 2016; Lind et al., 2017b; Ronaldson-Bouchard et al., 2018).
- Fig. 8
Engineered human skeletal muscle models. (A). Desmin and MyoD positive primary human myoblasts are generated by encapsulation in a hydrogel comprised of fibrinogen and matrigel matrix and subsequent casting into PDMS molds. (B and C) Differentiated human muscle fibers shorten over the course of 10 days in 3D culture (B = muscle cultures after gel polymerization, C = after 10 days in culture). (D and E) After compaction, the differentiated myofibers are densely packed and show characteristic striation of sarcomeric α-actinin (SAA). Furthermore, myogenin (MyoG)-positive nuclei and acetylcholine receptor (AChR)-positive domains can be seen. (F) Contractile force trace of a myoblast-derived myobundle showing that increased stimulation frequency results in stronger tetanic contraction. (G) Schematic depiction of a human motor unit on-a-chip formed by coculturing iPSC-derived motorneuron spheroids (MNs) and skeletal muscle fibers. Myoblasts were first injected into the right port of the chip. After 14 days of maturation, MN spheroids that were differentiated and maintained separately were injected into the left port, resulting in neural outgrowth, innervation of the differentiated muscle fiber constructs, and formation of neuromuscular junctions. (H) Comparison of motor units generated from control and ALS patient-derived stem cells. Note that the ALS motor unit has thinner neural fiber with less NMJ formed. Tuj1 (green; neuronal marker), F-actin (purple; muscle), and DAPI (blue; nuclei). Scale bars = 100 □m. (I) Optogenetic stimulation of neurons in ALS patient-derived motor units sometimes failed to translate into muscle contractions (red arrows). The number of such missed twitches could be reduced by cotreatment with rapamycin and bosutinib. Blue dashed lines represent light stimulation intervals (i.e., 200 ms). Figure modified with permission from (Chiron et al., 2012; Madden et al., 2015; Osaki et al., 2018).
- Fig. 9
Schematic depiction of bioprinting technology. The cells to be printed are embedded in a biocompatible matrix comprising polymers, nutrients, growth factors, and functional peptides jointly termed bioink. This solution can be deposited with high spatiotemporal resolution using different methodologically distinct modalities. In extrusion-based bioprinting (left), the cell-laden bioinks are deposited as continuous streams, whereas droplet-based bioprinting (middle) partitions the bioink into individual microdroplets. In contrast to both of these methods, laser-assisted bioprinting (right) deposits cells using a nozzle-free approach in which the bioink is locally heated using a UV laser and then drops onto the printing stage.








Tables
Cell Model Cell Type Mono-/Coculture Exposure Regimen Exposure Time TC50 Reference Toxicity Mechanism: Reactive Metabolites
Acetaminophen (Cmax = 0.136 mM; TC50 in mM)Spheroid PHH Monoculture Repeated 14 days 0.32–0.8 (Bell et al., 2018) Spheroid PHH Monoculture Repeated 14 days 0.46 (Bell et al., 2020) Spheroid PHH Coculture with NPCs Repeated 14 days ∼0.5 (Richert et al., 2016) Spheroid PHH Coculture with NPCs Repeated 14 days 0.57 (Proctor et al., 2017) Spheroid PHH Coculture with Kupffer cells Repeated 14 days ∼0.6 (Richert et al., 2016) Spheroid PHH Monoculture Repeated 14 days 0.64 (Bell et al., 2017) Spheroid PHH Coculture with NPCs Repeated 14 days 0.75 (Messner et al., 2013) Spheroid PHH Monoculture Repeated 14 days 0.88 (Hendriks et al., 2016) Spheroid PHH Monoculture Single 24 h 0.93 (Li et al., 2020) Spheroid HepaRG Monoculture Repeated 14 days 1.1 (Hendriks et al., 2016) Spheroid HepaRG Monoculture Single 24 h ∼1.1 (Leite et al., 2012) Spheroid PHH Monoculture Repeated 96 h 1.21 (Bell et al., 2020) Spheroid PHH Monoculture Repeated 8 days 1.3 (Hendriks et al., 2016) Spheroid PHH Coculture with NPCs Repeated 14 days 1.35 (Bell et al., 2020) Spheroid HepaRG Monoculture Repeated 8 days 1.8 (Hendriks et al., 2016) Spheroid PHH Coculture with NPCs Repeated 10 days 1.7 (Foster et al., 2019) Spheroid PHH Coculture with Kupffer cells Single 5 days 2.25 (Li et al., 2020) Perfused liver-chip PHH Coculture with LSECs Repeated 10 days 2.4 (Foster et al., 2019) Spheroid PHH Coculture with NPCs Repeated 96 h 2.46 (Bell et al., 2020) Spheroid HepaRG Monoculture Single 24 h 2.48 (Wang et al., 2016) Spheroid HepaRG Monoculture Single 24 h 2.7 (Gunness et al., 2013) Spheroid HepaRG Monoculture Repeated 6 days 2.879 (Ramaiahgari et al., 2017) Spheroid PHH Monoculture Single 5 days 3.28 (Li et al., 2020) Spheroid HepaRG Monoculture Repeated 7 days ∼4 (Liu et al., 2018a) MPCC iPSC-HLC Monoculture Repeated 6 days 4.48 (Ware et al., 2015) Perfused liver-chip PHH Coculture with NPCs Repeated 14 days 5.59 (Novik et al., 2017) Spheroid HepG2 Monoculture Repeated 4 days 7.21 (Gaskell et al., 2016) Spheroid HepaRG Monoculture Single 24 h 7.62 (Wang et al., 2015) Spheroid HepG2 Monoculture Repeated 6 days ∼9.4 (Ramaiahgari et al., 2014) Spheroid hiPSC/hESC-HLC Monoculture Single 24 h >10 (Tasnim et al., 2016) Spheroid hiPSC-HLC Monoculture Single 24 h >20 (Takayama et al., 2013) Spheroid HepG2 Monoculture Single 24 h >20 (Takayama et al., 2013) MPCC PHH Monoculture Single 24 h 35 (Khetani and Bhatia, 2008) Spheroid HepG2 Monoculture Single 24 h 40 (Fey and Wrzesinski, 2012) Toxicity Mechanism: Inhibition of Mitochondrial Respiration
Amiodarone (Cmax = 5 μM; TC50 in μM)Spheroid PHH Monoculture Repeated 28 days 1.6 (Bell et al., 2016) Spheroid PHH Monoculture Repeated 14 days 6.5 (Bell et al., 2016) Perfused liver-chip PHH Coculture with NPCs Repeated 14 days 9 (Novik et al., 2017) Spheroid PHH Monoculture Repeated 14 days 12 (Bell et al., 2017) Spheroid PHH Monoculture Repeated 14 days 12 (Hendriks et al., 2016) MPCC PHH Monoculture Repeated 14 days 14 (Khetani et al., 2013) Spheroid hiPSC-HLC Monoculture Single 24 h >25 (Takayama et al., 2013) Spheroid PHH Monoculture Single 5 days 26 (Li et al., 2020) Spheroid PHH Coculture with NPCs Repeated 14 days 32 (Proctor et al., 2017) Spheroid HepaRG Monoculture Single 7 days 49 (Ott et al., 2017) Spheroid HepG2 Monoculture Single 24 h ∼50 (Takayama et al., 2013) Spheroid PHH Coculture with NPCs Repeated 14 days ∼60 (Richert et al., 2016) Spheroid HepaRG Monoculture Single 24 h 83 (Ott et al., 2017) Spheroid HepaRG Monoculture Single 24 h 178 (Mueller et al., 2014) Spheroid HepG2 Monoculture Single 24 h >200 (Mueller et al., 2014) Spheroid HepG2 Monoculture Single 24 h 260 (Fey and Wrzesinski, 2012) Toxicity Mechanism: Cholestasis
Chlorpromazine (Cmax = 0.84 μM; TC50 in μM)Spheroid PHH Monoculture Single 5 days 4.4 (Li et al., 2020) Spheroid PHH Monoculture Repeated 14 days 4.6 (Bell et al., 2017) Spheroid PHH Coculture with NPCs Repeated 14 days ∼7 (Proctor et al., 2017) Spheroid PHH Coculture with only kupffer cells Repeated 14 days ∼9 (Richert et al., 2016) Spheroid PHH Coculture with NPCs Repeated 14 days ∼10 (Richert et al., 2016) Spheroid PHH Monoculture Repeated 14 days 11 (Hendriks et al., 2016) Spheroid PHH Monoculture Repeated 8 days 13 (Hendriks et al., 2016) Spheroid HepaRG Monoculture Repeated 14 days 13 (Hendriks et al., 2016) Spheroid HepaRG Monoculture Repeated 8 days 16 (Hendriks et al., 2016) Perfused liver-chip PHH Coculture with NPCs Repeated 14 days 28 (Novik et al., 2017) Spheroid HepaRG Monoculture Repeated 7 days ∼40 (Liu et al., 2018a) Spheroid HepG2 Monoculture Single 24 h 43 (Mueller et al., 2014) Spheroid HepaRG Monoculture Single 24 h 98 (Mueller et al., 2014) MPCC PHH Monoculture Single 24 h 100 (Khetani and Bhatia, 2008) Spheroid HepaRG Monoculture Single 24 h 120 (Wang et al., 2015) Toxicity Mechanism: Mitochondrial Depletion
Fialuridine (Cmax = 0.64 μM; TC50 in μM)Spheroid PHH Monoculture Repeated 28 days 0.1 (Bell et al., 2016) Spheroid PHH Monoculture Repeated 32 days 0.28 (Hendriks et al., 2019) Spheroid PHH Monoculture Repeated 14 days 0.7 (Bell et al., 2016) Spheroid PHH Monoculture Repeated 14 days ∼3 (Bell et al., 2018) Spheroid PHH Monoculture Repeated 7 days 7.86 (Hendriks et al., 2019) Spheroid PHH Coculture with NPCs Repeated 10 days 77 (Foster et al., 2019) Perfused liver-chip PHH Coculture with LSECs Repeated 10 days 84 (Foster et al., 2019) Spheroid PHH Coculture with NPCs Repeated 14 days >100 (Richert et al., 2016) Spheroid HepG2 Monoculture Repeated 4 days 200 (Gaskell et al., 2016) hESC, human embryonic stem cell.
- TABLE 2
Overview of studies using organotypic liver cultures for hepatic clearance predictions
Number of Compounds Predicted within 2-Fold of In Vivo Cl Culture Paradigm Low-Clearance Compounds Medium- to High-Clearance Compounds Prediction Method Results/Conclusion Reference PHH in MPCC 7/10 4/7 Conventional Medium- to high-Cl compounds mainly underpredicted (Chan et al., 2013) PHH in MPCC 9/15 3/10 Conventional High-Cl compounds underpredicted (Da-silva et al., 2018) PHH in MPCC 5/8 — Direct scaling No clear tendency (Kratochwil et al., 2017) PHH in MPCC 4/12 2/3 Conventional Clinical drug candidates (low Cl) were better predicted using direct scaling (Kratochwil et al., 2017) PHH spheroids 2/3 1/3 Conventional Compounds were underpredicted (Kanebratt et al., 2021) PHH in chip 0/2 3/4 Direct scaling Low-Cl compounds below LOQ, high Cl underpredicted (Chao et al., 2009) Cl, clearance; LOQ, limit of quantification.
Nephron Part Cell Origin Applications Pros Cons Proximal tubule Primary PTECs Drug transport and nephrotoxicity TEER similar to native tissue, metabolic enzymes, and drug transporters preserved Short-term cultures (up to 2 weeks max), limited access to primary material iPSCs Drug transport and nephrotoxicity Can be expanded, compatible with long-term culture Immature expression of uptake transporters Immortalized PTECs Drug transport and nephrotoxicity Can be expanded (30 passages), compatible with long-term culture Immature expression of uptake transporters Glomerulus Primary podoctyes Studies of podocytopathies Express slit diaphragm proteins, patient-derived podocyte cultures are possible Limited access to primary material iPSCs Study of podocytopathies Long-term cultures, generation of isogenic lines Loss of foot processes and slit diaphragms in most of models Distal tubule and collecting duct iPSCs Drug testing, disease model Expression of AQP2, response to toxins Most models show incomplete maturation of distal tubule and collecting duct Transwell Cultures Perfusion Systems Ussing Chambers Organoids Explant Cultures Medium dynamics Static Perfused Static Static Static Available drug permeability data +++ + +++ — ++ Cellular complexity Enterocytes +/− Goblet cells and M cells Enterocytes, Goblet cells, enteroendocrine cells, Paneth cells All intestinal cell types Enterocytes, Goblet cells, enteroendocrine cells, Paneth cells All intestinal cell types Molecular phenotype + ++ +++ ++ +++ Mucus Some, if Goblet cells are added Can reach physiologic levels Yes Some Yes Phenotypic stability Multiple weeks Multiple weeks Hours Multiple weeks, allow passaging 6–24 h Throughput +++ ++ + +++ + Ease of use +++ + ++ + ++ Costs +++ + ++ +++ ++ Model versatility ++ +++ + +++ ++ Microbiota No Yes No Yes No Peristalsis No Yes No No No Main applications Drug absorption Disease modeling Drug absorption Intestinal development, regenerative medicine Drug absorption, acute toxicity Disease Gene Cells Protocol Phenotype Reference Micro-cephaly CDK5RAP2 hiPSCs Cerebral cortex Smaller neuroepithelial regions, altered spindle orientation of radial glial cells, abundant neuronal outgrowth, smaller organoid size. (Lancaster et al., 2013) Seckel syndrome CPAP hiPSCs Cerebral cortex Extended G1‐S transition in NPCs from Seckel cell patient iPSCs. Premature NPC differentiation resulting in reduced cell number. (Gabriel et al., 2016) Macro-cephaly PTEN hESCs Cerebral cortex Promoted cell cycle re-entry and delayed neuronal differentiation, resulting in a pronounced expansion of the radial glia and intermediate progenitor cells and increased organoid size. (Li et al., 2017) Autism spectrum disorder — hiPSCs Dorsal telencephalon Abnormal proliferation of neural progenitor cells and an increased production of inhibitory GABAergic neurons, enhanced FOXG1 expression (Mariani et al., 2015) Schizophrenia DISC1 hiPSCs Forebrain Prolongs mitotic length and cell-cycle deficits in radial glial cells (Ye et al., 2017) Schizophrenia FGFR1 hiPSCs and hESCs Cerebral cortex Disorganized migration of proliferating cells and depletion of cortical neurons. Cortical malformation and altered FGFR1 signaling. (Stachowiak et al., 2017) Schizophrenia — hiPSCs Dorsal forebrain Differential expression of genes involved in synaptic biology and neurodevelopment. Subdued responses to stimulation and depolarization. Mitochondrial functional defects. (Kathuria et al., 2020) Alzheimer disease — hiPSCs Cortex Generation of pathologic amyloid β peptides (Lee et al., 2016) Alzheimer disease APP hiPSCs Neocortex β-amyloid aggregation, hyperphosphorylated τ, endosome abnormalities (Raja et al., 2016) Alzheimer disease — hiPSCs Triculture of neurons, astrocytes, and microglia Increased Aβ aggregation, phosphorylated τ accumulation, neuroinflammatory activity, and microglia recruitment (Park et al., 2018) Parkinson disease LRRK2 hiPSCs Midbrain Increased aggregation of α-synuclein and decreased dopaminergic neurite length (Kim et al., 2019) Miller-Dieker syndrome — hiPSCs Forebrain Increased apoptosis and horizontal divisions in neural stem cells, mitotic delay in outer radial glia-like cells (Bershteyn et al., 2017) Miller-Dieker syndrome LIS1 hiPSCs Forebrain Smaller organoids with fewer neuroepithelial loops, fewer symmetric ventricular radial glia cells divisions, disrupted cortical niche (Iefremova et al., 2017) Rett syndrome MeCP2 hiPSCs Undefined Aberrant neurogenesis and neuronal differentiation, increased ventricular area, and decreased radial thickness of organoid (Mellios et al., 2018) ZIKA virus infection ZIKV hiPSCs Forebrain Suppression of neural progenitor cell proliferation, decreased neuronal layer thickness and organoid size, enlarged lumen/ventricles (Qian et al., 2016) ZIKA virus infection ZIKV hESCs Cerebral cortex ZIKV-mediated TLR3 activation, reduction in organoid volume resembling microcephaly. (Dang et al., 2016) ZIKA virus infection ZIKV-NS2A hiPSCs Forebrain ZIKV-NS2A impairs radial glial cell proliferation and adherens junction formation (Yoon et al., 2017) hESC, human embryonic stem cell.
Pluripotent Stem Cells Primary Myoblasts Mature Myofiber Cultures Maturation method Transgene- or small molecule-based Transgene- or small molecule–based Not applicable Phenotype + ++ +++ Length of differentiation protocol + ++ Not applicable Stability in culture +++ ++ + Accessibility +++ ++ + Homogeneity of culture + ++ +++ Functionality + ++ +++ Current use in drug testing +++ ++ Not tested yet - TABLE 7
Overview of multiorgan-on-a-chip combinations used for pharmacological or toxicological applications
Application Tissues Type of Culture Type of Cells Findings References Toxicity Liver and heart 3D iPSC-derived Metabolic interaction underlying clomipramine toxicity (Yin et al., 2021) Toxicity Liver and heart 2D and 3D iPSC-derived and primary cells Metabolic interaction underlying cyclophosphamide and terfenadine toxicity (Oleaga et al., 2018) Toxicity Liver and heart 3D iPSC-derived and primary cells Tissue-specific toxicity of acetaminophen and doxorubicin (Zhang et al., 2017) Toxicity Liver and kidney 2D Cell lines and primary cells Metabolic interaction underlying ifosfamide and verapamil nephrotoxicity (Li et al., 2018c) Toxicity Liver and kidney 2D Cell lines Tissue interactions in vitamin D3 bioactivation (Theobald et al., 2019) Toxicity Liver and lung 3D and ALI Cell lines Liver cells reduce aflatoxin B1 pulmonary toxicity (Bovard et al., 2018) Toxicity Liver and lung 3D and ALI Cell lines Liver cells reduce aflatoxin B1 pulmonary toxicity (Schimek et al., 2020) Toxicity Liver, heart, and lung 3D iPSC-derived and primary cells Lung is essential in bleomycin-induced cardiotoxicity (Skardal et al., 2017) Toxicity Liver, heart, lung, endothelium, brain, and testes 3D iPSC-derived, cell lines, and primary cells Metabolic interaction underlying ifosfamide neurotoxicity (Rajan et al., 2020) Toxicity Liver, brain, pancreas, lung, heart, gut, and endometrium 2D Cell lines and primary cells Tolcapone metabolism and mechanism of action (Wang et al., 2019b) Toxicity Liver, cancer, bone marrow 3D Cell lines Hepatic bioactivation of capecitabine and tegafur (LaValley et al., 2021) Toxicity Liver and cancer 2D Cell lines Hepatic bioactivation of capecitabine and tegafur (Satoh et al., 2017) Toxicity Liver and cancer 2D and 3D Cell lines Hepatic bioactivation and inactivation of ifosfamide and temozolomide, respectively (Ma et al., 2012) Toxicity Liver and cancer 2D Cell lines Effects of hepatic metabolism on luteolin toxicity (Lee et al., 2017b) Toxicity Liver and cancer 2D Cell lines Hepatic bioactivation of irinotecan (Shinha et al., 2020) Toxicity Liver, intestine, and lung 2D Cell lines Hepatic bioactivation of cyclophosphamide or irinotecan (Kimura et al., 2015) Toxicity Liver, lung, kidney, and adipose tissue 3D Cell lines Tissue-specific effects of TGFβ (Zhang et al., 2009) Toxicity Liver, heart, lung, endothelium, testis, colon, and brain 3D Primary stem cells and primary cells Comparison of tissue-specific toxicity in coculture (Skardal et al., 2020) PK Liver and intestine 3D Cell lines and primary cells System retains drug absorption of panadol, mannitol, and caffeine (Chen et al., 2018) PK Liver and intestine 2D and 3D Cell lines and primary cells System retains drug permeability similar to monoculture (Esch et al., 2016) PK Liver and intestine 2D Cell lines Apigenin mertabolism in both intestine and liver (Choe et al., 2017) PK Liver and intestine 3D Cell lines and primary cells Estimation of diclofenac and hydrocortisone permeability and clearance (Tsamandouras et al., 2017) PK Liver and intestine 2D Cell lines Absorption of fatty acid and evaluation of antisteatotic effect of metformin and XL-335 (Jeon et al., 2021) Gut-brain-axis Brain and intestine 2D iPSC-derived Evaluation of the impact of the intestinal microflora on neurodegeneration (Raimondi et al., 2019) Gut-brain-axis Liver, intestine, and brain 2D and 3D Primary stem cells, cell lines, and primary cells Microbiome-derived short-chain fatty acids increase the expression of pathology-associated pathways in neurodegenerative disease (Trapecar et al., 2021) Metastasis formation Brain and lung 2D Cell lines NSCLC metastasizing propensity to brain (Liu et al., 2019) Glucose metabolism Liver and pancreas 3D Cell lines and primary cells Recapitulation of glucose metabolism and homeostasis (Bauer et al., 2017) Glucose metabolism Liver, pancreas, and skeletal muscle 2D Cell lines Recapitulation of glucose metabolism and homeostasis (Lee et al., 2019) ALI, air-liquid interface; NSCLC, non–small cell lung cancer; PK, pharmacokinetics.
Extrusion-
BasedDroplet-
BasedLaser-
AssistedCell viability 50%–70% 80%–90% 95% Printing precision + +++ ++ Printing resolution 100–200 µm 30–50 µm 10 µm Throughput + +++ ++ Cell density 108 cells/ml <106 cells/ml <106 cells/ml Bioink viscosity 30–107 mPa 3–12 mPa 1–300 mPa Single-cell deposition No Inconsistent Yes Possibility to print
scaffold-freeYes Yes No Cost Low Medium High Commercial availability +++ ++ + - TABLE 9
Advantages and disadvantages of commonly used materials for microphysiological devices
Glass Silicon Thermoplastics Thermosets Elastomers Material properties Knowledge base in biology High Medium Medium Medium Good Optical transparency High None Medium-good Medium-good Good Gas permeability Very low Very low Low Low Medium-high Mechanical robustness High Good Good-high Good-high Low Tunable mechanical property Low Low Medium Medium-good Low Chemical leachability None None Low-medium Low-medium High Hydrophobic molecule sorption Very low Very low Low-medium Low-medium High Cell attachment Very high High Medium-high Medium-high Low-medium Manufacturing properties Common fabrication methods Photolithography Photolithography, E-beam lithography Injection molding Soft lithography, reaction injection molding Soft lithography Ease of rapid prototyping Very low Low Low Medium-high Medium-high Cost of prototyping (material and fabrication) Very high High Medium Low-medium Low Industrial microstructuring capability High High High Medium-high Low Scalability Medium-high Medium-high High Good Low
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- I. Introduction
- II. Liver
- III. Kidney
- IV. Intestine
- V. Brain
- VI. Heart
- VII. Skeletal Muscle
- VIII. Fluidic Integration of Organotypic Tissue Models
- IX. Bioprinting as a Novel Modality for the Generation of Organotypic Cultures
- X. Technical Considerations for Culture Devices
- XI. Conclusions and Perspectives
- Authorship Contributions
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters