



Abstract
We have experienced an enormous cohesive effort of the scientific community to understand how the immune system reacts to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and how to elicit protective immunity via vaccination. This effort resulted in the development of vaccines in record time with high levels of safety, efficacy, and real-life effectiveness. However, the rapid diffusion of viral variants that escape protective antibodies prompted new studies to understand SARS-CoV-2 vulnerabilities and strategies to guide follow-up actions to increase, and maintain, the protection offered by vaccines. In this review, we report the main findings on human immunity to SARS-CoV-2 after natural infection and vaccination; we dissect the immunogenicity and efficacy of the different vaccination strategies that resulted in products widely used in the population; and we describe the impact of viral variants on vaccine-elicited immunity, summarizing the main discoveries and challenges to stay ahead of SARS-CoV-2 evolution.
Significance Statement This study reviewed findings on human immunity to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), analyzed the immunogenicity and efficacy of the various vaccines currently used in large vaccination campaigns or candidates in advanced clinical development, and discussed the challenging task to ensure high protective efficacy against the rapidly evolving SARS-CoV-2 virus. This manuscript was completed prior to the emergence of the Omicron variant and to global vaccine boosting efforts.
I. Introduction
Nearly two years have passed since the discovery of a novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) causing coronavirus disease (COVID-19) in Wuhan, China. The sudden emergence and rapid spread around the globe of SARS-CoV-2 drove many of the world’s governments to implement far-reaching health measures such as mandatory mask wearing, restrictions on public gathering and travel, and requirements for physical distancing to stop uncontrolled viral transmission, all in an effort to safeguard the public’s health. Significant funds were committed by both public authorities and private enterprises to find interventions against COVID-19. Large-scale efforts were conducted by the scientific community to understand the biology, pathology, and epidemiology of SARS-CoV-2 and to identify potential determinants of immunity in the human host. This rapidly growing trove of scientific knowledge was made publicly available at unprecedented levels and provided the foundation for the development of new treatments and vaccines with the clear objective of stopping the pandemic. In this review, we gather evidence of natural and vaccine-induced immunity against SARS-CoV-2, the role of vaccine platforms in eliciting protective immunity, the impact of emerging virus variants with increased infectiveness and resistance to antibody-mediated neutralization, and current and future challenges that threaten the progress the world has made in stopping the pandemic.
II. Infection and Disease
Viruses have evolved to colonize and replicate within their hosts, deploying multiple strategies that ensure their survival. Similarly, our immune system has developed a multiplicity of mechanisms to counteract viral infection (Abbas et al., 2012). In Fig. 1, we summarize the main features of antibody- and T cell-mediated immunity, which are important for understanding the immune response to SARS-CoV-2 in the following sections of this review.
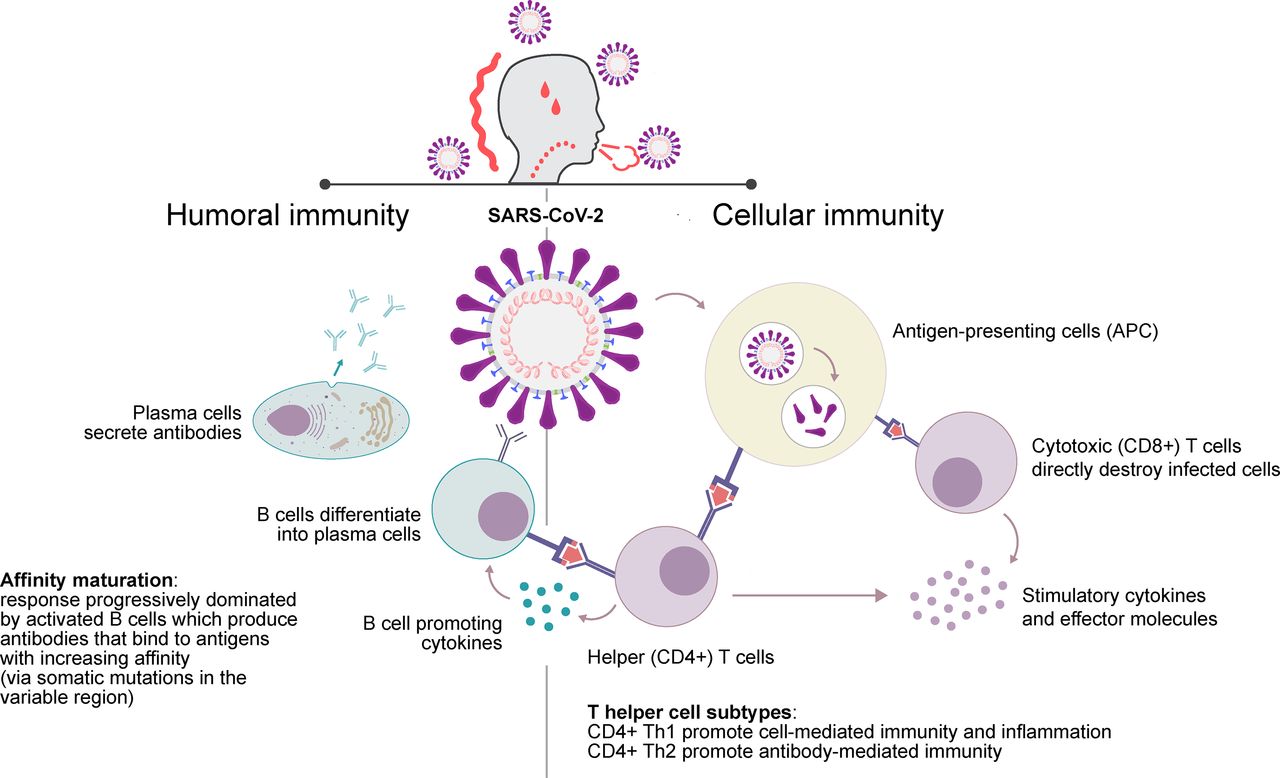
Basics of antibody and cell-mediated immunity. After encounter with the virus, immune cells known as antigen-presenting cells activate antigen-specific T cells that act either as support to other cell types by producing cytokines and stimulatory molecules (CD4+, also referred to as helper T cells) or attack the infected cells participating in infection clearance (CD8+, also referred to as cytotoxic T cells). Helper T cells can be further divided in Th1, Th2, or other subtypes, which differentially affect the immune effector response. Antigen-specific B cells, also acting as antigen-presenting cells, are activated by helper T cells and either immediately start producing antibodies or undergo affinity maturation into the germinal center of secondary lymphoid tissues (not represented in the figure) before differentiating into plasma cells that produce high affinity antibodies that neutralize the virus (Abbas et al., 2012).
A. Emergence of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
The newly emerged SARS-CoV-2 belongs to the family of coronaviruses, highly infectious viruses of zoonotic origin that can be both endemic, with seasonal patterns, and epidemic, with sudden appearance in the human population after spillover from animal hosts. Particularly, severe acute respiratory syndrome coronavirus (SARS-CoV-1) and Middle East respiratory syndrome coronavirus (MERS-CoV) originated in market civets and dromedary camels causing fatal outbreaks of respiratory illness in 2002–2003 and 2012, respectively [reviewed in (Cui et al., 2019)]. Other seasonal coronaviruses, HCoV-NL63, HCoV-229E, HCoV-OC43, and HKU1, induce mild upper respiratory disease in immunocompetent human hosts (Su et al., 2016). Importantly, no licensed vaccines against coronaviruses were available in humans when SARS-CoV-2 suddenly emerged.
All coronaviruses are enveloped by a lipid bilayer membrane of spherical shape with a diameter of approximately 150 nm and decorated by the spike (S) trimeric protein, the main viral receptor that gives the virus the typical sun-like shape when observed using an electron microscope [reviewed by (Chen et al., 2020)]. The S protein is divided into the S1 and S2 subunits, which are responsible for binding to the human angiotensin-converting enzyme 2 (ACE2) and fusing with the cellular membrane of alveolar epithelial cells and other cell types, respectively, determining viral entry. The envelope (E) protein, involved in virion assembly and envelope formation, and the membrane (M) protein, which stabilizes the envelope and binds the nucleocapsid on its internal side, are also part of the envelope. Within the envelope, the nucleocapsid (N) protein is complexed with the positive, single-stranded RNA genome encoding for the aforementioned functional proteins, the replicase (ORF1a/ORF1b) and several accessory species-specific proteins relevant to viral replication (Chen et al., 2020) (Fig. 2A).

Immunity to SARS-CoV-2. (A) Immunologic targets of SARS-CoV-2, highlighting the four main viral antigens. (B) Plot of putative correlation of neutralizing antibody levels with protection from infection and disease. A first wave of IgM and IgA antibodies might confer a degree of protection against SARS-CoV-2, but their levels rapidly decrease. On the contrary, IgG antibodies, appearing later during natural infection, are mostly correlated with protection. In case of active immunization, neutralizing IgG responses are induced in naïve individuals, with levels varying depending on the vaccine platform and immunization strategy used. In the case of vaccination of previously infected individuals, neutralizing IgG levels are much higher, boosting memory B cell responses developed during natural infection. In time, IgG levels might decrease below a protective threshold, requiring booster vaccination.
The novel virus SARS-CoV-2 is closely related to SARS-CoV-1—with around 80% genome homology—and other coronaviruses of bat origin [reviewed by (Hu et al., 2021)] (Chan et al., 2020). SARS-CoV-2 was first reported in December 2019 in Wuhan, China, and isolated in January 2020 from patients suffering from viral pneumonia (Zhu et al., 2020c). Its viral genome was sequenced using a metagenomics approach and, within a month, the sequence was shared globally via the Global Initiative on Sharing Avian Influenza Data initiative, prompting a rapid response from the scientific community (Rappuoli et al., 2021). Along with many similarities to SARS-CoV-1, the new virus had a distinct S protein with a four-amino acid residues insertion in the junction between S1 and S2, which generated a polybasic cleavage site enabling furin cleavage (Coutard et al., 2020). Like other coronaviruses, SARS-CoV-2 requires proteolytic processing of the S protein to induce cell-cell fusion, thus the insertion of a functional furin cleavage site was associated with enhanced cell entry and virulence (Andersen et al., 2020).
B. Coronavirus Disease (COVID-19) Caused by SARS-CoV-2
The pathogenesis of COVID-19 starts with SARS-CoV-2 binding to epithelial cells in the respiratory tract, expressing ACE2. Rapid viral replication after migration to the lungs triggers an immune response. Individuals infected with SARS-CoV-2 can be asymptomatic, effectively controlling the infection, or can present mild symptoms such as chest pain, respiratory discomfort, and pneumonia, potentially progressing to severe clinical manifestations, such as respiratory compromise, systemic inflammatory syndrome, and finally multiorgan failure and death (summarized in Fig. 3) (Brodin, 2021; Hu et al., 2021; Sudre et al., 2021).

Clinical features of COVID-19. SARS-CoV-2 infection is characterized by three stages of acute infection of progressive severity. After the acute phase, a period of persistent disease can occur (Brodin, 2021; Hu et al., 2021; Sudre et al., 2021). *Postacute sequalae in <5% of patients (Sudre et al., 2021).
Dysregulation of inflammatory cytokines with the elevated release of interleukins IL-1, IL-6, IL-8, and tumor necrosis factor alpha, defined as “cytokine storm,” is a hallmark of coronaviruses dissemination correlated with severe disease and poor prognosis (Channappanavar and Perlman, 2017; Huang et al., 2020b). In particular, virus-induced proinflammatory levels of IL-6 have been hypothesized to increase the risk of fatal disease (Santa Cruz et al., 2021). The cytokine storm has been mostly associated with patients over 60 years of age and with pre-existing medical conditions (Liu et al., 2020b; Ragab et al., 2020).
After remission of severe illness, some individuals, the so called COVID long-haulers, suffer from debilitating symptoms for extended periods of time, often lasting for several months. This condition, dubbed “long COVID”, resembles postdisease syndromes associated with chikungunya and Ebola virus infections (Brodin, 2021; Korompoki et al., 2021).
III. Immunity to SARS-CoV-2
In the first months of the COVID-19 pandemic, the identification of determinants of immunity to SARS-CoV-2 became paramount for guiding patient treatment strategies and vaccine design. Emerging data gathered from animal models and from patients recovering from COVID-19 revealed the importance of both humoral and cellular immunity to protect from SARS-CoV-2 and other coronaviruses infection [reviewed by (Dai and Gao, 2021; Huang et al., 2020a; Poland et al., 2020)].
A. Targets of Antibody-Mediated Immunity
Antibodies are the primary defense mechanism against viruses, blocking viral receptor binding and inhibiting viral entry and replication. Mechanistically, antibodies against the SARS-CoV-2 S protein neutralize the virus by blocking its interaction with ACE2 on the surface of alveolar epithelial cells in the lower respiratory tract and other cell types, a phenomenon observed both for SARS-CoV-1 and SARS-CoV-2 (Ju et al., 2020; Lan et al., 2020; Song et al., 2018; Wan et al., 2020; Wrapp et al., 2020). Evidence of immunodominance of the S protein came from both SARS-CoV-1 and SARS-CoV-2 studies, which identified the receptor binding domain (RBD) in the S1 subunit of the S protein as the main target of antibody-dependent neutralization (Wong et al., 2004; Woo et al., 2005; Premkumar et al., 2020). Indeed, antibodies targeting RBD accounted for 90% of the convalescent serum neutralizing activity (Piccoli et al., 2020). The kinetics of the antibody response in COVID-19 patients is characterized by rapid seroconversion 7–14 days after symptom onset (Norman et al., 2020). In longitudinal studies, IgM and IgA antibodies are the first antibodies to be detected, which peak at 10–12 days (IgM) and 20–22 days (IgA) and then decline, followed by IgG antibodies, which increase within the first three weeks and persist in the blood for long periods of time (Adams et al., 2020; Lou et al., 2020; Ni et al., 2020; Okba et al., 2020; Padoan et al., 2020; Bartsch et al., 2021). Similarly, asymptomatic individuals rapidly seroconverted after SARS-CoV-2 infection, with elicitation of primarily IgG antibodies (Jiang et al., 2020) (Fig. 2B). Antibody responses mainly target S and N proteins, and S-specific IgG levels were shown to correlate with neutralization (Okba et al., 2020). In two studies, serum IgG levels were found to rapidly decay 2–3 months postinfection in asymptomatic or mild COVID-19 patients, raising concerns about the duration of immunity (Ibarrondo et al., 2020; Long et al., 2020). On the contrary, persistence of IgG was longer, beyond three months, in other studies assessing patients with moderate or severe disease (Isho et al., 2020; Iyer et al., 2020; Wajnberg et al., 2020). Importantly, disease severity was associated with earlier seroconversion and higher antibody titers, a phenomenon observed both for SARS-CoV-2 and MERS-CoV (Ko et al., 2017; Wang et al., 2020c). This evidence could imply a role of virus-specific antibodies in the worsening of the disease, a phenomenon known as antibody-dependent enhancement (ADE), previously described for respiratory syncytial virus (RSV) infection and occurring in vitro in SARS-CoV-1 experiments. However, current clinical data are insufficient to implicate ADE in COVID-19 (Arvin et al., 2020; Graham, 2020).
Strong evidence of viral neutralization by S-specific IgG and the need for quick remedies for COVID-19 patients while vaccines were still in development led to a focus on the isolation of monoclonal antibodies (mAbs) from COVID-19 convalescent patients (Cohen, 2020). Several research groups deployed state-of-the-art technologies that were rapidly adapted from the human immunodeficiency virus (HIV), RSV, and Ebola research fields (Cohen, 2020). Indeed, several potent mAbs were isolated against the S protein, most of them targeting the RBD domain, which help defining critical characteristics of antibody-mediated protection to SARS-CoV-2 (Barnes et al., 2020a; Barnes et al., 2020b; Baum et al., 2020b; Brouwer et al., 2020; Hansen et al., 2020; Ju et al., 2020; Piccoli et al., 2020; Pinto et al., 2020; Tortorici et al., 2020; Wang et al., 2020a; Wu et al., 2020b; Andreano et al., 2021). Additional studies determined that the IgG heavy-chain variable region 3-53 (VH3-53) was used most frequently among RBD-targeting antibodies (Barnes et al., 2020a; Yuan et al., 2020). These antibodies, defined as class 1, represent a public antibody response to the RBD epitope in the “up” conformation, which mediates binding to the ACE2 receptor. Class 2 antibodies exhibit more diverse VH-gene usage and bind the RBD in both “up” and “down” conformations. A second S protein neutralization target, the N-terminal domain (NTD), was less immunogenic than RBD, possibly due to extensive glycosylation (Chi et al., 2020; Piccoli et al., 2020; Rogers et al., 2020; Sun et al., 2020a; Walls et al., 2020b; GSK, 2021). Potent human neutralizing antibodies were also isolated against the NTD “i” supersite, displaying similar potency to RBD-directed mAbs (McCallum et al., 2021). More recent data indicate that the convalescent plasma IgG response to SARS-CoV-2 is directed overwhelmingly toward non-RBD epitopes, including potently neutralizing antibodies against the NTD (Voss et al., 2021). Importantly, given the constant selective pressure on the RBD and the NTD domains by the immune system, SARS-CoV-2 variants of concern emerged, harboring frequent mutations in these two sites, as described below.
The functionality of potent neutralizing mAbs was demonstrated in animals, primarily rodents and nonhuman primates (NHPs), which were protected from SARS-CoV-2 infection after mAb injection (Baum et al., 2020a; Hansen et al., 2020; Rogers et al., 2020; Tortorici et al., 2020; Zost et al., 2020; Jones et al., 2021). Importantly, a strong correlation between neutralizing antibodies and reduction of viral load was determined in NHPs (Guebre-Xabier et al., 2020; Mercado et al., 2020; van Doremalen et al., 2020; Zost et al., 2020; Jones et al., 2021). Aside from mechanistically blocking viral cell entry, animal studies indicated that effector functions mediated by the antibody Fc domain (i.e., antibody-dependent cellular cytotoxicity, phagocytosis, and cell recruitment) contributed to optimal protection against SARS-CoV-2 (Schafer et al., 2021; Winkler et al., 2021).
In humans, transfusion of hyperimmune plasma with high titers of neutralizing antibodies from convalescent persons to hospitalized patients was initially considered for early COVID-19 therapy (Liu et al., 2020a). Several studies have reported some level of benefit for plasma therapy; however, due to the complexity and risks linked to this practice, attention was shifted to the isolation and use of human mAbs (Renn et al., 2020; Katz, 2021). The use of mAbs demonstrated efficacy in preventing disease progression of outpatients with COVID-19 and enhancing viral clearance (Celltrion, 2021; Chen et al., 2021a; Weinreich et al., 2021) (Eom et al., preprint, https://www.researchsquare.com/article/rs-296518/v1). This evidence supported the Emergency Use Authorization (EUA) of bamlanivimab (AbCellera/Eli Lilly; combination of LY-CoV555 with etesevimab LY-CoV016) and REGN-COV2 (Regeneron; a combination of two mAbs, casirivimab REGN10933 and imdevimab REGN10987) by the U.S. Food and Drug Administration (FDA), and the approval of regdanvimab (Celltrion; monotherapy with CT-P59 mAb) by the European Medicines Agency; several other mAbs are in clinical development (Taylor et al., 2021). Sotrovimab (Vir Biotechnology/GSK; VIR-7831) is a mAb isolated from a 2003 SARS-CoV-1 survivor, targeting the RBD glycan N343 that is highly conserved among Sarbecoviruses. N343 is distinct from the receptor-binding motif, thus not competing for ACE2 binding in solution (Pinto et al., 2020) (Cathcart et al., preprint, https://www.biorxiv.org/content/10.1101/2021.03.09.434607v1). VIR-7831 has received EUA (U.S. Food and Drug Administration, 2021a) after the evidence of profound efficacy (85% reduction in hospitalization or death) in phase 3 clinical trial (GSK, 2021). Studies are now expanding to determine whether mAbs could be used to prevent SARS-CoV-2 infection and as an alternative to vaccination for people who cannot take vaccines, are immunodeficient, need more immediate prophylaxis, or as a rapid postexposure treatment (Cohen, 2021b).
After the extensive analysis of neutralizing immunity to the S protein, studies evaluating the antibody response to other SARS-CoV-2 antigens were conducted but did not provide compelling evidence of neutralizing immunity.
The N antigen is a highly abundant and immunogenic protein expressed during viral replication, which participates in RNA transcription, amplification, and packaging into virions (Chang et al., 2006; Sariol and Perlman, 2020). It is the target of both antibodies and T cells, responsible for virus clearance in SARS-CoV-1 infections in mice (Zhao et al., 2010) and of SARS-CoV-2-specific CD8+ and CD4+ T cells in humans (Grifoni et al., 2020). Prior SARS-CoV-1 immunogenicity studies demonstrated that active immunization could elicit both CD4+ and CD8+ T cell responses in mice (Liu et al., 2006). However, N-specific immune serum did not protect mice against SARS-CoV-2 infection (Sun et al., 2020b). Thus, the N protein is regarded as a target for viral clearance.
The M and E proteins are required for virus assembly (Schoeman and Fielding, 2019). Due to their small ectodomain, they are poorly immunogenic (Du et al., 2008). In mice, sera elicited against M or E protein did not protect against SARS-CoV-2 infection (Sun et al., 2020b).
B. T Cell Responses
T cell responses are often neglected when studying viruses as it is generally assumed antibodies are necessary for virus neutralization. However, they play an important role for B cell maturation, antibody production, and viral clearance. In COVID-19 patients, virus-specific CD4+ and CD8+ T cell responses were associated with mild disease, suggesting the potential of cellular immunity in protecting against SARS-CoV-2 (Peng et al., 2020; Rydyznski Moderbacher et al., 2020; Tan et al., 2021). Indeed, an ideal vaccine would be expected to elicit both humoral and cellular immune responses. Using peptide pools, several studies showed that CD4+ T cell responses were primarily directed to M, N, and S proteins and, to a lesser extent, to nonstructural viral proteins, suggesting a broad reactivity against multiple viral targets (Grifoni et al., 2020; Le Bert et al., 2020; Peng et al., 2020; Nelde et al., 2021). Low and colleagues reported that 34% T cell clones from 34 COVID-19-recovered individuals recognized a conserved immunodominant region within the RBD (Low et al., 2021). Virus-specific CD4+ T cells were also present in 40%–60% healthy controls, reflecting a certain degree of crossreactivity with pre-existing coronavirus immunity (Grifoni et al., 2020; Le Bert et al., 2020; Nelde et al., 2021). The S protein was a main target of CD8+ T cell responses (Grifoni et al., 2020). These findings were in line with another study which detected robust virus-specific T cells activated in acute COVID-19 and in 28% of unexposed healthy donors (Sekine et al., 2020). Crossreactivity to seasonal coronavirus and other viruses could affect vaccination outcomes, leading to a faster or better immune response, or might be detrimental, through mechanisms of dampening of the de novo immune response (“original antigenic sin”) or via ADE of disease, and require to be further investigated (Sette and Crotty, 2020).
IV. Vaccine-Elicited Immunity
A. Overview on SARS-CoV-2 Vaccines
After the global spread of SARS-CoV-2, it quickly became apparent that stopping the pandemic and achieving herd immunity could only be accomplished via vaccination. Employing technologies and lessons learned from other infectious diseases such as HIV, influenza, RSV, and Ebola, a globe-spanning effort was launched to develop efficacious SARS-CoV-2 vaccines with overlapping—rather than sequentially—nonclinical and clinical studies that ensured fast development without compromising on safety and quality (Kelley, 2020; Rappuoli et al., 2021). Considering the scale of the pandemic, researchers agreed that several vaccines would need to be developed in parallel to facilitate the production and distribution of billions of vaccine doses. To address this significant challenge, diverse vaccine types, including inactivated/attenuated virus vaccines, nucleic acid-based vaccines, nonreplicating vector, and recombinant subunit vaccines, underwent rapid development. The general description of these different approaches is detailed in several reviews (Dong et al., 2020; Krammer, 2020; Dai and Gao, 2021). In the following sections, we will focus on the immunogenicity and efficacy against COVID-19 of the most advanced SARS-CoV-2 vaccines (approved for use or in phase 3 clinical trials).
The development of these vaccines was incredibly rapid, with some being approved by regulators for emergency use in less than one calendar year from the onset of the pandemic, thanks to unprecedented levels of international collaborations, funding, data sharing, and use of synthetic gene expression technologies. One of the key accelerators was the complete and open access to the viral genome sequences that facilitated viral spread monitoring and the design of diagnostic tests and vaccine candidates. The development of vaccines and treatments for other coronaviruses provided the basis for the development of SARS-CoV-2 vaccines (Park et al., 2020; Sariol and Perlman, 2020). However, prior to the COVID-19 pandemic, no coronavirus vaccine was approved for human use, with the most advanced SARS-CoV-1 and MERS-CoV candidates still in phase 1 clinical trial (NCT00099463, NCT03399578, NCT04170829) (Li et al., 2020b; Tizard, 2020). Nonetheless, instrumental to the success of new SARS-CoV-2 vaccines were previous findings from structural vaccinology studies that identified key amino acid substitutions for stabilizing the RSV viral fusion protein in its prefusion conformation, which is considered highly important for the induction of functional neutralizing antibodies (McLellan et al., 2013a; McLellan et al., 2013b). Structure-based design was previously used to stabilize the MERS-CoV spike protein as well, which elicited more potent antibody responses (Pallesen et al., 2017). Learnings from these studies were eventually applied to the SARS-CoV-2 S protein, resulting in a S-2P antigen bearing two proline substitutions that stabilize the protein in the prefusion conformation, the antigen of choice for some of the most advanced vaccines (Wrapp et al., 2020). In addition to structural vaccinology approaches, reverse vaccinology, synthetic biology, and platform technologies were also instrumental in the accelerated development of SARS-CoV-2 vaccines (Rappuoli et al., 2021). A considerable amount of research was focused on the design of gene-based vaccines. In oncology, research on synthetic gene delivery contributed for decades to the development of therapies allowing gene transfer to repair nonfunctional alleles (Anguela and High, 2019). In the infectious disease space, transfer of DNA encoding viral antigen and viral RNA delivery via lipid nanoparticles were previously developed for vaccination (Martinon et al., 1993; Ulmer et al., 1993). The synthetic gene delivery approach obviates the need for expressing large quantities of viral proteins required for immunization; however, this had never been tested beyond phase 2 and never on a global population level. Evidence on gene-based vaccines in humans was accumulated for the first time during the COVID-19 pandemic.
B. Immunogenicity and Efficacy of the Different Vaccine Platforms Targeting Spike Protein
Even though most vaccines incorporate the viral S protein as the primary target of the immune system, the use of different technologies resulted in marked differences in terms of level of spike-specific immunity and vaccine efficacy against COVID-19 (summarized in Table 1).
Characteristics, immunogenicity, and efficacy of the most advanced SARS-CoV-2 vaccines
For each vaccine, different methods were used to assess immunogenicity and reactogenicity; the data reported in the table are for reference only and not for comparison. Vaccines clinical trial data were retrieved from the World Health Organization COVID-19 vaccine tracker (http://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines). Vaccine approval status was retrieved from regulatory agencies websites and McGill University’s COVID-19 vaccine tracker (http://covid19.trackvaccines.org/vaccines/). Where available, vaccine efficacy estimates assessed in certain countries or against a specific variant are provided.
1. Inactivated and Attenuated SARS-CoV-2 Vaccines
Virus inactivation is an established and relatively easy approach for producing stable vaccines, which has contributed to the near eradication of polio and prevention of other diseases. A drawback for inactivated vaccines against COVID-19 is that biosafety level 3 containment facilities are required to grow the live virus (World Health Organization, 2020a), which limits manufacturing capability.
To face the COVID-19 pandemic, vaccines based on inactivated SARS-CoV-2 virus were readily developed and BBIBP-CorV (Sinopharm) and CoronaVac (Sinovac Biotech) were approved for human use. The immunogenicity of BBIBP-CorV was first tested in animals including rodents and NHPs. Two 2 μg/dose immunizations with BBIBP-CorV provided protection against SARS-CoV-2 challenge in NHPs (Wang et al., 2020b). In humans, BBIBP-CorV was safe and well tolerated in a phase 1/2 study conducted in China, with mild or moderate adverse reactions in 23% of participants (76/336 in phase 2) and no reported serious adverse events (Xia et al., 2021). Vaccine-elicited neutralizing antibody titers were highest with two 4 μg doses set 21 days apart, with a geometric mean titer (GMT) of 282.7 at 28 days postsecond dose. Convalescent neutralizing levels were not reported for comparison. CoronaVac had a similar safety profile in phase 1/2 clinical trials, with detection of any adverse reactions in 33% of participants (40/120 in phase 2) for the 0, 14-day 3 μg/dose schedule (Zhang et al., 2021b). The GMT of neutralizing antibodies for this cohort was 27.6, compared with 163.7 in convalescent asymptomatic patients. To this date, vaccine efficacy in phase 3 trials in China, United Arab Emirates, Turkey, and Brazil was reported to be between 50% and 91% for BBIBP-CorV and CoronaVac (Kim et al., 2021; Tanriover et al., 2021).
BBV152 (Covaxin, Bharat Biotech) was evaluated in a phase 2 trial in adults and adolescents 12–65 years of age in India receiving 3 and 6 μg doses administered 28 days apart. The first results of the study indicate that at four weeks postvaccination, GMTs of neutralizing antibodies were higher in the 6 μg group and comparable to those in convalescent patients. Both formulations were well tolerated, with around 40% of participants experiencing solicited adverse events and no serious adverse events reported (Ella et al., 2021). Another inactivated vaccine, PiCoVacc (Sinovac Biotech) was immunogenic and safe in animal models and showed protection against SARS-CoV-2 challenge in macaques (Gao et al., 2020).
Live-attenuated SARS-CoV-2 vaccines were evaluated in preclinical studies. YF-SO, composed of a yellow fever virus 17D backbone expressing SARS-CoV-2 spike protein, induced protective immunity in hamsters and macaques after one dose (Sanchez-Felipe et al., 2021).
Traditionally, inactivated vaccines are stable at refrigerator temperature (2–8°C) and affordable, making them particularly suitable for developing countries, and they are therefore the target of a considerable part of the SARS-CoV-2 vaccine effort. However, weakly neutralizing titers and Th2-skewed responses associated with inactivated vaccines warrant further investigation and close follow-up in case of emergence of viral variants (Jeyanathan et al., 2020).
2. Viral Vector SARS-CoV-2 Vaccines
Multiple replication incompetent vector vaccines carrying the SARS-CoV-2 S protein have been licensed in several countries for human use. Vectored vaccines attract the immune system’s attention to the viral protein without the need of adjuvants. However, pre-existing and newly elicited vector specific immunity can result in the blunting of immunogenicity, thus, lowering the efficacy of the vaccine. To address this, researchers have employed modified vectors from viruses less common in humans or adapted from other species. Adenoviruses have been studied for almost two decades as vehicles for HIV, malaria, influenza, Ebola, and tuberculosis antigens (Zhang and Zhou, 2016).
CanSino Biologics and Beijing Institute of Biotechnology developed an adenovirus type 5 vectored SARS-CoV-2 vaccine (Ad5-nCov; Convidecia). In phase 1/2 studies conducted in China, the vaccine elicited neutralizing antibodies (GMT of 18.3) and specific T cell responses with a single-dose regimen of 5 x 1010 viral particles. For this regimen, solicited adverse reactions were reported by 74% participants (96/129 in phase 2) (Zhu et al., 2020a; Zhu et al., 2020b). Based on an interim analysis of the phase 3 trial, vaccine efficacy was reported in a press release to be 65.3% at preventing symptomatic COVID-19 (CanSino, 2021).
A similar strategy, with a recombinant adenovirus type 26 encoding a full-length prefusion stabilized SARS-CoV-2 S protein (Ad26.COV2.S) was employed by Janssen/Johnson & Johnson. This is the only vaccine currently employed that is administered as a single dose. Ad26 vector was previously used to deliver the Ebola vaccine Ad26.ZEBOV approved for human use, in combination with a modified vaccinia Ankara vector (Pollard et al., 2021). Immunization with Ad26.COV2.S induced robust neutralizing antibody responses and provided complete protection from SARS-CoV-2 challenge in NHPs, with almost no detectable virus in the lungs and nose (Mercado et al., 2020). In humans, a 5 × 1010 viral particles dose resulted in a neutralizing antibody GMT of 288, which increased to 827 after a boost at eight weeks (Sadoff et al., 2021b). In phase 3 trials, worldwide efficacy was 66.1% against moderate to severe COVID-19 (at ≥28 days postvaccination), with vaccinees reporting primarily local reactions, whereas serious adverse events occurred in 0.4% of vaccinated individuals, comparable to the placebo group (Sadoff et al., 2021a).
In contrast, a heterologous prime-boost approach of recombinant adenovirus type 26 (rAd26-S) and type 5 (rAd5-S), both carrying the SARS-CoV-2 S protein gene, was developed by Gamaleya Research Institute of Moscow (Gam-COVID-Vac, Sputnik-V). In phase 1/2 studies, heterologous prime-boosting with Gam-COVID-Vac elicited strong humoral responses with neutralizing antibody titers of 49.25, which was comparable to titers measured in convalescent plasma. The vaccine also induced antigen-specific CD4+ and CD8+ T cells cellular immune responses (Logunov et al., 2020). The efficacy against polymerase chain reaction (PCR)-confirmed COVID-19 from three weeks after receiving the first vaccine dose was reported to be 91.6% in an interim analysis of phase 3 data (Logunov et al., 2021).
Simian adenovirus was used to develop the ChAdOx1 vaccine (AZD1222, Covishield/Vaxzevria) by Oxford University and AstraZeneca. The ChAdOx1 vector was previously used to develop, among other candidates, a malaria vaccine that reached phase 1/2 clinical testing (NCT03203421). Optimal dosing for the COVID-19 vaccine has been the subject of extensive investigations. In a phase 1/2 trial, median neutralizing titers were higher after the second dose (87.9 after one dose, 162.9 after two doses), and similar to the ones observed in convalescent plasma from patients recovered from COVID-19 (Folegatti et al., 2020). A second dose of ChAdOx1 correlated with enhanced Fc-mediated functional antibody responses (Barrett et al., 2021). In the interim analysis of the pivotal phase 3 clinical trial conducted in the United Kingdom, Brazil, and South Africa, vaccine efficacy against laboratory-confirmed, symptomatic COVID-19 more than 14 days after the second dose was 62.1% in participants who received two standard doses and was 90% when a half-dose was followed by a full dose after at least one month. Overall vaccine efficacy across both groups was 70.4% (Voysey et al., 2021a). A subsequent pooled analysis from the same trial suggested that allowing longer time between doses, a feature of the latter subgroup, was more important than lowering the priming dose (Voysey et al., 2021b). Indeed, since its EUA, vaccine administration was conducted 4–12 weeks apart, with the World Health Organization (WHO) recommended interval of 8–12 weeks (World Health Organization, 2021b). Longer prime-boost intervals increased spike-specific IgG levels and vaccine effectiveness, reaching 81.3% in case of an interval of 12 weeks or more versus 55.1% for an interval of less than six weeks (Voysey et al., 2021b). Among controversies on data analysis of a subsequent phase 3 trial in the US, Peru, and Chile, which included two doses administered four weeks apart, preliminary assessment marked the efficacy at 76% at preventing PCR-confirmed, symptomatic COVID-19 and 100% at preventing severe disease. Notably, efficacy was higher in older individuals, 65 and over (AstraZeneca, 2021). Even though no major safety concerns were raised during clinical development, adenovirus-based vaccines were associated with very rare thrombotic thrombocytopenia events, resulting in public concern (Ledford, 2021). From an immunologic standpoint, these rare events could be associated to development of antiplatelet factor 4 antibodies, although further research is required to fully understand this phenomenon (Greinacher et al., 2021).
Replication competent viruses have been tested in preclinical studies but had not reached clinical stage yet. A vesicular stomatitis virus vaccine expressing a modified form of the S protein (VSV-eGFP-SARS-CoV-2) elicited high antibody titers and reduced viral infection and lung inflammation after challenge in mice, indicating protection against COVID-19 (Case et al., 2020). Similarly, the vesicular stomatitis virus-vectored vaccine ConVac expressing SARS-CoV-2 S1 subunit demonstrated protection against severe COVID-19 in hamsters (Malherbe et al., 2021).
3. Nucleic Acid SARS-CoV-2 Vaccines
Vaccines based on mRNA have shown great promise and were the first to be authorized for human use by the FDA and European Medicines Agency. Extremely fast in vitro production of these vaccines was essential to achieve the rapid scale-up that brought them to the clinics in less than 6 months. Moreover, recent advances in RNA nucleoside modification, technological improvement for in vivo delivery, and spike-stabilizing mutations, described before, have greatly enhanced their immunogenicity (Chaudhary et al., 2021). Particularly, engineering of the RNA sequence, including codon optimization, capping and tail regulatory sequences modification, and the use of immuno-silent nucleosides such as pseudouridine and 1-methylpseudouridine, greatly enhanced stability and level of protein expression (Pardi et al., 2018). Vaccines developed by Moderna (mRNA-1273; Moderna COVID-19 vaccine) and BioNTech/Pfizer (BNT162b2; Comirnaty), based on sequence optimized nucleoside-modified mRNAs formulated in lipid nanoparticles, received EUA in December 2020. BNT162b2 became the first FDA-approved vaccine for the prevention of COVID-19 disease in individuals 16 years of age and older (Food and Drug Administration, 2021b). In the case of mRNA-1273, the EUA allows vaccination of individuals 18 years of age and older.
Moderna’s mRNA-1273, jointly developed with the National Institutes of Health, was first demonstrated to be immunogenic and protective in NHPs, which received two doses of 10 or 100 μg four weeks apart (Corbett et al., 2020). In a phase 1 study, participants were immunized with either 25, 100, or 250 μg mRNA-1273 at 28 days apart (Jackson et al., 2020). Serum neutralizing titers were highest after two 100 μg doses (GMT of 654.3), and in the upper range of the neutralization response observed in convalescent samples. T cell responses were biased toward Th1 cytokines. In a follow-up study, vaccine immunogenicity in the elderly was assessed, with high neutralizing antibody titers detected in both the age groups 56–70 and 71 and older (GMTs of 402 and 317, respectively) (Anderson et al., 2020). Phase 3 data from a trial conducted in the United States and using the 100 μg dosage showed a vaccine efficacy of 94.1% against COVID-19 illness with onset at least 14 days after the second vaccine dose (Baden et al., 2021).
During the development of the Pfizer/BioNTech vaccine, two sets of mRNA vaccines coding either for the RBD-only domain (BNT162b1) or full-length S protein (BNT162b2) were initially evaluated. Findings from phase 1 studies showed that two consecutive 30 μg doses of BNT162b1/2 elicited high SARS-CoV-2 neutralizing antibody titers and robust levels of antigen-specific CD8+ and Th1-type CD4+ T-cell responses (Mulligan et al., 2020; Sahin et al., 2020; Walsh et al., 2020). Both vaccines elicited dose-dependent SARS-CoV-2 neutralizing antibody titers (GMTs of 437 for BNT162b1 and 163 for BNT162b2 at 14 days postsecond dose for the 30 μg doses, in participants 18–55 years of age), although BNT162b2 was less reactogenic, probably due to a lower number of mRNA copies in the formulation. These neutralizing antibody GMTs were 1.7 to 4.6 times higher than the average neutralizing response of convalescent serum. Analysis of phase 3 clinical trials showed a vaccine efficacy of 95.0% against laboratory-confirmed COVID-19 with onset at least 7 days after the second dose for BNT162b2 (Polack et al., 2020). In a subsequent clinical trial designed to assess protection of adolescents 12–15 years of age, which are particularly exposed to the virus due to in-person learning and socialization, vaccine efficacy was 100% (Frenck et al., 2021).
After the beginning of the mass vaccination campaign in Israel, real-world vaccine effectiveness of 97.0% against symptomatic COVID-19 was observed between January 24 and April 3, 2021 (Haas et al., 2021). In the United States, vaccine effectiveness was reported to be 88.8% and 96.3% for BNT162b2 and mRNA-1273, respectively (Pilishvili et al., 2021).
A third mRNA vaccine, CVnCoV (CureVac), composed of nonmodified codon-optimized mRNA encoding for prefusion stabilized S protein, was recently assessed in a pivotal phase 2b/3 trial after positive phase 1 data demonstrating induction of high neutralizing titers with two 12 μg vaccine doses administered 28 days apart (Kremsner et al., 2021). However, the phase 2b/3 study reported a vaccine efficacy of 47% against COVID-19 of any severity across all age groups, markedly lower than the other mRNA vaccines (CureVac, 2021). This study was conducted in the context of a high prevalence of SARS-CoV-2 variant strains and an almost complete absence of the wild-type virus (see section V.B).
Despite being the product of a relatively new technology that had never before been tested in a large population, all mRNA COVID-19 vaccines demonstrated a high safety profile both in clinical trials and in real-world settings. After a mass vaccination campaign in more than 2.4 million people, BNT162b2 vaccination was reported not to be associated with an elevated risk of most of the adverse events examined, except for a possible excess risk of myocarditis (1–5 events per 100,000 persons) (Barda et al., 2021). The latter evidence calls for the need of further studies to better estimate long-term effects of this class of vaccines.
DNA vaccines are also being developed against SARS-CoV-2, although not yet approved for human use. DNA vaccines need to be delivered into cell nuclei, so they require particular type of injection devices. INO-4800 (Inovio Pharmaceuticals), AG0301 (AnGes and Takara Bio), and ZyCoV-D (Zydus Cadila) are currently in phase 2/3 trials. In phase 1, INO-4800 elicited neutralizing antibodies in 78% and 84% of participants when electroporated at two 1 mg and 2 mg doses, respectively; GMTs were 102.3 and 63.5 for the respective groups (Tebas et al., 2021).
4. Recombinant Protein SARS-CoV-2 Vaccines
Recombinant spike-protein-based vaccines, recombinant RBD-based vaccines and viruslike particle (VLP)-based vaccines are currently under clinical investigation. As previously mentioned, the development process for these vaccines is more complex and time consuming, but well-known adjuvants can be used to boost the immune response. Adjuvants are vaccine components able to enhance magnitude, breadth, and durability of the immune response (Del Giudice et al., 2018; Pulendran et al., 2021).
NVX-CoV2373 (Covovax, Novavax) is a protein-based vaccine candidate composed of the SARS-CoV-2 S protein and the saponin-based Matrix-M adjuvant. After intranasal and intratracheal challenge with SARS-CoV-2, macaques were protected against upper and lower infection and pulmonary disease when immunized with two 5 or 25 μg doses of NVX-CoV2373 (Guebre-Xabier et al., 2020). In phase 1/2 trials, NVX-CoV2373 elicited immune responses that exceed levels in COVID-19 convalescent serum (Keech et al., 2020). Two 5 μg vaccine doses induced a neutralizing antibody GMT of 3906, four times higher than the one detected in symptomatic COVID-19 outpatients (837) and comparable to levels observed in hospitalized patients (7457) (Keech et al., 2020). In this clinical study, Matrix-M1-adjuvanted regimens induced antigen-specific T cell responses with a strong bias toward Th1 phenotype. Mild reactogenicity to the vaccine, more pronounced at the second dose, was consistent with previous experience with adjuvanted vaccines.
Other recombinant protein vaccine candidates are being developed. The SARS-CoV-2 prefusion stabilized S protein vaccine CoVLP (Medicago/GSK), expressed as VLPs produced in plant cells (Nicotiana benthamiana), elicited neutralizing antibody titers comparable to or 10 times higher than COVID-19 convalescents when adjuvanted with CpG1018 or AS03, respectively, and it is currently in phase 2/3 trial (NCT04636697) (Ward et al., 2021). Another protein-based vaccine, developed by Sanofi/GSK and containing AS03-adjuvanted soluble S-2P trimer, showed preliminary positive results in NHPs using a refined formulation after the first one elicited an insufficient immune response in older adults, likely due to an insufficient concentration of the antigen (Francica et al., 2021; Goepfert et al., 2021). In a phase 2 trial, strong rates of neutralizing antibody response in all age groups (18–95 years old) were reported for all two-dose regimens used (antigen dose levels of 5, 10, and 15 µg) (Sanofi, 2021). This vaccine candidate is currently in phase 3 trial (VAT00008/NCT04904549). Lastly, SCB-2019 native trimeric S protein (Clover Biopharmaceuticals) was tested at three different dosages (3, 9, and 30 μg), administered as two doses 21 days apart using AS03, CpG/Alum, or no adjuvant formulations. Neutralizing antibody GMTs were 3948 and 1050 for the AS03 and the CpG/Alum groups at 36 days after the second 30 μg dose and, respectively, 1522 and 1159 at 50 days in 18–54 years of age (Richmond et al., 2021). The frequency of local and general adverse events was higher in the AS03 group (≤75% and ≤62.5%) compared with CpG/Alum (≤62.5% and ≤37.5% after each dose). Based on these results, a phase 2/3 clinical trial with a 40 μg dose plus CpG/Alum adjuvant was conducted, reporting an efficacy of 79% against COVID-19 and 100% against severe disease, in a context where all sequenced strains were variants, mostly (73%) Delta (Clover, 2021) (see section V.B).
Self-assembling protein nanoparticles, which have been shown previously to significantly improve potency and breadth of antibody response (Brouwer et al., 2019; Marcandalli et al., 2019), have also been used as templates for the design of SARS-CoV-2 vaccines. The two-component I53-50 nanoparticle, coming from studies in the HIV field, was used to display 60 RBDs in an array that elicited antibody titers 10-fold higher than the prefusion-stabilized S protein despite a lower dose (Walls et al., 2020a).
C. Vaccines Targeting Other SARS-CoV-2 Antigens
Vaccines against SARS-CoV-2 have been mainly focused on the viral S protein due to the strong correlation between neutralization of viral entry and protection from disease. Due to low immunogenicity, the M and E proteins have not been identified as targets of vaccines. The highly immunogenic N protein was also not exploited as primary vaccine candidate. Th2-biased response and enhanced infection-induced pneumonia were observed in previous SARS-CoV-1 studies in N protein-immunized mice (Yasui et al., 2008). Moreover, N protein-directed antibodies were associated with COVID-19 immunopathology. High levels of IgG against N protein were associated with higher risk of admission to intensive care units (Batra et al., 2021). As reported in this study, the proposed mechanism behind this phenomenon is the generation of immune complexes on infected cells expressing N protein, resulting in proinflammatory cytokines and inflammation.
V. SARS-CoV-2 Notable Variants
A. Emergence of SARS-CoV-2 Variants
The first generation of COVID-19 vaccines was designed with the spike protein of the SARS-CoV-2 strain circulating during the early phases of the pandemic in 2020, Wuhan-Hu-1. During the course of the pandemic, several mutated lineages of SARS-CoV-2 emerged and abruptly spread globally (Fig. 4) (PANGO, 2021; World Health Organization, 2021c). A summary of terminology used to describe SARS-CoV-2 variants is reported in Fig. 5.

Origin and timeline of appearance for notable SARS-CoV-2 variants. The origin of viral variants across the world and the date of their earliest documentation during the COVID-19 pandemic (World Health Organization, 2021c).

Terminology of SARS-CoV-2 variants. Brief description of the terminology used in the manuscript referred to SARS-CoV-2 virus (Rambaut et al., 2020; Mascola et al., 2021).
Compared with other RNA viruses like influenza or HIV, SARS-CoV-2 genome variation is slow to occur, probably because of a proofreading enzyme that corrects copying mistakes. Nonetheless, mutations arise at a rate of roughly 2 point mutations per month, and new variants can emerge via recombination between strains from different host species, including humans (Singh and Yi, 2021). Together with selective pressure by the immune system, this has given rise to numerous viral variants with concerning altered transmissibility that promptly activated monitoring of impact on immunity and disease severity (Koyama et al., 2020). Importantly, mutation speed does not change in the new variants, but the growing number of infected people increases the likelihood of even more variants to appear. In this paragraph, we report evidence related to notable variants that carry mutations implicated in immune evasion.
Amino acid substitution D614G in S1 subunit of the S protein of the original Wuhan-Hu-1 SARS-CoV-2 strain was the first to emerge in late January or early February 2020 and became the dominant virus strain circulating globally by June 2020. Studies in human respiratory cells and animal models demonstrated that, compared with the original strain, D614G substitution enhanced virus transmission due to increased S protein shift toward a more ACE2-binding competent state (Korber et al., 2020; Yurkovetskiy et al., 2020; Ozono et al., 2021). Subsequently, in September 2020, a SARS-CoV-2 variant linked to infection in farmed mink was identified in Denmark, referred to as “Cluster 5” (World Health Organization, 2020b). There was concern that this variant could result in reduced virus neutralization in humans, but in the end it did not spread widely.
Starting with September 2020, in the United Kingdom, a cluster of viruses phylogenetically separated from the original viral strain was identified during continuous epidemiologic surveillance (Public Health England, 2020). This strain gave a negative result on the PCR targeting spike protein in a three-target test (S, ORF1ab, and N), suggesting the presence of multiple mutations in the viral receptor. This variant was initially termed “variant under investigation” VUI-202012/01 and subsequently became “variant of concern” (VOC) 202012/01 or B.1.1.7 (WHO label Alpha) when it was demonstrated to spread more rapidly in the population, although no evidence of increased disease severity has been reported (Public Health England, 2020). Alpha spread quickly in the United Kingdom with a modeled increased transmissibility of 53%–56% (Davies et al., 2021; Volz et al., 2021) and being 43%–90% more transmissible than the previous lineage (Davies et al., 2021). Genome sequencing revealed a total of 23 mutations, including amino acid substitutions and deletion in the spike RBD domain (Fig. 6). Three mutations, N501Y, P681H, and H69-V70 deletion were linked to increased ACE2 binding and potential escape from neutralizing antibodies. Particularly, N501Y was demonstrated to increase in vitro binding to ACE2-expressing cells (Starr et al., 2020).

SARS-CoV-2 spike mutations in notable viral variants. (A) SARS-CoV-2 spike trimer and biologically relevant mutations, modeled on PDB 7C2L. One protomer is highlighted, showing the NTD in green and RBD in two shades of blue. Residues with dotted lines are hidden in this view. (B) Summary of SARS-CoV-2 variant-defining mutations and major biologic effects.
In December 2020, national authorities in South Africa announced the spread of a new variant, named 501Y.V2 or B.1.351, and subsequently labeled as Beta by the WHO, which became dominant in the region and was eventually classified as VOC (Fig. 4) (Tegally et al., 2021a). During continuous genomic surveillance, several other SARS-CoV-2 variants were reported in South Africa (Tegally et al., 2021b). The Beta variant is characterized by eight lineage-defining mutations in the spike protein, including three substitutions of functional significance, K417N, E484K, and N501Y, as described below. Lineage P.1, also known as 20J/501Y.V3 or the Gamma variant (first documented in November 2020), arose in Brazil in January 2021, causing widespread infection in Manaus, despite high seroprevalence (76%) against the wild-type strain in the population (Faria et al., 2021; Sabino et al., 2021). Gamma has 17 unique amino acid changes and shares with Beta the acquisition of N501, K417, and E484 mutations, being now classified as VOC (Fig. 4). A separate study identified the P.2 (Zeta) lineage, which also harbors the spike E484K mutation but not the N501Y and K417T mutations, which concomitantly appeared in several locations in Brazil but evolved independently from the Gamma lineage from Manaus (Naveca et al., 2021).
Since its earliest detection in October 2020, variant B.1.617.2 or Delta has rapidly spread from India, and in less than a year, it became the dominant SARS-CoV-2 variant in circulation worldwide, outcompeting pre-existing lineages including Alpha. The B.1.617 lineage was initially identified as carrying two mutations that might be related to increased transmissibility and immune evasion: E484Q (similar to E484K substitution in South African and Brazilian strain) and L452R (found in the California variant CAL.20C [B.1.427/1.429], WHO label Epsilon) (Fig. 4) (Ferreira et al., 2021; Pattabiraman et al., 2021). One important feature of Delta was the significant replication advantage observed in vitro in comparison with Alpha, both in human airway epithelial model and airway organoids (Mlcochova et al., 2021). Viral cell entry is dependent on the polybasic cleavage site between S1 and S2 (Peacock et al., 2021). The Alpha spike bearing the P681H mutation was reported to have significantly higher fusogenic potential than a D614G Wuhan virus, which resulted in increased spike-mediated cell entry (Meng et al., 2021). Similarly, mutation P681R in the B.1.617 spike was linked with a higher portion of cleaved spike compared with Alpha and increased in vitro replication fitness (Mlcochova et al., 2021). This results in the high observed infectivity of the Delta variant, with reported higher viral burden in infected individuals and resulting in a higher probability of person-to-person transmission (Li et al., 2021; Scientific Pandemic Influenza Group on Modeling Operational subgroup, 2021; Teyssou et al., 2021). Indeed, the Delta reproductive number (R0) was calculated to range between 3.2 and 8, which is up to three times more contagious than the original SARS-CoV-2 strain (R0 = 2.79) (Liu and Rocklov, 2021).
Other SARS-CoV-2 variants are currently being monitored. The Epsilon variant, first recorded in South California with the S protein mutation L452R, accounted for a significant fraction of samples collected in the region at the beginning of 2021, in a retrospective study (Zhang et al., 2021a), and is now virtually disappearing. The C.37 variant, also known as Lambda, was first detected in Peru in December 2020, bearing the same L452R mutation and, on the contrary, spread to many countries (Wink et al., 2021). One of the more recently emerging variants of COVID-19, Mu (also known as B.1.621), first detected in January 2021, quickly became the dominant strain in Colombia and spread to South America and Europe. The Mu variant contains several amino acid substitutions including E484K, N501Y, and P681H, which could confer a synergistic effect on vaccine resistance, transmissibility, and disease severity (Laiton-Donato et al., 2021). Even though these variants did not spread as largely as the VOCs, they are currently being monitored for their potential immune-evasive and infection-enhancing mutations.
After the identification of these variants, a main objective became the assessment of their ability to evade protective immune response. The expression of S protein mutations found in Alpha on viral vectors (pseudoviruses) resulted in decreased monoclonal antibody neutralization; however, only minor potency loss was observed for polyclonal sera (Rees-Spear et al., 2021). On the contrary, in vitro assays with pseudoviruses expressing the spike of the Beta variant, which contains substitutions in the two immunodominant domains RBD and NTD, revelated that the neutralizing capacity of antibodies from convalescent patients was diminished against this variant (Wibmer et al., 2021). Substitution K417N induced reduced binding and neutralization from representative class 1 mAbs (that bind RBD in “up” position) from South African donors due to loss of a key hydrogen bond. Similarly, representative class 2 antibodies failed to bind Beta RBD, due to a missing contact point. Moreover, deletions in the NTD domain abrogated binding of N5-loop-directed antibodies. These studies provided evidence that the Beta variant can evade antibodies targeting highly immunogenic antigenic sites in the spike protein. Tested against sera from convalescent patients, Beta variant mutations induced a 10- to 15-fold reduction in mean neutralization titer (Cele et al., 2021; Wibmer et al., 2021). On the contrary, the Alpha variant was relatively more resistant to NTD-directed than RBD-directed mAbs and showed only partial reduced neutralization by convalescent serum (one out of 20 patients) (Wang et al., 2021a). This relative resistance can thus be associated largely to the E484K mutation, shared by Beta and Gamma variants. In areas where these viruses were common, there was concern of reinfection due to a second wave of variant strains infecting a population seropositive for the wild-type virus, as reported in Manaus, Brazil, which could pose a significant safety risk (Sabino et al., 2021). Importantly, the appearance of a E484K mutation was reported in a small fraction of Alpha viruses (11 out of 214159 sequences), suggesting that the United Kingdom variant could potentially acquire this change (Public Health England, 2021). More recently, the Delta variant was shown to be 4- to 6-fold less sensitive than other variants to serum neutralizing antibodies from recovered individuals (Mlcochova et al., 2021; Planas et al., 2021). In vitro, viruses with Delta spike exhibited higher resistance to neutralization with mAbs, denoting that the rapid spread of Delta was associated with the potential to escape antibody-mediated protection (Mlcochova et al., 2021; Planas et al., 2021).
The immune escape of E484K-bearing viruses was observed also for mAbs used in the clinic. LY-CoV555 alone and in combination with CB6 was inactive against Beta and, similarly, the activity of REGN10933 alone was impaired (Wang et al., 2021a). Due to resistance of viral variants, in April 2021, FDA revoked its previously issued EUA for bamlanivimab alone, which remained approved as part of a combination therapy with etesevimab (Food and Drug Administration, 2021a). Decreased binding to spike proteins containing E484K, D614G, and N501Y mutations was also observed for other mAbs (Chen et al., 2021b). Additionally, S protein mutations N234Q, L452R (detected in Epsilon and Delta variants), A475V, and V483A were found to induce escape from mAbs (Li et al., 2020a). The reasons behind the origin of these antibody escape mutants could be twofold: the high level of SARS-CoV-2 transmission rates, likely resulting in higher chances of emergence of mutated viruses, and the large proportion of commonly shared public antibodies expressing VH3-53/66 and VH1-24 that induced selective pressure on key S protein RBD and NTD residues (Wibmer et al., 2021). Importantly, many mAbs under clinical development derived from the same public antibodies, raising concerns on the future efficacy of these treatments (Marovich et al., 2020).
B. Implications for Vaccines
The emergence of SARS-CoV-2 variants had important implications for vaccine development. The impact of SARS-CoV-2 variants was observed on the clinical trials conducted in countries where the variants emerged during the course of the study and prompted the vaccine developers to initiate extensive studies to understand the effect on vaccination campaigns (Abdool Karim and de Oliveira, 2021).
The first mutation to be recorded, D614G, was suggested not to be an obstacle to vaccine development due to enhanced vulnerability to neutralization (Weissman et al., 2021). On the contrary, neutralizing antibody activity of the two mRNA vaccines was reported to be lower against the Beta variant’s mutations. Using pseudovirus neutralization assays, reduction of neutralization by a factor of 6.4 was detected in serum obtained from mRNA-1273 vaccinees, whereas neutralization was reduced 0.8- to 9-fold in serum from BNT162b2 vaccinees (Chen et al., 2021b; Liu et al., 2021; Wu et al., 2021; Xie et al., 2021). Reduced binding to Beta-associated mutations was also determined for eight out of 12 (67%) serum samples from recipients of the Gam-COVID-Vac vaccine (Ikegame et al., 2021). Evidence from in vitro analyses resulted in only a minor effect of the Alpha mutations, highlighting again the escape effect of the K417N, E484K, and N501Y substitutions (Garcia-Beltran et al., 2021).
A decrease in efficacy was observed for viral vectored, recombinant protein, and mRNA vaccines tested during the insurgence of the SARS-CoV-2 variants. The vaccine efficacy of ChAdOx1 was 75% when determined before the emergence of SARS-CoV-2 variants. A preliminary analysis reported that the vaccine provided a similar, yet reduced, level of neutralization against the Alpha variant mutations expressed on pseudoviruses. The mean fold change of the Alpha variant versus wild-type was 3.2 and 1.9 for the first and second dose vaccine sera (Collier et al., 2021). However, in a phase 1/2 study in South Africa, protection against mild-to-moderate COVID-19 was 21.9% in the overall study population, and it was calculated to be only 10.4% against the Beta variant. Impact of this variant on hospitalization rate was not observed, likely due to the exclusion of 65 and older people from the trial (Madhi et al., 2021).
In a phase 2b trial conducted in South Africa, where approximately 90% of the cases were attributed to the Beta variant, a vaccine efficacy of 49.4% was reported for NVX-CoV2373 against PCR-confirmed, symptomatic COVID-19, or 60% if HIV-positive individuals in the trial population were removed from the analysis. On the contrary, interim analysis of a phase 3 trial in the United Kingdom, in which Alpha was detected in over half of the PCR-confirmed symptomatic cases, resulted in overall 89.3% estimated vaccine efficacy. Efficacy by strain was calculated to be 95.6% against the original SARS-CoV-2 strain and 85.6% against the United Kingdom variant (Callaway and Mallapaty, 2021; Novavax, 2021). Efficacy for a second recombinant protein vaccine, SCB-2019, was reported to be 79% in a context where 100% of SARS-CoV-2 strains were variants, with Delta as the predominant strain. Interestingly, in this randomized trial, SCB-2019 also showed a 64% reduced risk of reinfection in previously infected individuals (Clover, 2021).
For Ad26.COV2.S, a phase 3 study conducted in areas of high COVID-19 incidence and where variants were emerging demonstrated a global efficacy of 66.1% (at ≥28 days postadministration) against PCR-confirmed moderate to severe disease. In the United States, where the D614G strain is dominant, efficacy was 72%, whereas in Brazil, with a high prevalence of Zeta variant, efficacy was 68.1%. In South Africa, where almost all cases were caused by Beta, efficacy was 52.0% (Sadoff et al., 2021a).
For CVnCoV, a vaccine efficacy of 47% was reported against PCR-confirmed COVID-19 disease of any severity, in the context of high prevalence of SARS-CoV-2 variants (CureVac, 2021). In similar settings, other mRNA vaccines showed reduced effectiveness although maintaining a high degree of protection against COVID-19. For instance, in Qatar, an effectiveness of 87.0% and 100% was estimated against the Alpha variant and 72.1% and 96.4% against the Beta variant for BNT162b2 and mRNA-1273 (Abu-Raddad et al., 2021; Chemaitelly et al., 2021b). This apparent difference in vaccine performance could be due to the insufficient immunogenicity resulting from the use of unmodified mRNA bases [CVnCoV does not use uracil-modified mRNA, which is deemed to be critical in inducing high protein expression (Karikó et al., 2005)] or from the low dose used (12 µg compared with 100 µg for mRNA-1273 and 30 µg for BNT162b2). A next-generation vaccine with optimized noncoding regions, CV2CoV, also using nonmodified RNA bases, is currently being developed in collaboration with GSK. In NHPs, CV2CoV induced substantially higher immune responses compared with the first-generation CVnCoV, including more potent neutralization against SARS-CoV-2 variants (Gebre et al., 2021).
Reduction of vaccine effectiveness in a real-world setting was also reported after the rapid global spread of the Delta variant. In vitro, Delta was reported to be eight-fold less sensitive to vaccine-elicited antibodies as compared with the Wuhan D614G strain after two doses of ChAdOx1 or BNT162b2 vaccines (Mlcochova et al., 2021). Vaccine effectiveness against Delta was lower when compared with that against Alpha: 88% versus 93.7% and 67% versus 74.5%, after two doses of BNT162b2 and ChAdOx1, respectively, as reported in a test-negative case-control study in the UK (Lopez Bernal et al., 2021). In studies from Israel, Qatar, and the UK, BNT162b2 vaccine efficacy was observed to decrease with time, suggesting an increased risk of Delta variant infection months after vaccination (Chemaitelly et al., 2021a; Goldberg et al., 2021; Pouwels et al., 2021).
VI. Discussion
A. Considerations on Immune Response to Natural Infection and Vaccination
The induction of neutralizing antibodies is a key objective of vaccination strategies against viral targets. A detailed understanding of the human antibody response to SARS-CoV-2 is therefore critical to develop effective COVID-19 vaccines (Burton and Walker, 2020). Potent neutralizing antibodies have been identified by several research groups, most of which target the RBD domain of S protein and interfere with ACE2 receptor binding. In contrast to many other viral infections for which the accumulation of somatic hypermutation and clonal B cell selection is often observed in time, SARS-CoV-2 neutralizing antibodies isolated from convalescent patients showed limited accumulation of mutations (Kreer et al., 2020). This is likely due to a rapid viral clearance and the limited amount of available antigen, or the limited access to the germinal center (Kreer et al., 2020). Another explanation could be prior exposure to other human coronaviruses (Rydyznski Moderbacher et al., 2020). Importantly, the antibody response to RBD appears to be dominated by near-germline sequences coded by the VH3-53 gene, binding conserved residues such as K417 and E484 (Wu et al., 2020a; Yuan et al., 2020), which represent a pool of shared antibody responses among individuals. Despite their limited affinity maturation requirement to achieve a potent neutralization, these sequences could be ideal for vaccine design; unfortunately, the residues targeted by these public antibodies are the ones mutated in emerging escape viral variants, which significantly reduced the neutralization potency of sera from both convalescent and vaccinated individuals (Andreano and Rappuoli, 2021). With these considerations in mind, the use of immunogens that elicit other germline sequences less affected by viral escape could be a valuable strategy. In line with this, the selection of mAbs targeting highly conserved spike regions and the update of existing mAb cocktails could be envisioned to chase and stop escape mutants.
After the induction of a protective immune response, durability of immunity is a critical parameter to consider either after infection or vaccination. Immunity to SARS-CoV-2 infection was studied up to one year after infection. Antibodies against S protein and RBD declined moderately over time. Memory B cells increased between 1 and 8 months after infection, whereas memory CD4+ and CD8+ T cells declined with an initial half-life of 3–5 months (Dan et al., 2021). In another study, memory B cell levels were consistent with antigen persistence, suggesting robust activation of germinal centers, with a dominance of antibodies encoded by the VH3-53 gene (Gaebler et al., 2021). In case of active immunization, neutralizing antibodies have been reported to persist up to 6 months after a second dose of mRNA-1273 (Doria-Rose et al., 2021) and BNT162b2 (with a more rapid decline in the first 3 months from vaccination) (Levin et al., 2021; Naaber et al., 2021). Robust memory B cell responses were shown to last at least 6 months after mRNA vaccination (Goel et al., 2021b; Wang et al., 2021c). Strong recall memory was also demonstrated after one mRNA vaccine dose in individuals previously exposed to the virus (Goel et al., 2021a). These observations are crucial for recall responses after natural infection and vaccination. Indeed, vaccination of previously infected individuals has been shown to generate a strong humoral and cellular recall response, with evidence of extremely high neutralizing antibodies titers (Fig. 2B). Several research groups have reported that a single mRNA vaccine dose boosted infection-acquired immunity, with neutralizing antibodies up to a thousand times higher than the ones in virus-naïve individuals (Krammer et al., 2021; Prendecki et al., 2021; Stamatatos et al., 2021). Moreover, antibody titers were substantially boosted and compared with previous infection or vaccination alone (Reynolds et al., 2021). This boosting effect also involved cellular immune responses, with recall of S1 and S2 specific T cells (Prendecki et al., 2021). Importantly, the recall antibody response induced by mRNA vaccination in individuals that experienced SARS-CoV-2 infection was demonstrated to be crossneutralizing with the Beta variant, even at lower titers (Stamatatos et al., 2021). Interestingly, crossneutralizing antibody responses were due to anti-RBD antibodies whereas the NTD domain, which carries mutations/deletions in SARS-CoV-2 variants, did not appear to be a target of crossneutralizing antibodies. In another study, mRNA vaccination after SARS-CoV-2 infection increased the number of RBD-binding memory cells by more than an order of magnitude and recruited new B cell clones, expanding breadth of response (Wang et al., 2021b). Boosting of immune response with single-dose immunization and elicitation of crossprotective immunity has important implications for vaccines. This phenomenon, termed “hybrid immunity,” is proposed to derive from vaccine-induced activation of abundant memory B and T cells previously generated during natural infection (Crotty, 2021). Looking ahead, a combination of vaccine modalities and design of booster immunizations could be envisioned to achieve this effect also in virus-naïve individuals.
B. Considerations for Vaccines
Efforts to rapidly develop a first wave of COVID-19 vaccines focused on the elicitation of antibody responses to the spike protein, achieving high efficacy against the original SARS-CoV-2 virus (Hacisuleyman et al., 2021). However, mutations caused by either natural evolution or selective pressure by the immune system impacted the effectiveness of some vaccines (Karim, 2021).
The appearance of novel SARS-CoV-2 variants greatly complicated the measurement of vaccine efficacy in clinical trial, which saw rapidly changing scenarios when a new immune-resistant strain suddenly emerged. Despite good results in the United Kingdom, where ChAdOx1 exhibited a vaccine efficacy of 70%, the vaccine’s distribution was halted in South Africa due to a low efficacy of 22% as a result of a high prevalence of the Beta variant (Madhi et al., 2021). This opened question on the targets of the vaccine-elicited immunity. The extreme drop in neutralization titer against Beta, bearing three distinct mutations in the RBD region, suggested that much, though not all, of the ChAdOx1 vaccine-elicited neutralization was directed against the RBD (Madhi et al., 2021). Similarly, the neutralizing activity of the two mRNA vaccines against the Beta variant was reported to be 0.8–9 times lower, whereas a minor difference was evident against the Alpha variant. Other vaccines, NVX-CoV2373 and Ad26.COV2.S, registered decreased efficacy when tested in countries where Beta is present. Initial breakthrough cases of vaccine-elicited immunity were reported in the case of mRNA vaccination, when two fully vaccinated individuals developed COVID-19 caused by variant virus (Hacisuleyman et al., 2021). These findings underlined the importance of eliciting a diverse crossprotective antibody immunity via vaccination, which is critical to decrease the probability of viral escape from common antibody lineages elicited by first-generation vaccines. Wu and colleagues reported that 6 months after mRNA vaccination, all individuals retained potent serum neutralizing titer against wild-type SARS-CoV-2, but only 50% of them had measurable neutralizing titers against Beta and Gamma variants (Wu et al., 2021). Importantly, waning immunity was observed, with a significant decline in spike-specific antibody levels as early as 2 months after full vaccination (Shrotri et al., 2021). Vaccine efficacy estimated over 2-month periods was also reported to decline in time, from 96.2% (starting with 7 days after two BNT162b2 doses) to 83.7% (starting with 4 months postvaccination) (Thomas et al., 2021). A recent modeling study in Israel showed a significantly higher risk for infection for early vaccinees compared with those vaccinated later (Mizrahi et al., 2021). With the spread of Delta variant, which rapidly became the dominant SARS-CoV-2 strain, more breakthrough infections were reported even in countries with high vaccine coverage (Bergwerk et al., 2021). These events prompted the deployment of two strategies to enhance effectiveness against viral variants and boost immunity of the first wave of COVID-19 vaccines. The first one, homologous booster vaccination, was used to potentiate the two-dose mRNA vaccination schedule and counteract waning immunity. Higher neutralizing titers is thought to be sufficient to stop emerging variants such as Delta, against which serum neutralization capacity is decreased. After rapid implementation of booster vaccination in the population, a recent study from Israel showed that a booster dose administered to individuals aged ≥60 years at >5 months after the two-dose series lowered the rates of confirmed infection and severe illness by a factor of 11.3 and 19.5, respectively, compared with those not receiving a booster (Bar-On et al., 2021). Deployment of booster doses in at-risk people at least 6 months after full vaccination was indeed rapidly authorized by the FDA (Food and Drug Administration, 2021c). However, an important consideration is that boosting vaccinated individuals with currently available mRNA vaccines might produce strong responses that mirror their initial vaccine responses to wild-type virus but with potentially low coverage against variants, as observed by Cho and colleagues (Cho et al., 2021). The second strategy, heterologous vaccination, aims at developing a broader immune response, similarly to the hybrid immunity observed for vaccination of previously infected individuals. A booster dose with a modified mRNA vaccine designed to target the Beta variant is being developed and induced a broader crossreactive response (Choi et al., 2021). This evidence supports the development of second-generation vaccines tailored against emerging and future variants, exploiting the rapid development pace of mRNA vaccines. Alternatively, heterologous prime-boost with adeno-vectored and mRNA vaccine elicited 4.5 and 2.6 times higher neutralizing titers against Alpha and 8.6 and 5.8 times higher neutralizing titers against Beta compared with homologous prime-boost with ChAdOx1 and BNT162b2, respectively (Hillus et al., 2021). This approach could be used to enhance protection of individuals previously immunized with vaccines that are more likely to be escaped by emerging variants. Despite the two booster strategies described, it remains clear that determining the real efficacy of vaccines against viral variants, and particularly translating immunity to efficacy against disease, is challenging; thus, the identification of correlates of protection is still paramount to guide coordinated vaccination efforts.
C. Correlates of Protection
Correlates of protection have yet to been defined for COVID-19 vaccines, but they are critically important to rapidly evaluate vaccine efficacy and to enable timely reactions to the emergence of concerning variants (Karim, 2021). Accumulating data from animal models and clinical trials on the relationship between antibody levels and efficacy suggest that antibodies play a major role in protecting from SARS-CoV-2 infection. Indeed, passive transfer of neutralizing antibodies was reported to prevent SARS-CoV-2 infection in NHPs (McMahan et al., 2021). In addition, studies in NHPs also pointed toward the role of antibody-dependent effector functions, particularly when CD8+ T cells are elicited (McMahan et al., 2021).
Due to the rapid evolution of SARS-CoV-2, vaccine efficacy has become increasingly difficult to demonstrate in clinical trial settings, as data are only generated for the variants dominant at a particular time and place. Emergence of SARS-CoV-2 variants also affects real-world vaccine effectiveness (likely to vary with everchanging distribution of variants) and vaccine-induced protection, which will also decrease due to waning immunity. Determining correlates of protection in humans is complicated due to the absence of standardized clinical trial endpoints and assays for evaluation of immune response (World Health Organization, 2021a). Nonetheless, using current data from vaccines trials and convalescents, two independent studies found a strong correlation between neutralizing antibodies levels and protection from SARS-CoV-2 infection. Earle et al. determined correlation between postimmunization neutralizing titer and vaccine efficacy (Earle et al., 2021). Moreover, Khoury and colleagues estimated that a neutralizing titer approximately 20% of the average convalescent level was necessary to achieve 50% protection from disease, and that an even lower level (3%) was sufficient to protect from severe infection (Khoury et al., 2021). The role of vaccine-induced cellular mediated immunity is still under investigation. SARS-CoV-2 spike mRNA vaccines mediate protection from severe disease early after the first dose, when antibody titer is low. In this early window, functional vaccine-induced CD8+ T cells could be responsible for part of the protective efficacy of mRNA vaccines (Oberhardt et al., 2021). Structure-based network analysis was used to define mutationally constrained CD8+ T cell epitopes across the SARS-CoV-2 proteome, a finding that could be used to design novel T-cell-based vaccines against emerging variants and other coronaviruses (Nathan et al., 2021).
VII. Perspective: The Challenges Ahead
The first generation of COVID-19 vaccines demonstrated efficacy significantly above the 50% lower limit recommended by the WHO and considered by FDA for licensure, with all candidates protecting from COVID-19 disease and up to 95% vaccine efficacy estimates. A vaccine that has at least 50% efficacy may in theory provide protection in vaccinated individuals (Krause et al., 2020). However, the appearance of SARS-CoV-2 variants dramatically reduced the effectiveness of some vaccines and shifted the requirements for herd immunity, prompting new research strategies against emerging viral mutants. Aside from boosting existing vaccine-induced immunity, a second generation of COVID-19 vaccines could be deployed to achieve enhanced protection against identified variants and potentially new ones (Choi et al., 2021). For this reason, active research for a “pan-corona” vaccine is pushing the field toward new solutions (Cohen, 2021a). Broad protection and crossreactive immunity will likely have to be assessed in case of vaccination of previously infected or previously vaccinated people. A critical aspect would be a better understanding of the plasticity of the immune response in these individuals. Although homologous boosting increases both neutralizing antibody titers and cellular immunity, studies will have to be focused on determining how a primed immune system can be effectively reprogrammed toward new specificities, or whether pre-existing immunity will dominate recall responses. The phenomenon of continuous recall of B cells activated by past immunizations was observed for influenza vaccines (“original antigenic sin”) and will be an important consideration for future vaccination strategies against SARS-CoV-2. For this reason, studying the evolution of the human B cell repertoire after immunization is critical to elucidate the underlying parameters that contribute to the broadening of immune responses. Optimal localization of the immune response is also an important parameter to consider. An increased understanding of how to elicit robust mucosal immunity will provide a more robust first line of defense against the virus. To achieve this, intranasal vaccination could be used, a route of immunization being considered for ChAdOx1 and other vaccines (Lund and Randall, 2021).
A second consideration for a new generation of COVID-19 vaccines is the assessment of evolution of viral epidemiology and its impact on human health. A better understanding of viral mutation, escape, and distribution in the biosphere will inform scientists on the most appropriate vaccination strategies. Continuous mapping of viral mutations and data integration with laboratory experiments will be needed. In this regard, the isolation of mAbs will allow precise mapping of escaping epitopes, thus allowing rational design of optimized antigens (Correia et al., 2014). Together, accumulation of structural and epidemiology data will provide an unparalleled understanding of viral protein evolution. The 5-year strategic plan recently launched by the Coalition for Epidemic Preparedness Innovations might address most of these issues, including surveillance of new emerging SARS-CoV-2 variants and development of broadly protective vaccines, with a proposal to compress vaccine development timelines from 1 year to 100 days (Samarasekera, 2021). Similar strategic plans are also considered by the United States and European Union governments, aimed at funding multibillion dollar end-to-end solution from surveillance to rapid development and manufacture of vaccines, treatments, and diagnostics.
A third consideration for future SARS-CoV-2 vaccines is safety and reactogenicity. Although current vaccines have an acceptable reactogenicity profile, the frequency of adverse events may vary between age groups, and safety concerns were raised for both adenovirus and RNA vaccines (Angeli et al., 2021). A better safety profile will be necessary for a second generation of vaccines. Particularly, continuous monitoring of anaphylaxis and other adverse events in the population is needed, together with assessment of long-term safety and efficacy. Refined and improved formulations will likely allow a better safety profile and even an improved storage and handling of vaccines.
This leads us to the last point to consider, which is not the least in terms of importance. Most of the industrialized world is quickly rolling out vaccines, with access being broadened to all age groups. However, developing countries are lagging, with difficult access to costly, degradable, highly efficacious vaccines. The WHO estimated that if all available doses were distributed equitably, all health workers and older people worldwide would have been already vaccinated. Despite the availability of a sufficient number of doses globally, lack of access to vaccines in low- and lower-middle–income countries means that WHO’s target of stopping the pandemic by vaccinating ≥40% of people in every country by the end of 2021 and ≥70% by the first half of 2022 will not be reached (World Health Organization, 2021e). Equitable access to vaccination needs to be placed at the center of all current and future vaccination strategies, and worldwide roll-out capability will have to be an important comparator of next-generation vaccines. Fast and equitable supply requires discussion in the regulatory domain supporting implementation of common principles and initiatives for equitable access (Azevedo Soares, 2020). From the manufacturing perspective, it will be important to scale up operations and to define quality specifications, together with a strong stability strategy and comparability of analytical assays and product validation from multiple manufacturing sites (World Health Organization, 2021d).
In conclusion, in less than 2 years we have experienced a massive effort to map regions of vulnerability of SARS-CoV-2 virus and to understand how the immune system reacts to it. The accumulation of knowledge was used to parallel-develop highly effective vaccines in record time, making them available to the general population, yet disparities in vaccine access emerged. At the same time, viral evolution into variants that escape common antibody responses elicited by the original strain prompted a new wave of development effort, which will likely result in even more effective and safe second-generation vaccines. Learnings from the COVID-19 pandemic is ultimately changing the way we approach vaccinology and will hopefully result in a better understanding of our immunity and in a technological leap that could be used to design effective treatments for other infectious and noninfectious diseases.
VIII. Methods
This review is based on SARS-CoV-2 and COVID-19 related data published in English encompassing the time period between late 2019, when the virus was first reported, up to September 2021. Studies related to SARS-CoV-2 immunology and vaccines from PubMed, Biorxiv, and Medrxiv, as well as certain press releases were considered for inclusion. Studies of seasonal and epidemic coronaviruses, including SARS-CoV-1 and MERS-CoV, were considered for inclusion. Both animal and human studies related to SARS-CoV-2 were considered for inclusion.
Acknowledgments
The authors thank Dr. Roberta Cozzi for rendering the SARS-CoV-2 S protein in Fig. 2. The authors also thank Modis for editorial assistance and manuscript coordination, on behalf of GSK. Petronela M. Petrar provided editorial support, and Sander Hulsmans and Stephanie Deroo coordinated the manuscript development.
Authorship Contributions
Wrote or contributed to the writing of the manuscript: Pecetta, Kratochvil, Kato, Vadivelu, Rappuoli.
Footnotes
This work was supported by GlaxoSmithKline Biologicals SA, which was involved in all stages and covered the costs associated with the development and publishing of this manuscript.
S.P., K.V., and R.R. are full-time employees of the GSK group of companies. K.V. and R.R. hold shares in the GSK group of companies as part of their remuneration. S.K. and Y.K. do not have an actual or perceived conflict of interest with the contents of this article.
Abbreviations
- ACE2
- angiotensin-converting enzyme 2
- ADE
- antibody-dependent enhancement
- COVID-19
- infectious disease caused by the SARS-CoV-2 virus
- E
- envelope
- EU
- European Union
- EUA
- emergency use authorization
- FDA
- U.S. Food and Drug Administration
- GMT
- geometric mean titer
- M
- membrane
- mAb
- monoclonal antibody
- MERS-CoV
- Middle East respiratory syndrome coronavirus
- N
- nucleocapsid
- NHP
- nonhuman primate
- NTD
- N-terminal domain
- PCR
- polymerase chain reaction
- RBD
- receptor binding domain
- RSV
- respiratory syncytial virus
- S
- spike
- SARS-CoV-1
- severe acute respiratory syndrome coronavirus
- SARS-CoV-2
- severe acute respiratory syndrome coronavirus 2
- VH3-53
- IgG heavy-chain variable region 3-53
- VLP
- viruslike particle
- VOC
- variant of concern
- WHO
- World Health Organization
- Copyright © 2022 The Author(s)
This is an open access article distributed under the CC BY Attribution 4.0 International license.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
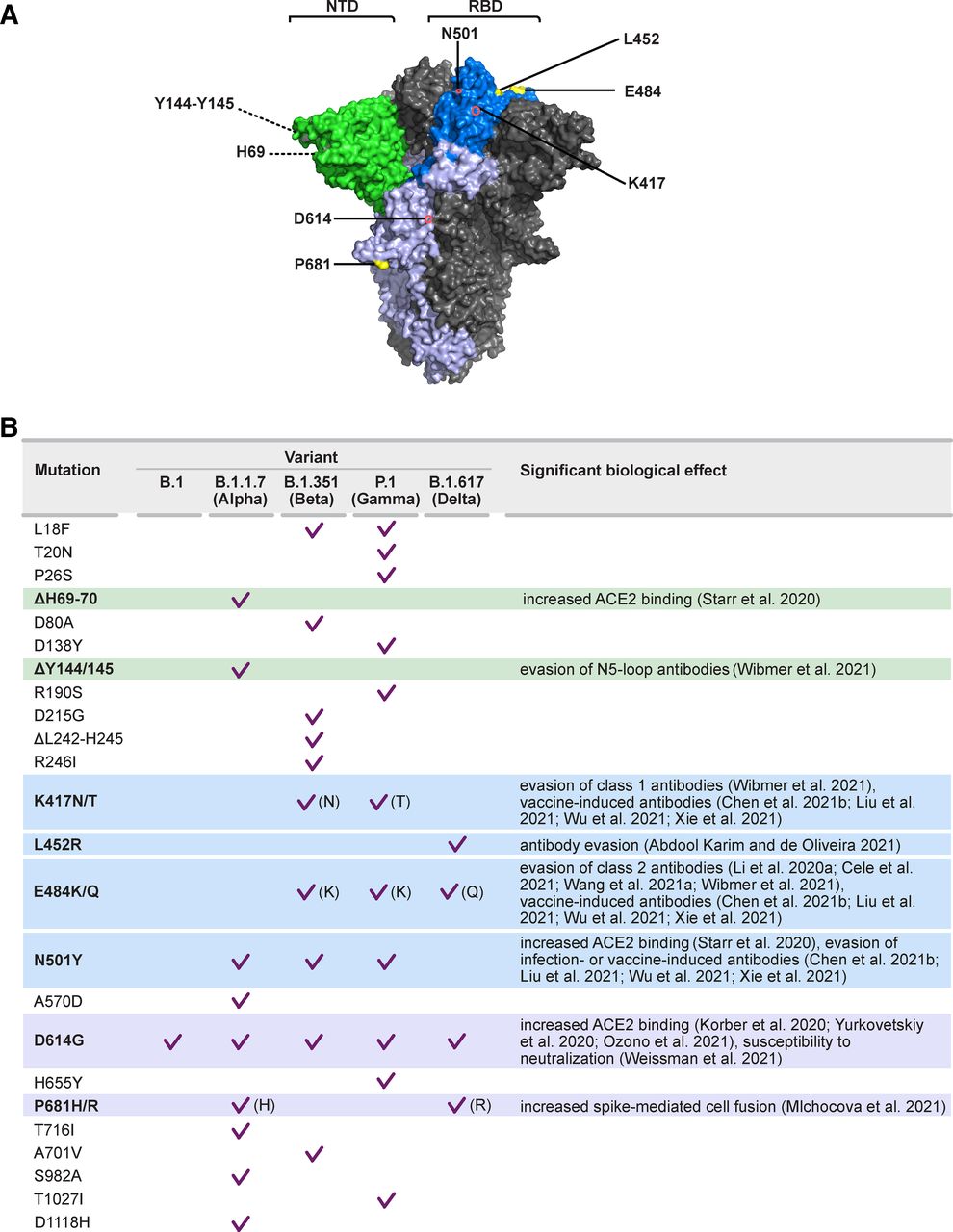{kind=link}