


Abstract
There is a vital need to understand mechanisms contributing to susceptibility to depression to improve treatments for the 11% of Americans who currently suffer from this debilitating disease. The adaptive immune system, comprising T and B cells, has emerged as a potential contributor to depression, as demonstrated in the context of lymphopenic mice. Overall, patients with depression have reduced circulating T and regulatory B cells, "immunosuppressed" T cells, and alterations in the relative abundance of T cell subtypes. T helper (Th) cells have the capacity to differentiate to various lineages depending on the cytokine environment, antigen stimulation, and costimulation. Regulatory T cells are decreased, and the Th1/Th2 ratio and the Th17 cells are increased in patients with depression. Evidence for changes in each Th lineage has been reported to some extent in patients with depression. However, the evidence is strongest for the association of depression with changes in Th17 cells. Th17 cells produce the inflammatory cytokine interleukin (IL)-17A, and the discovery of Th17 cell involvement in depression evolved from the well established link that IL-6, which is required for Th17 cell differentiation, contributes to the onset, and possibly maintenance, of depression. One intriguing action of Th17 cells is their participation in the gut-brain axis to mediate stress responses. Although the mechanisms of action of Th17 cells in depression remain unclear, neutralization of IL-17A by anti-IL-17A antibodies, blocking stress-induced production, or release of gut Th17 cells represent feasible therapeutic approaches and might provide a new avenue to improve depression symptoms.
Significance Statement Th17 cells appear as a promising therapeutic target for depression, for which efficacious therapeutic options are limited. The use of neutralizing antibodies targeting Th17 cells has provided encouraging results in depressed patients with comorbid autoimmune diseases.
Introduction
Immunity is a critical guardian of health involving both protection against pathogens and maintenance of homeostasis. The immune response is often viewed as the cooperation between the innate immune system, which responds quickly and often unselectively to an insult, and the adaptive immune system, which takes time to be activated and requires antigen selectivity. Immune responses are often considered peripheral events in response to pathogen infections or injuries; nonetheless, they are not without effects on the central nervous system. This review will focus on the adaptive immune responses associated with depression.
Depression is a widespread and debilitating disorder. 17.3 million (7.1%) of adults and 2.3 million (9.4%) of adolescents aged 12–17 had at least one major depressive episode in the United States in 2017 (NIMH data). Women are more affected than men, and around two-thirds of patients with depression also experience severe emotional and cognitive impairments and difficulty handling daily activities such as sleeping, eating, or working. A major depressive episode is defined by the Diagnostic and Statistical Manual of Mental Disorders (DSM)-5 and is diagnosed if a person experiences a constellation of symptoms associated with depressed mood for at least 2 weeks. The etiology of depression remains mainly unknown, and the manifestation of the disease is heterogeneous. The risk factors for depression include a combination of genetic, environmental, biologic, and psychologic factors, in addition to comorbid disorders (e.g., diabetes, cardiovascular diseases, etc., and of relevance for this review autoimmune diseases). Multiple treatment options are available including medications, psychotherapy, and electroconvulsive therapies. Although efficient in around one-third of patients, antidepressant medications have the disadvantage of taking time to work (usually 2–4 weeks), and many patients develop treatment resistance (Beurel et al., 2020).
I. Dysregulation of the Adaptive Immune Response in Depression
A. Adaptive Immune System and Depression
Immune responses in patients with depression have attracted attention after findings that patients with depression have a variety of elevated inflammatory markers called cytokines, compared to healthy controls (Raison et al., 2006). Cytokines are produced by the innate immune system and participate in the quick resolution of the insult(s). In clinical trials for depression, patients have found benefit after selective anti-cytokine therapies, such as neutralization of tumor necrosis factor (TNF) or interleukin (IL)-6, although these results are not always reproduced and the sample size is small, mainly due to the expensive nature of these therapies. Yet the adaptive immune system also produces cytokines, and some promising therapies have emerged from targeting cytokines produced by the adaptive immune system (e.g., anti-IL-23 or anti-IL-17A therapies), which will be discussed in more detail later in this review.
The adaptive immune system has been well described in textbooks (Murphy et al., 2022). Briefly, it is composed of lymphocytes T and B cells and is activated when the innate immune system is overwhelmed by an infection or an insult, which is often the case of infections with pathogens. The microenvironment produced by the innate immune response to the infection or insult provides the adequate conditions to activate the adaptive immune system in an antigen specific manner. T cells are circulating in the bloodstream after they complete their development in the thymus and reach peripheral lymphoid organs through the lymphatics, where they usually encounter processed antigens by the antigen presenting cells. T cells that have not encountered antigens yet are called naïve T cells, whereas effector T cells are ready to perform their function as soon as they encounter their specific antigen. Every effector T cell can differentiate in several functional classes specialized in several different activities. Thus, cluster of differentiation (CD)8+ T cells recognize pathogens presented by major histocompatibility complex (MHC) class I molecules, which are expressed by nearly every mammalian cell type, and differentiate into cytotoxic effector T cells to kill infected cells. In contrast, CD4+ T cells recognize pathogens presented by MHC class II, in which expression is restricted to cells of the immune system and have the capacity to differentiate in a continuum of T helper (Th) lineages. B cells are in charge of the humoral immunity and when differentiated into plasma cells, produce antibodies. Antibodies often survey extracellular space to eliminate any pathogens using a variety of mechanisms (direct blockade of pathogen infectivity, induction of the complement cascade leading to phagocytosis, etc.). Altogether the adaptive immune system is critical to resolve the insult, which explains why mounting immune responses to self-antigens often lead to the development of autoimmune diseases.
Unmedicated patients with major depressive disorders often exhibit lymphopenia (reduced number of circulating lymphocytes) (Kronfol et al., 1984; Schleifer et al., 1984; Irwin et al., 1990; Herbert and Cohen, 1993; Zorrilla et al., 2001). When examining leukocyte enumeration, the data are more controversial with reports of decrease of T cells (Schleifer et al., 1984,1985), B cells (Schleifer et al., 1984; Maes et al., 1992b; Petitto et al., 1993; Ahmetspahic et al., 2018), and increase of the CD4/CD8 ratio (Darko et al., 1988; Tondo et al., 1988; Müller et al., 1989) or no change in any lymphocyte subsets (Calabrese et al., 1986; Darko et al., 1989; Kronfol and House, 1989; Schleifer et al., 1989). Other cells from the innate immune system (e.g., natural killer cells, neutrophils, and monocytes) have also been shown to be differentially present in patients with major depressive disorder (MDD) (Evans et al., 1992; Ravindran et al., 1999). Furthermore, recent studies have shown an association between low white blood cell counts and severity of depression (Beydoun et al., 2016; Köhler et al., 2017), whereas others report in larger cohorts of patients a genetic association between polygenic score and increased white blood cell counts (Shafiee et al., 2017; Sealock et al., 2021). Whether these controversial reports result from the analysis of different subpopulations of T or B cells remains to be determined. It is indeed only recently that advances in the immunophenotyping of immune cells have allowed the discovery of subsets of T or B cells associated with major depressive disorder. For example, CD24+CD38hi transitional B cells, which are associated with a regulatory phenotype, are reduced in MDD (Ahmetspahic et al., 2018). Transitional B cells originate from the bone marrow to reach the lymphoid tissues to complete maturation (Sims et al., 2005; Palanichamy et al., 2009). Similarly, memory CD4+ T cells and CD69+CD19+ activated B cells have been found reduced among peripheral blood mononuclear cells exposed to plasma of unmedicated patients with MDD, compared with exposure to plasma of healthy control subjects (Syed et al., 2018). Furthermore, telomeres of three lymphocyte subpopulations [CD8+ (T cytotoxic cells), CD20+ (B cells), and CD4+ (T helper) cells] are shorter in female patients with MDD compared with age matched female controls (Karabatsiakis et al., 2014), suggesting that these subpopulations of lymphocytes age faster in patients with depression. The different subtypes of Th cells will be discussed in greater details in the next section.
In mouse models, acute stress seems associated with increased immune responses, whereas chronic stress in contrast reduces immune responses including reduced leukocyte trafficking, neutrophil phagocytosis, and reduction of peripheral lymphocytes (Padgett and Glaser, 2003; Glaser and Kiecolt-Glaser, 2005; Fan et al., 2019). Yet, Rag1−/− or Rag2−/− mice, which are deficient in T and B cells, have been shown to exhibit attenuated depressive-like and anxiety-like behaviors (Rattazzi et al., 2013; Clark et al., 2015, 2016 ,Rilett et al., 2015; Fan et al., 2019). Furthermore, adoptive transfer of splenocytes/lymphocytes from chronically stressed mice protects naïve mice from exhibiting anxiety-like and depressive-like behaviors (Lewitus et al., 2009; Brachman et al., 2015). T cells are also necessary to promote the resolution of inflammation-induced depressive-like behaviors (Laumet et al., 2018). The role of B cells in depressive-like behaviors has been less studied, but some reports have shown an amelioration of depressive symptoms in patients receiving B cell neutralizing antibodies (Wittenberg et al., 2020), whereas others have reported increased incidence of depression in patients with autoimmune disorders treated with the anti-B cell activating factor (BAFF) antibodies (Belimumab) (Minnema et al., 2019), which deplete B cells, as BAFF is a B cell survival cytokine belonging to the TNF superfamily. In addition, autoantibody levels are elevated in MDD patients (Iseme et al., 2014; Postal and Appenzeller, 2015). Overall, these findings suggest that the apparent controversial effects of T and B cells in depression might in fact result from the existence of various T and B cell subtypes that might respond differentially to the environment (e.g., cytokine milieu, stress hormones, microbiome composition, etc.).
B. T Helper Cells and Depression
In response to the milieu of cytokines, the activation by an antigen, and a costimulatory signal, CD4+ T cells differentiate into a specific lineage belonging to a continuum of phenotypes. The main lineages studied in depression are: Th1, Th2, Th17, cells and regulatory T cells (Treg) (Fig. 1). Simplistically, Th1 cells require IL-12 and interferon (IFN)γ to differentiate in vitro and produce IFNγ; Th2 cells require IL-4 and produce IL-4, IL-5, and IL-13; Th17 cells require IL-6 and transforming growth factor (TGF)β and produce IL-17A; and Treg cells require TGFβ and express forkhead box P3 (Foxp3) (Zhu et al., 2010).
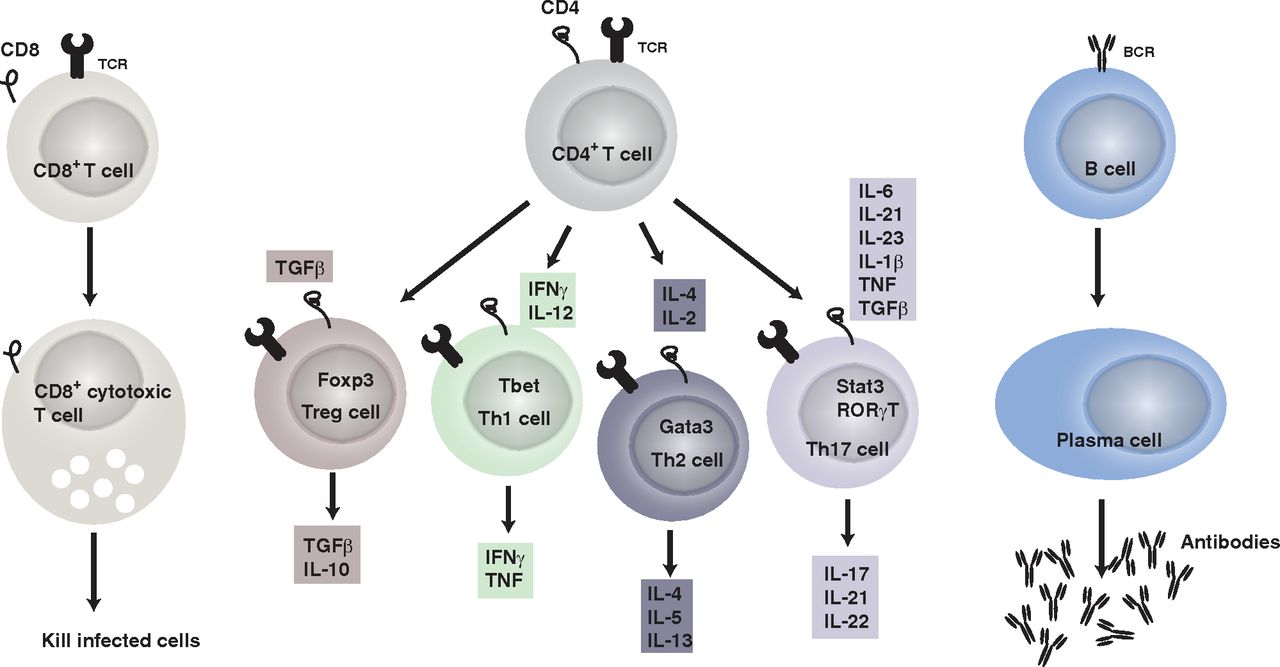
The adaptive immune system. It includes T (CD4+ and CD8+) and B cells. CD8+ T cells differentiate into cytotoxic T cells after recognizing antigen presented by MHC class I. CD4+ T cells in contrast recognize antigens presented by MHC class II on antigen-presenting cells. Once activated by their antigen and the cytokine milieu (each lineage of CD4+ T cells has a combination of cytokines required to differentiate), as well as upon receiving costimulatory signals, CD4+ T cells differentiate in the different lineages: Treg, Th1, Th2, and Th17 cells, which produce a combination of cytokines as indicated. Finally, CD4+ T cells help B cells to differentiate into plasma cells once they encounter a specific antigen and plasma cells produce antigen specific antibodies.
1. T Helper 1/T Helper 2 Cells
Although data are limited, there is growing evidence of alterations of Th cells in MDD. Some findings include evidence in patients with MDD of increased Th cells relative to T cytotoxic cells (Darko et al., 1988; Tondo et al., 1988; Müller et al., 1989), and elevated ratio of Th1/Th2 cells (Maes et al., 1992a) that is reversed by antidepressants (Kubera et al., 2001). It is generally thought that acute mild stress increases Th2 activation, whereas chronic or repeated stress leads to Th1 activation (Maes et al., 2012). Th1 cells are increased in the brains of mice subjected to learned helplessness, although their role in promoting learned helplessness is less clear (Beurel et al., 2013, 2018a). Nonetheless, Th1 cell specific transcription factor Tbet knockout mice do not exhibit depressive-like behaviors (Kim et al., 2011), suggesting a positive role of Th1 cells in promoting depressive-like behaviors. One of the possible mechanisms whereby Th1 cells promote depressive-like behaviors is through the activation of the enzyme indoleamine 2,3-dioxygenase (IDO). Indeed, IDO converts tryptophan to kynurenine, depleting the pool of tryptophan required for the synthesis of serotonin. IDO is activated in depression (Wichers and Maes, 2004) and antidepressants reduce Th1 cell level (Mohr et al., 2001). In addition, polymorphisms in the Th1 gene Tbet predict diagnosis of MDD (Wong et al., 2008). These suggest that the proinflammatory Th1 cells promote depression. In contrast, Th2 cells attenuate sepsis-associated encephalopathy-induced anxiety-like behaviors (Saito et al., 2021). Reciprocally, Th2-associated cytokine IL-4-deficient mice exhibit enhanced depressive-like behaviors (Wachholz et al., 2017). IL-4 inhibits depressive-like behaviors induced by IL-1β-mediated-astrocytic activation and IL-1β-induced alterations of serotonin and norepinephrine levels (Park et al., 2015). These suggest that Th2 cells might be anti-inflammatory and protect from depression.
2. T Helper 17 Cells
Recently, most of the studies have focused on the newly discovered Th17 cell subset of T helper cells (Harrington et al., 2005). There is accumulating evidence that Th17 cells are increased in depression and contribute to the pathology of depression. Thus, patients with MDD have elevated blood levels of Th17 cells (Chen et al., 2011; Ghosh et al., 2020), and patients at high risk of suicide exhibit the highest level of Th17 cells (Schiweck et al., 2020). Furthermore, CD4+ T cells isolated from patients with generalized anxiety disorder when activated in vitro acquire a Th17 phenotype (Vieira et al., 2010; Ferreira et al., 2011). It is important to note patients with Th17 cell-associated autoimmune diseases often exhibit comorbid MDD (Kurd et al., 2010; Patten et al., 2017; Singh et al., 2020; Sales et al., 2021). Consistent with these findings, the signature inflammatory cytokine produced by Th17 cells, IL-17A, is elevated in some patients with MDD (Chen et al., 2011; Liu et al., 2012; Kim et al., 2013a; Davami et al., 2016), but not all (Liu et al., 2012; Kim et al., 2013b), and in patients with post-traumatic stress disorder (Zhou et al., 2014). In addition, the treatment response to certain antidepressants (bupropion and escitalopram combination) is predicted by IL-17A levels (Jha et al., 2017). Depressive symptoms are reduced after anti-IL-17A treatment in 40% of patients with psoriasis experiencing MDD (Griffiths et al., 2017), whereas blocking the downstream effectors of IL-17A in patients with psoriasis by blocking its receptor using anti-IL-17RA therapy has been associated with increased psychiatric disorders and suicidality risk (Lebwohl et al., 2018).
Studies in rodents provide corroborative evidence of these detrimental effects of Th17/IL-17A in depression, such as administration of IL-17A in rodents promotes depressive-like behaviors (Nadeem et al., 2017). Stress increases IL-17A levels (Lu et al., 2017; Cheng et al., 2018; Gu et al., 2018; Zhang et al., 2019). Th17 cells accumulate in the brain of learned helpless mice or mice subjected to chronic restraint stress (Beurel et al., 2013, 2018; Medina-Rodriguez et al., 2020). Th2 and Th17 responses are associated with exacerbation of fear in a severe allergen induced airway hyperreactivity mouse model (Lewkowich et al., 2020). Splenic Th17 cells are increased after stress induced by social defeat (Ambrée et al., 2019), and Th17/Treg ratio is increased in the liver and ileum of mice subjected to chronic stress (Westfall et al., 2021). The involvement of Th17 cells in promoting depressive-like behaviors has been demonstrated by using a variety of approaches: stimulation by IL-17A (Nadeem et al., 2017), neutralization of IL-17A (Beurel et al., 2013; Nadeem et al., 2017; Kim et al., 2021), pharmacological inhibition (Beurel et al., 2013), or knockout approach where the master transcription factor of Th17 cells is depleted (Beurel et al., 2013). Th17 cells increase microglial activation in the hippocampus, amygdala, and prefrontal cortex of cumulative mild prenatal stress, and anti-IL-17 treatment rescues depressive-like and anxiety-like behaviors after perinatal stress (Kim et al., 2021). Moreover, the transcription factor Retinoic acid receptors (RAR)-related orphan receptor gamma T (RORγT) is required for Th17 induced depressive-like behaviors (Beurel et al., 2013, 2018a). Little is known about the mechanisms of action of Th17 cells, as well as their mode of infiltration into the brain (Fig. 2). Nonetheless, the expression of the chemokine receptor C-C motif chemokine receptor (CCR)6 is required for Th17 cells to infiltrate the brain (Beurel et al., 2018a). Th17 cells in the brain also express the follicular marker C-X-C motif chemokine receptor 5, whose contribution to depression is less clear (Beurel et al., 2018a). The possible mechanisms of action of Th17 cells include induction of neuroinflammation by activated microglia and astrocytes and/or induction of neuronal cell death [e.g., increased IL-17A-mediated astrocytic glutamate excitotoxicity (Kostic et al., 2017)] as well as inhibition of neurogenesis by IL-17A as shown by IL-17A knockout, which increases neurogenesis (Liu et al., 2014) (Fig. 2).

Potential mechanisms whereby Th17 cells promote depressive symptoms. Upon stress, Th17 cells enter the brain. CCR6 and IL-17A seem critical for the entry of pathogenic Th17 cells into the brain after stress. Various effects of Th17 cells have been proposed in the brain. Neurons, microglia, and astrocytes express IL-17RA and are therefore regulated by the Th17 cell signature cytokine IL-17A. In astrocytes, for example, IL-17A activates glutamate excitotoxicity. IL-17A also inhibits neurogenesis. Th17 cells have also been reported to trigger neuronal cell death via direct contact leading to increased intracellular Ca2+ level or through activation of neuronal Fas by FasL produced by Th17 cells.
Th17 cells are present in healthy mice in the lamina propria of the small intestines because of the presence of the bacteria Segmented filamentous bacterium (SFB), which is required to induce the differentiation of Th17 cells (Ivanov et al., 2009). SFB level predicts sensitivity to the learned helplessness paradigm in a Th17 cell-dependent manner. Mice deficient in SFB are resistant to the learned helplessness (Medina-Rodriguez et al., 2020). Furthermore, CD4+ T cells specific to SFB are sufficient to promote depressive-like behaviors and accumulation of Th17 cells in the hippocampus of Rag2−/− mice (Medina-Rodriguez et al., 2020). This suggests that microbiome alterations might be responsible for the increased Th17 cell response in depression.
3. Treg Cells
In contrast to those inflammatory Th cell types, anti-inflammatory Treg cells are decreased in patients with MDD (Li et al., 2010; Chen et al., 2011; Grosse et al., 2016b; Jahangard and Behzad, 2020) or adolescents with high risk for mood disorders (Snijders et al., 2016), whereas other studies find increased regulatory T cells in patients with MDD (Suzuki et al., 2017; Patas et al., 2018). Treg cells are decreased after chronic unpredictable stress in rodents (Hong et al., 2013). Depletion of Treg cells is associated with higher rate of depressive-like behaviors in mice (Kim et al., 2012). Depletion of Treg cells is also associated with increased levels of Th1 (IFNγ), Th2 (IL-4), Th17 (IL-17) cytokines, and reduced serotonin level (Kim et al., 2012). In addition, Treg cells are increased after effective antidepressant treatment (Himmerich et al., 2010; Grosse et al., 2016a). Altogether, findings on T helper cells suggest that depression is associated with more proinflammatory Th cells and fewer immunosuppressing regulatory T cells.
II. Possible Origins of These Immune Dysregulations in Depression
Although it remains to be determined whether all the dysregulations of the adaptive immune system observed in patients with depression occur in the same individual at the same time, it is possible to envision that factors specific to depression might affect the adaptive immune response. Thus, for example, it is possible that the plasticity of Th cells is responsible for some of the differences observed between the different Th subtypes in depression. Late stage Th17 cells can indeed differentiate into Th1 cells (Lee et al., 2009) or regulatory T cells (Gagliani et al., 2015), for example. It is also possible that previous treatments with antidepressants affect the proportion of T helper cells. In addition, Th cells are differently affected by stress, including the type of stress (acute versus chronic), the stress hormones, as well as indirect effects of stress such as alterations of the microbiota, which is known to control immunity. Thus, acute stress promotes leukocyte mobilization to the blood (Dhabhar et al., 2012) and infiltration into the site of inflammation (Viswanathan and Dhabhar, 2005) and mostly activates proliferative responses of immune cells (Rinner et al., 1992). In addition, increased activity of natural killer and CD8+T cells is found in humans subjected to acute stress. Chronic stress in contrast is immunosuppressant in animals and humans (Kiecolt-Glaser et al., 1996; Dhabhar, 2000). Some of the pathways contributing to these different actions of stress are described below (Fig. 3).

Effects of glucocorticoids, catecholamines, and microbiota on T helper cells.
A. Glucocorticoids
Glucocorticoids are small lipid hormones that are produced by the adrenal glands and are immunosuppressant in periphery and, therefore, have been used as anti-inflammatory and immunosuppressive therapies. Stress and circadian alterations change the level of glucocorticoids to regulate metabolism, neuronal and cardiovascular functions, as well as immune activity. Exogenous glucocorticoid administration such as corticosteroid treatment in mice provides immunosuppression in part by inhibiting the secretion of T cell growth factors; blocking the gene expression of a wide variety of cytokines including IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-8, IL-10, IL-13, granulocyte-macrophage colony-stimulating factor (GM-CSF), TNF, and IFNγ; and shaping the cytokine milieu required for T cell differentiation (for review Taves and Ashwell, 2021). Endogenous glucocorticoids, however, can promote or suppress immunity. Glucocorticoids have been implicated in the selection of the appropriate T cell receptor repertoire in the thymus, suppressing Th1 cells (Franchimont et al., 2000) while moderately inhibiting Th2 cells and favoring Th17 cells (Schewitz-Bowers et al., 2015), regulating T cell trafficking, and promoting T cell memory (Taves and Ashwell, 2021). Glucocorticoid receptors are expressed on all T cells, but different T cells have different sensitivity to glucocorticoids. The difference is mainly attributed to the glucocorticoid-induced loss of expression of B-cell lymphoma 2 (Bcl-2), and greater expression of Bim in Th1 cells, whereas Bcl-2 is moderately affected in Th2 cells and increased in Th17 cells (Banuelos et al., 2017). Furthermore, Treg cell differentiation is also promoted by glucocorticoids due to the upregulation of TGFβ receptors and Foxp3 (Karagiannidis et al., 2004), and the promotion of regulatory T cells by glucocorticoids might be one of the major mechanisms mediating immunosuppression (Fig. 4). These wide effects of glucocorticoids on immunity might explain the enrichment in certain T cell subtypes during depression, e.g., Th17 cells are the most resistant cells to glucocorticoid-induced apoptosis and are found elevated in depression. Depression is associated with increased level of cortisol/corticosterone in humans and animal models (Pariante, 2009). It is important to note, that although of potential benefit to patients with MDD because glucocorticoids reduce inflammation and inflammation is elevated in patients with MDD, glucocorticoid treatment only provides minimal improvement to patients with MDD with short-term treatments (DeBattista et al., 2000). Furthermore, patients with MDD exhibit a dysregulated response to glucocorticoids (Carvalho et al., 2015), which complicates the interpretation of the role of glucocorticoids in T cell-mediated behavioral alterations. Nonetheless, several studies have pointed toward a blunted leukocyte responsiveness to glucocorticoids in patients with MDD (Bauer et al., 2002, 2003; Raison and Miller, 2003; Carvalho et al., 2014; Grosse et al., 2015; Pariante and Miller, 2001), whereas another study has not (Hasselmann et al., 2018). Similarly, T and B cell responses are modulated by exposure to stress and glucocorticoids in animal models (Roque et al., 2011; Clark et al., 2019), suggesting a possible mechanism whereby lymphocytes become immunosuppressed in depression.
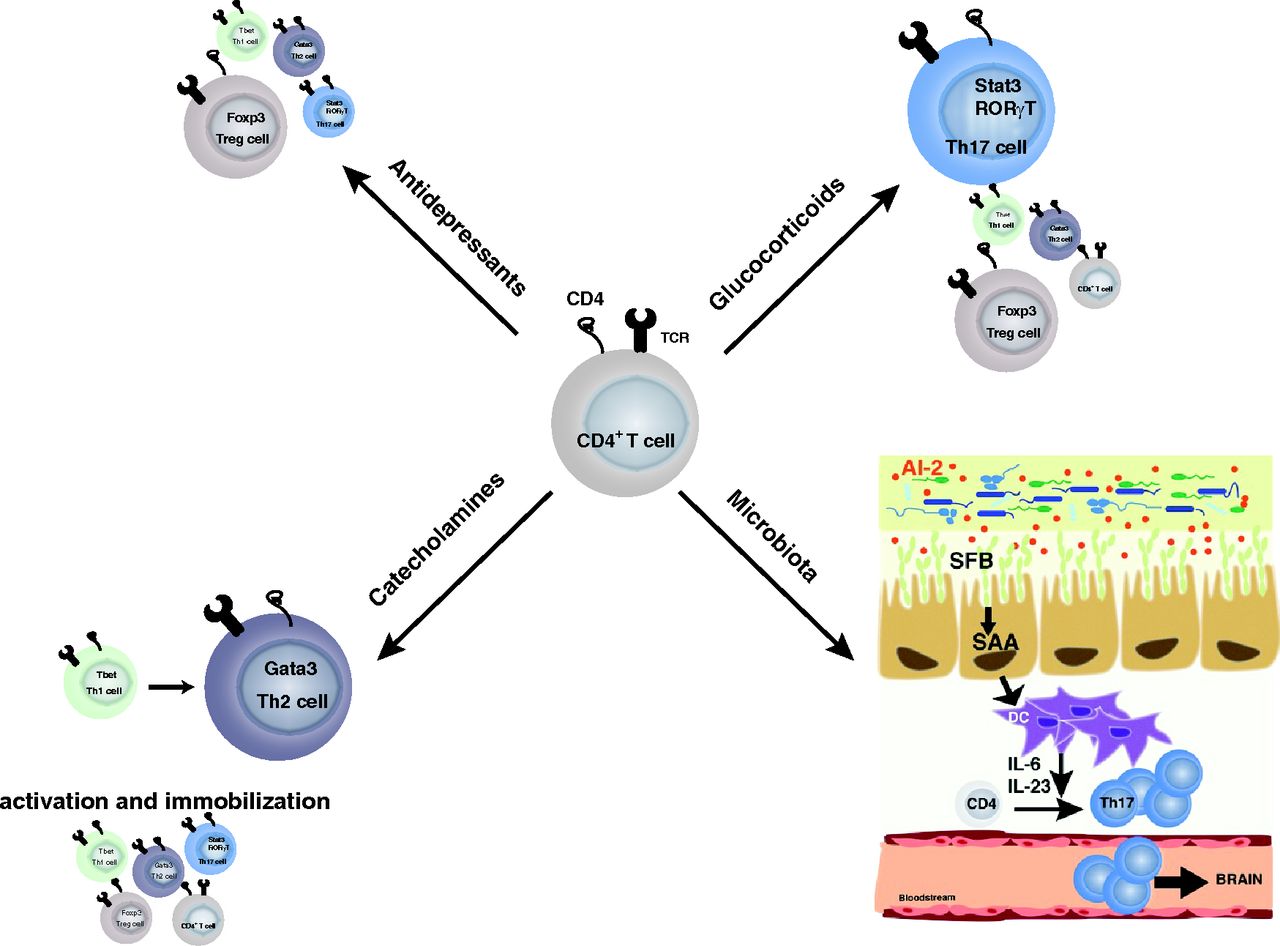
Overall selection of T cells associated with depression. Glucocorticoid treatment and microbiota alterations in depression have been associated with a shift toward Th17 cells. Catecholamines seem to favor Th2 cells. However, catecholamines promote T helper cell activation, whereas they block leukocyte migration. Antidepressant treatments in contrast seem to reduce T helper cells in a nonspecific manner and favor regulatory T cells.
B. Catecholamines
The catecholamines include neurotransmitters such as epinephrine, norepinephrine, and dopamine and act through the autonomic nervous system. Traditionally, the sympathetic nervous system is activated after stress to increase blood epinephrine and norepinephrine levels. Immune cells express both α-adrenergic and β-adrenergic receptors, and their level of expression depends on the state of the cells, their maturation, and activation (Elenkov et al., 2000; Scanzano and Cosentino, 2015). Consistent with this, norepinephrine promotes a shift from Th1 cells to Th2 cells (Wahle et al., 2006; Hou et al., 2013), whereas the overall role of catecholamines on Th17 cells is less clear. Epinephrine promotes Th17 cells via secretion of cytokines by the dendritic cells (Kim and Jones, 2010; Manni et al., 2011), whereas norepinephrine inhibits Th17 cells directly through β2-adrenergic receptors (Liu et al., 2018). Furthermore, effector T cells are activated, whereas naive splenic T cells are suppressed by increased levels of catecholamines (Case and Zimmerman, 2015). These suggest that catecholamines affect activation of T cells. Nevertheless, the duration and concentration of exposure to the catecholamines, the interaction with other factors induced during stress (glucocorticoids, etc.), and the role of adrenergic receptors in the various T cell subtypes need to be also taken into consideration. Maybe the most striking effect of catecholamines on T cells is their effects on migration (Fig. 4). Indeed, T cell migration is temporarily paralyzed in response to stress-induced sympathetic nervous system activation (Devi et al., 2021), which might explain the effect of acute stress on leukocyte migration. Inflammation has been shown to reduce dopamine synthesis and availability (Felger and Treadway, 2017), suggesting that in depression, where inflammation is increased and dopamine level reduced, T cell migration might be enhanced. Other nervous systems also affect T cells, including the parasympathetic nervous system with the release of acetylcholine and the sensory nervous system with the release of neuropeptides and glutamate, which are also known to modulate T cell responses. Of interest for this review, skin sensory neuron activation induces IL-17 response (Cohen et al., 2019).
These suggest that T cells are the point of convergence of many pathways associated with the stress response.
C. Microbiome
Host microbiota interactions influence the development of the adaptive immune system. These interactions are required for homeostasis. Yet, changes in the environment or genetic defects lead to disruption of these interactions and promote diseases. Depression has been associated with changes in the relative abundance of Firmicutes, Actinobacteria, and Bacteroidetes compared to healthy individuals [(Zheng et al., 2016); for review, see (Cheung et al., 2019)], but the pathways and underlying mechanisms used by the microbiota to promote diseases are just emerging.
The adaptive immune system is in close contact with the microbiota, and mutual regulatory mechanisms are in place. Thus, to protect the gut mucosal epithelial layer from microbes, B cells secrete microbial antigen specific IgA into the gut lumen. The production of IgA is critical for shaping the microbial communities and enhancing regulatory T cell expansion (Kawamoto et al., 2014). Th17 cells, which are only present in the small intestinal lamina propria of healthy mice, also participate in the maintenance of the epithelial barrier by producing low amounts of IL-17A (Cypowyj et al., 2012). Th17 cells are absent in germ-free mice, showing a major impact of the microbiota on Th17 cells (Amatya et al., 2017) and suggesting that Th17 cells might be one of the major Th subtypes dependent on the regulation by the microbiota. SFB is one of the first discovered inducers of Th17 cells (Ivanov et al., 2009). Other bacteria [e.g., Escherichia coli (Viladomiu et al., 2017), Bifidobacterium adolescentis (Tan et al., 2016), Staphylococcus aureus (Zielinski et al., 2012), and Candida albicans (Hernández-Santos and Gaffen, 2012)] also increase Th17 cells. However, it is important to note that these different microbes induce different flavors of Th17 cells (Tan et al., 2016). This suggests that other signals are required besides the microbial antigen to promote the differentiation of Th17 cells (Yang et al., 2014). Thus, for example, transdifferentiation of Th17 cells into regulatory T cells is partially dependent on the aryl hydrocarbon receptor (Ahr), whose agonists are produced by the microbiota to promote anti-inflammatory responses comprising of tryptophan metabolites (Hirota et al., 2011; Wlodarska et al., 2017). The abundance of aryl hydrocarbon receptor agonists in the microbiota is often diet dependent. Although Th17 cells might mediate certain actions of the microbiome in depression, the role of the microbiota-induced Th17 cells in depression remains largely unclear. The limited data show Th17 cells mediate SFB-dependent depressive-like behaviors. Stress increases SFB, which promotes the production of the quorum sensing molecule autoinducer-2, which in turn enhances the production of serum amyloid A (SAA)1/2 that is required for Th17 cell differentiation and promotion of depressive-like behavior (Medina-Rodriguez et al., 2020) (Fig. 4).
In contrast, spore forming Clostridium clusters IV and XIVa, Escherichia, Akkermansia, Bacteroides, Lactobacillus, and Streptococcus are potent inducers of regulatory T cells (Atarashi et al., 2013; Ahern et al., 2014; Geva-Zatorsky et al., 2017). Microbial metabolites (e.g., short chain fatty acids, especially butyrate) also promote regulatory T cells (Smith et al., 2013). Butyrate is mainly produced by Clostridiales species such as Ruminococcaceae and Lachnospiraceae. Polysaccharide A produced by Bacteroides species promotes Treg anti-inflammatory effects while inhibiting Th1 and Th17 cell differentiation (Round and Mazmanian, 2010; Round et al., 2011). The overall effect of the microbiota on the balance between Th17 and Treg cells depends on the composition of the microbiota. Thus, by promoting Th17/Treg imbalance microbiota metabolites promote depressive-like behaviors (Westfall et al., 2021). Although Th17 cells and Treg cells seem to be the main Th subtypes modulated by the microbiota, other Th cells have been associated with changes of the microbiota, such as helminth infection, which promotes a Th2 response (Reynolds et al., 2015).
Although the understanding of the effects of the microbiota on T cell plasticity remains to be fully understood, it is conceivable that small changes in the microbiota can have dramatic consequences on the adaptive immune cells that in turn impact mood. Although stress is known to affect microbiota composition, other factors affect the microbiota composition such as diet, disease, environment, etc. and will need to be considered. Recent evidence has pointed toward a dysregulated gut-brain axis including a choroid plexus vascular barrier contributing to the mental symptoms associated with inflammatory bowel disease (Carloni et al., 2021).
Overall, various stress signals converge on the adaptive immune cells, and it remains to be determined which signal(s) can be targeted to improve depression.
III. Implications for Patients with Depression
A. Increased Infections and Comorbidities
“We tend to get sick after stressful events” (Elstad and Vabø, 2008), prompts the question of whether the dysregulated adaptive immune response is responsible for higher sensitivity of patients with depression to infections, since the adaptive immune system is decisive in eradicating infections. A past history of depression is in fact associated with increased risks of infections (Troidle et al., 2003; Adams et al., 2008; Irwin et al., 2011; Seminog and Goldacre, 2013; Andersson et al., 2016). The effectiveness of the immune activity in depression is reduced and has been associated with increased mortality (Wulsin et al., 1999). Similarly, there is an increased risk of developing autoimmune diseases (multiple sclerosis, psoriasis, inflammatory bowel disease, rheumatoid arthritis, and systemic lupus erythematosus) in patients with depression (Kurina et al., 2001; Dickens et al., 2002; Andersson et al., 2015; Euesden et al., 2017; Patten et al., 2017). The autoreactive T cells, especially Th1 and Th17 cells, have been thought to be the culprit, and this is consistent with anti-IL-17A treatment ameliorating depression symptoms in patients with psoriasis (Griffiths et al., 2017). Reciprocally, antidepressant treatments (e.g., Sertraline, psychotherapy, and cognitive behavioral therapy) are associated with Th1-dependent improvement of multiple sclerosis (Mohr et al., 2001). Ketamine reduces the differentiation and proliferation of Th17 cells in a mouse model of experimental autoimmune encephalitis (Lee et al., 2017). Furthermore, there are reports of depression affecting the recovery from surgery (Cheng et al., 2020), and B regulatory cells are reduced in patients with hip fracture who develop depression compared with non-depressed patients with hip fracture or healthy individuals (Duggal et al., 2016). These suggest that the alterations of the immune system, especially of the adaptive immune system in depression are associated with negative outcomes in the long-term.
B. How Does the Adaptive Immune System Promote Depression?
Reciprocally, it also raises the question on how the adaptive immune alteration promotes depression. The adaptive immune system produces cytokines that can impede neuronal circuitries involved in mood regulation. Thus, for example IL-17A, by activating IL-17RA and IL-17RC receptors, leads to the production of cytokines and chemokines, matrix metalloproteinases, and anti-microbial peptides via the activation of TNF receptor associated factor (Traf)6, Traf4, nuclear factor kappa light chain enhancer of activated B cells (NF-κB), CCAAT-enhancer-binding proteins (C-EBP)β, C-EBPδ, and mitogen-activated protein kinase (MAPK) pathways [(for review (Milovanovic et al., 2020)]. Cytokines, especially IL-6, are sufficient to promote depressive-like behaviors. In addition, IL-17RA receptors are expressed on a variety of neuron subtypes, astrocytes, microglia, and endothelial cells in the brain (Das Sarma et al., 2009), broadening even more the effects of IL-17A (Fig. 2). However, it remains to be determined whether these cell specific pathways are all contributing to depression. Astrocytes mediate Th17 cell effects in a mouse model of multiple sclerosis (Kang et al., 2010). Th17 cells disrupt the blood brain barrier by acting on endothelial cells (Kebir et al., 2007), whereas they promote the production of cytokines by microglia (Murphy et al., 2010). Th17 cells promote neuronal damage (Siffrin et al., 2010) (Fig. 2). Although most of these findings were associated with autoimmune diseases for which the number of Th17 cells is higher than in depression, recent studies in autism suggest a Th17 cell-dependent neuronal damage in psychiatric diseases even when the number of Th17 cells is lower. Thus, IL-17A produced by Th17 cells induces abnormal cortical phenotype in a mouse model of maternal immune activation-inducing autistic traits (Choi et al., 2016; Reed et al., 2020), and the abnormal cortical phenotype is microbiota dependent (Kim et al., 2017). Similarly, IL-17A produced by meningeal γδT cells has been shown recently to promote anxiety-like behaviors by activating IL-17RA receptors on cortical glutamatergic neurons (Alves de Lima et al., 2020). Furthermore, Th17 cells induced by high salt diet promote neurovascular dysfunction in an IL-17A-dependent reduction of nitric oxide production by cerebral endothelial cells (Faraco et al., 2018). These data suggest that the action of Th17 cells might be specific depending on the disease and the brain region they infiltrate. Although similar mechanisms of action of Th17 cells in depression are conceivable, the specific mechanisms of action of Th17 cells in depression remain to be determined.
C. Potential Pharmacological Outcomes
Lastly, antidepressant drugs affect directly T cell proliferation and activity. Thus, fluoxetine reduces the proliferation of T cells (Fazzino et al., 2009; Ahern, 2011), leading to a decrease in number of blood T cells (Rosso et al., 2016). Fluoxetine (Himmerich et al., 2010; Grosse et al., 2016a) and ketamine (Hou et al., 2016) increase anti-inflammatory regulatory T cells. Ketamine also decreases Th2 cell differentiation (Gao et al., 2011), inhibits leukocyte adhesion and migration (Larsen et al., 1998; Kawasaki et al., 2001), and induces T cell apoptosis (Braun et al., 2010). Therefore, detangling the contribution of T cells in depression might be compromised by the use of an antidepressant treatment, and further studies will be required to understand the role of the adaptive immune cell response in medication-free depression.
Maybe one of the most promising approaches is the use of cytokine neutralizing antibodies targeting Th17 cells in depression. Indeed, few anti-IL-17A antibodies (Secukinumab, Ixekizumab) and anti-IL-17RA (Brodalumab) are available for psoriasis and have been tested on depressive symptoms associated with autoimmune diseases. Ixekizumab reduces by 40% depressive symptoms in patients with psoriasis (Griffiths et al., 2017). Brodalumab used in psoriasis, in contrast, promotes risk of suicide and of developing psychiatric illnesses, leading the Food and Drug Administration (FDA) to issue a warning (Lebwohl et al., 2018). Furthermore, anti-IL-12/IL-23 (Ustekinumab) treatment significantly improves depressive symptoms compared with placebo in patients with psoriasis (Wittenberg et al., 2020). Using monoclonal antibodies to deplete B cells, such as anti-BAFF (anti-BLγS, Belimumab) or anti-CD20 (Ofatumumab), to treat systemic lupus erythematosus and rheumatoid arthritis, respectively, have been shown to slightly improve depressive symptoms (Wittenberg et al., 2020). However, other reports have found, in contrast, an increase in risk to develop depression and suicidal ideation in patients treated with Belimumab or Ofatumumab (Minnema et al., 2019). These suggest that monoclonal neutralizing antibodies are potential therapies to alleviate depressive symptoms. However, a better understanding of the contribution of the adaptive immune system in depression is a prerequisite to design the most efficacious drug.
Some probiotics regimens have also been shown to ameliorate depressive symptoms (Yong et al., 2020). Of relevance for this review, oleic acid, one of the major components of olive oil, provides antidepressant effects (Lin et al., 2012; Kiecolt-Glaser et al., 2013) and reduces Th17 cell accumulation into the brain (Medina-Rodriguez et al., 2020). This suggests that targeting Th17 cells in periphery might be sufficient to induce antidepressant actions. Other Th17 cell-targeting compounds such as the RAR-related orphan receptor gamma T inhibitor SR1001 block Th17 cell-induced depressive-like behaviors (Beurel et al., 2013). Although all the characteristics of Th17 cells associated with depression remain to be determined, it is conceivable that other features of pathogenic Th17 cells could be targeted in depression to block Th17 cell entry to brain, for example (e.g., blockade of CCR6). Indeed, blocking all Th17 cells might have a negative impact on intestinal mucosal health.
Although limited, the arsenal of drugs to target Th17 cells in depression is expanding.
IV. Conclusion
Altogether, there is substantial evidence for a contribution of the adaptive immune system to depression. On one hand, there is a reduction of the lymphopoiesis in depression due to elevated level of norepinephrine and glucocorticoids (Wohleb et al., 2016). This is accompanied with a switch toward inflammatory T helper cells at the expense of the anti-inflammatory Treg cells, which might be exacerbated by gut microbial changes, which favor proinflammatory Th17 cells in depression. Th17 cells appear to be central in promoting depressive symptoms, and although the mechanisms of action of Th17 cells in depression remain to be fully elucidated, neutralizing the signature cytokine of Th17 cells, IL-17A, provides unexpected antidepressant effects.
Authorship Contributions
Wrote or contributed to the writing of the manuscript: Beurel, Medina-Rodriguez, Jope.
Footnotes
The work was supported by National Institutes of Health National Institute of Mental Health [Grant R01-MH104656] and [Grant R01-MH110415] (to E.B.) and by a Merit Award from the Veterans Administration [BX003678] (to R.S.J.).
No author has an actual or perceived conflict of interest with the contents of this article.
Abbreviations
- BAFF
- B cell activating factor
- CCR6
- chemokine receptor C-C motif chemokine receptor
- CD
- cluster of differentiation
- IDO
- indoleamine 2,3-dioxygenase
- IFN
- interferon
- IL
- interleukin
- MDD
- major depressive disorder
- MHC
- major histocompatibility complex
- SFB
- segmented filamentous bacterium
- TGF
- transforming growth factor
- Th
- T helper
- TNF
- tumor necrosis factor
- Treg
- regulatory T cells
- Copyright © 2022 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}