


Abstract
Neuroimaging studies of psychedelics have advanced our understanding of hierarchical brain organization and the mechanisms underlying their subjective and therapeutic effects. The primary mechanism of action of classic psychedelics is binding to serotonergic 5-HT2A receptors. Agonist activity at these receptors leads to neuromodulatory changes in synaptic efficacy that can have a profound effect on hierarchical message-passing in the brain. Here, we review the cognitive and neuroimaging evidence for the effects of psychedelics: in particular, their influence on selfhood and subject-object boundaries—known as ego dissolution—surmised to underwrite their subjective and therapeutic effects. Agonism of 5-HT2A receptors, located at the apex of the cortical hierarchy, may have a particularly powerful effect on sentience and consciousness. These effects can endure well after the pharmacological half-life, suggesting that psychedelics may have effects on neural plasticity that may play a role in their therapeutic efficacy. Psychologically, this may be accompanied by a disarming of ego resistance that increases the repertoire of perceptual hypotheses and affords alternate pathways for thought and behavior, including those that undergird selfhood. We consider the interaction between serotonergic neuromodulation and sentience through the lens of hierarchical predictive coding, which speaks to the value of psychedelics in understanding how we make sense of the world and specific predictions about effective connectivity in cortical hierarchies that can be tested using functional neuroimaging.
Significance Statement Classic psychedelics bind to serotonergic 5-HT2A receptors. Their agonist activity at these receptors leads to neuromodulatory changes in synaptic efficacy, resulting in a profound effect on information processing in the brain. Here, we synthesize an abundance of brain imaging research with pharmacological and psychological interpretations informed by the framework of predictive coding. Moreover, predictive coding is suggested to offer more sophisticated interpretations of neuroimaging findings by bridging the role between the 5-HT2A receptors and large-scale brain networks.
I. Introduction
Psychedelics constitute a class of drugs with origins in certain plants, animals, and fungi that have a long history of human use. Records indicate that their consumption in ancient cultural rituals—to achieve altered states of consciousness and spiritual insight—date back millennia (Wasson, 1978; Carod-Artal, 2015; Byock, 2018). Their inception into mainstream Western culture followed the introduction of mescaline in the early 1900s and the synthesis of a novel psychedelic, lysergic acid diethylamide (LSD), in 1938 (Nichols, 2016; Nichols and Walter, 2021) (The psychoactive effects of LSD were not discovered until 5 years later, in 1943). This latter compound, which mimics the activity of its naturally derived counterparts, attracted the interest of scientists and clinical researchers, who perceived similarities between the effects of psychedelics and psychosis and considered their potential therapeutic applications (Swanson, 2018). Unregulated mainstream use of psychedelics, particularly LSD, led to the counterculture of the 1960s that challenged indoctrinated societal values and political agendas, leading to restrictions on psychedelic research by regulatory institutions (Dyck, 2005). Shortly after, psychedelics were designated by the US government, then globally, as illicit substances—possessing no medicinal or therapeutic value—and human clinical psychedelic research ground to a halt (Nichols, 2016) (Preclinical animal and cell culture studies continued throughout the hiatus of human clinical psychedelic research, helping psychedelic science persist). However, interest in the neurobiological basis of psychosis continued, and reference to early psychedelic research led to studies in healthy adults before the turn of the millennium (Hermle et al., 1992; Strassman et al., 1994; Vollenweider et al., 1997).
In the new millennium, restrictions on psychedelic research proceeded to relax. Experimentation in the United States on healthy adults resumed, showing that psychedelics can produce lasting changes in a personal sense of meaning and spiritual significance (Griffiths et al., 2006), a notion resonant with their ancient use for existential guidance. In the past decade, there have been further developments in psychedelic research in the setting of psychiatry. For example, the application of psychedelics to cancer-related distress marked a crucial milestone, showing that psychedelics could produce a clinically significant reduction in depression and anxiety where alternative treatments failed (Griffiths et al., 2016).
A. Advances in Neuroscience
Meanwhile, neuroscience advanced considerably over the period of restricted psychedelic research. Techniques such as positron emission tomography (PET), developed in the late 1970s, and functional magnetic resonance imaging (fMRI) in the 1990s improved our understanding of the molecular and physiologic processes in the central nervous system (Cowan et al., 2000). The predominant conceptualization of functional neuroanatomy at the time characterized structure-function relationships within the brain in terms of segregated areas of brain activation, namely the principle of functional segregation (Zeki, 2005). Subsequently, neuroimaging addressed the coordination of distributed neuronal responses—among functionally segregated regions—in terms of functional integration (Zeki and Shipp, 1988; Friston, 2011; Razi and Friston, 2016).
Our current understanding of network activity in the brain is informed, at least in part, by the identification of resting-state brain networks, namely intrinsic networks that remain active in the absence of goal-directed attention (Raichle, 2015). Early research, in the resting state, found that the metabolic consumption of the brain at rest requires 20% of the body’s energy, and brain consumption during tasks increased this value a mere 5% (Fox and Raichle, 2007). Findings of this sort led to the notion that resting-state activity may be an important measure of functional integration and led to the eventual identification of multiple resting-state networks. Their role in mentation was confirmed by research showing that activity in resting-state networks could predict cognitive performance (Arieli et al., 1996; Tsodyks et al., 1999; Tavor et al., 2016) and identify psychopathology (Fox and Raichle, 2007).
The reintroduction of psychedelics to scientific research offers neuroscience new opportunities to use brain imaging to further our understanding of neurologic processes underlying brain function and mental health. Evidence for therapeutic outcomes following psychedelic treatment suggests that therapeutic outcomes will be accompanied by measurable changes in brain connectivity. Moreover, measurement of acute changes in connectivity under psychedelics may offer an opportunity to better understand the neural correlates of sentience and consciousness.
B. Importance of Brain Connectivity for Subjective Experience
The fundamental importance of brain connectivity for perception may appear obvious; however, a close analysis serves as a reminder that objective reality is quintessentially subjective (i.e., dependent on brain connectivity). This is demonstrated by brain disorders and injuries that impair perception, such as face perception, known as prosopagnosia (Wolland and Hagelsteen, 1991), and personality, described in the famous case of Phineas Gage (O'Driscoll and Leach, 1998). Damage to the brain can also alter representations of space and time (Buzsáki and Llinás, 2017), with dramatic consequences for perception, such as in cases of akinetopsia (Zeki, 1991). Disorders such as schizophrenia and psychosis provide complementary evidence, suggesting that altered patterns of brain connectivity can affect perception and selfhood (Griffa et al., 2019; Li et al., 2019). Research has also identified brain regions and connectivity underlying altered subjective experiences, including out-of-body experiences (Blanke and Arzy, 2005), spiritual experiences (Miller et al., 2019), and experiences of self-transcendence (Urgesi et al., 2010), self-awareness (Ham et al., 2014), and self-reference (Bréchet et al., 2018). Damage and disorders reliably indicate the function of brain connectivity in aspects of perception and behavior. These examples provide a strong parallel to the subjective effects of psychedelics, which may act similarly by altering circuitry.
C. Subjective Effects of Psychedelics
Classic psychedelics are a group of perception-altering drugs classified as entheogens (Ruck et al., 1979). Entheogens are substances that, when ingested, manifest an altered state of consciousness considered to have spiritual or existential significance (Winkelman and Hoffman, 2015). This differentiates them from related entactogen drugs such as 3,4-methylenedioxymethamphetamine (commonly known as ecstasy), known to induce effects related to empathy, such as emotional warmth and connectedness (Nichols, 1986; Bedi et al., 2010; Wagner et al., 2017; Byock, 2018).
Classic psychedelics exist in natural sources and chemically synthesized forms. The most widely recognized classic psychedelics belong to three main chemical classes: indoleamines [psilocybin and N,N-dimethyltryptamine (DMT)] (Psilocybin and DMT are often referred to as tryptamines, which is an indolamine metabolite of the essential amino acid tryptophan), phenylalkylamines (mescaline), and semisynthetic ergolines (LSD) (Nichols, 2004, 2018; Geyer et al., 2017). Psilocybin, considered the prototypical psychedelic, is the prodrug of the psychoactive compound psilocin that occurs naturally in the psychoactive Psilocybe and related genera of mushrooms, known colloquially as magic mushrooms. Like DMT, the psychoactive ingredient in the psychedelic brew ayahuasca, and mescaline, found in some species of cacti, psilocybin has a long history of use in medicinal and religious cultural contexts (Nichols, 2016). The contemporary counterpart to plant- and fungi-based psychedelics, LSD, which is technically semisynthetic [LSD-25 (Lysergsäure-diethylamid) was the 25th compound synthesized by Albert Hofmann in 1938 in a drug discovery program. The program investigated ergot fungus alkaloids, which include lysergic acid. LSD’s psychedelic effects were later accidentally discovered by Hofmann in 1943] has been of primary interest in modern psychedelic culture and research—though much recent attention has shifted to psilocybin. These entheogenic drugs share the capacity to alter the sense of identity and perspective in addition to causing hallucinations at sufficient doses (Swanson, 2018). Cognitive and metacognitive effects include shifts in attention and perspective, reframing of the relationship of self with the world and an altered perception of time and space, sense of transcendence, clarity and insight, positive emotional state and an increased sense of unity (Tagliazucchi et al., 2014; Lebedev et al., 2015; Preller et al., 2015; Zamaria, 2016; Byock, 2018). Psychedelic experiences have been characterized as mystical-type experiences (Griffiths et al., 2006) and peak experiences (Cummins and Lyke, 2013). However, no characterization subsumes and epitomizes the psychedelic experience more precisely than the fundamental alteration of self, known as ego dissolution.
D. Ego Dissolution
Ego dissolution is the definitive quality of the subjective experience resulting from ingesting a sufficient dose of a classic psychedelic. The term ego dissolution emerged in the mid-20th century to denote the effects of LSD and mescaline (Millière, 2017) and has been variously described as the disintegration of boundaries between one’s self and the world, a breakdown of one’s sense of self (Lebedev et al., 2015), cognition unbounded by models of the self (Letheby and Gerrans, 2017), a compromised sense of an integrated and distinct identity (Tagliazucchi et al., 2016), and antithetical to self-assuredness (Nour et al., 2016). Stanislav Grof—well known for his early studies of LSD and its effects on the psyche—similarly expressed ego dissolution as “an ecstatic-state, characterized by the loss of boundaries between the subjective and the objective world, with ensuing feelings of unity with other people, nature, the entire Universe, and God” (Grof, 1980).
Our theoretical treatment of ego dissolution reflects gestalt psychology, which views consciousness as an emergent whole greater than the sum of its parts. Similarly, we suggest ego dissolution as an emergent shift in consciousness demarcated as the change to the observer in contrast to other psychedelic perceptual effects characterized by the change to the observed.
Despite its high level of abstraction, ego dissolution has been suggested as a valid, measurable construct (Nour et al., 2016), with evidence suggesting that therapeutic outcomes are tied to its occurrence (Garcia-Romeu et al., 2014; Carhart-Harris and Goodwin, 2017; Roseman et al., 2018b; Yaden and Griffiths, 2020), although this is debated (Carhart-Harris and Goodwin, 2017; Olson, 2020).
Ego dissolution overlaps with other descriptions of psychedelic experience, such as the aforementioned mystical-type experiences (Griffiths et al., 2006) or peak experiences (Cummins and Lyke, 2013), and remains somewhat confounded with other facets of the psychedelic experience, such as the unitive state, which is described as a sense of oneness (Nour et al., 2016; Carhart-Harris et al., 2018a). The early onset of ego dissolution and overlap with peak experiences suggest that it is a prerequisite to access the range and richness of psychedelic experiences. Interestingly, ego dissolution is thought to be a binary state, an all-or-nothing phenomenon (Carhart-Harris and Friston, 2019), suggesting a dose-response related threshold that, once surpassed, realizes a phenomenological shift in consciousness. PET research endorses the notion of a nonlinear, dose-response relationship through investigation of plasma psilocin (the metabolite of psilocybin) occupancy in the brain and reported subjective intensity of experience (Madsen et al., 2019). However, the ego dissolution dose-response relationship is still being explored [see Safron (2020) for theoretical review of dose-response relationship] (Holze et al., 2021). It is influenced by external stimuli, particularly music (Kaelen et al., 2018), and some psychedelics, such as psilocybin, are reported to produce ego dissolution more reliably than others, such as LSD (Liechti, 2017). Following the onset of ego dissolution, psychedelic experiences are thought to catalyze spiritual and personal insight through introspection (Letheby and Gerrans, 2017; Malone et al., 2018; Dos Santos and Hallak, 2020). The ensuing insights can result in lasting therapeutic effects and transformation of self-related beliefs (Lyons and Carhart-Harris, 2018).
E. Measures of Subjective Effects of Psychedelics
Acute subjective effects of psychedelics have been measured in experimental studies using the hallucinogen rating scale (Strassman et al., 1994), the mystical experiences questionnaire (Maclean et al., 2012), and the abnormal mental states questionnaire, renamed the altered states of consciousness rating scale (OAV) (Studerus et al., 2010) and revised as the 5 dimensions of altered states of consciousness scale (5D-ASC) (Dittrich, 1998). This scale includes three primary dimensions: oceanic boundlessness or positively felt ego dissolution, dread of ego dissolution or negatively felt ego dissolution, and visionary restructuralization or visual alterations and altered meaning of precepts. The two other (secondary) dimensions are referred to as auditory alterations and a vigilance state. Subsequent psychometric evaluation of the OAV/5D-ASC revealed 11 factors that correlated with the original OAV scales: spiritual experience, changed meaning of percept, insightfulness, anxiety, impaired control and cognition, experience of unity, complex imagery, elementary imagery, blissful state, audio-visual synaesthesia, and disembodiment (Studerus et al., 2010; Nichols, 2016) [Oceanic boundlessness (i.e., ego dissolution) and visionary restructuralization (e.g., changed meaning of precepts) are factors that can be combined at a high level of the 5D-ASC construct hierarchy]. A more recent (and less widely explored) inventory—that validates ego dissolution as an independent construct—focuses on the phenomenon of ego dissolution and is aptly named the ego dissolution inventory (EDI) (Nour et al., 2016). Although the precise phenomenological classification or taxonomy of the subjective effects of psychedelic drugs are incomplete, the factors identified in the hallucinogen rating scale and 5D-ASC provide formal deconstructions of the subjective experience under classic psychedelics. Ego dissolution, however, as exemplified by the development of the EDI and emphasis in the 5D-ASC, is perhaps the best characterization of the primary subjective effect of psychedelics.
F. Psychedelic Subjective Experience and Therapeutic Outcomes
Interest in psychedelics has been renewed in part due to their tolerability (Dos Santos et al., 2018), safety (Studerus et al., 2010), and the provision of guidelines for safe administration (Johnson et al., 2008). This interest includes considering psychedelics as an adjunct to psychotherapy. Unlike traditional psychotherapy, which relies primarily on interpersonal discourse, psychedelic treatment relies on an introspective, endogenous experience as the patient typically lies silent, blindfolded, and in a state of rest (Nutt and Carhart-Harris, 2021). After the therapeutic session, patients are usually invited to a more traditional psychotherapeutic session coined integration, in which the patient and therapist, often a psychiatrist or psychologist, discuss the experience in the hope of deriving insights that may help cement positive life changes. Under supportive conditions, changes to patterns of thought and behavior—underlying psychiatric conditions such as anxiety, addiction, and depression—can show significant improvement from as little as one or just a few psychedelic sessions (Griffiths et al., 2011; Nichols et al., 2016; Letheby and Gerrans, 2017).
The transformative experience psychedelics produce offers a therapeutic tool that differs from traditional pharmacotherapy, which relies on neurochemical mechanisms to produce therapeutic effects. Although there is some evidence that certain psychiatric conditions, such as obsessive compulsive disorder (Majic et al., 2015), may respond more to the physiologic effects of psychedelics, the clinical literature suggests that the psychologic experience elicited by psychedelics is integral to transformative therapeutic change [Preclinical research also demonstrates therapeutic change and suggests a caveat to psychological mediation of therapeutic outcomes. Please refer to Separating Subjective Effects from Therapeutic Outcomes for our review] (Roseman et al., 2018b; Yaden and Griffiths, 2020; Nutt and Carhart-Harris, 2021). This is reflected in the dependence of lasting therapeutic outcomes upon the spiritual or personally meaningful quality of ego dissolution, here read as mystical experiences (Griffiths et al., 2011, 2016; Garcia-Romeu et al., 2014; Johnson et al., 2014; Bogenschutz et al., 2015; Carhart-Harris et al., 2016a; Ross et al., 2016; Hartogsohn, 2018; Roseman et al., 2018b; Russ, et al., 2018; Russ et al., 2019; Yaden and Griffiths, 2020).
Transformative outcomes from the psychedelic experience can be linked to the early 19th century psychologic theory known as filtration theory. Filtration theory suggests that information entering consciousness is winnowed by selection processes that filter the subconscious material (Marshall, 2005; Swanson, 2018). The term psychedelic—which originates from the ancient Greek word for psyche (ψυχή, “soul”) and dēloun (δηλου˜ν, “to make visible, to reveal”), translating to “mind manifesting”—was inspired by this theory to describe the inhibition of filtration processes in the brain, relaxing constraints on perception, emotions, thought, and sense of self (Osmond, 1957; Swanson, 2018). Under this theory, psychedelics may be considered to facilitate and synergize with the psychoanalytic process. In the patient-practitioner relationship, as in the relationship with oneself, some thoughts and feelings may be openly identified or disclosed while others are inhibited or avoided. The disinhibition of the normal constraints of mentation by psychedelics may allow activity—usually suppressed to the subconscious and constrained by conscious processes—to surface into conscious awareness. The description of this process accords with modern accounts of therapeutic applications of psychedelics. For example, the introspective experiences of addicts treated with psychedelics featured personally relevant emotional content, otherwise inhibited from conscious awareness, to surface, and the resulting nuanced personal insight led to reduced substance dependency (Malone et al., 2018). Similar accounts are also documented in the unmonitored use of psychedelics (Zamaria, 2016). Although the merit of insights derived under psychedelics warrants some skepticism (Letheby, 2016), evidence that ego dissolution targets the most abstract levels of processing—responsible for selecting channels that provide input to conscious awareness and their capacity to produce therapeutic insight (Hartogsohn, 2018)—endorses filtration theory as a rudimentary model of the action of psychedelics that is consistent with changes in hierarchical connectivity, as we will see below.
G. Ego Resistance
Psychedelic phenomenology has recently been described under the free energy principle (FEP). The FEP explains the innate tendency of living organisms to minimize surprise and will be explored later in detail. Elaborations of the FEP, applied to psychoanalysis, suggest that the tendency to minimize surprise can lead to maladaptive beliefs that are resistant to change (Carhart-Harris and Friston, 2019) and that psychedelics may dissolve recalcitrant beliefs. Psychedelic ego dissolution appears to feature in this (therapeutic) process by removing a hurdle and relaxing psychologic resistance. The mechanism of resistance-release is thought to be a central feature of ego dissolution associated with the therapeutic effects of psychedelics.
The term psychologic resistance is typically used to denote psychotherapeutic exchanges in which patients avoid particular uncomfortable topics. However, resistance may be fundamental to mental well-being and is taken more broadly as the trigger of ego defenses; namely, invoking beliefs and attitudes in response to perceived threats. Freudian psychoanalytic theory helps frame this process as the ego being maintained in a solved condition rather than an unsolved state (Solms, 2018). Freud described this preference as the source of resistance that elicits defense (Solms, 2018). Therefore, occurrences of resistance may be considered as the departure of the ego from the solved state. The ego, wishing to return to the solved state, may manifest self-beliefs that provide a form of emotional catharsis, or the best attempt at the solved state, in lieu of more authentic solutions, reminiscent of Freud’s wish fulfillment.
If the beliefs are maladaptive, they may lead to further experiences of resistance that produces incongruence, or dissonance, between the innate desire for positive emotional self-worth and the experiences. This incongruence or dissonance maintains the unsolved state and is similar to Festinger’s cognitive dissonance that describes the discomfort of conflict between action and cognition (Festinger, 1962). Here, the concept of dissonance is extended to describe the conflict between emotional self-worth and experiences that the ego resolves through beliefs.
Maladaptive beliefs are those which fail to reduce experiences of resistance and instead may perpetuate dissonance that in turn perpetuates resistance, leading to the development of rigid, ingrained patterns of thinking (Fig. 1). In more extreme cases, these unhelpful beliefs may develop into symptoms of psychopathological disorders; the exact presentation of disorder may resemble how the dissonance between experiences and emotion is resolved through the cognition. In the case of depression, a bullied individual may cathartically solve dissonance between damaging experiences and the innate emotional desire for self-worth by forming beliefs about their inferiority, thereby enabling the ego to enter a proxy solved state. In the case of narcissism, dissonance between emotional self-worth and conflicting experiences may be solved through belief in the inferiority of others. Alternately, a more straightforward instance of resistance may be evinced in the exaggerated emotional response to disappointment, characteristic of borderline personality disorder (Houben et al., 2018).
Elementary model of resistance leading to rigid or inflexible beliefs. Resistance that leads to ego defense may be accompanied by rationalizations in the form of higher-order beliefs. Higher-order beliefs that are maladaptive may lead to further experiences of resistance that evoke dissonance between emotions and experiences, which fortify maladaptive beliefs leading to belief rigidity.
Although dissonance between self-worth and experience is also underlined in this scenario, resistance can also be seen as resistance to experience (and implicitly, belief updating). In all cases, resistance leads the ego into an unsolved state that is dissonant and can form maladaptive beliefs that are characterized by further resistance. Psychedelics target this psychologic ego resistance and introduce a permissive flexibility in cognition that dissolves rigid patterns of thinking and is reflective of the psychoanalytic process [Working through resistance in psychotherapeutic exchanges between practitioner and patient and in successful psychedelic ego dissolution experiences contributes to lasting therapeutic changes (LaFarge, 2012; Roseman et al., 2018b; Russ et al., 2019)]. From the perspective of FEP, psychedelics dissolve precise belief states where that dissolution can be quantified as an increase in entropy or decrease in precision (Carhart-Harris, 2018). Technically, the precision of a (probabilistic or Bayesian) belief corresponds to its negative entropy. This dissolution has been connected to the personality trait of openness (Lebedev et al., 2016) and is arguably antithetical to resistance. The dissolving of resistance through successful ego dissolution, therefore, appears to be commensurate with the relaxation of precise (priors) beliefs and ego defenses that enable deeply ingrained patterns of thoughts to be critically evaluated and revised in therapeutic settings (Carhart-Harris et al., 2014a). However, effective ego dissolution requires a mindset willing to forgo resistance to the deconstruction of self and beliefs. When an individual has traits of absorption, openness, acceptance, and surrender, they are more likely to have positive experiences, whereas preoccupied or apprehensive mindsets indicate possible adverse reactions (Aday et al., 2021). For example, preoccupation or an attitude of surrender at the time of psilocybin ingestion explained anxious and positive responses to psilocybin experiences (Russ et al., 2018, 2019). These findings reflect the parallel between the optimal state of mind for psychedelic experiences and meditative (i.e., mindful) state of mind.
H. Psychedelics and Meditation
The disarming of ego resistance is also elemental to the practice of nonjudgemental awareness through meditation. Like the use of psychedelics, meditation is another practice rooted in spirituality, dating back millennia. Meditation is a nonpharmacological intervention with a wealth of evidence supporting positive therapeutic outcomes (Keng et al., 2011). Meditation and psychedelics share similar processes of dissolving the self and attaining insight, and both demonstrate the capacity to inspire insights leading to personal growth and improved well-being (Griffiths et al., 2018). Similarities in the brain connectivity patterns underlying states of meditation and psychedelic states have been reported, particularly in resting-state networks (Brewer et al., 2011; Carhart-Harris et al., 2013; Lebedev et al., 2015; Heuschkel and Kuypers, 2020). Recent studies of psychedelics and meditation suggest that the practice of meditation may be beneficial to the psychedelic experience under psilocybin by reducing anxiety (Smigielski et al., 2019a). Similarly, psychedelics may enhance meditation and spiritual practice.
Single doses of psilocybin administered to expert meditators were found to improve depth of meditation and long-term well-being beyond the beneficial effects of exclusive long-term meditation practice (Smigielski et al., 2019a,b). Another study exploring psilocybin—in conjunction with (nonsecular) spiritual activities—suggested that their combination boosts lasting prosocial attitudes and sustains personal spiritual practice (Griffiths et al., 2018). Further studies suggest a complementary effect of psilocybin and mindfulness meditation in the treatment of depression (Heuschkel and Kuypers, 2020). Reference to meditation in psychedelic research is both common and valuable by helping to define similarities and differences between psychedelic phenomenology and brain connectivity. One of the most prominent differences is the phenomenological richness of the psychedelic experience; see Millière et al. (2018) for an in-depth review and exploration of their potential synergy; Heuschkel and Kuypers (2020) for a review of meditation complementing psilocybin in the treatment of depression; and (Payne et al., (2021) for a review of clinical synergies. The effectiveness of meditation through nonjudgement and nonresistance may reflect an innate self-corrective tendency of the mind toward the reduction of resistance to engender well-being. This tendency often fails under ordinary control of the ego and appears to be facilitated in a similar fashion by both psychedelics and meditation.
I. Outline of the Review
A comprehensive understanding of psychedelics requires consideration of historical, cultural, sociological, spiritual, philosophical, psychologic, and physiologic perspectives. Our focus will be on the effects of psychedelics on brain connectivity and the mediation of subjective effects, particularly ego dissolution. Psychedelic studies that used noninvasive brain imaging, including fMRI, PET, electroencephalography (EEG), and magnetoencephalography (MEG), are surveyed with a special emphasis on how modeling and estimating brain connectivity may shed light on the neural mechanisms that underlie the psychedelic experience.
The remainder of this review begins with the molecular and pharmacological action of psychedelics and their effect on cognition. The effects of psychedelics on neurotransmitter receptors are central to formulations of their action under hierarchical models of brain connectivity. These models explain the ability of psychedelics to increase cognitive flexibility and speak to the importance of context and administration setting. Building on the pharmacological action at the neuronal level, we examine hierarchically organized brain regions and networks with functions spanning the faculties of perception.
The role of neuronal populations in subcortical structures is reviewed. In particular, the thalamus is a highly connected structure attributed with a gating function that filters information exchanged with the cortex (Torrico and Munakomi, 2019). Its function under psychedelics is well aligned with filtration theory and suggests psychedelic-induced altered information processing open novel thalamic connectivity pathways to the cortex. Next, we consider medial temporal lobe (MTL) connectivity. MTL regions are involved in memory and emotion (Steriade and Llinas, 1988; Stein et al., 2000; Voets et al., 2015) and may share an association with ego dissolution and the therapeutic engagement of emotion under psychedelics (Kraehenmann et al., 2015a; Carhart-Harris et al., 2017). Moving to the cortex, altered “resting-state” brain connectivity across distinct networks—typically involved in abstract and executive functions—are considered. Resting-state connectivity at the top of the cortical hierarchy underlie consciousness and sentience. Psychedelic connectivity change here may deconstruct belief constructs that play a role in our sense of identity and identification with the world. This deconstruction may diminish top-heavy hierarchical balance of brain organization and, in therapeutic contexts, afford alternate hypotheses underwriting beliefs about the self and the world. Finally, integration of these systems and unifying theories of psychedelic effects are explored before considering future research directions.
J. Pharmacology, Plasticity, and Context
Classic psychedelics such as LSD, psilocybin, mescaline, and DMT are characterized by agonist activity at the serotonin 5-hydroxytryptamine (5-HT) 2A receptor (5-HT2AR) (Nichols, 2016; Dinis-Oliveira, 2017). The serotonergic system is one of the earliest evolved neurotransmitter systems and innervates a diversity of receptors (Hoyer, 2007). The dorsal and median raphe nuclei of the brainstem synthesize 5-HT and project to subcortical, limbic, and neocortical parts of the brain (Azmitia and Gannon, 1986; Wilson and Molliver, 1991; Jacobs and Azmitia, 1992). 5-HT receptors are classified into 14 subtypes within 7 groups and include one serotonin transporter protein, 5-hydroxytryptamine transporter (5-HTT), that typically transports the neuromodulator serotonin from the synaptic cleft to the presynaptic neuron (Amara and Kuhar, 1993; Serretti et al., 2006; Deco et al., 2018). The abundant 5-HT2A receptor expression throughout the cortex suggests the involvement of 2A subtype in higher cognitive and integrative functions (Hensler, 2012). Psychedelic compounds share a common affinity for the 5-HT2AR (Nichols, 2012). 5-HT2AR can mediate pathologies such as stress and depression (Mann, 1999), and stimulation of 5-HT2AR, among other receptors, may help explain the potential for psychedelics to mediate behavioral change (Carhart-Harris and Nutt, 2017; Preller et al., 2018).
The binding kinetics and occupancy of the 5-HT2AR predict the subjective effects of classic psychedelics (Glennon et al., 1992; Madsen et al., 2019). Although the molecular structures of psychedelics vary [For a detailed review of the neurobiology of psychedelic drugs, see Vollenweider and Preller (2020)]—and many act at non–5-HT receptors [LSD acts at dopaminergic and adrenergic receptor sites (Nichols, 2004)] (Tylš et al., 2014; Nichols, 2016)—the importance of the 5-HT2AR has been established using antagonists such as ketanserin, which blocks the receptor and inhibits subjective effects (Quednow et al., 2012; Preller et al., 2018a,b). Subjective psychedelic effects may also be precluded by the rapid desensitization of the 5-HT2AR following exposure to psychedelics (Damjanoska et al., 2004). The 5-HT2AR is a metabotropic serotonin receptor located postsynaptically on the apical dendrites of layer 5 pyramidal neurons (Weber and Andrade, 2010). Pyramidal neurons act as the primary output from cortical regions projecting to distal cortical and subcortical regions (Spruston, 2008). Accordingly, postsynaptic 5-HT2A receptors are profuse in the cortex and are especially abundant in the prefrontal cortex (Hall et al., 2000; Saulin et al., 2012; Celada et al., 2013b). A smaller proportion are located presynaptically on thalamocortical afferents projecting to the neocortex (Marek, 2018). 5-HT2A receptors exist in several types of excitatory and inhibitory neurons and nonneurons (e.g., glial cells) within the brain; however, they are expressed at their highest densities within excitatory pyramidal cells (Jakab and Goldman-Rakic, 1998; Meller et al., 2002). Layer 5 pyramidal neurons require agonist activity to reach an action potential threshold to communicate with other neurons (Varley et al., 2019). Psychedelic binding to the 5-HT2AR only depolarizes a small percentage of deep cortical cells; the majority do not (Andrade, 2011). Only small ensembles of cells, about 5%–10%, have demonstrated direct activation by psychedelics. These ensembles represent a mixture of excitatory neurons and specific subtypes of interneurons, glia, and astrocytes (Martin and Nichols, 2016). Moreover, at least two different subpopulations of excitatory neurons exist in the cortex. Five percent to 10% of excitatory neurons within these subpopulations express significantly more 5-HT2A receptors than the remaining 90%–95%. These 5%–10% are the excitatory neurons that directly depolarize under psychedelics (Martin and Nichols, 2016; Vollenweider and Preller, 2020). 5-HT2ARs are also found in GABAergic interneurons of the cortex and subcortical structures (Celada et al., 2013b). GABA is a primary inhibitory neurotransmitter of the nervous system. LSD is a powerful partial agonist at cortical and subcortical 5-HT2A receptors of GABAergic interneurons (Marek and Aghajanian, 1996; Inserra et al., 2021a,b). Psychedelics also stimulate 5-HT1A receptors (Strassman, 1996; Carter et al., 2005; Pokorny et al., 2016) and 5-HT2B receptors (Kozlowska et al., 2021). Stimulation of the 5-HT1A receptor and GABA interneurons by LSD both inhibit and disinhibit prefrontal pyramidal cell activity (Lladó-Pelfort et al., 2012; Celada, et al., 2013a; Lladó-Pelfort et al., 2018) and show downstream effects on dopaminergic (Vollenweider et al. 1999) and glutamatergic systems (Mason et al., 2020; Vollenweider and Preller, 2020). It is evident that the global lowering of the resting membrane potentials of layer 5 pyramidal cells is insufficient to explain how psychedelics influence connectivity and circuitry function. However, despite their partially inhibitory mechanisms, psychedelics like LSD result in a net-excitatory effect on layer 5 pyramidal neurons (Martín-Ruiz et al., 2001; Puig et al., 2003; Lambe and Aghajanian, 2006) [The net excitatory effects of psychedelics contrast the effects of anesthetic drugs, such as propofol, that increase inhibition in neural connections and reduce consciousness (Rudolph and Antkowiak, 2004)]. Variation in changes to profiles of neuronal ensembles inhibitory-excitatory activation across brain regions may underwrite the connectivity changes observed between brain regions under psychedelics (Martin and Nichols, 2016; Vollenweider and Smallridge, 2022).
K. Free Energy Principle and 5-HT2A Receptor Agonist Activity
Altered serotonin transmission has been linked to neuronal responses to unpredicted or surprising events (Matias et al., 2017). Learning from unexpected outcomes is important for the survival of living organisms and confers a clear evolutionary advantage. The FEP provides a framework to understand responses to surprise—and the influence of psychedelics. The FEP posits that all living systems survive by minimizing the difference between sensory input and predictions based upon an internal (generative) model of the world (Friston, 2009, 2010). This difference is described as self-information, surprisal, or more simply, surprise, which can also be written as a (log) model evidence in Bayesian statistics (for which the free energy is an upper bound). This means that minimizing free energy is the same as maximizing model evidence, sometimes known as self-evidencing (Hohwy, 2016).
A popular way of understanding the minimization of free energy is in terms of minimizing prediction error or, more precisely, precision weighted prediction error. Precision refers to the predictability of some random variable. This means that prediction errors should only be treated as surprising if they convey precise information. In other words, a prediction error is not very useful if something is not predictable. The encoding of predictability or precision is thought to be mediated by synaptic efficacy: in other words, the sensitivity of gain of postsynaptic responses to predictions and prediction errors. Because psychedelics have a selective and profound effect on postsynaptic gain, they can be understood as rebalancing the influence of sensory evidence and prior beliefs during belief updating (mediated by neuronal message passing) in response to sensory input.
In this framework, 5-HT2AR modulation by psychedelics is understood to alter the sensitivity to prediction errors at various hierarchical levels in the cortical hierarchy. In brief, psychedelics are thought to relax the precision of high-level prediction errors so that lower level (e.g., sensory) prediction errors have a greater influence. This allows multiple hypotheses and explanations for sensory input to be entertained by the brain, endowing the implicit self-evidencing with a greater flexibility (Carhart-Harris, 2018). Neurophysiological measures can express this in terms of a greater entropy or complexity of neuronal activity that underwrites belief updating.
In probability theory, entropy is a measure of uncertainty and, in some settings, complexity. Agonist activity at 5-HT2AR is thought to increase neuronal entropy (Herzog et al., 2020). It serves as a metric of neuronal activity that has been used to quantify the influence of psychedelics via modulation of 5-HT2AR neurotransmission (Carhart-Harris et al., 2014a; Bouso et al., 2018; Carhart-Harris, 2018). Recent findings suggest that the increase in the entropy of neuronal activity is topographically organized, exhibiting increases in some brain regions and decreases in others under psychedelics (Herzog et al., 2020). The varied expressions of entropy across the brain suggest that complex patterns of cellular excitation-inhibition are induced by psychedelics. Psychedelic-induced entropy changes in the brain have also been read as a state of increased plasticity (Carhart-Harris and Nutt, 2017; Ly et al., 2018) manifesting as cognitive flexibility (Matias et al., 2017) and an increased repertoire of possible brain states (Atasoy et al., 2017). The psychedelic enabling of neural plasticity is a key mechanism used to explain the therapeutic effects of psychedelics (Inserra et al., 2021a).
Under the FEP, psychopathology is usually interpreted in terms of false inference. For example, inferring something is present when it is not (e.g., hallucinations and delusions) or inferring something is not present when it is (e.g., agnosia and neglect syndromes). Usually, this is interpreted as inference under aberrant precision. In other words, too much or too little precision is afforded by sensory prediction errors, relative to prediction errors higher in the hierarchy that mediate prior beliefs. If prior precision is unduly high, inference about states of affairs causing sensations may proceed under inflexible priors that are maladaptive (Carhart-Harris and Friston, 2019). 5-HT2AR modulation under psychedelics is thought to relax overly precise prior precision and enable the revision of maladaptive priors through experience-dependent learning (e.g., of the sort that may be leveraged in psilocybin-assisted psychotherapy).
The ensuing effects on 5-HT2AR signaling has been suggested to mediate active coping with distress and support cognitive-emotional revision (Carhart-Harris and Nutt, 2017). The notion of inflexible, maladaptively precise priors is compatible with psychoanalytic theory in which rigid priors may be thought of as manifesting strong resistance. Overlap between the distributions of high-density 5-HT2AR and the cortical regions considered to underlie self-related beliefs also speaks to the notion that psychedelics may target the substrate of high-order (Bayesian) beliefs (Carhart-Harris et al., 2014a; Beliveau et al., 2017; Komorowski et al., 2017). This formulation offers a neuronally plausible account of neuromodulation and plasticity in sentient behavior that is supported by the FEP (Griffiths et al., 2011; Tagliazucchi et al., 2014; Letheby and Gerrans, 2017; Carhart-Harris, 2018). Increased flexibility following psychedelic-induced plasticity also explains the context sensitivity that individuals exhibit under psychedelics (Carhart-Harris et al., 2015).
L. Psychedelics Amplify Set and Setting
Contextual factors such as states of the environment, individual traits, and psychologic variables are crucial in shaping the psychedelic experience (Carhart-Harris et al., 2018b, 2018c; Tylš et al., 2016; Carhart-Harris and Nutt, 2017; Roseman et al., 2018b). Some early experiments with psychedelics created conditions that elicited adverse and traumatic reactions from participants (Carhart-Harris et al., 2018b). Such outcomes may have contributed to early Western characterization of psychedelics as psychotomimetic, referring to properties that mimic psychosis (Müller et al., 2018). The evolving understanding of psychedelics nuanced this terminology, later describing them as hallucinogens (Nichols et al., 2016). However, neither term recognized their potential merit in therapeutic settings, leading to the evolution of the modern term psychedelic. This term encompasses the mind-manifesting nature of psychedelics as “nonspecific amplifiers of consciousness” (Grof, 1980) and speaks to the importance of set and setting. This is consistent with the effect of psychedelics on the synaptic encoding of precision in terms of synaptic efficacy or postsynaptic gain, where increases in the sensitivity to prediction errors at various levels in cortical hierarchy has been associated with attentional selection or the instantiation of a particular attentional set (Feldman and Friston, 2010; Ainley et al., 2012; Ainley et al., 2016; Fotopoulou and Tsakiris, 2017).
Psychedelics, through 5-HT2AR agonist activity, may amplify exogenous influences of the setting and endogenous influences of attentional set. The ensuing state of plasticity can be likened to feeling “lost” (e.g., in the forest), which, depending on mindset and setting, can manifest opposing (emotional) responses (Fig. 2). Recognition of the influence of set and setting has led to the development of guidelines for the safe administration of psychedelics. These include measures for screening, preparation, and supportive environmental conditions (Johnson et al., 2008). Under such guidelines, subjects typically respond well to even high psychedelic doses, evidenced by the large number of modern clinical trials and applications of psychedelics. To date, there are no reports of serious adverse reactions (Johnson et al., 2018). However, even in supportive settings, challenging experiences should be anticipated. These may be viewed as part of the therapeutic self-discovery process—under the premise that challenging contents and experiences manifest from the disinhibition of ego restraints—that usually precludes therapeutically relevant emotion and cognition from entering conscious awareness (Carbonaro et al., 2016).

Lost in the bush (forest). This schematic illustrates the opposing psychologic responses to psychedelic-induced uncertainty dependent on the context of mindset and setting. Adapted from a photo taken at the rainforest gallery, Warburton, Victoria, Australia.
However, unregulated use of psychedelics remains potentially hazardous: phenomenological studies of unregulated ingestion show that users generally prepare both mindset and environmental settings before the dose intake by ensuring supportive environmental conditions generally (Zamaria, 2016). This preparation reflects the tenets of psychedelic-assisted therapy and the customs of ancient ritualistic use that involve care and preparation.
Set and setting are crucial factors when explaining negative psychotomimetic and positive therapeutic reactions to psychedelics. Identifying the influence of set and setting is an important subfield of research (Fig. 3).

Extrapharmacological model. Traits and setting influence mindset prior to administration. Mindset, setting (environment), and dosage contribute to the psychedelic experience (state) and subsequent therapeutic outcomes. Purple-colored boxes represent psychedelic influenced states. Adapted from extra-pharmacological model by Carhart-Harris and Nutt (2017).
Manipulation and measurement of the impact that preparatory methods and environmental stimuli have upon psychedelic experiences has yet to be widely explored. For example, the influence of music on the psychedelic experience has been found to affect the induction of ego dissolution and insightfulness (Kaelen et al., 2018) and enhance these experiences (Barrett et al., 2017). Other visual manipulations—such as comparing brain activity between eyes-closed and eyes-open states—speaks to the mediation of the psychedelic experience by exogenous stimuli (Mediano et al., 2020). Furthermore, recent studies of expert meditators suggest that meditation may be an effective preparation for psychedelic experiences (Smigielski et al., 2019a). Meditators are trained to cultivate an open, unattached, and nonjudgemental awareness of internal mentation. This practice of nonresistance and release resembles the clinical guidelines in psychedelic therapy (Wolff et al., 2020). Meditation training may reduce challenging experiences in the state of uncertainty accompanying ego dissolution and may offer a promising mode of preparation for therapeutic applications. The link between the quality of psychedelic experiences and therapeutic outcomes (Roseman et al., 2018b; Russ et al., 2018; Russ et al., 2019) suggests that mindset preparation through meditation may be an effective means to moderate anxiety. Moreover, engagement with meditation may predict the quality and therapeutic utility of psychedelic ego dissolution.
Although rare, reports of adverse reactions to meditation exist (Farias et al., 2020). Research is required to determine whether adverse reactions to meditation predict adverse reactions to psychedelics. Modern screening practices implemented in clinical studies prioritize screening for personal or family history of psychosis-related disorders (schizophrenia or bipolar disorder). However, this does not entirely mitigate the risk associated with psychedelic experiences. Psychedelics reduce vigilance (Bayne and Carter, 2018) and create impairment in memory tasks while paradoxically enhancing the vividness and recollection of autobiographical memories (Healy, 2021). Combined with their capacity to enhance suggestibility (Carhart-Harris et al., 2015), psychedelics' effects on memory may indicate a risk of false memories.
A related concern is that psychedelic experiences may be overinterpreted as offering some truth about the nature of existence (Móró et al., 2011; Letheby, 2016). These revelations may justifiably be met with skepticism and may be explained as spiritual bypassing. (Masters, 2010; Carhart-Harris and Friston, 2019). Spiritual bypassing defines the interpretative leaps an individual may take in the attempt to formulate explanations from the uncertainty or surprise that comes from profound experiences. In the absence of explanation, attempts to explain away (i.e., reduce) uncertainty can lead to ungrounded beliefs that deviate from reason and the tenets of scientific thinking. A similar effect is noted in acute psychosis, in which an overload of information is expressed as accounts of delusional thinking and beliefs (Carhart-Harris and Friston, 2019).
Mindset and setting at the outset of psychedelic experiences may predispose a subject to spiritual bypassing. For example, subjects administered psilocybin in the 1962 “Good Friday experiment” framed their interpretation of the experience according to their prior religious beliefs and the religious setting (Doblin, 1991). Development of preparation and integration programs may be required to guard against radical, secular, or superstitious interpretations. These programs may require cultural and subject-specific sensitivity that considers the intent of psychedelic use and evaluates the potential influence of the subject's prior beliefs on the psychedelic experience. We estimate that the need for such programs will depend on the intention of the psychedelic experience and agree with other authors that approaches developed from depth psychology or the nonsecular wisdom found in Buddhism may be appropriate (Carhart-Harris and Friston, 2019). We also suggest Maslow's hierarchy of needs as an optimal framework to understand the target of psychedelic use and measure how psychedelics facilitate personal growth. Psychosocial preparation and subsequent integration that focus on the meaning and utility of the experience to personal development during participant-led integrative discourse may be formulated on this model.
Overinterpretation may also be proportional to the clinical populations for whom psychedelics are recommended. Those with excessively rigid thinking, defined under filtration theory, may be at reduced risk compared with those with imaginative and superstitious thinking. The risk of overinterpretation may also be increased when no therapeutic intent is determined prior to psychedelic use. However, this may also depend on context and requires further investigation. Although ameliorating interpretative leaps may require preparation and guidance, guarding against false beliefs should not overshadow the broad pattern of successful therapeutic outcomes.
The evidence of contextual influences under psychedelics offers another research opportunity to explore neural plasticity and cognitive flexibility relating to 5-HT2AR agonist activity. Investigating different combinations of set and setting may help direct interventions and inform clinical procedures related to set and setting. How 5-HT2AR–dependent changes in distributed neuronal activity are mediated by changes in connectivity between subcortical systems and the cortex is based on insights obtained from functional neuroimaging of healthy adults as reviewed next.
M. Brain Connectivity
Different parts of the brain are connected to realize certain functions such as cognition, perception, and decision making. The subjective effects of psychedelics on perception suggest that psychedelics may preferentially target brain regions and connections involved in associative functions related to sentience and perception. This conjecture is supported by the high densities of 5-HT2AR expressed throughout associative brain regions (Jakab and Goldman-Rakic, 1998). However, cortico-subcortical networks and limbic structures also express 5-HT2AR. 5-HT2AR–mediated change of subcortical circuitry by psychedelics alters their functions and may account for aspects of the psychedelic subjective experience. Evidence from imaging studies can, in principle, be used to test hypotheses for the mechanism of action of psychedelics.
N. Subcortical Connectivity
1. Thalamic Signal Filtration
The early understanding of psychedelic mechanisms—as described in filtration theory—suggests that consciousness is winnowed by selection processes that exclude subconscious material (Marshall, 2005; Swanson, 2018). The thalamus is thought to serve such a function by filtering sensory information reaching the cortex. The thalamus is an information filter and relay hub located in the center of the brain, with projections in all directions, responsible for conveying signals between the brain and body (Torrico and Munakomi, 2019). The thalamus is also involved in the regulation of consciousness level, including sleep, wakefulness, alertness, and sensorimotor arousal (Torrico and Munakomi, 2019). It shares connectivity profiles with limbic structures (Steriade and Llinas, 1988; Stein et al., 2000; Voets et al., 2015) and is implicated in medial temporal lobe epilepsy (Barron et al., 2014), which produces psychedelic-like sensory illusions and déjà vu experiences. Disruptions to the thalamus—within the cortico-striato-thalamo-cortical loop—speak to the role of the thalamus in schizophrenia (Behrendt and Young, 2004; Halberstadt, 2015), sometimes referred to as a cognitive dysmetria. Certain portions of the thalamus (e.g., the pulvinar) have been associated with setting the precision of cortical message passing, of the sort implicated in selective attention and feature ground separation in the visual system (Kanai et al., 2015).
Commonalities—between the positive symptoms of schizophrenia and hallucinations produced under psychedelics—suggest that the cortico-striato-thalamo-cortical circuitry may be involved in hallucinations and mediate increased bottom-up connectivity from the thalamus to the cortex (Geyer and Vollenweider, 2008; Tylš et al., 2016). Examinations of functional connectivity (FC), under LSD, confirm an increase in correlations between the thalamus and cortical hubs (Tagliazucchi et al., 2016; Müller et al., 2018). FC analysis under LSD also found increases in correlated activity between the thalamus and insula and fusiform gyrus, thought to be related to the perceptual effects of psychedelics (Müller et al., 2017). Functional connectivity refers to correlations between neuronal activity in different systems that is mediated by effective connectivity, namely the directed influence that one neuronal system exerts over another.
Dynamic causal modeling (DCM) for resting-state fMRI (Friston et al., 2014; Razi et al., 2015; Razi et al., 2017) quantified the effective connectivity of the thalamus connectivity under LSD. The results showed increased effective connectivity from the thalamus to the posterior cingulate cortex (PCC) and decreased top-down connectivity from the PCC to the thalamus (Preller et al., 2019). These findings support the general notion of psychedelic-enabled increase in bottom-up effects on the cortex (Vollenweider and Preller, 2020). This is consistent with a relaxation of prior precision in high levels of cortical hierarchies, to which the thalamus projects. In short, the thalamus appears to play a key role in enabling ascending or bottom-up connectivity under psychedelics by opening a “thalamic filter” (Fig. 4). Further review of thalamic connectivity changes under psychedelics is featured in Unifying Neuroimaging Evidence of Psychedelic Mechanisms.

Opening the thalamic filter under psychedelics. Flatheads represent top-down inhibition of bottom-up signals, and arrowheads represent uninhibited signals. Reduced top-down inhibition from the cortex enables increased bottom-up connectivity to the cortex.
O. Midbrain Connectivity
1. Medial Temporal Lobe
The MTL regions involve limbic structures that are associated with emotion and memory, which may relate to the therapeutic effects of psychedelics (Carhart-Harris, 2007; Ritchey et al., 2019). Cortical midline structures such as the PCC, anterior cingulate cortex (ACC), and medial prefrontal cortex (mPFC) are connected to MTL structures, including the parahippocampus (PHC), retrosplenial cortex (RSC), and amygdala (AMG) (Milad and Quirk, 2002; Etkin et al., 2011; Andrews-Hanna et al., 2014). Like the thalamus, increased signaling from the MTL following psychedelics is thought to affect the influence of MLT on higher associative areas and subsequently conscious awareness (Carhart-Harris et al., 2014a). Abnormal MTL activity can produce dreamy states and a sense of depersonalization, resembling psychedelic subjective effects (Halgren et al., 1978; Lemche et al., 2016). This suggests that psychedelic-induced alterations of the MTL may produce similar effects. However, the MTL’s importance in memory and emotion point to its connectivity with other cerebellar and cortical regions that may play a role in acute and lasting therapeutic effects. Moreover, decoupling between cortical regions and the MTL has been suggested to play a role in ego dissolution (Lebedev et al., 2015). Understanding connectivity changes of the MTL under psychedelics may help discern its contribution to subjective and therapeutic effects. Partitioning of the MTL into its constituent systems, including the AMG, PHC and RSC, provides a more granular view of their role under psychedelics.
2. Amygdala
The AMG is involved in perception and emotional processing (Phelps and LeDoux, 2005; Kraehenmann et al., 2015a) and identifying emotionally salient stimuli. Activity of the AMG can be summarized as eliciting experiences of heightened emotional arousal mediated by top-down connectivity. The AMG comprises substructures with distinct functions and connectivity to various brain regions. Although few psychedelic studies report the change to substructure connectivity of the AMG—likely due to the challenges of accurately locating and aligning group connectivity changes—investigating the differential roles of amygdala substructures under psychedelics for their role in emotion and therapeutic outcomes is crucial to illustrate the neural mechanisms of psychedelics accurately. For example, the basolateral amygdala is the largest substructure and projects to layer V pyramidal neurons (Marek, 2018). An increase in basolateral AMG pyramidal and stellate neurons is implicated in chronic stress (Lu et al., 2021), and its projections to the cortex are cited in preclinical research of addiction (Roura-Martínez et al., 2020). Its connectivity with the prefrontal cortex is also important to extinction learning (Quirk and Mueller, 2008; Herry et al., 2010; Milad and Quirk, 2012; Baldi et al., 2021), and dysfunction of this connectivity marks fear disorders such as PTSD (Rauch et al., 2006; Shin et al., 2006). Under psilocybin, the basolateral AMG demonstrates an elevated expression level of c-Fos (a marker of neural activity) (Davoudian et al., 2022) and is implicated in mice under nonclassic psychedelics such as 3,4-methylenedioxymethamphetamine (Glavonic et al., 2022). However, destruction of the basolateral AMG had a negligible effect on prefrontal 5-HT cortical excitation in preclinical research. The negligible effect suggests that the AMG is not a bottom-up pathway to prefrontal serotonergic excitation in the rat brain (Marek et al., 2001). Future psychedelic work may also wish to investigate the differential roles of amygdala substructures under psychedelics for their role in emotion and therapeutic outcomes.
Whole AMG analyses indicate top-down AMG connectivity may play a role in mediating visual salience. This emotional processing circuitry can be sensitive to the effects of psychedelics. The AMG receives top-down projections from midline cortical regions, like the ACC with indirect connections to the mPFC (Etkin et al., 2011; Robinson et al., 2013; Kraehenmann et al., 2015b). For example, top-down connectivity between the ventral ACC and AMG influences emotional regulation and reappraisal (Etkin et al., 2011). AMG circuitry is thought to respond to threats to both beliefs and physical safety (Kaplan et al., 2016). This implicates the AMG in ego resistance and defenses that may require resolution to produce ego dissolution experiences. However, the AMG’s association with anxiety and fear should not overshadow its more general role in the regulation of emotions (Price et al., 2009) and detection of emotional salience (Adolphs, 2010; Santos et al., 2011). AMG activation does not determine the emotional valence of subjective experiences. For example, increased AMG activation was found after treating (treatment-resistant) depression with psilocybin (Roseman et al., 2018a). This effect was attributed to acceptance and emotional reconnection reported by patients (Watts et al., 2017; Roseman et al., 2018a). Conversely, another psilocybin study associated positive mood change in healthy subjects, with decreased AMG response measured acutely (Kraehenmann et al., 2015a). This suggests that AMG deactivation was associated with a reduction in negative emotion and aligns with other psilocybin investigations showing a reduction of cerebral blood flow in the AMG that correlates with a reduction of depressive symptoms (Carhart-Harris et al., 2017).
Such contradictory associations between AMG activation and behavioral outcomes are evident throughout imaging literature. The reduced structural volume of the AMG in meditators is thought to represent decreased negative affect (Gotink et al., 2018), whereas the same finding in borderline personality disorder is believed to indicate decreased emotional regulation (Weniger et al., 2009). These findings suggest that the structural volume and activation of the AMG in isolation do not disambiguate the quality of psychologic experience. More accurate associations between the AMG and behavior require attention to the subject’s experience during measurement and the time when AMG activity is assessed: the AMG appears to deactivate under acute psychedelic effects in response to negative emotional stimuli in participants in supportive conditions. Increased activation of the AMG, however, appears in the period following clearance of the drug.
Understanding top-down connectivity may help disambiguate the changes in AMG relating to psychologic function and therapeutic outcomes. Top-down connectivity generally has an inhibitory influence on bottom-up projections. Disruption caused by psychedelics to top-down connectivity is seen clearly in changes in the visual-limbic-prefrontal network involved in detecting visual threat cues. This circuitry is thought to evince increased top-down influences in pathologic conditions (Disner et al., 2011). Under acute effects of psilocybin, reduced top-down threat processing in healthy adults is demonstrated by decreased effective connectivity from the AMG to the primary visual cortex (Kraehenmann et al., 2015b). Cortical connectivity to the AMG also mediates emotional threat processing, and LSD reduces AMG responses to fearful faces in healthy adults under acute effects (Mueller et al., 2017).
As previously mentioned, an opposite pattern of increased AMG activity is seen using similar task conditions 1 day after psilocybin in the (right) AMG (Roseman et al., 2018a), whereas individuals with borderline personality disorder—which is characterized by over-reactive responses to threat—show hyper-AMG activation to emotional face tasks (Donegan et al., 2003). This signifies the important role of top-down connectivity when interpreting AMG activity. Follow-up research measuring psilocybin effects 1 day after intake suggests that AMG activity increases are accompanied by decreased connectivity between the prefrontal cortex and the AMG (Mertens et al., 2020). Although this research did not determine the direction of influence, the authors propose connectivity involving the AMG may be an important mechanism underlying previously reported therapeutic outcomes of emotional reconnection (Watts et al., 2017).
AMG and affect responses to negative faces have also been measured up to 1 month postpsilocybin in healthy participants. This research shows sustained reduction of AMG responses to negative faces lasting up to 1 month. Moreover, this research indicated that increased positive responses to emotionally conflicting (resistance arousing) stimuli, lasting up to 1 week, were associated with cortical projections to the dorsolateral prefrontal and medial orbitofrontal cortex (Barrett et al., 2020a). Positive responses to emotionally conflicting (resistance arousing) stimuli—involving the cortex and changes to the AMG—are evidence that psychedelics can have enduring effects on neuroplasticity, lasting well beyond the half-life of psilocybin. These results also speak to the importance of emotional regulation of the AMG by the cortex in long-lasting therapeutic outcomes. Reduced acute activation of the AMG by negative stimuli, under psychedelics, may contribute to reduced ego resistance and positively felt ego dissolution.
3. Parahippocampus and Hippocampus
The PHC is an associative hub of the MTL involved in memory retrieval, associative memory, contextual processing and detection of familiarity (Aminoff et al., 2013). The hippocampus (HC) is involved in decision making, learning, and memory consolidation and can be further divided into the dentate gyrus, the hippocampus proper, and the subiculum (Anand and Dhikav, 2012; Fogwe et al., 2021). The PHC is connected with the HC and may mediate HC connections with the associative cortex (Ward et al., 2014; Millière, 2017). This connectivity is relevant in limbic-cortical dysregulation impacting mood regulation (Zeng et al., 2012), and increased FC between the HC and prefrontal cortex (e.g., mPFC) is seen in depression (Kaiser et al., 2015). Decreased resting-state FC between the PHC and prefrontal cortex was identified under psilocybin and was predictive of treatment response in a study of psilocybin for treatment-resistant depression (Carhart-Harris et al., 2017). Moreover, these connectivity changes also predicted ego dissolution (Carhart-Harris et al., 2017). Dysconnectivity of the PHC is also implicated in medial temporal lobe epilepsy and déjà vu experiences (Illman et al., 2012), suggesting that, under psychedelics, PHC activity in the detection of familiarity may contribute to disruption of the narrative self (discussed later). Following this notion, information theoretical analysis of fMRI brain connectivity identified that reduced connectivity (as measured by lower diversity coefficient) between the anterior PHC and cortical regions increased the likelihood of experiencing ego dissolution under psilocybin (Lebedev et al., 2015). Speculatively, reduced connectivity may sensitize neuronal message-passing to the effects of 5-HT2AR activation.
Music also influences PHC connectivity with cortical regions under psychedelics and may relate to PHC function in associative memory and contextual processing. A study using DCM for fMRI data reported increased connectivity of the PHC to visual regions and attenuated top-down connectivity under LSD (Kaelen et al., 2016). This suggests that psychedelic-induced dissolution of hierarchical processing may decouple the PHC and the visual system (Kaelen et al., 2016). It also suggests free interactions lower in the cortical hierarchy, labeled anarchy (Carhart-Harris et al., 2014b). Moreover, hierarchical decoupling identified by reduced hippocampal glutamate levels under psilocybin was associated with positively felt ego dissolution (Mason et al., 2020). Although it remains uncertain whether reduced glutamate levels are a cause or consequence of ego dissolution, some authors propose that the decoupling of hippocampal connectivity may inhibit semantic autobiographical memory from reaching the cortex (Millière et al., 2018).
Oscillatory changes are also seen in psychedelic states. Cortical oscillations are rhythmic electrical activity or waves that are produced by interactions among neurons. Synchronous patterns of neuronal firing are commonly associated with states of arousal and sleep (Poulet and Crochet, 2019). EEG recordings under psilocybin show decreased PHC oscillations—also observed in coherence between the ACC and PCC (Kometer et al., 2015). These findings suggest that changes to cortical FC and the PHC contribute to the psychedelic experience. The PHC also contributes to spatial navigation functions of the RSC (Epstein, 2008; Vann et al., 2009).
4. Retrosplenial Cortex
The RSC supports a sense of head orientation and cognitive capacities such as visualization, autobiographical memory, future-oriented thinking, retrieval of memory, and navigation. The RSC’s role in navigation enables awareness of self-location, relative to spatial context, and the translation of spatial context to cognitive maps, which are independent of self-reference (Vann et al., 2009). The role of the RSC in generating reference-independent self-orientation has also been suggested to extend to representations of permanence, both of environmental features (Auger et al., 2012) and representations of permanence more generally (Auger and Maguire, 2018; Kim and Maguire, 2018). These representations may underwrite a sense of self, time, and space that are affected by ego dissolution. A study of LSD, integrating fMRI, MEG, and arterial spin labeling–MRI, showed decreased communication between the RSC and PHC, which correlated with subjective measures of ego dissolution and the maintenance of a sense of self (Carhart-Harris et al., 2016b). Desynchronization of δ-band oscillations, measured between the RSC, PHC, and lateral orbitofrontal areas, was also seen under psilocybin: this desynchronization was associated with spiritual experience and insight, which may be considered aspects of ego dissolution (Kometer et al., 2015). It was conjectured that the implicit effects on connectivity may facilitate insight-enhanced reprocessing of autobiographical memories (Kometer et al., 2015). Conversely, memory retrieval tasks under psilocybin do not support this supposition (Carhart-Harris et al., 2012a). However, it is worth noting that the memory retrieval studies included only a few subjects (n = 10) and did find lasting therapeutic effects of psilocybin (Carhart-Harris et al., 2012a). Despite the absence of clear evidence for the functional correlates of psychedelic effects on the RSC, its role in spatial orientation and representation of permanence suggests a role in ego dissolution.
5. Subcortical Summary
Evidence relating hierarchical connectivity changes between subcortical limbic regions and the cortex under psychedelics is consistent across studies and may relate to ego dissolution and therapeutic effects. However, further investigations are needed to identify the specific effects on directed (effective) connectivity and their functional role in sentience and ego dissolution. The effects on thalamic connectivity appear to be an increase in bottom-up connectivity, which may facilitate hallucinations/hallucinosis through reduced sensory filtration. The role of the thalamus in ego dissolution and the cognitive aspects of psychedelic experiences is uncertain and may be indirect. The connectivity of the AMG with cortical and visual regions speaks to an effect of psychedelics on emotional regulation. This connectivity also appears to be rendered increasingly plastic by psychedelics. In positively felt ego dissolution, connectivity between the AMG and cortical regions may facilitate reduced threat responses (defenses) and ameliorate ego resistance required for therapeutic outcomes. The effect of psychedelics on associative processes in the PHC may have a role in ego dissolution and mediate therapeutic effects through routing of neuronal signals between limbic structures and the cortex. Psychedelics may also decouple the PHC from regions lower in the hierarchy due to reduced top-down inhibition from higher levels of the cortex. The ensuing flexibility, or anarchy, may also account for increased context sensitivity experienced by subjects under psychedelics. A role of the RSC in the dissolution of ego permanence is conjectured. The RSC is related to sense of permanence and may mediate some of the therapeutic effects of psychedelics.
Changes in subcortical connectivity, with higher cortical areas, indicate the importance of hierarchical message-passing under psychedelics. Cortical regions support high-level processes, including self-related thinking, salience, and attention, and are responsible for top-down connectivity, which influences bottom-up projections from subcortical regions. The major cortical networks—that feature in the psychedelic imaging literature—are considered next.
P. Connectivity within Large-Scale Resting-State Networks
1. The Default Mode Network
The default mode network (DMN) is a brain network that shows increased endogenous fluctuations when an individual is awake but not engaged in a task requiring attention (Buckner et al., 2008). It was originally identified using PET and fMRI, which showed a decrease in activity from baseline—in certain brain regions—during goal-directed behavior. This decreases regional activity in the absence of attentional- and stimulus-bound processing (Raichle et al., 2001). Advances in our understanding of the role of the DMN rest on its connectivity to other parts of the association cortex (Hagmann et al., 2008; Bonnelle et al., 2012). DMN connectivity is believed to emerge over the course of human evolution and at a developmental timescale over the lifespan (Van Essen and Dierker, 2007; Supekar et al., 2010).
The free energy principle, and associated Bayesian brain hypotheses, rests on the notion that the brain is trying to fit or invert an internal model that generates the sensory consequences of hidden or latent states in the world. On this view, the brain embodies a hierarchical generative model of how sensations are generated. In terms of evolution, hierarchical brain organization is thought to reflect recently evolved high-level associative functions that have a predictive, supervisory and directive role over subordinate primal regions (Holmes and Nolte, 2019). The notion of hierarchy suggests an increased complexity of representations from sensory to associative regions of the brain (Felleman and Van Essen, 1991). Furthermore, complex sensory events involving longer sequences of representations are processed in superordinate levels of the hierarchy capable of generating ordinal sequences and narratives (Kiebel et al., 2008). In predictive coding formulations of the Bayesian brain, the highest levels of the hierarchy are thought to generate top-down predictions of sensory information. The DMN is believed to be situated at the top of the cortical hierarchy and may control bottom-up (prediction error) signals from lower levels of the hierarchy (e.g., the MTL) (Andrews-Hanna et al., 2010; Menon, 2011). The cycle of updating top-down predictions based on the flow of bottom-up prediction errors is referred to as hierarchical predictive coding (Rao and Ballard, 1999; Kanai et al., 2015; Friston, 2018).
2. Default Mode Network Function and Priors (Beliefs)
The function of the DMN centers on adaptive and internally oriented mental processes that allow humans to be aware of environmental threats, such as predators, in the absence of goal-directed attention (Raichle et al., 2001). The DMN’s role in directing attention inward—that manifests as self-oriented thoughts—also suggests its relationship to personal identity and the construct of narrative self, which is closely related to the Freudian concept of ego (Buckner et al., 2008; Carhart-Harris and Friston, 2010; Lebedev et al., 2015). In Freud’s theory, the ego regulates behavior by suppressing the free energy of underlying subordinate structures, such as the limbic and paralimbic systems (Cieri and Esposito, 2019). The DMN similarly attenuates free energy and regulates behavior by controlling the selection of information channels.
The DMN is rich in 5-HT2AR (Beliveau et al., 2017). Mounting evidence suggests that psychedelics decrease DMN connectivity; see Table 1, which provides a summary of connectivity changes under various psychoactive compounds (Ruban and Kolodziej, 2018). Acute decrease in DMN connectivity is observed under both psilocybin and LSD (Carhart-Harris et al., 2012b; Müller et al., 2018). Decreased DMN connectivity likely relates to ego dissolution and may involve the dissolution of self-related priors (beliefs) sustained by the DMN. These priors may support a narrative sense of self that disintegrates during ego dissolution. Self-related priors in the DMN are also thought to control the top-down inhibition of prediction errors from subordinate hierarchical levels (Dillon and Pizzagalli, 2007; Carhart-Harris and Friston, 2010; Carhart-Harris et al., 2014a). If top-down inhibitory mechanisms are disengaged by psychedelics, then DMN regions may become more receptive to bottom-up influences (e.g., from the MTL) that enable revision and updating of beliefs (Carhart-Harris and Friston, 2019).
DMN connectivity under classic psychedelics, inspired by Ruban and Kolodziej (2018)
Decreased DMN activity under psychedelics is a common finding across psychedelic literature. For DMN functional connectivity with other regions, see Ruban and Kolodziej (2018).
3. Default Mode Network and Priors in Mental Health
Maladaptive priors are surmised to be part of ego resistance. Disinhibition of ascending prediction errors by psychedelics may reduce ego resistance and play a permissive and foundational role in the updating of priors. Reported psychedelic therapeutic outcomes and increased communication between the subcortex and cortex is reflected in their role in the cognitive control of emotion (Ochsner and Gross, 2005). The interaction between cognition and emotion may help disarm maladaptive ego defenses. In the context of psychedelic therapy, supportive preparation and prior application of behavioral therapies may also disarm maladaptive ego defenses and help guide positively felt ego dissolution and facilitate cognitive-emotional engagement (Carhart-Harris et al., 2018a), emotional reconnection (Watts et al., 2017), and a sense of being attuned with one’s emotions (Roseman et al., 2018a).
The importance of DMN connectivity in mental well-being is evidenced in psychopathology research. For example, connectivity variability between the mPFC and PCC is greater in depressed individuals than in healthy controls (Wise et al., 2017), and increased connectivity of the DMN is associated with ruminative thought, low mood, and depression (Speth et al., 2016). Furthermore, reduced task-related mPFC suppression and increased functional connectivity of the DMN has been identified in schizophrenia (Whitfield-Gabrieli et al., 2009; Whitfield-Gabrieli and Ford, 2012), and decreased anterior and increased posterior DMN connectivity is reported in addiction (Zhang and Volkow, 2019).
These reports speak to the importance of DMN connectivity for psychologic well-being. For example, decreases in DMN functional connectivity is related to reduced references to the past by subjects post-LSD in task-based studies, suggesting that the influence of psychedelics on DMN function may relate to rumination (Speth et al., 2016). This could mean that psychedelics—similar to meditation—cultivate a more present-centered focus (Brewer et al., 2011). Moreover, this present-centered focus may represent reduced defense mechanisms.
Defense mechanisms associated with psychologic threats may have evolutionary origins in DMN monitoring for physical threats. To minimize surprise, the DMN remains aware of the environment and can mentalize temporally extended sequences of events (Andrews-Hanna et al., 2014). The DMN also responds to nonphysical forms of threat. The DMN displayed activity increases in subjects whose political beliefs have been challenged (Kaplan et al., 2016). These findings suggest that DMN mentalizing functions predict the consequences of threats to beliefs like physical threats. Defense mechanisms may have evolved to mitigate psychosocial hazards and minimize surprise. However, defense mechanisms also mark instability and suggest the difficulty of the ego to effectively estimate the value of immediate relative to temporally delayed action or gratification (i.e., regulate behavior) in a dynamic environment (Dayan and Daw, 2008; Dohmatob et al., 2020). At their extreme, defense mechanisms may form maladaptive beliefs that underwrite psychopathologies.
Investigation of the brain regions that form the DMN and their relationship with self-related processes may help discern mechanisms of action of psychedelics that mediate ego dissolution and therapeutic outcomes.
4. Posterior Cingulate Cortex
Within the DMN, a cardinal structure associated with the self is the PCC. The PCC absorbs around 20% more metabolic energy than most other brain regions (Raichle et al., 2001; Buckner et al., 2008). Its prominent role may be reflected in its structurally fortified location and access to ample blood supply (Raichle et al., 2001). The PCC remains active, automatically and continuously alert to surroundings (Raichle et al., 2001), and there is robust evidence of PCC involvement in processes of self-reflection (Vogt and Laureys, 2005) and a sense of self (Sampedro et al., 2017). This suggests that its activity underwrites a narrative sense of self (the ego) (Carhart-Harris and Friston, 2010; Carhart-Harris et al., 2012b). The PCC is also a hub that routes and regulates neuronal message-passing throughout the brain and has been shown to be abnormal in schizophrenic patients (Liang et al., 2020).
The importance of the PCC in the preservation of ego was suggested by an fMRI study of regional decreases of cerebral blood flow in the PCC (by up to 20%) that were correlated with ego dissolution induced by psilocybin (Carhart-Harris et al., 2012b). The relationship between the PCC and ego dissolution was further characterized by effective connectivity analysis of MEG data using DCM. DCM analysis demonstrated that psychedelics desynchronized cortical oscillatory rhythms of the PCC by decreasing α power (Muthukumaraswamy et al., 2013; Carhart-Harris et al., 2016b). This finding suggested that the α (8–13 Hz) frequency band of the PCC may be a correlate of ego integrity (Carhart-Harris et al., 2014)). This observation was reinforced by later findings, from fMRI under LSD, which also detected reduced α power in the PCC in association with ego dissolution (Carhart-Harris et al., 2016b).
PCC functionality is differentiated by its ventral and dorsal segments. The ventral PCC shares connections with the DMN during internally focused states and is thought to orient attention toward the self, whereas the dorsal PCC—a node within multiple resting-state networks—is involved in the dynamic coordination of attentional focus between internal and external thoughts by altering the metastability of resting-state networks (Leech and Sharp, 2014; Preller et al., 2019). The connectivity of the dorsal and ventral PCC with other brain regions—in conjunction with evidence of its alteration by psychedelics—suggests that it may be important to track these pathways to understand the mechanisms of psychedelic effects. Evidence of connectivity between the PCC and another cardinal node of the DMN, the mPFC, has also been highlighted in accounts of ego dissolution.
5. Connectivity to the Medial Prefrontal Cortex
The mPFC is implicated in executive cognitive functions, involving memory and decision making (Euston et al., 2012), and contextual associations between memory and responses by relating times, places, and events to adaptive emotional and physical responses (Euston et al., 2012). The mPFC integrates bottom-up internal and external information, conveyed by connectivity to the PCC, which may play a self-reflective role in decision making (Gusnard et al., 2001; Northoff et al., 2006; Brewer et al., 2011). Relatedly, functional connectivity between the PCC and mPFC is identified in postdecision dissonance (Tompson et al., 2016). The mPFC also has a role in selecting which bottom-up signals reach the DMN, which points to its significance in emotional processing (Gusnard et al., 2001; Ochsner and Gross, 2005). Under LSD, evidence of mPFC emotional mediation is demonstrated by reduced reactivity of right mPFC and left AMG functional connectivity in response to fearful faces (Mueller et al., 2017). Neuroimaging studies support the role of the mPFC in the suppression of limbic signals (e.g., in posttraumatic stress disorder) (Davidson et al., 2000; Hariri et al., 2000; Milad and Quirk, 2002; Phillips et al., 2003; Shin et al., 2004; Etkin et al., 2006; Hopper et al., 2007; Carhart-Harris and Friston, 2010). In short, changes in the mPFC facilitation of bottom-up signals, under psychedelics, may be important in ego dissolution and therapeutic outcomes.
Mindfulness research has identified reduced mPFC and PCC connectivity in meditators following training compared with control subjects (Brewer et al., 2011; Simon and Engström, 2015). Similarities between psychedelic experiences and meditation practice provides a reference for psychedelic-induced connectivity changes and therapeutic outcomes, and moreover suggest a potential synergy. Smigielski et al. (2019b) report a study in which experienced meditators practiced an open-awareness style of meditation under a single dose of psilocybin during a 5-day mindfulness retreat (Smigielski et al., 2019b). These meditators then underwent fMRI imaging 1 day after the retreat. Compared with controls, meditators exhibited reduced mPFC-PCC connectivity that correlated with ego dissolution. Moreover, lasting therapeutic outcomes were evident 4 months after administration. These results are significant because, despite being trained with an average 5000+ hours, the treatment group showed lasting psychologic gains beyond the benefits of extensive meditation alone (Smigielski et al., 2019a,b). Interestingly, during the follow-up fMRI, meditators in the psilocybin group were able to better modulate experiences of self-transcendence through their meditation associated with the mPFC-PCC decoupling (Smigielski et al., 2019b), suggesting psychedelic-induced neural plasticity in these projections. This study also noted reduced connectivity between the mPFC and angular gyrus, another node of the DMN, that was associated with a loosening of self-reference (Smigielski et al., 2019b). Increased self-transcendence has also been associated with cortical thinning of the PCC in regular ayahuasca users (Bouso et al., 2015). These research findings again highlight that psychedelics may facilitate lasting changes—in this case, self-transcendence through mPFC-PCC connectivity under meditation—that may also facilitate longer-term psychologic well-being.
An earlier fMRI psilocybin study found decreased functional connectivity between the PCC and mPFC. The link was interpreted as a reduced PCC influence on the mPFC and was related to ego dissolution (Carhart-Harris et al., 2012b). However, this assertion was not supported by an effective connectivity analysis of the directed influence. Another study using ayahuasca failed to find significant decoupling between PCC and mPFC using fMRI. Instead, and similar to the research of mindfulness meditators (Brewer et al., 2011), the PCC and mPFC showed reduced self-connectivity (Palhano-Fontes et al., 2015). The deactivation of mPFC has been related more generally to the subjective effects of psilocybin (Carhart-Harris et al., 2012b; Ruban and Kolodziej, 2018). In contrast, increased mPFC activation—which shares association with hyperfrontality—identified by higher levels of glutamate was associated with anxious ego dissolution (Mason et al., 2020). Decreased connectivity within or between the mPFC and PCC may indicate self-related attention, contextual judgement, emotional regulation, and suppression of bottom-up signals as possible mechanisms of ego dissolution. This evidence points to the importance of the midline regions of the DMN under psychedelics. However, further specification of the roles of these connections is required before any more definitive conclusions can be drawn.
6. Salience Network
The salience network (SN) serves important roles in sentience and conscious awareness. It detects and evaluates salient events (Menon and Uddin, 2010), monitors the environmental features relevant to goal-directed thinking (Seeley et al., 2007), and acts as the switching mechanism, coordinating attention between internal and external stimuli (Liang et al., 2015; Seeley et al., 2007). Moreover, the SN contributes to self-awareness (Menon, 2015). The SN is usually considered to be composed of the dorsal anterior cingulate cortex (dACC) and anterior insula (AI). Both are consistently coactivated across cognitive tasks (Swick et al., 2011), although the dACC is more involved in response selection and conflict monitoring (Menon, 2011; Ide et al., 2013). The AI receives more multimodal sensory input (Vogt and Pandya, 1987; Averbeck and Seo, 2008), detects behaviorally relevant stimuli (Menon, 2015), and coordinates the dynamic interactions of networks involved in external-oriented attention and internally self-oriented mental processes (Sridharan et al., 2008; Menon and Uddin, 2010). The SN is also associated with reasoning under uncertainty (Singer et al., 2009; Donoso et al., 2014; Lebedev et al., 2015), perception of time (Craig, 2009), and self-agency (Farrer and Frith, 2002). Many of these associations share a similarity to ego dissolution effects under psychedelics. Psychedelic-induced alterations of self-agency, the perception of time (Shebloski and Broadway, 2016), and reasoning under uncertainty indicate a possible role of the SN in the awareness of psychedelic experiences.
7. Anterior Cingulate Cortex
The ACC is a cortical midline hub region of the SN involved in cognitive control and is frequently highlighted in connection with psychedelic-induced altered states. It is highly connected to hub regions of the DMN and plays a role in self-referential processing (Northoff et al., 2006). It is also situated between emotional and cognitive domains, suggesting that it may bridge and mediate emotional responses (Liotti et al., 2000; Etkin et al., 2011; Stevens et al., 2011). Similar to the PCC, the ACC is a large region with dorsal and ventral subdivisions.
The dACC is connected to regions implicated in cognition, such as the (lateral) prefrontal cortex, parietal cortex, and premotor and supplementary motor areas (Asemi et al., 2015; Heilbronner and Hayden, 2016). dACC connectivity may be expressed in social pain. A task-based fMRI study exposed controls and participants under psilocybin to social exclusion cues. Under psilocybin, negative responses to social exclusion were reduced with decreased dACC connectivity (Preller et al., 2015). The dACC is a large cortical region with multiple functions, including performance monitoring (Ham et al., 2014). This may suggest that reduced dACC activity represents abnormal self-monitoring in response to social cues under psilocybin. Changes in AMG responsiveness under psychedelics suggests that changes to top-down mechanisms may indeed mediate emotional responses (Preller et al., 2020), with the ACC implicated (Ghashghaei et al., 2007; Fusar-Poli et al., 2010). Moreover, the rostral ACC (rACC) subarea of the ACC is related to emotional expression (Etkin et al., 2006) and—under psilocybin—has been related to subjective scores on emotional subcategories of the 5D-ASC (more on this below) (Lewis et al., 2020). Magnetic resonance spectroscopy, after ingestion of ayahuasca, showed increased connectivity between the ACC and MTL regions associated with enhanced self-compassion (Sampedro et al., 2017). Increased connectivity between the ACC and PCC was also detected. This study suggested that the ACC mediated the interaction between cognition and emotion that contributes to psychedelic afterglow effects and the enhanced mindfulness capacities measured in study participants. Mindfulness capacities cultivate a more present-centered default mode (Brewer et al., 2011), develop increased awareness of mental contents (Schooler et al., 2011; Dahl et al., 2015), and reduce maladaptive DMN activity associated with various forms of psychopathology (Soler et al., 2014; Sampedro et al., 2017; Ramirez Barrantes et al., 2019). Mindfulness functions may also help reduce ego resistance, and their increase following psychedelics again suggests evidence of neuroplasticity.
Despite its large size and differing functions, ACC subdivisions are not always reported in psychedelic literature (Stevens et al., 2011). This may contribute to conflicting findings. For example, past PET imaging evidence describes increased ACC and MTL activity under psilocybin (Vollenweider et al., 1997). However, contrasting evidence of reduced ACC neural oscillations in a network involving the PHC, PCC, and ACC, measured by EEG, have also been reported under psilocybin (Kometer et al., 2015), and further evidence suggests increased synchronization and entropy between the ACC and hippocampus under psilocybin (Tagliazucchi et al., 2014). The lower spatial resolution of PET and EEG may explain these contradictory findings. Spatial specificity of ACC changes under psychedelics may help identify associations between its connectivity, function, and phenomenology under psychedelics. For example, the ventral stream of ACC connects to affective brain regions, including the AMG (Etkin et al., 2011; Andrews-Hanna et al., 2014), that could be investigated in relation to therapeutic outcomes under psychedelics.
The structural size of the ACC has also been related to psychedelics. The subjective effects of a standardized psychedelic dose can vary widely across participants, and this variance may be related to the structural volume of the rACC, previously noted in subjective scores on emotional subcategories of the 5D-ASC. rACC volume correlated with interindividual subjective effects, suggesting that its structural variation across volunteers may predict feelings of unity, bliss, spiritual experience, and insightfulness (Lewis et al., 2020). Structural increases of the ACC were noted in long-term ayahuasca users and may be related to long-term psychedelic use that is thought to preserve psychedelic-induced neuropsychological changes (Bouso et al., 2015). This sample also showed reduced cortical thickness in the PCC. Notably, these long-term users of ayahuasca displayed high levels of mindfulness, which may suggest a relationship between structural change of the SN and DMN following psychedelics that is related to the increased levels of mindfulness. These structural differences may have existed prior to exposure to ayahuasca and disposed participants toward mindfulness and psychedelic use. This highlights the potential bias in recruitment samples in psychedelic studies.
The ACC and PCC structural differences do, however, align with evidence of the reliance of mindfulness capacity on SN control over the DMN. Although the DMN is responsible for mind wandering, awareness of mind wandering can be regulated by the SN. This is reflected in functional connectivity increases during awareness of mind-wandering in meditation examined in fMRI (Hasenkamp et al., 2012). SN hierarchical dominance over the DMN has been validated using spectral DCM analysis (Friston et al., 2014; Zhou et al., 2018), and its control over the DMN is cited as a crucial component in cognitive health and performance (Putcha et al., 2016). Its breakdown results in dysregulated DMN function (Bonnelle et al., 2012). SN role in performance monitoring and emotional mediation may account for the influence of psychedelics on self-awareness and affective regulation and encourage a more mindful or present-centered focus.
8. Frontoparietal Control Network
The frontoparietal control network (FPCN) is an attention control network, also known as the central executive network or the frontoparietal central executive network (Menon, 2011). It encompasses the dorsolateral and anterior prefrontal cortices, inferior parietal lobes, AI, and ACC (Vincent et al., 2008; Rens et al., 2017). Notably, the latter two regions (AI and ACC) overlap with the SN. The lateral prefrontal cortex and inferior parietal lobule are associated with cognitive control and decision making (Lord et al., 2018). The FPCN is also associated with access to conscious information (Dehaene and Naccache, 2001) and is inhibited in states of deep sleep (Boly et al., 2008; Tagliazucchi et al., 2016). The functional role of the FPCN can be summarized as evaluations over time, involving uncertainty or abstraction in voluntary choices (Andrews-Hanna et al., 2014; Fan, 2014; Fan et al., 2014; Rens et al., 2017) and includes the dorsal prefrontal and posterior parietal cortices as key regions tasked with negotiating acute uncertainty (Huettel et al., 2005). Abnormalities in this network, following brain injury, have been associated with impaired self-awareness (Ham et al., 2014), which may indicate its relevance to ego dissolution.
Decreases of connectivity within the FPCN are detected under both LSD and psilocybin (Lord et al., 2018, 2019; Barnett et al., 2019). However, the decrease of within-network connectivity is accompanied by increases in between-network connectivity (i.e., deviation of functional connections from network pathways). This pattern of reduced-within and increased-between connectivity—coined as disintegration and desegregation (Carhart-Harris et al., 2016b)—is documented across resting-state networks under psychedelics (Barnett et al., 2019; Varley et al., 2019) and alters the functional integration of brain regions (Petri et al., 2014) (Fig. 5). Cognitive function depends on the dynamic, context-sensitive regulation of functional segregation and integration in the brain and is mediated by neural gain modulation. Neural gain by projections from ascending neuromodulatory nuclei has the computational capacity to elicit global fluctuations, altering the balance between segregation and integration in the brain (Shine et al., 2018; Shine, 2019). Disintegration and desegregation highlight the role of the sensitization (i.e., the neuromodulation) of high-level serotonergic populations under psychedelics underwriting changes to cognition. FPCN desegregated connectivity is highlighted by several studies (Tagliazucchi et al., 2016; Lord et al., 2018, 2019) and speaks to the importance of connectivity in the FPCN in psychedelic experiences.
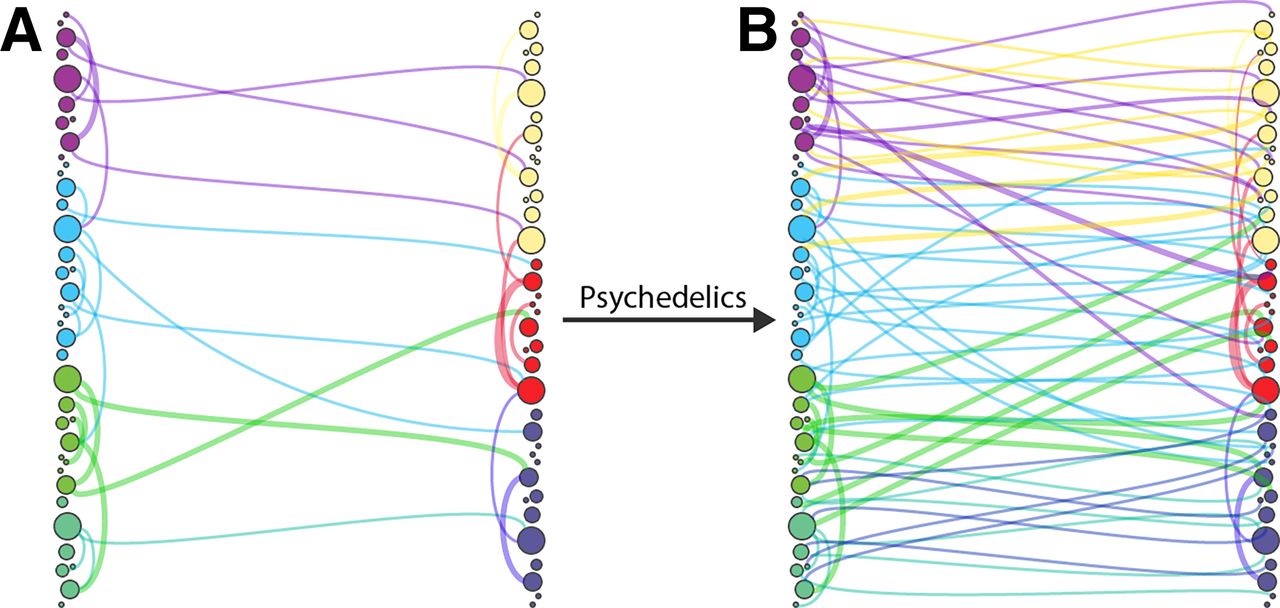
Illustration of desegregated connectivity under psilocybin, inspired by Petri et al. (2014). (A) Integration between communities—organized by color—observed in healthy adults. (B) Greater integration and reduced constraint of connections between communities observed under psilocybin. For original schematic and methods, see Petri et al. (2014).
“Connectome harmonics” research has revealed that patterns of psychedelic brain dynamics preserve new, complex forms of coherence and stability (Atasoy et al., 2017; Lord et al., 2019; Varley et al., 2019). Increased power and energy of connectome harmonic brain states, observed under psilocybin, display higher spatial and temporal variability yet maintain novel forms of stability and have been related to an increased repertoire of brain states (Atasoy et al., 2017). Links between these changes and their relationship to disintegrated and desegregated connectivity across resting-state networks are not fully understood; however, theories of their role upon the FPCN can be surmised. For example, evidence of FPCN desegregated connectivity (Tagliazucchi et al., 2016; Lord et al., 2018, 2019) may influence consciousness by enabling access to connectivity usually restricted from the FPCN. Desegregation may also be mechanistic to increased repertoires of brain states made available under psychedelics (Tagliazucchi et al., 2014; Atasoy et al., 2017). Both observations might also relate to the phenomenological richness of psychedelic experience and therapeutic outcomes thought to depend on increased access to (precise) bottom-up prediction errors necessary for the revision of priors. The coinciding disintegration of the FPCN (Lord et al., 2018, 2019; Barnett et al., 2019) and relaxation of priors (Carhart-Harris and Friston, 2019) would also be mechanistic in this process; however, they may contribute to cognitive aspects of ego dissolution. Untangling the mechanistic relationship between FPCN changes under psychedelics and their role in ego dissolution needs more research; however, existing evidence points to the potential importance of its role in cognitive aspects of the psychedelic experience.
Correlating cortical oscillatory changes with psychedelic experiences and therapeutic introspection may also help elucidate the role of the FPCN under psychedelics. Under psilocybin, an MEG study reported decreased (bilateral) β-band activity in the FPCN (Muthukumaraswamy et al., 2013). β-Band connectivity is suggested to encode long-term priors of behavior and environmental stimuli (Betti, et al., 2021). Decreased oscillatory power of the FPCN may be a measure related to change and revision of the association between behavior and stimuli. θ-band connectivity is proposed to support internally directed attention between the DMN and FPCN (Kam et al., 2019) and thus important in psychedelic research regarding brain oscillations. However, FPCN connectivity changes are not always reported under psychedelics (Müller et al., 2018), and the source of inconsistencies—which are discussed later in detail—remain uncertain, although are not entirely unusual in imaging research. Added caution is further suggested by the variable functional topography of the FPCN between individuals (Marek and Dosenbach, 2018). The importance of psychedelic effects upon the self can be explored through large-scale integrated dynamics.
Q. Connectivity between Large-Scale Resting-State Networks
1. The Self in Networks
The SN is related to the minimal aspects of conscious self-awareness (Limanowski and Blankenburg, 2013), also known as the embodied aspect of self (Seth, 2013; Lebedev et al., 2015). The minimal or embodied aspect of self-awareness requires bottom-up processing to define physical boundaries, proprioception, interoception, and the experience of oneself being rooted in (sensory) motor processes (Blanke and Metzinger, 2009; Legrand and Ruby, 2009). This aspect of self may be important for ego dissolution, indicated by decreased within-SN connectivity associated with ego dissolution in an fMRI psilocybin study (Lebedev et al., 2015). The DMN, in contrast, is related to the narrative sense of self commensurate with personal identity, described in Bayesian terms as the brain predicting its own contribution to sensory input by top-down inferences about the causes (i.e., latent or hidden states of the world) of sensory information (Metzinger, 2003; Friston, 2010; Clark, 2013).
Aberrant DMN-SN connectivity is important in clinical and theoretical models of various mood disorders, including depression, anxiety, and schizophrenia (Menon, 2011). Mood disorders associated with this connectivity suggest its importance in therapeutic outcomes, and change to this connectivity under LSD is associated with positive mood and arousal (Atasoy et al., 2017). However, DMN-SN functional connectivity is more commonly linked to ego dissolution and shows reduced anticorrelation under psilocybin (Carhart-Harris et al., 2013) and decreased segregation under LSD (Carhart-Harris et al., 2016b). The relationship of DMN-SN connectivity to respective aspects of self suggests that psychedelic influence on these networks may contribute to ego dissolution. Furthermore, evidence of their relationship to psychopathology speaks to the overlap between networks involved in the self and mental well-being. Further analysis of this connectivity and psychologic changes under psychedelics may help determine the contribution of this network to ego dissolution and normal waking consciousness. These possibilities invite further investigation of the well established anticorrelation between these networks.
2. Coordinated Balance between Anticorrelated Networks
An important feature of intrinsic brain networks is their coordinated and balanced activity. Patterns of correlations and anticorrelations between networks constitute an aspect of functional integration supporting consciousness (Bak et al., 1987; Demertzi et al., 2019). For example, the DMN focuses attention inward in a task-free exploratory manner that—upon stimulation from the environment—rapidly deactivates in deference to externally focused networks like the DAN. The dorsal attention network (DAN) is composed of the frontal eye fields and inferior parietal sulcus and is responsible for orientation to behaviorally salient cues (Ptak and Schnider, 2010). Altered DAN connectivity has been reported in the psychedelic literature and has been related to an increased repertoire of brain states under psilocybin (Tagliazucchi et al., 2014).
The SN mediates the anticorrelation of the DMN and DAN (Fox et al., 2005) through dynamic mechanisms responsible for the coordination of attention between bottom-up perceptual features of the environment and top-down goals (Menon, 2015). The SN is hierarchically above the DAN and DMN, suggesting that it may be a control network capable of selecting between these anticorrelated networks (Zhou et al., 2018). SN-meditated switches enable attentional resources to transition efficiently from internal focus of the DMN to an external focus (i.e., sensory attention) of DAN. Psilocybin reduces the anticorrelation between DMN-DAN connectivity deployment (Carhart-Harris et al., 2013). Reduced anticorrelation indicates that attentional resources may not be divided between external and internal realms of attention in the usual manner. Descriptions of ego dissolution that describe the dissolution of boundaries between the objective and subjective world signify the importance of investigations of the DMN-DAN anticorrelation (Stoliker et al., 2021). Like SN-DMN connectivity, competition between the DMN and DAN is correlated with disorders such as schizophrenia (Whitfield-Gabrieli et al., 2009), highlighting the potential importance of DMN-DAN anticorrelation in mental health and perceptual synthesis. A certain form of meditation, known as nondual awareness, tries to reduce the separation between external and internal reality, which, intriguingly, reduces the anticorrelation between brain areas that respond to intrinsic and extrinsic stimuli (Josipovic et al., 2012). This finding reinforces the suggestion that DMN-DAN anticorrelation may be crucial to the loss of boundaries between the subjective and the objective world (Grof, 1980) that characterize ego dissolution.
Alterations to anticorrelated brain networks may also be involved in therapeutic outcomes following psychedelics; however, evidence is scarce. Whether therapeutic outcomes represent a direct result of altered anticorrelation or changes to connectivity between other systems such as the MTL under acute effects is unclear. It may be possible that the altered anticorrelations contribute to ego dissolution and promote neuroplasticity, leading to psychologic changes. Furthermore, the role of these networks in higher-order processing and attentional orientation could indicate access to information that is usually suppressed and may contribute to therapeutic processes that reorganize patterns of thought. Evidence of DMN-DAN connectivity changes associated with enhanced mindfulness capacities may represent a link to reduced reactivity, judgmental thinking and enhanced self-kindness (Sampedro et al., 2017). Moreover, anticorrelations under control of the SN also involve the FPCN (Zhou et al., 2018), which is known to be anticorrelated with the DMN (Menon, 2011, 2018; Andrews-Hanna et al., 2014; Chand et al., 2017; Bolton et al., 2020).
Control of the FPCN is managed by the SN recruitment of the AI, previously noted to be involved in the interactions between internal and external mental processes (Sridharan et al., 2008; Menon and Uddin, 2010) and detect behaviorally relevant stimuli (Menon, 2015). Specifically, activation of the right AI engages the FPCN, resulting in coactivation of the SN and FPCN (Raichle et al., 2001; Greicius and Menon, 2004; Menon, 2015) [Recent evidence suggests that nodal measurements are more sensitive to the severity of disorders of consciousness than global measurements (Martínez et al., 2020)]. Although normally anticorrelated, correlated activation of the FPCN and DMN has been documented to correlate with symptoms of psychosis that relate to a disturbed sense of self and uncertainty of what is and is not real (Leptourgos et al., 2020). Changes to the functional integration of the FPCN, DMN, and SN are associated with meditation practice (Doll et al., 2015) and the control of attention (Corbetta and Shulman, 2002). These observations speak to the putative role of anticorrelations in ego dissolution that involve networks that orient attention and contribute to self and perspectival belief updating.
Subcortical brain regions that evince anticorrelation under psychedelics have also been investigated. For example, the claustrum is a subcortical region that has strong connectivity with the prefrontal cortex (Brown et al., 2017). It has been shown to activate with demanding cognitive tasks and the engagement of the FPCN (Krimmel et al., 2019). A recent study showed that under psilocybin, connectivity between the right claustrum and the DMN decreased, whereas connectivity to the FPCN increased (Barrett et al., 2020b). Claustrum connectivity changes may contribute to functional changes in cortical networks that support attention and self-consciousness. Further investigations comparing anticorrelation of brain networks in psychosis, meditation, and classic psychedelics may help determine the functional importance of brain anticorrelations in ego dissolution, therapeutic outcomes, and the consciousness of self.
R. Methodological Considerations
1. Conflicts within Psychedelics Findings
The association between subjective effects and brain connectivity is often inconsistent among psychedelic studies. A brief overview of these discrepancies characterizes the present state of understanding in psychedelic neuroscience. Ego dissolution is often associated with DMN hub regions under psychedelics. This is demonstrated by mPFC-PCC decoupling in fMRI (Smigielski et al., 2019b) and decreased PCC α power in MEG under psilocybin (Muthukumaraswamy et al., 2013). More generally, subjective effects under psilocybin are associated with decreases across the ACC/mPFC (Carhart-Harris et al., 2012b). However, brain-wide connectivity changes are also associated with ego dissolution and extend beyond the DMN under LSD (Tagliazucchi et al., 2016) and have been linked to connectivity between the anterior PHC and cortical communities, including visual, somatosensory, salience, prefrontal, and control areas under psilocybin (Lebedev et al., 2015). Furthermore, not all psychedelic investigations find changes to the DMN. For example, the expected changes in complexity of brain signals—measured with the fractal dimension—from DMN nodes in fMRI, under LSD and psilocybin, were not found (Varley et al., 2019).
An LSD study employing fMRI, MEG, and arterial spin labeling identified decreased RSC and PHC connectivity that correlated with ego dissolution and noted that activity of resting-state networks such as the DMN was not correlated with ego dissolution (Carhart-Harris et al., 2016b). These findings challenge the reliability of DMN changes associated with ego dissolution. An EEG study also found cortical oscillations in the RSC, PHC, and lateral orbitofrontal areas; however, in this study, these changes were associated with spiritual experience and insightfulness and not with ego dissolution (Kometer et al., 2015). Other resting-state networks suspected to have a role in psychedelic effects such as the FPCN are also sometimes reported as absent: in this case, in fMRI scans under LSD (Müller et al., 2018). Furthermore, effective connectivity involving the PCC was found to correlate with subjective effects of psilocybin in a DCM analysis of MEG (Muthukumaraswamy et al., 2013), yet evidence of its alteration in an fMRI study of LSD was absent (Müller et al., 2018). See Table 2 for a summary of connectivity results associated with ego dissolution (Millière et al., 2018).
2. Conflicts between Psychedelic Findings and Nonpsychedelic Findings
Occasionally, psychedelic-induced changes in connectivity—cited in association with subjective effects—are seen in nonpsychedelic studies without subjective effects. For example, decreased activation within the DMN is cited in association to ego dissolution (Carhart-Harris et al., 2016b). However, this also resembles the patterns of activity following administration of sertraline, a nonpsychoactive drug (Klaassens et al., 2015; Müller et al., 2018). Within the DMN, the deactivation and decoupling of the PCC and mPFC has been suggested to underlie ego dissolution (Carhart-Harris et al., 2012b; Smigielski et al., 2019b; Carhart-Harris et al., 2014b). Although, this deactivation is also identified in response to antidepressants, with no noticeable psychoactive effects (Goldstein-Piekarski et al., 2018), and a similar activation pattern is also elicited by anxiety (Zhao et al., 2007). If DMN connectivity changes underlie alterations to perception reported under psychedelics, then we might expect similar connectivity changes to produce psychedelic-like subjective effects. Psychedelic experimental designs are at risk of inducing anxiety, especially when rapid onset of a psychedelic through intravenous or intranasal route is employed. It is, therefore, important to note known confounds when trying to identify neural mechanisms.
3. Signal from the Noise
The results of neuroimaging studies suggest connectivity associations with ego dissolution—and other subjective effects of psychedelics—that have yet to be clearly determined. Examining and integrating cellular, regional, network, and whole-brain connectivity changes may best account for observed phenomenological changes in consciousness (Varela et al., 2001; Palva et al., 2005; Melloni et al., 2007; Busch et al., 2009; Dehaene and Changeux, 2011; Hipp et al., 2011; Fahrenfort et al., 2012). Psychedelic subjective effects will likely involve widespread changes to hierarchical connectivity subtending associative and sensory processing across the cortex and subcortex. Holistic accounts of brain changes under psychedelics—that identify reliable correlations between connectivity and subjective experience—may be followed by more thorough and advanced examinations, such as those using effective connectivity analyses. However, the accurate modeling of psychedelic brain connectivity is encumbered by challenges.
The anatomic coordinates of regions—such as the PCC—may be specified using various methods for subsequent region of interest–based analysis. For example, independent component analysis can be used to detect the peak activity of cardinal regions and their corresponding coordinates, or these coordinates may be borrowed from previous literature. Recent alternate methods have also been proposed, involving probabilistic mapping to improve consensus and inferences (Dworetsky et al., 2021). These strategies may produce different connectivity results when referring to the same regions. Variability due to individual differences in brain anatomy may further complicate accurate determination of anatomic coordinates. Commonly-referred-to networks may also be composed of different regions. Moreover, the application of global signal regression has been considered a cause of anticorrelated brain connectivity, and debate over its application questions interpretations of anticorrelations (Murphy et al., 2009). Concerns also exist with the interpretation of the blood oxygen level–dependent signal and whether the blood oxygen level–dependent signal is sensitive to changes in conscious experience (Laumann et al., 2017).
These concerns aside, understanding the mechanistic unfolding of processes after psychedelics can be advanced by imaging and analytic approaches. This requires increased sample size and the standardization of image processing pipelines, including the methods of drug administration, timing of imaging, environmental setting, balance of participant group–averaged neuroanatomy, and data preprocessing pipelines to clean (physiologic) signals. Variation in preprocessing pipelines in particular can be a significant barrier to the accurate comparison of findings across neuroimaging studies. For comparison of motion correction pipelines, see Parkes et al. (2018); for global signal regression see Almgren et al. (2020) and Aquino et al. (2020); and for motion correction in diffusion imaging pipelines, see Oldham et al. (2020). The optimization and standardization of these pipelines—in conjunction with replication and comparison between psychedelics—may be essential. Fortunately, growing interest in psychedelic therapeutic potential may motivate standardization and replication.
Lastly, we consider the measurement of ego dissolution in imaging studies. Ego dissolution is a phenomenon that varies across subjects provided a standard dose of a classic psychedelic. Therefore, the available connectivity findings do not precisely indicate the changes underlying psychedelic ego dissolution. Retrospective self-report measures also limit the measurement of ego dissolution. Self-reporting, typically taken after the effects have subsided, is inherently subjective and an issue of reliability. Subjects may have no prior basis for the experience of ego dissolution. This suggests variability in the self-scoring of ego dissolution across subjects who may have experienced similar subjective effects. An additional measurement consideration is that the onset of subjective effects can vary across subjects. This means that the time at which imaging occurs may not align with psychedelic peak effects. To offset these issues, we recommend subjects complete qualitative behavior reports detailing their experience with special attention to the imaging portion of the session. Researchers may also record observations as an additional means of assessing the degree of ego dissolution and the time of its occurrence. Standardized timing of imaging procedures may also be reconsidered by initiating procedural “time zero” at the point that subjective effects first become noticeable. This may perhaps be achieved by relying on subject reports, for example, of closed eye visual alterations. Development of such onset measures may help ensure imaging reliably records optimal subjective effects. Furthermore, researchers may wish to discard data from outlier subjects who experience very mild subjective effects from the research dose of a psychedelic to improve analyses and consider building a registry of subjects with high subjective effects for future imaging studies.
4. Unifying Neuroimaging Evidence of Psychedelic Mechanisms
Closing gaps in the understanding of the neural mechanisms of subjective effects of psychedelics and formulating a unifying theory of 5-HT2R activity across the brain remains a goal of psychedelic research. Psychedelics have been suggested to drive the brain away from order toward a state of disorder in the entropic brain theory (Carhart-Harris et al., 2014). The flexibility of conscious states, spatial and temporal changes in connectivity, and increased sensitivity to intrinsic and extrinsic perturbations under psychedelics supports this theory (Carhart-Harris et al., 2014a; Carhart-Harris, 2018; Lord et al., 2018, 2019). Observed criticality characteristics have been used to understand the stable and coherent organization of brain dynamics under psychedelics (Atasoy et al., 2017). Criticality is a widespread phenomenon in multiscale and complex systems such as the brain. Criticality arises between states of order and disorder. It is marked by scale-free fluctuations that stretch from the finest to the coarsest scale and that can spontaneously jump between diverse spatiotemporal patterns (Cocchi et al., 2017). Although criticality has also been challenged as an unnecessarily complex explanation of drug-induced states (Muthukumaraswamy and Liley, 2018), it illustrates that psychedelics may temporarily exercise neuronal and behavioral flexibility.
An empirical approach to measuring psychedelic mechanisms across the brain comes from a recent imaging study explaining broad FC patterns leading to subjective effects. This study measured time-dependent changes of psilocybin over the course of onset to peak effects at three timepoints (20, 40, and 70 minutes post–oral administration) (Preller et al., 2020). Resting-state FC was quantified using a data-driven global brain connectivity method, which suggested that psilocybin reduces the connectivity of associative regions and increases connectivity in sensory regions across subcortical and cortical areas (Preller et al., 2020; Vollenweider and Preller, 2020). The authors suggest that the disintegration of associative activity and synchronization of sensory activity may explain subjective sensory and self-related experiences under psychedelics.
Amounting basic and clinical evidence also indicates that subcortical alteration of thalamo-cortical gating alters the bottom-up flow of information (Vollenweider and Geyer, 2001; Geyer and Vollenweider, 2008). Thalamic gating is controlled by glutamatergic cortico-striatal and cortico-thalamic pathways and is modulated by serotonergic and dopaminergic projections (Vollenweider and Smallridge, 2022). Vollenweider and Smallridge (2022) and work from their laboratory have established a working model of thalamic cortical circuitry identifying psychedelic stimulation of 5-HT2A receptors on CTSC cortical pyramidal cells and GABA interneurons on thalamocortical information flow. For illustrations of their working hypothesis of CTSC and cortico-cortical circuits, see Vollenweider and Smallridge (2022). The result is altered cortico-cortical integration of the distributed neuronal activity underlying the integration of external and internal information reaching the cortex. Thalamic changes are proposed to underwrite psychedelic perceptual alterations and suggest a neural mechanism of disintegrating subject-object boundary (Vollenweider and Smallridge, 2022). The thalamic gating account suggests that psychedelics increase bottom-up prediction errors, which may share mechanisms with the disinhibition of top-down associative connectivity and related desegregated patterns of connectivity. These theoretical and empirical accounts offer useful summaries of psychedelic subjective effects that complement an evaluation of psychedelics offered by the framework of hierarchical predictive coding.
5. Hierarchical Predictive Coding
Hierarchical predictive coding is a mechanistic account of how the brain processes information (Friston, 2008). It is supported by empirical evidence (Mumford, 1992; Rao and Ballard, 1999) and informed by our understanding of synaptic communication (Adams et al., 2013; Friston, 2020; Hobson et al., 2021). Noting the neuromodulatory role of serotonergic neurotransmission (Picard and Friston, 2014) and the influence of psychedelics upon these receptors, it provides a unifying account of psychedelic subjective and therapeutic effects. The premise of hierarchical predictive coding is that the brain is a constructive organ, explaining (i.e., inferring) sensory impressions or the causes of input on sensory epithelia. Sensory signals from the outer world are transduced into electrical signals that ascend the brain’s hierarchical architecture — in the form of prediction errors—to update (Bayesian) beliefs about hidden states in the world. These (subpersonal) beliefs form the basis of perception and can be regarded as the brain making predictions about the causes of its sensations. As new impressions from the sensory epithelia ascend the hierarchy, the accuracy of prediction is tested against the new sensory evidence in a cyclical process described variously as predictive coding, Bayesian filtering, or Bayesian belief updating. In short, bottom-up sensory evidence carries information (i.e., prediction errors) that updates expectations about states of affairs (i.e., posterior perceptions based on prior beliefs). These expectations then provide revised predictions that descend through the hierarchy to resolve prediction errors at lower levels.
The computational formulation of hierarchical predictive coding goes on to consider neuromodulation in the construction of predictions, suggesting that gain control (i.e., sensitivity) is a key player in selecting the most reliable or precise prediction errors for belief updating at higher hierarchical levels. Synaptic gain is thought to encode precision, and synaptic gain control reflects predictions of precision that implement this (attentional) selection. Computationally, precision is the predictability of some random variable and may be interpreted as a level of confidence placed in a prediction or prediction error. At high levels of the hierarchy that may entail (declarative or experienced) conscious processing, precision refers to confidence in an inference or belief. The neuromodulation or sensitization of neural populations is thought to determine the attenuation or augmentation of precision (i.e., confidence) in a source of information. Precision control, therefore, provides a synaptic mechanism describing the selection and attenuation of sensory channels and is proposed as a neurologic explanation for many psychopathologies (Brown et al., 2013). For example, in psychosis, abnormal neuromodulation or sensitization of neural populations may confound selection and attenuation of sensory channels, resulting in false inferences (Adams et al., 2013; Kanai et al., 2015).
The role of psychedelics in neuromodulation derives from binding to serotonergic 5-HT2A receptors. Serotonergic receptors serve a crucial role in precision control underlying sentience and attention that is necessary to infer both the cause of sensory impressions and the consequences of self-initiated actions (Brown et al., 2013; Parr and Friston, 2017, 2019). Please see (Parr and Friston, 2017, 2018; Parr et al., 2018) for recent theoretical treatments of neuromodulation and precision under active inference. Imaging evidence and brain connectivity findings are consistent with the preferential stimulation of 5-HT2AR on deep pyramidal cells, which are densely distributed in visual and associative areas of the cortical hierarchy (Carhart-Harris et al., 2012b; Saulin et al., 2012; Roseman et al., 2016; Carhart-Harris et al., 2016b; Preller et al., 2018b; Carhart-Harris and Friston, 2019; Preller et al., 2020; Vollenweider and Preller, 2020). Under predictive coding, these cells are thought to be the cells-of-origin of descending predictions to subcortical systems or lower hierarchical levels.
Consistent connectivity findings across visual and associative areas have helped account for psychedelic hallucinogenic effects and ego dissolution, respectively, and can be framed within the hierarchical predictive coding framework. For example, subcortical connectivity to sensory and associative areas is dependent on synaptic gain control. Furthermore, links between pharmacology and psychology can be considered in terms of coordinated message-passing of belief-updating cortical hierarchies. Expression of 5-HT2AR agonism serves a role in the active coping with stress and uncertainty (Carhart-Harris and Nutt, 2017; Murnane, 2019). Increased entropy and changes to synaptic gain control at the top of the hierarchy may influence the executive functions of cognition and priors. In short, hierarchical predictive coding provides a biologic starting point to understand the effect of pharmacological interventions at the synaptic level in terms of sentience and planning.
The leading synthesis of hierarchical predictive coding and psychedelic effects to date is the relaxation of beliefs under psychedelics (REBUS) model. REBUS describes the relaxation of precision at high levels of the hierarchy, leading to a greater influence of ascending prediction errors. In other words, a rebalancing of prior and sensory precision that enables new prior hypotheses and narratives to be engaged. In virtue of these expectations being at the apex of a hierarchical generative model, they are necessarily concerned with agency and active sentience of a multimodal sort and, on one reading, entail an implicit or explicit notion of self as agent.
Physiologically, the relaxation of high-level precision corresponds to a reduction in the synaptic gain or efficacy of particular regions in the cortical hierarchy. These changes are relatively consistent with neuroimaging findings, which show decreased within-network connectivity and altered oscillatory rhythms. The REBUS model may explain both the mechanisms of psychedelic subjective (Stoliker et al., 2022) and therapeutic effects; however, it remains to be validated (Fig. 6). Understanding that synaptic efficacy depends upon neuromodulation and that neuromodulation (of synaptic gain) changes the inhibition-excitation balance of synaptic integration suggests how changes in serotonergic agonist activity can influence internalizing disorders—disorders that are characterized by rigid ways of thinking and unduly precise prior beliefs. This is particularly relevant to psychiatry by suggesting psychedelics as a neuromodulator that affects the selection of ascending information—and thereby may be capable of subverting overly precise beliefs (i.e., ego resistance).
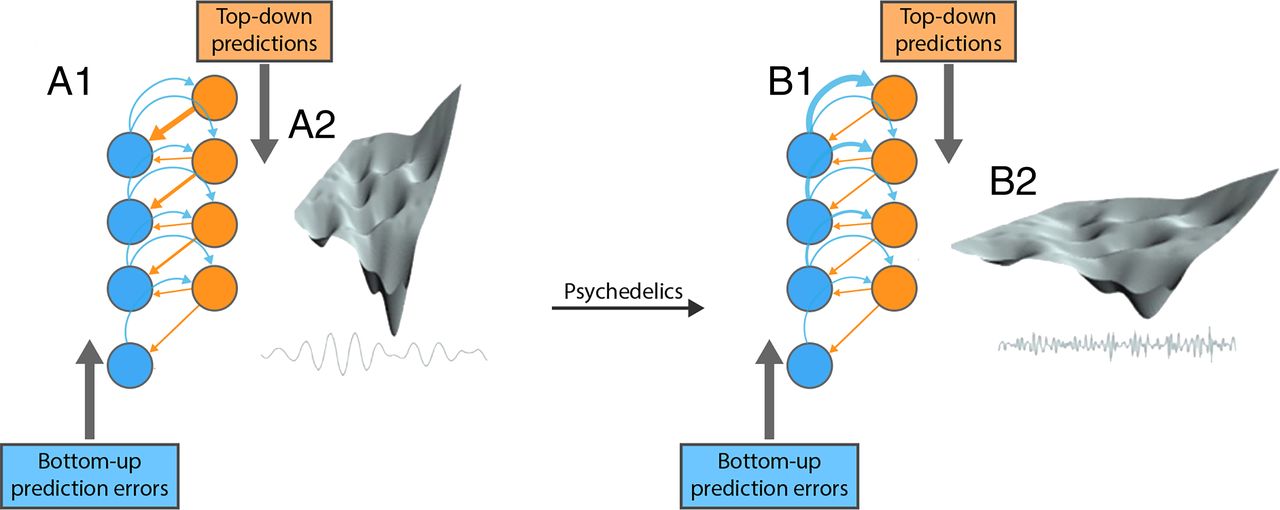
(A1) Sensory input is compared with top-down predictions to form prediction errors that are passed onto higher levels of the hierarchy to revise Bayesian beliefs. These beliefs or representations then supply top-down predictions, which resolve the prediction errors at the lower level. This process is repeated to minimize the prediction error at each level. The predictive coding hierarchy tries to construct the best top-down explanation for bottom-up sensory input at each level of the hierarchy. (B1) Psychedelics are thought to reduce precision and flatten the energy landscape of beliefs generated in high levels of the hierarchy supporting self-related beliefs, thereby producing the dissolving of self-related priors (i.e., ego dissolution). (A2 and B2) Dissolution of precision of high-level priors flatten the curvature of the free energy landscape, enabling neural dynamics to escape their local minima or basins of attraction, allowing greater attention to the sensory input and prediction errors (computationally expressed as a free energy landscape). The cognitive-therapeutic result of ego dissolution is the reduced precision or commitment to higher-level beliefs in the high levels of the hierarchy that affords an opportunity to explore a landscape of alternative hypotheses of the causes of sensory impressions and the consequences of self-initiated actions. Change to these explanations can be therapeutic by enabling new ways to make sense of the world and lived exchanges with it. This notion of free energy landscapes is endorsed by empirical studies of electrical physiologic responses and functional anatomy (Bastos et al., 2012). Adapted from Carhart-Harris and Friston (2019).
Clearly, we have somewhat simplified the account of how psychedelics mediates its effects through changing precision or postsynaptic gain. The effects of psychedelics are complex, and the precise pathways and synaptic mechanisms remain unclear. For a detailed account of the specific signaling pathways engaged by psychedelics, see Marek (2018) and López-Giménez and González-Maeso (2018). The particular 5-HT2AR–expressing populations also need to be considered. For example, 5-HT2AR–expressing deep-layer pyramidal neurons appear to be those that are most likely to be directly activated by psychedelics (Martin and Nichols, 2016). Psychedelics affect many types of neurons and produce complex transcriptional responses within the brain (Martin and Nichols, 2016). Generally speaking, activation of 5-HT2ARs induces an increase in excitatory postsynaptic currents and discharge rates in pyramidal neurons (Celada et al., 2008), causing an asynchronous mode of glutamate release (Aghajanian and Marek, 1999; Aghajanian, 2009) and spike-field decoherence (Celada et al., 2008). This decoherence has also been observed in vivo using dynamic causal modeling of MEG data, implicating psilocybin-induced increases in deep pyramidal cell excitability as the cause of broadband decreases in oscillatory power (Muthukumaraswamy et al., 2013). This effect of psilocybin implicates inhibitory interneurons, in the sense that oscillatory dynamics rest upon the interaction between pyramidal cells and fast-spiking inhibitory interneurons (Lisman and Buzsáki, 2008; Sohal et al., 2009; Lisman, 2012).
Similar adaptations of the REBUS model are described by the altered beliefs under psychedelics (ALBUS) model and related strengthened beliefs under psychedelics (SEBUS) model, suggesting that the strength of a dose calibrates the flexibility or rigidity of prior beliefs (Safron, 2020). The ALBUS model details possible dose-dependent differential effects of psychedelics on prior belief through their influence on inhibitory interneurons and the transmission of hippocampal/entorhinal system signals, (Safron, 2020). We suggest the alteration and strengthening of beliefs at specific doses may also be mediated by interaction with psychologic and environmental context, which is a point acknowledged by the ALBUS model. Furthermore, the mechanisms underlying altered beliefs may also be informed by work investigating reality monitoring (Dijkstra et al., 2022). The REBUS model is also reflected in neural annealing as reduced energy sinks (https://opentheory.net/2019/11/neural-annealing-toward-a-neural-theory-of-everything/) and are surmised to enable new directions for thought patterns under psychedelics without a complete breakdown of cognition (Petri et al., 2014; Atasoy et al., 2017). On this view, precision plays the role of an inverse temperature, where decreasing the precision of prior beliefs (i.e., increasing their temperature) makes it easier for higher-level beliefs to jump from one to another. In other words, the energy landscape is flattened. A useful analogy that captures the flattened landscape of priors under ego dissolution—and subsequent promotion of flexibility and plasticity—is that of a skier descending a hill after a fresh snow fall: Tracks (i.e., ruts), which were preferentially traveled previously (likened to prior belief updating), are leveled by the snowfall (i.e., the grooming action of psychedelics on high-level precision), resulting in a flattened landscape that enables travel in new directions without prior resistance. In short, psychedelics may simply flatten the “rut” and stop belief updating getting “stuck in a rut.”
Finding itself contained in a dynamic environment, the brain must possess a hierarchical generative model that can represent the world's contents, causes, and contexts (Bastos et al., 2015). We view ego dissolution as a breakdown of the generative model, particularly at the highest levels involved in our sense of identity and identification with the world. Psychedelics' net-excitatory effect on layer 5 pyramidal neurons at the upper levels of the hierarchy may, at its extreme, strip down the precision of the hierarchical generative model's priors of the world's contents, causes, and contexts. This makes sense because neurons in deep cortical layers represent perceptual hypotheses. We suggest this temporary breakdown alters the observer's sense of agency (i.e., own contribution to sensory input) and manifests a sense of heightened existential experience and primordial awareness.
Stated in alternative terms, the hierarchical generative model places boundaries on the physiologic states of living systems to support the behavior that ensures biologic survival (Friston, 2008). These boundaries need to be flexible enough to adapt to the dynamic environment. Belief updating supports adaptation by resolving mismatches in bottom-up prediction errors and top-down predictions and is itself supported by the coordination of hierarchical signals and the balance of integrated activity across distributed networks and regions that respond to internal and external contexts. At doses sufficient to induce ego dissolution, increased entropy may elicit a state of criticality wherein the ordinary cognitive boundaries of biologic system states are temporarily exceeded. This suggests reduced Bayesian model evidence and the extension of the distribution of hypotheses that underlie beliefs that make sense of the self and world. Cognitively, this may generate alternative models of the world and illustrate how our evolved, specialized faculties (i.e., connectivity) can construct it.
Top-down disinhibition may also factor into alternative models and may enable the desegregation of connectivity, which we suggest may further extend the probability distribution and cognitive flexibility at the level of consciousness and sentience. Together, beliefs organized along the hierarchical generative model may be afforded alternate hypotheses, and mentation may be afforded the exploration of alternate states of thought, feeling, and perception. Therefore, serotonergic psychedelics may expose the possible constructions of the world, and these possibilities may offer alternative therapeutic pathways for beliefs about the relationships within it.
This gives some indication of how the effect of serotonergic psychedelics may alter the generative model's role in sentience and planning, which speaks to the function of serotonin and the layer 5 apical dendrites in the emergence of consciousness. Advancements in psychedelic imaging analyses are uncovering more specific mechanisms underwriting connectivity changes proposed by unifying theories. For example, entropy changes in the brain, once described as uniformly elevated across the brain, are now suggested to vary and decrease across brain regions (Herzog et al., 2020). Similarly, recent work using “template” independent component analysis revealed the mediodorsal and pulvinar thalamic nuclei connectivity between visual connectivity and the DMN in association with subjective effects under psilocybin (Gaddis et al., 2022). The thalamus is composed of as many as 60 nuclei (Cassel and de Vasconcelos, 2015) and provides one example of the precision required to identify the neural mechanisms of psychedelics. As research moves forward and the need for more precise estimations of connectivity changes are demanded, combining methods that isolate highly specific regions with the precision DCM can advance our understanding of the brain-behavior relationship under psychedelics.
6. Future Research Priorities
Neuroimaging research provides a basis for interpreting psychedelic effects on consciousness and lasting behavioral outcomes. As has been described above, psychedelic changes span the hierarchical organization of the brain—from binding to serotonergic receptors to the downstream effects of oscillatory patterns on specialized regions and network connectivity. Measurement of correlates of psychedelic changes to neurons that form brain connectivity can contribute to our understanding of the biologic underpinnings of psychedelic subjective effects. Analysis methods such as measuring the cytoarchitecture of the brain’s hub regions may be a focus of future research. Cytoarchitecture research suggests that rich links between brain hubs can be explained by genetic factors, whereas peripheral links are better explained by environmental influences (Fulcher and Fornito, 2016). The desegregation of FC from network pathways under psychedelics (Lord et al., 2018; Varley et al., 2019; Luppi et al., 2021) also demonstrates deviation of connectivity mediating rich links that is consistent with the observations of increased sensitivity to intrinsic and extrinsic perturbations (i.e., environmental influences) under psychedelics.
Hierarchical interactions are necessarily directed (i.e., bottom-up and top-down). Functional connectivity measures are blind to directed connectivity. This suggests that more focus should be placed on the analysis of effective connectivity changes under psychedelics. Correlation-based whole-brain FC techniques can be complemented by the ability of DCM to infer the directed causes of neuronal responses. This technique is capable of parcellating the brain hierarchy and identifying the direction and strength of message-passing across subcortical and cortical levels. Crucially, unlike functional connectivity, the effective connectivity estimates supplied by DCM include intrinsic self-connections, namely the intrinsic excitability of different regions or neuronal populations. This is important in terms of estimating the effective precision or synaptic gain of a population under a hierarchical predictive coding formulation. In short, DCM can link synaptic level mechanisms described in hierarchical predictive processing to the directed connectivity changes of region level neural substrates.
a. Separating subjective effects from therapeutic outcomes
The necessity of subjective effects—for therapeutic outcomes—remains a debate that can be informed by further neurobiological study. Most clinical psychedelic research suggests that the psychologic aspect of experience is crucial to therapeutic outcomes (Yaden and Griffiths, 2020). This position can be understood through the influence of serotonin on precision control and suggests that psychedelics act upon the very mechanisms of sentience and attention, which may lead to therapeutic outcomes. Furthermore, the efficacy of these mechanisms is argued to depend on the psychologic mindset of individuals and the setting of administration to successfully ameliorate ego resistance (suggested here to be a prerequisite to positively felt ego dissolution). However, despite these factors, some research suggests that the biologic effects of psychedelics may be divorced from the subjective psychedelic experience while retaining therapeutic properties (Olson, 2020).
Preclinical research has demonstrated that rats can achieve long-lasting antidepressant effects from a single administration of a psychedelic (Hibicke et al., 2020). The authors suggest that although we do not know the mind of rodents, it is reasonable to suspect they do not use insight in the same manner as human subjects. Their evidence indicates that the subjective experience may be unnecessary and only a correlate of doses sufficient to alter cellular and structural changes. In this case, the subjective experience of ego dissolution might be a marker predicting a successful therapeutic outcome.
Differential dosing in preclinical and clinical trials may help evaluate the necessity of subjective experience in humans. Measured by body weight, the lowest doses in preclinical studies typically resemble the highest doses in clinical studies (de Vos et al., 2021). The typical psilocybin oral administration of 25 mg used in human clinical trials roughly equates to a 3–3.5 mg intravenous delivery, and human imaging research reports extreme intense subjective effects in some participants when administered 2.0 mg for the total body weight of a human (Carhart-Harris et al., 2012b). Hibicke et al. (2020) injected rats with a much more significant amount of psilocybin, 1.0 mg/kg. However, body weight–adjusted dosing overlooks the importance of allometric scaling, which accounts for differences in pharmacokinetics and pharmacodynamics among species (Nair and Jacob, 2016). Using allometric scaling, the dosing between Hibicke and colleagues’ preclinical study and the oral doses used in human research that elicits subjective effects may be roughly equivalent.
Moreover, structural and cellular changes at extreme doses may be sufficiently therapeutic in humans and preclude the need for psychologic mediation. For example, a case study of a 15-year-old female individual who had previously suffered from 3 years of chronic depression accidentally consumed 10 times the standard dose of LSD (1200 µg). Following the incident, she reported no symptoms of depression for the next 13 years (Haden and Woods, 2020). A separate case also found that chronic pain was alleviated following a massive overdose (550 times the standard dose) of LSD (Haden and Woods, 2020). Neurobiological mechanisms may have therapeutic utility at unsuitable doses for humans. However, it remains plausible that the psychedelic experience facilitates underlying neurobiological changes. The role of psychologic mediation is reinforced by considering the effect of context on the subjective quality of the psychedelic experience. Supportive contexts can help elicit positively felt ego dissolution and ameliorate resistance that encourages a therapeutic response and associated revision of self-beliefs, whereas suboptimal environment and preparation may produce null results or be psychologically detrimental (Mashour, 2007; Carhart-Harris et al., 2018b). The capacity of environmental conditions and peer support to direct the psychedelic experiences point to a biopsychosocial model of psychedelics and signifies the importance of psychologic mediation to the therapeutic effects at doses safely administered to humans.
The theory that psychedelic therapeutic effects depend upon psychosocial contributions will be falsified if equal efficacy in the same range of pathologies can be achieved without subjective effects. Such an outcome suggests that the psychedelic aspect of psychedelics may be superfluous. This appears unlikely in the face of reported therapeutic experiences and outcomes, such as in the case of end-of-life psilocybin treatments (https://maps.org/news/bulletin/my-experience-as-a-guide-in-the-johns-hopkins-psilocybin-research-project/). Psychedelics change how we make sense of the world. It may then also be reasoned that, insofar as our thoughts guide actions, psychedelic change to beliefs in response to subjective experiences may reorient behavior and serve an important role in mediating the therapeutic outcomes. We still have much to learn about how experiences of ego dissolution influence long-term behavioral outcomes. Being rated among the most meaningful experiences of one's life suggests that even as the emotional and therapeutic effects subside, the experience may remain a meaningful impression that helps shape personal identity, world view, and values as much as any other meaningful life experience. Likewise, a bad experience may also shape mental health outcomes and perception of reality (Bienemann et al., 2020). However, quantification of long-term effects of ego dissolution is lacking, and longitudinal studies from breakthrough therapeutic experiences are encouraged. Palliative care experiences of unity suggest that ego dissolution is exceptionally meaningful (https://maps.org/news/bulletin/my-experience-as-a-guide-in-the-johns-hopkins-psilocybin-research-project/). These existential experiences can bring about personal meaning, closure, and peace for those facing end of life. In these instances, ego dissolution's emotional and psychologic value is arguably immeasurable. Similar outcomes are also observed in clinical outcomes outside of palliative care (Malone et al., 2018; Noorani et al., 2018) and provide evidence that psychologic mediation is an essential aspect of psychedelic therapeutic effects.
Research efforts to understand the relationship between subjective effects and therapeutic qualities and remove the psychedelic experience while preserving therapeutic effects are a growing area of pharmacological research. Psychedelic mechanisms separated from their psychologic component may have utility, although a more plausible outcome may be a reduced efficacy in pathologies currently recommended for psychedelic therapy or findings that suggest the efficacy of drugs derived from psychedelics in alternate domains of medicine. In these eventualities, it may be argued that a unique class of drug has been invented and that the biopsychosocial approach to classic psychedelic administration for therapeutic uses remains efficacious.
Future pharmacology studies are sure to test this hypothesis and determine the limits of separating subjective experience from biologically induced therapeutic benefits. One avenue may be combining of anesthesia and psychedelic doses considered unsafe in the waking state. Neuroimaging research can also contribute to this line of inquiry. Research at the macroscopic level of brain regions and networks, and their integration can address this question by delineating precise mechanisms. For example, measuring the effective connectivity between brain regions influenced by the 5-HT2AR involved in longitudinal studies may indicate whether changes associated with regions involved in consciousness correlate with well-being. This may indicate how therapeutic effects depend on changes in brain connectivity associated with consciousness and their separability from subjective effects. Meanwhile, interest in the manipulation of psychedelic molecules and research building upon pretreatments acting as selective inhibitors of subjective effects (Vollenweider et al., 1998; Quednow et al., 2012; Preller et al., 2019) will likely provide stronger understanding of the reliance of therapeutic outcomes upon subjective effects.
b. Determinants of dose-response and behavioral measures of ego dissolution
Uncertainty exists whether ego dissolution involves a gradual onset or a binary (i.e., all or nothing) experience. Evidence suggesting the binary (i.e., nonlinear) dose-response hypothesis is supported by PET investigation of plasma psilocin occupancy (the active metabolite of the prodrug psilocybin) and the reported subjective intensity of experience (Madsen et al., 2019). However, visual schematics illustrating subjective responses of oceanic boundlessness (i.e., ego dissolution) counter this assertion by providing evidence of a gradual onset (Fig. 7) (Hirschfeld and Schmidt, 2020).
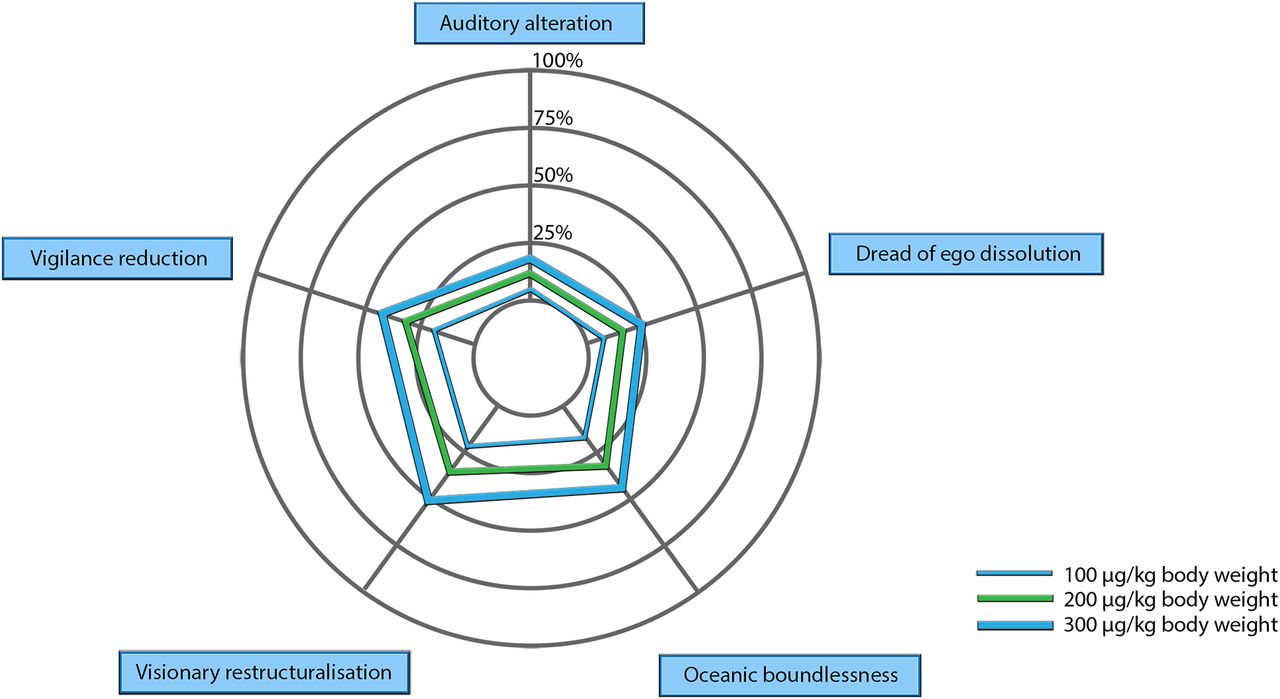
Ego dissolution rating by body weight–adjusted psilocybin dose, adapted from Hirschfeld and Schmidt (2020)’s review of psilocybin studies using the 5D-ASC. Psilocybin doses assigned by varying body weights suggest ego dissolution (oceanic boundlessness) may be amplified in a linear, dose-dependent manner (i.e., gradual) (Hirschfeld and Schmidt, 2020).
We approach the conflict between gradual and binary dose-response curves by proposing that alterations to high-level observer constructs (i.e., self or ego and meaning) occur as a binary shift. In contrast, lower-level observed constructs encounter gradual dose-dependent changes, for example, in visual alterations. See Safron (2020) for related views. Paradoxically, psychedelic desegregation of connectivity pathways and the collapse of the predictive hierarchy may reduce differentiation between hierarchical levels and contribute to blurred subject-object boundaries. Our review characterizes ego dissolution as the emergent shift in the observer. Like the mystery of consciousness, ego dissolution is suggested to be a phenomenon that is greater than the sum of its parts. Those parts represent psychedelic altered object phenomena such as visual, temporal, and emotional alterations. We also recognize the limitations of comparing self-reports across subjects with different prior experiences and references to base their responses. To measure the binary onset of ego dissolution, researchers conducting imaging studies may wish to note momentary metacognitive lapses that subjects report in their stream of consciousness—for example, experiences of disembodiment.
Time-dependent connectivity mechanisms of ego dissolution may be helpful to answer the dose-response question and have been used in the work of Preller et al. (2020). However, this study used a body-weight dose equivalent to 14 mg per 70 kg, whereas the standard “full” dose used in therapeutic trials is typically 25 mg. Sequential time-dependent scanning and behavioral measures across high-dose onset may require psychedelic-experienced participants and enable better measurement of the correspondence between connectivity changes and the onset of ego dissolution. Alternatively, imaging efforts may attempt to demarcate shifts in consciousness using the high temporal specificity of EEG and MEG modalities. Follow-up analysis may then observe whether a transition in brain connectivity accompanies reports of ego dissolution during brain imaging.
A related controversy pertains to psychedelic doses adjusted by body weight. Weight-adjusted doses have been the standard form of dose administration across neuroimaging trials; however, recent evidence suggests no influence of body weight on subjective effects of psilocybin (Garcia-Romeu et al., 2021). This research also marks the need for greater understanding of the mechanisms controlling the relationship between dose and response. Moreover, subjective responses can vary significantly following a standard or weight-adjusted dose of a psychedelic (Preller et al., 2019). Several approaches to the dose-response question may be entertained. For example, greater understanding of relationship between subjective effects and the density and distribution of serotonin receptors may be informative. Measures of the volume and functional connectivity profiles of individual brain structures may also predict the dose-response relationship and warrants future investigation. For example, baseline global connectivity may be associated with subjective responses to psilocybin (Preller et al., 2020) and intensity of ego dissolution has been linked to a low diversity of connections in the anterior parahippocampus (Lebedev et al., 2015). Structural predictors of dose responses are also intriguing, following evidence that psychedelics can influence particular region volumes in long-term users (Bouso et al., 2015) and rACC thickness can predict emotional subjective effects (Lewis et al., 2020). To date, no method has been established to predict dose-response relationships in psychedelic naive participants. Neuroimaging studies sensitive to associations between the density and distribution of receptors, region structural volume, measures of connectivity and the subjective response of participants may be valuable in determining predictors of dose-responses and enable the tailoring of doses for clinical applications. Post-hoc analyses can also contribute to answering the dose-response question by determining how effective connectivity pathways between key brain regions and hierarchical strength (calculated by summing efferent and afferent region connectivity) (Zhou et al., 2018) between networks predict aspects of the experience reported on behavioral measures. Understanding the connectivity mechanisms of the dose-response relationship may provide biomarkers that determine the optimal dose in therapeutic contexts and, in turn, identify the mechanisms of subjective effects and ego dissolution.
Furthermore, improvements in existing behavioral measures administered after psychedelic ingestion are crucial to advancing research. At present, aspects of the psychedelic experience such as ego dissolution are lumped into a single measure of changes to sense of self and the relationship between subject and object. However neuroscientific and consciousness research suggest that the self can be dichotomised into different aspects related to distinct networks, such as the SN (i.e., minimal self) and DMN (i.e., narrative self). In associating connectivity patterns to ego dissolution, it may be essential to implement subjective measures sensitive to discriminations between minimal and narrative aspects of the self. These independent behavioral measures may then be associated with underlying connectivity and inform existing theories of SN and DMN contributions to ego dissolution. This will help determine network-dependent changes to consciousness under psychedelics with greater accuracy.
c. Comparative research
Our knowledge of psychedelics may also benefit from protocols that compare connectivity changes observed across different classic psychedelics and a variety of other altered states of consciousness including psychosis and meditation (or even sleep). Parallels between the early stages of psychosis and psychedelic reports was one of the first insights into the nature of psychedelic-induced conscious states. However, this parallel may have been biased by reports of negatively felt ego dissolution that was more likely in studies which did not appreciate the importance of supportive set and settings (Carhart-Harris et al., 2018b). Findings from these studies were more likely to resemble psychotic disturbances. Appreciation of the influence of context—on the valence of psychedelic experiences— motivates further study of both set and setting. Research testing the influence of factors contributing to the administration settings could manipulate imagery, lighting, sound, interactions with stimuli (e.g., walks in nature or experiences in virtual reality) and whether the administration of psychedelics occurs in an individual or group setting. Mindset may also be manipulated by controlling for subjects with specific characteristics, dispositions and training. The importance of mindset can also be demonstrated by three key manners in which psychedelic experiences differ from experiences of psychosis. These include consent that is given prior to administration, targeted use of psychedelic experience that may involve an intention for administration, and the temporary duration of the psychedelic experience. Provision and assurance of these factors are important contextual determinants expected to promote positively felt loss of self that differentiates psychedelic states from psychosis. Moreover, preparing mindset may influence the organized coherence of dysregulated and disintegrated connectivity under psychedelics (Atasoy et al., 2017). This hypothesis can be tested by testing for correlations between connectivity and participants’ behavioral reports of positively felt ego dissolution (i.e., oceanic boundlessness on the 5D-ASC scale) and dread of ego dissolution (i.e., dread of ego dissolution on the 5D-ASC scale). Between subject studies can also be used to identify connectivity mechanism underlying specific facets of experience such as blissful state and sense of unity (Carhart-Harris, 2019) that may demonstrate unique forms of coherence or stability of brain states (Atasoy et al., 2017) during ego dissolution.
The importance of supporting mindset factors speaks to the value of comparative research between meditation and psychedelics. Many authors have highlighted similarities and differences between meditation and psychedelics that provide a basis for future neuroimaging studies. The utility of meditation as an adjunct to psychedelic therapy is supported by research that found nominal anxiety in meditators (Smigielski et al., 2019a) and enhanced depth of meditation (Smigielski et al., 2019b). This synergistic approach—also possible with music (Kaelen et al., 2018)—can be used to evaluate the benefits to behavioral outcomes and accompanying connectivity changes. The importance of integrating meditation in psychedelic study design is further highlighted by evidence that the quality of psychedelic experiences may determine therapeutic outcomes (Roseman et al., 2018b). Future study designs could integrate meditation training prior to psychedelic use to prepare participants, enhanced positive ego dissolution and measure connectivity changes associated with the inclusion of meditation. They may also test the effect of meditation programs after the psychedelic experience as a form of integration to cement hypothesized neuroplastic changes that may underwrite behavioral change. Some work in this direction (previously discussed) has been undertaken by Griffiths et al. (2018) (Smigielski et al. 2019a,b); however, relatively less focus has been placed upon programs implemented post hoc. In summary, equipping neuroimaging protocols with groups assigned to meditation programs can help identify the connectivity differences based on meditation and psychedelic synergy.
d. Consciousness research
Psychedelics show value as a tool for a neuroscientific understanding of consciousness. The search for the neural correlates of ego dissolution require the minimally sufficient neural prerequisites of consciousness to be identified to claim that the neural activity correlated with consciousness has been determined (Chalmers, 2000). Ego dissolution may not encompass the minimally sufficient neural prerequisites of consciousness, because some important aspects of consciousness are retained in psychedelic experiences. However, they may still reveal some neural prerequisites of consciousness if change to connectivity induced by psychedelics is found to operate exclusively in conscious states (Kalat, 2014; Millière, 2017). Future neuroimaging research could identify neural mechanisms underlying psychedelic subjective effects and states of ego dissolution that are distinct from other states of transcendence, such as meditation.
e. Plasticity
Apart from understanding the brain connectivity that underlies ego dissolution and its potential to inform our understanding of consciousness, the related idea of psychedelic-induced neuroplasticity proposes a fruitful avenue for future psychedelic research. The notion that psychedelics leave the brain in a more plastic (i.e., malleable) state has a substantial body of evidence. Preclinical evidence shows that psychedelics render the brain in a more malleable state. For example, C-Fos expression (a marker of neural activity) is identified across brain regions in mice under psilocybin (Davoudian et al., 2022), DMT developed new neurons in the hippocampus of mice (Morales-Garcia et al., 2020), and the development of dendritic spines in neurons and synaptic connectivity are observed in the prefrontal neurons of rats under DMT (Ly et al., 2018). Dendritic change can alter the enhancement and suppression of membrane excitability, which may create opposing actions on dendritic excitability that promote plasticity (Savalia et al., 2021). Although the cellular mechanisms leading to therapeutic efficacy are not yet understood, serotonergic psychedelics stimulation of neural and glial cells may reduce inflammation and reduce oxidative stress and act as disease-modifying therapeutics in neurodegenerative disorders (Kozlowska et al., 2021). Biomarkers of plasticity are also demonstrated in clinical research, showing elevation of blood plasma brain-derived neurotrophic factor (BDNF) after administration of 200 μg of LSD compared with placebo (Holze et al., 2021). BDNF is a protein molecule that promotes the survival and growth of neurons and plays a crucial role in plasticity (e.g., learning and memory) (Miranda et al., 2019). Interestingly, this study also found that the 200 μg of LSD required to increase plasma BDNF went above the ceiling of therapeutic (i.e., positively felt) ego dissolution (Holze et al., 2021). It remains possible that a dissociation exists between psychedelic neurobiological and psychologic therapeutic mechanisms.
Similarly, abundant evidence suggests lasting behavioral changes, including behavioral flexibility in rats (Hibicke et al., 2020). However, further research is required to identify the synaptic changes and ensuing effects on connectivity in humans and determine what biologic changes exist after psychedelics have been fully eliminated. Understanding this could elucidate the synaptic mechanisms supporting lasting impressions on consciousness that underwrite therapeutic benefits. One line of inquiry—connecting neural mechanisms to lasting behavioral change—is measures of entropy induced by psychedelics that have been related to personality change (Lebedev et al., 2016; Liechti, 2017). Psychedelic-induced entropy may relate to increased openness, a personality trait connected to schizotypy and antithetical to ego resistance. Schizotypy describes an imaginative state of mind that, in extreme presentations, shares similarities with psychosis. However, this facet of personality is also associated with beneficial outcomes (Mohr and Claridge, 2015; Schultze-Lutter et al., 2019).
Plasticity corresponds to measures of dynamic flexibility in brain networks. When flexibility in the brain exists within metastable ranges, it can be associated with positive adaptive behaviors, such as learning (Bassett et al., 2011) and the desire for cognitive endeavors (i.e., a personality trait known as the need for cognition) (He et al., 2019). However, beyond metastable ranges, elevated flexibility is a biomarker of schizophrenia and increased risk of psychosis (Braun et al., 2016). The parallels between psychedelic states and psychosis may derive from the elicitation of criticality that induces neural and behavioral plasticity. Observations associating behavioral responses to psychedelics (i.e., positive or negatively felt ego dissolution) and measures of flexibility may offer clinical insight. For example, measures of flexibility can be measured as disjointed or cohesive flexibility and have been associated with different behavioral outcomes (Safron et al., 2022). Uncovering the specific dynamics of neural flexibility in regions and networks and their correspondence to behavioral properties may be vital to understanding the effects of psychedelics. For further review of forms of flexibility in brain dynamics and their functional significance, readers are directed to Safron et al. (2022).
Neuroimaging research can clarify the link between connectivity and behavior (associated with openness) by estimating connectivity before and after psychedelic use. This may provide an indication of neuroplastic changes in the brain. Similar efforts already exist showing evidence of connectivity changes in the AMG lasting up to 1 month (Barrett et al., 2020a) and mPFC-PCC in meditators performing meditation in the scanner post–administration of psilocybin (Smigielski, et al., 2019b). These findings suggest that underlying neuroplastic mechanisms may account for lasting connectivity changes and behavioral outcomes; however, the dose-dependent window of plasticity and long-term structural changes remains largely unknown (Banks et al., 2021). Thorough investigations of neural plasticity across the brain remain a direction of future neuroimaging research and may discover important clinical applications such as advancing the understanding of the pathophysiology and prevention of neurodegeneration. These future studies can study the influence of psychedelics on regional connectivity changes over time after administration. This research can also be refined by investigating how contextual determinants and psychologic support influence connectivity changes indicative of plasticity and how biologic processes may translate into psychologic growth. These changes may also benefit nonclinical populations by enhancing social and cognitive skills, empathy, and creativity (Mason et al., 2019), as well as general well-being (Smigielski et al., 2019a; Yaden and Griffiths, 2020; de Vos et al., 2021). Optimized integration programs following psychedelics that cement insights developed during ego dissolution may capitalize on hypothesized neuroplastic benefits of reduced ego resistance. Measuring connectivity differences following effective psychedelic integration can advance our ability to discern and alter brain connectivity for well-being.
However, it is crucial to recognize that psychedelic-induced neural plasticity is undirected and not inherently therapeutic (Branchi, 2011). Early psychologic research demonstrates that emotional arousal can be stressful or pleasurable based on the cognitive appraisal (Folkman et al., 1986). Similarly, the psychologic response to psychedelic effects is likely to mediate the therapeutic utility of psychedelic-induced plasticity. Psychedelic-reduced belief constructs may open the range of alternative hypotheses entertained by beliefs while not guaranteeing a therapeutic direction. How mindset and environment prepare, support, and influence resistance or surrender to ego dissolution is a crucial determinant of the valence of ego dissolution. For example, research shows that anxious ego dissolution does not produce the same therapeutic quality as positively felt ego dissolution (Roseman et al., 2018b; Russ et al., 2018; Russ et al., 2019). When ego dissolution is accepted, psychedelics can reduce belief constructs bound to the self. We argue that, in humans, these changes to beliefs may outlast any physiologic or neurobiological effects and speak to the importance of the interaction between psychosocial influences and altered synaptic processes, at apical dendrites, in how we make sense of the world. Neuroplasticity suggested by brain changes and sensitivity to context may procure opportunities for personal and existential reflection most optimally when a privileged (i.e., prepared, supportive, and worry free) environment allows the disarming of ego and concomitant dissolving separation of the self from the world.
II. Conclusions
Historical records indicate the use of classic psychedelics to alter consciousness and manifest insight has existed in cultures around the world since antiquity. Their popularization in Western culture during the mid-20th century affected the thoughts and feeling of many who used them, leading to a revision of sociopolitical beliefs. The backlash of this threat to hierarchical political authority was the sensationalism of fears and legislation that cast shade over psychedelic research until the turn of the millennium. Motivated by academic psychiatry, researchers revisited the signs of potential indicated by early psychedelic research. Their findings challenged the legislation of psychedelics as substances with no therapeutic or scientific value. Since this ground-breaking research, which demonstrated the efficacy of psychedelics, research into their therapeutic applications has accelerated, and consciousness research has followed to understand the underlying mechanisms of therapeutic outcomes and subjective effects.
In the late 1970s, it was discovered that 5-HT2 receptors have a vital role in regulating responses to psychedelics. Subsequent pharmacological research in recent decades has cloned the 5-HT2A receptor and identified the 2A subtype as the primary target for psychedelic's effects on consciousness. 5-HT2AR agonist activity results in entropic changes influencing the synaptic gain of neural populations, which affects precision control underwrites inferences about the causes of sensation and consequences of actions. Understanding the effect of serotonergic activity on whole-brain connectivity poses significant challenges. Different brain regions have specialized functions, and these regions are functionally integrated into networks that serve roles underlying conscious processes. Applying an understanding of regional and network-level function in consciousness to psychedelic subjective effects, underwritten by changes to oscillatory rhythms (i.e., connectivity) within and between brain substrates, is a promising direction for future psychedelic neuroimaging research. Here, we reviewed regional and network-level analyses of psychedelic changes to brain connectivity, which identified mechanisms that may contribute to ego dissolution and therapeutic effects. Neural mechanisms and hypotheses of altered brain connectivity under psychedelics such as entropy, criticality, and plasticity reinforce each other and help explain behavioral flexibility. For example, lasting personality change is demonstrated as increased openness to experience. These features of psychedelic effects are also harmonious with observations of reduced connectivity and desegregation of associative resting-state networks. Moreover, synchronous patterns of acute psychedelic effects reformatting consciousness and sentience are reflected in more specific mechanisms. The disintegration of the DMN and midline cortical connectivity—and their inhibitory function—and reduced thalamic gating also synthesize to give a sense of how the loss of self experienced in ego dissolution may occur and afford alternate pathways that facilitate the generation of meaning and revision of beliefs.
Neuroimaging endeavors are required to address a range of questions probing the nature of psychedelics. Answers to these questions depend on (repeated) neuroimaging experiments that test hypotheses of connectivity changes and advancement of behavioral measures. We will then be able to more precisely discriminate features of subjective experience associated with brain connectivity and provide further empirical evidence for unifying frameworks such as hierarchical predictive coding.
Hierarchical predictive coding provides a biologically plausible computational model of sentience and serves as a model of neuronal function that is an essential building block to understanding neural processes. Neuronal processes of hierarchical predictive coding form the structure of the generative model that is able to represent dichotomy between self and contents and causes and contexts of the world. Hierarchical predictive coding explains serotonergic synaptic mechanism’s role in making sense of the world through top-down predictions (or inferences) and bottom-up prediction errors (that drive belief updating). These neural mechanisms offer a means of synthesizing psychologic and neuroscientific interpretation of psychedelic neuroimaging observations. Understanding the link between psychology and neurology is crucial, following behavioral evidence that reaction to a dose of a psychedelic substance depends upon the environment in which is it is taken and mindset of the subject. This evidence indicates the importance of understanding the role of biopsychosocial factors. These factors may also be crucial to predicting and supporting therapeutic efficacy. Future therapeutic research is encouraged to support positively felt ego dissolution by cultivating reduced ego resistance. Cultivating this attitude to the onset of psychedelics sets the stage for ego dissolution and surrender to the psychedelic experience. This may entail manipulating and optimizing a subject’s preparation, the administrative setting, and integration programs designed to cement psychedelic insights into lasting well-being. Further understanding of the interactions of biopsychosocial influences of serotonergic neural mechanisms can be aided by neuroimaging analyses that observe differences between the brain connectivity of successful and unsuccessful psychedelic interventions. This may help explain neural mechanisms underlying well-being, hypothesized to be underwritten by neuroplasticity, and suggests the important contribution of psychedelic neuroimaging to psychiatry.
Hierarchical predictive coding also offers a framework for understanding the neural mechanisms of consciousness. Psychedelics provide hierarchical predictive coding with a means of testing its modeling of sentient behavior and advancing the understanding of serotonergic predictive processes in consciousness and attention. The alteration of these synaptic processes by psychedelics results in subjective effects, argued here to be an essential component of therapeutic outcomes. Hierarchical predictive coding explains subjective and therapeutic mechanisms at the neural level, which suggests that further investigations that can quantify the changes to the hierarchical organization of connectivity under psychedelics are essential. Dynamic causal modeling offers a method of testing changes to hierarchically organized connectivity and can be synthesized with hierarchical predictive coding to help model change to brain connectivity involved in consciousness and attention. Applying the understanding of predictive processes may also help synthesize interpretations across various neuroimaging analysis methods that describe macro- and microscale changes to the brain and their association with subjective effects.
Through psychedelic alteration of serotonergic receptors, the hierarchical relationship between top-down associative processes and bottom-up sensations are collapsed. This collapse inhibits associative functions of regions and networks involved in conscious beliefs and perceptions and enables a landscape in which bottom-up influences can update beliefs about interactions with the world. This temporary collapse is reflected in the breakdown of self, evidenced by ego dissolution, and the updating of beliefs is demonstrated in therapeutic insights leading to changes in patterns of thought and perspective. Psychedelic ego dissolution deconstructs the generative model’s function to maintain beliefs and how we make sense of the world. When this occurs with limited resistance on the part of the observer, personal and existential reflections can realize alternate pathways of thought and behavior, which can be incredibly imaginative and potentially therapeutic. Plasticity is hypothesized to underlie these changes, whereas the exact nature of the relationship between beliefs, behavior, and structural changes is still in the early stage of investigation. Fully realized therapeutic ego dissolution can involve the overcoming of fear—fear of loss or change, adapting to psychologic change, and forming novel connections—that may be underwritten by serotonergic-facilitated amplified, disintegrated, and desegregated connectivity changes. The therapeutic effects of psychedelics may depend on physiologic changes. However, the relationship between subjective experience, belief change, and mental health outcomes appears complex and may be mediated by biopsychosocial interactions that follow psychedelic experiences.
Our review suggests that ego dissolution is the shift in consciousness that manifests from an array of brain changes, primarily to the disintegration of the self-related associative connectivity and desegregation of connectivity involved in space, time, and various other subjective effects that occur with ego dissolution. We refer to gestalt psychology to explain ego dissolution. The whole of various observed psychedelic effects may manifest as greater than the sum of their parts in changes to the observer. We suggest that the shift in the experience of self and psychedelic effects relates to the hierarchical breakdown of the distribution of priors that compose the generative model, which affords the generation of alternate models of the world and boundaries of the self.
Interestingly, the introduction of psychedelics to Western culture reflects a similar process of reidentification through challenge to the hierarchical authority of top-down beliefs from political leadership and bottom-up grassroots influences. This suggests a common effect of psychedelics across neurology and sociology, in which boundaries limiting perception are broken. Moreover, it indicates a reflection between the hierarchical modeling of the brain and social organization. Surmising that the experience of ego dissolution exists at the center of the psychedelic experience and that its neural mechanisms may be close to the roots of the subject-object dichotomy may explain why phenomenological encounters with this experience can be intimate and fundamentally change self-identification and interactions with the world. Continued investigation of the neural mechanisms of ego dissolution and the dichotomy of separation between the self and the world may advance scientific understanding of sentience. The value of psychedelics may then, under appropriate conditions, be their power to catalyze meaningful philosophical and scientific understanding of the “u” (i.e., self) in the universe.
Acknowledgments
A.R. is a CIFAR Azrieli Global Scholar in the Brain, Mind, and Consciousness Program. A.R. and K.F. are affiliated with The Wellcome Centre for Human Neuroimaging.
Authorship Contributions
Wrote or contributed to the writing of the manuscript: Stoliker, Egan, Friston, Razi.
Footnotes
- Received September 29, 2021.
- Revision received June 26, 2022.
- Accepted June 29, 2022.
This work was supported by the Australian Research Council [Grants DE170100128 and DP200100757], the Australian National Health and Medical Research Council Investigator Grant [1194910], and the Wellcome Trust [Grant 203147/Z/16/Z].
The authors declare no conflicts of interest.
Abbreviations
- 5D-ASC
- 5 dimensions of altered states of consciousness scale
- 5-HT
- 5-hydroxytryptamine
- 5-HT2AR
- 5-HT2A receptor
- ACC
- anterior cingulate cortex
- AI
- anterior insula
- ALBUS
- altered beliefs under psychedelics
- AMG
- amygdala
- BDNF
- brain-derived neurotrophic factor
- dACC
- dorsal anterior cingulate cortex
- DAN
- dorsal attention network
- DCM
- dynamic causal modelling
- DMN
- default mode network
- DMT
- N,N-dimethyltryptamine
- EDI
- ego dissolution inventory
- EEG
- electroencephalography
- FC
- functional connectivity
- FEP
- free energy principle
- fMRI
- functional magnetic resonance imaging
- FPCN
- frontoparietal control network
- HC
- hippocampus
- LSD
- lysergic acid diethylamide
- MEG
- magnetoencephalography
- mPFC
- medial prefrontal cortex
- MTL
- medial temporal lobe
- OAV
- altered states of consciousness rating scale
- PCC
- posterior cingulate cortex
- PET
- positron emission tomography
- PHC
- parahippocampus
- rACC
- rostral anterior cingulate cortex
- REBUS
- relaxation of beliefs under psychedelics
- RSC
- retrosplenial cortex
- SN
- salience network
- Copyright © 2022 by The Author(s)
This is an open access article distributed under the CC BY-NC Attribution 4.0 International license.
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}