
Abstract
Nicotinic acetylcholine receptors (nAChRs) have been implicated in a number of neurological disorders. 5-Iodo-3-[2(S)-2-azetidinylmethoxy]pyridine (5-I-A-85380) is a novel nAChR marker, binding predominantly to the α4β2 subtype. This in vitro autoradiography study describes the distribution of 5-[125I]-A-85380 binding in post-mortem brain tissue from normal elderly individuals and from cases with age-associated dementias of both neurodegenerative and vascular types. The binding distribution of 5-[125I]-A-85380 in normal brain tissue was found to be consistent with the reported distribution of other high-affinity nicotinic ligands. In addition to high thalamic and moderate striatal and temporal cortex density, moderate 5-[125I]-A-85380 binding was also seen in white matter tracts in cingulate, occipital, and temporal areas, indicating the presence of nAChRs along nerve fiber tracts, which has not been reported in other high-affinity nicotinic agonist distribution studies. In Parkinson's disease (PD), loss of striatal 5-[125I]-A-85380 binding closely parallels the loss of nigrostriatal dopaminergic markers previously observed. In dementia with Lewy bodies (DLB) reduced striatal 5-[125I]-A-85380 binding density, comparable to that in PD, may be a marker of early degeneration in nigrostriatal inputs, while in Alzheimer's disease (AD) reduced striatal 5-[125I]-A-85380 binding could be related to reduced cortical inputs. The reductions of nAChRs seen in AD, DLB, and PD were not apparent in vascular dementia (VaD). In conclusion, 5-I-A-85380 is clearly a useful ligand for both in vitro and in vivo single photon emission tomography human studies investigating disease symptoms and progression, response to acetylcholinesterase-inhibiting drugs and in differentiating primary degenerative dementia from VaD.
Similar content being viewed by others

INTRODUCTION
Nicotinic acetylcholine receptors (nAChRs) are ligand-gated ion channels that are widely distributed in the human brain where they have a modulatory function associated with numerous transmitter systems. The major nAChR subtypes present in the mammalian brain are α7 and α4β2 (Lindstrom et al, 1995; Holladay et al, 1997). The majority of brain nAChRs with a high affinity for agonists contain α4 and β2 subunits (Picciotto et al, 1995; Marubio et al, 1999). [3H]cytisine and [3H]ABT-418 predominantly bind to α4β2, whereas [3H]epibatidine and [3H]nicotine bind in addition to α3 and possibly other subunit-containing receptors (Flores et al, 1992; Sihver et al, 1998; Zoli et al, 1998).
Reductions in nAChR density have been identified in a number of neurodegenerative disorders including Alzheimer's disease (AD), dementia with Lewy bodies (DLB), and Parkinson's disease (PD) (Rinne et al, 1991; Perry et al, 1995; Court et al, 2000a). In AD, binding assays with [3H]ABT-418 and [3H]epibatidine (Warpman and Nordberg, 1995) and immunochemical analysis (Martin-Ruiz et al, 1999; Wevers et al, 1999; Guan et al, 2000; Burghaus et al, 2000) indicate that the major contributor to loss of nAChRs in cortical areas is the α4 subunit. Deficits in [11C]nicotine binding in the hippocampus, temporal and frontal cortex have also been reported in vivo in AD (Paterson and Nordberg, 2000). In PD, the majority of reports indicate deficits in high-affinity nicotinic agonist binding in both the caudate and putamen (Perry et al, 1990a; Rinne et al, 1991; Aubert et al, 1992a; Court et al, 2000b), in parallel with the decline in dopaminergic markers (Piggott et al, 1999). Reduced [3H]nicotine binding in the substantia nigra (SN) pars compacta and ventral tegmental area has also been reported in PD (Perry et al, 1995). In DLB, losses in [3H]nicotine binding have been reported in the striatum, the SN pars compacta and pars reticulata, and the dorsolateral tegmentum (Perry et al, 1995; Court et al, 2000b).
5-Iodo-3-[2(S)-2-azetidinylmethoxy]pyridine (5-I-A-85380) is a high-affinity nAChR ligand that demonstrates high subtype selectivity for α4β2 nAChRs (Mukhin et al, 2000). A recent study has also shown 5-I-A-85380 binding to α-conotoxin MII-sensitive (α6β2) nAChRs in the rodent and monkey striatum (Kulak et al, 2002). The development of an in vivo α4β2 subtype-selective nAChR imaging agent will allow studies following disease progression, identifying where and when during the disease process nAChR deficits occur, and also provide information on how nAChR's expression relates to symptomatology and therapeutic response.
The present study investigates the distribution of 5-[125I]-A-85380 binding in the midbrain, neocortex, and cerebellum in normal elderly brain autopsy tissue by autoradiography. In addition, comparison of 5-[125I]-A-85380 binding was made between cases of AD, mild AD, DLB, PD, vascular dementia (VaD), and normal elderly aged-matched controls. The study aims to provide reference data for use in clinical research SPECT neuroimaging studies using 5-[123I]-A-85380.
MATERIALS AND METHODS
Materials
The trimethylstannyl precursor (TMS precursor) of 5-[125I]-A-85380 and cold 5-I-A-85380 was obtained from ABX, Germany. Chemicals used in radiolabelling and in autoradiography assays were obtained from Sigma Aldrich, UK and Vectabond was from Vector Labs, UK. Na125I and 125I autoradiographic microscale standards were obtained from Amersham Pharmacia Biotech UK Ltd, UK. High-performance liquid chromatography (HPLC) solvents were obtained from Rathburn Chemicals Ltd, UK and HPLC columns were obtained from Phenomenex, UK. Autoradiography film was obtained from Perkin-Elmer, Belgium and developer and fix was obtained from Kodak, HA West, UK.
Radiochemistry
5-[125I]-A-85380 was synthesized via a modification of a previously published method by Zoghbi et al (2001). Aliquots (300 μg) of TMS precursor were stored in nitrogen-filled, silanized vials at −18°C. A measure of 74 MBq, carrier free, Na125I in ∼18 μl of 0.01–0.1 M NaOH was obtained and was made up to 200 μl using 0.01 M NaOH. The radioiodination reaction was prepared by adding 20 μl 2 M HCl to the activity followed by addition of a mixed solution of 300 μg of precursor in 100 μl ethanol. The reaction was initiated by adding 50 μl (50 μg) of Chloramine T and the reaction mixture was then mixed by vortex. Mixing was continued during an incubation period of 15 min at 65°C. The reaction was quenched by addition of 100 μl (100 μg) NaHSO4. Deprotection was achieved by addition of 200 μl of trifluoroacetic acid and mixing. Mixing was continued during an incubation period of 15 min at 65°C.
Purification of 5-[125I]-A-85380 was carried out by injection of the reaction mixture onto a reverse phase C-18 semi-preparative column (Synergi Hydro 4 μm 10 × 150 mm) at a flow rate of 3 ml/min. The fraction containing 5-[125I]-A-85380 was collected and the solvent removed by rotatory evaporation. Reconstitution of the purified 5-[125I]-A-85380 was carried out with sterile 0.9% saline solution through a 0.22 μm filter. The 5-[125I]-A-85380 obtained was then stored at −20°C. Analytical HPLC of an aliquot of the final product was carried out on a reverse phase C-18 analytical column (Synergi Hydro 4 μm, 4.6 × 150 mm) at a flow rate of 1 ml/min.
Stability of 5-[125I]-A-85380
The stability of 5-[125I]-A-85380 in saline was investigated over time at −20°C. Samples were stored and analyzed at 5½, 9, and 13 weeks. Samples were injected on to a reverse phase C-18 analytical column (Synergi Hydro 4 μm, 4.6 × 150 mm) at a flow rate of 1 ml/min.
Brain Tissue
Joint Ethics Committee of Newcastle and North Tyneside Health Authority rules and MRC brain banking procedures were followed and consent of next of kin was obtained in the collection of all brain tissue. Coronal slices from the left hemisphere were obtained postautopsy and were rapidly frozen in liquid arcton cooled over liquid nitrogen. The slices were stored at −70°C.
For the distribution study, tissue from 16 control cases was used. Demographic details of control cases are shown in Table 1. Smoking histories were obtained, one smoker was excluded from the study and ex-smokers identified. The control group studied had not been exposed to neuroleptic medication. Tissue blocks were obtained containing striatum, globus pallidus, insula and cingulate cortices as well as blocks of anterior thalamus, temporal cortex, occipital cortex, frontal cortex, and cerebellum. Not all areas studied were available in every case.
For the disease study, tissue from 16 controls cases, 24 DLB cases, 8 AD cases, 4 mild AD cases, 10 VaD cases, and 10 PD cases was used. Demographic details of cases are shown in Table 1. No significant differences in age were seen between groups. Smoking histories were obtained for all groups, one smoker was excluded from the study and ex-smokers identified. Neuroleptic drug histories were also identified. None of the cases used in this study were exposed to cholinesterase inhibitors. Tissue blocks were obtained containing striatum, globus pallidus, insula and cingulate cortices as well as blocks of anterior thalamus and temporal cortex. The majority of areas were obtained from most cases.
For autoradiography, sections were cut by cryostat (20 μm) and dried on to Vecta-bond-coated slides at room temperature before storage in sealed boxes at −70°C.
Neuropathological Diagnosis
At post-mortem, the right hemisphere was formalin fixed for at least 3 months. AD pathology was evaluated using quantitative techniques to determine plaque and tangle densities (Perry et al, 1990b). Sections of cortical areas were stained with a modification of Palmgren's silver technique for neurofibrillary tangle quantification (Cross, 1982). The von Braunmühl silver impregnation technique was used to demonstrate plaques. For each neocortical area, the mean tangle density was obtained in consecutive fields (0.61 mm2/area) through the full width of the cortical ribbon in five randomly marked positions around gyri; the mean plaque density was calculated from counts in fields (3.1 mm2/area) at five similarly marked points. Tangle and plaque densities in the whole of the neocortex were expressed as mean values per mm2. Braak staging was determined (Braak and Braak, 1991) and AD was defined as Braak stage V and VI. Mild AD was defined as cases with no clinical confirmation of dementia but demonstrating senile plaque densities diagnostic for possible AD and neurofibrillary changes characteristic of early AD (Braak stage ⩽III). α-Synuclein immunohistochemistry (Novocastra Laboratories Ltd, Newcastle, UK) was used to demonstrate Lewy bodies (LB) and Lewy neurites. Cortical LB frequency was assessed according to the consensus guidelines for diagnostic rating protocol of DLB (McKeith et al, 1996), which have been modified for α-synuclein, an LB pathology marker more sensitive than the earlier recommended ubiquitin. Scoring for each cortical area for α-synuclein was: 0=no LB, 1=up to five LB, 2=5–20 LB plus few neurites, 3=20–40 LB plus moderate neurites, 4=over 40 LB plus dense neurites. Controls had no evidence of neurological disease. Controls were assessed neuropathologically, and did not show AD pathology or LB commensurate with a diagnosis of AD or other neurodegenerative disease. No significant vascular pathology was seen in controls.
Autoradiography Procedures
Prior to assay, sections were left to air dry at room temperature. Adjacent sections were prewashed in Tris-HCl pH 7.4 buffer, containing 120 mM NaCl, 5 mM KCl, 2.5 mM CaCl2, and 1 mM MgCl2, for 20 min at room temperature to remove endogenous ligand. For total binding, sections were incubated in the same buffer as prewash and 22 pM 5-[125I]-A-85380 for 4 h at room temperature. The Kd of 5-[125I]-A-85380 has been reported to be 12 pM in the human brain (Mukhin et al, 2000); therefore, at the above concentration binding approaches Bmax. A measure of 1 μM epibatidine was added to at least one adjacent section from each case to give a measure of nonspecific binding. After incubation, sections were washed for 2 × 10 min in the same buffer at 4°C and dipped into distilled water at 4°C. Sections were then left to air dry for 1 h before exposure to Kodak Biomax MR film (24 × 30 cm) with 125I autoradiographic microscale standards at 4°C. After 48 h exposure, the films were warmed to room temperature (2 h) before development using 500 ml D19 for 5 min, stopped using 500 ml 1% aqueous acetic acid for 1 min, fixed using 500 ml 25% Unifix for 6 min, and washed for 20 min in running water. Films were left to air dry for 2 h. Binding was quantified to 125I standards using MCID M5+ image analysis system (Imaging Research Ltd, Interfocus, UK) to give binding in fmol/mg of tissue. Specific binding was calculated by subtraction of non-specific binding from mean total binding.
Statistical Analysis
Binding data in control and disease groups were analyzed by one-way ANOVA with post hoc analysis using Fisher's multiple comparison of means. Differences, between groups, in age and post-mortem delay were analyzed by one-way ANOVA with post hoc analysis using Fisher's multiple comparison of means, and correlations between binding and demographic factors were analyzed by Pearson's product moment correlation. Binding data in ex-smokers and non-smokers, and neuroleptic exposed and non-exposed, were analyzed by one-way ANOVA with post hoc analysis using Fisher's multiple comparison of means. Analysis was carried out using Minitab (Release 13, Minitab Inc.)
RESULTS
Radiochemistry
Modification of a previously reported method (Zoghbi et al, 2001) has allowed radiolabelling and purification of 5-125IA via a more rapid, one-pot synthesis method. 5-[125I]-A-85380 was successfully obtained with a radiochemical purity of >98% and an isolated radiochemical yield of 60.2%. The specific activity of the product was calculated to be 2580 Ci/mmol.
Stability of 5-[125I]-A-85380 in Saline
After 9 weeks, the radiochemical purity of 5-[125I]-A-85380 stored at −20°C was found to be 96.9%±3.0 (n=2). By 13 weeks, the radiochemical purity of 5-[125I]-A-85380 stored at −20°C was reduced to 61.1%±2.6 (n=4). All autoradiography assays were completed within 6 weeks after the synthesis of 5-[125I]-A-85380.
Effects of Demographic Variables, Neuroleptic and Smoking Histories
There were no significant correlations in 5-[125I]-A-85380 binding with age and post-mortem delay. In addition, no significant differences in 5-[125I]-A-85380 binding were found between ex-smokers and non-smokers in all areas studied in the control group in the distribution study and in all groups in the disease study. Binding in the striatum and insular cortex in ex- and non-smokers is shown in Table 2. Exposure to neuroleptic medication was also shown not to affect 5-[125I]-A-85380 binding, with no significant differences seen between exposed and non-exposed cases within disease groups (Table 3).
Autoradiography
Distribution in normal tissue
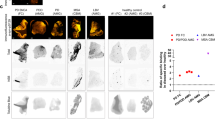
5-[125I]-A-85380 binding in normal human brain tissue was highest in the thalamic nuclei (Figure 1b and c), in particular the lateral geniculate (LG) nucleus and anteroprincipal (APr) thalamic nucleus (1.50±0.31 and 1.49±0.64 fmol/mg, respectively) where binding is 3- to 4-fold higher than in average cortical binding. Binding in the cerebellum (Figure 1a) was five-fold higher in the molecular layer (1.17±0.20 fmol/mg) than the white matter (0.20±0.06 fmol/mg), with moderate binding in the granular layer (0.48±0.06 fmol/mg). The highest cortical density was in the entorhinal cortex (0.82±0.55 fmol/mg) (Figure 1f). Moderate binding was also found in the visual area (Brodmann Area (Ba) 17) and secondary visual area (Ba18), in particular middle and lower layers (0.50±0.08 and 0.48±0.126 fmol/mg), in the occipital lobe (Figure 1e) and moderate binding was also found in the auditory association area (Ba21), Ba20, and Ba36 in the temporal cortex (0.44±0.11, 0.45±0.17, and 0.46±0.08 fmol/mg, respectively) (Figure 1f). The striatal binding was moderate with slightly higher binding in the caudate (0.57±0.17 fmol/mg) than putamen (0.56±0.19 fmol/mg), whereas binding in the globus pallidus (GP) was largely absent (Figure 1b). Also highlighted with moderate binding were white matter tracts in the occipital (Figure 1e), frontal (Figure 1c), and temporal lobes (Figure 1f) where binding was generally 3- to 5-fold greater than white matter (0.26±0.06 and 0.48±0.13 fmol/mg). No measurable non-specific binding of 5-[125I]-A-85380 was apparent in any areas studied when binding was displaced by 1 μM epibatidine. An example of non-specific binding in a section of striatum, anterior thalamus, and adjacent tissue is illustrated in Figure 1d.

Autoradiographs of 5-[125I]-A-85380 binding in human tissue sections: (a) cerebellum, (b) striatal section showing caudate, putamen, anterior thalamus, subthalamic nucleus, and the edge of the substantia nigra, (c) thalamic section showing lateral geniculate and cingulate, entorhinal and insular cortices, (d) (c) when displaced with 1 μM epibatidine, (e) occipital lobe, (f) temporal lobe. (AD=anterodorsal, APr=anteroprincipal, CM=centromedian, GPe=external globus pallidus, GPi=internal globus pallidus, Iml=internal medullary lamina, L.=lateral, LD=lateral dorsal, MD=medial dorsal, N.=nucleus, PF=parafascicular, PV=paraventricular, sm=stria medullaris, SN=substantia nigra, STH=subthalamic nucleus, Thal.=thalamic, VA=ventroanterior, VIM=ventrointermedius, VLA=Ventrolateral anterior, VPM=Ventroposterior medial).
Disease comparisons
Striatal 5-[125I]-A-85380 binding in AD, DLB, and PD tended to be lower than both control and VaD cases (Table 4), reaching significance in the caudate (F(6,58)=6.8, p<0.05, <0.001, and <0.001, respectively) and in the putamen in only the DLB and PD groups (F(6,60)=6.94, p<0.001 and <0.001, respectively). In addition, 5-[125I]-A-85380 binding in the putamen was significantly lower in PD than in AD (F(6,60)=6.94, p<0.05). In the entorhinal cortex, significantly lower 5-[125I]-A-85380 binding was shown in AD, DLB, and PD compared to controls (F(5,11)=2.03, p<0.05),. The DLB group also showed significantly lower 5-[125I]-A-85380 binding than controls in the ventrolateral posterior thalamic (VLP) nucleus (F(6,18)=1.16, p<0.05) and the SN (F(6,38)=4.06, p<0.01). In the SN, 5-[125I]-A-85380 binding was also significantly reduced in DLB compared to AD and VaD cases (F(6,38)=4.06, p<0.05 and <0.001, respectively). In the SN, significantly lower 5-[125I]-A-85380 binding was observed in PD compared with control, AD. and VaD cases (F(6,38)=4.06, p<0.0, <0.01, and p<0.001, respectively). The PD group also was shown to have significantly reduced 5-[125I]-A-85380 binding than controls in the subthalamic (STH) nucleus (F(3,35)=1.58, p<0.05), the reticular nucleus (F(6,62)=1.02, p<0.05), and the APr thalamic nucleus (F(6,29)=1.67, p<0.05). 5-[125I]-A-85380 binding in the STH nucleus and insular cortex in PD was shown to be significantly lower than in DLB (F(3,35)=1.58, p<0.05 and F(6,60)=2.16, p<0.01, respectively). No significant difference in 5-[125I]-A-85380 binding was seen between the AD and mild AD groups in all areas studied.
DISCUSSION
Effect of Drug and Smoking Histories on 5-[125I]-A-85380 Binding
The density of 5-[125I]-A-85380 binding in ex-smokers was similar to that in non-smokers in all areas and groups investigated, in agreement with the reported similar [3H]nicotine binding in human brain in non- and ex-smokers (Court et al, 1998). In addition, there were no significant differences in 5-[125I]-A-85380 binding between patients exposed and not exposed to neuroleptic medication in all groups and areas studied. Previous post-mortem studies have shown neuroleptic medication to be associated with accentuation of [3H]nicotine binding deficits in the caudate nucleus (Court et al, 2000b). Results from this study suggest that neuroleptic medication may affect nAChRs other than the α4β2 subtype.
Distribution in Normal Tissue
In vitro visualization of 5-[125I]-A-85380 receptor distribution has previously been carried out in rats (Mukhin et al, 2000) and mice (Musachio et al, 1998; Horti et al, 1999). In the present study, the binding distribution of 5-[125I]-A-85380 in human post-mortem tissue was found to be consistent with [3H]cytisine (Pabreza et al, 1991), [3H]epibatidine (Marutle et al, 1998), and [3H]nicotine (Spurden et al, 1997; Court et al, 2000a) binding, with highest levels appearing in the thalamus, in particular the LG nucleus and APr nucleus, and the entorhinal cortex.
5-[125I]-A-85380 binding in the caudate and putamen was of intermediate density, similar to reported [3H]epibatidine (Marutle et al, 1998) and [3H]nicotine (Court et al, 1998, 2000a) binding in human brain tissue. A recent study by Kulak et al (2002) has shown 5-[125I]-A-85380 binding to α-conotoxin MII-sensitive nAChRs in the rodent and monkey striatum. These α-conotoxin MII-sensitive sites contain the α6 and β2 subunits (Grady et al, 2001; Champtiaux et al, 2002) and therefore striatal binding evident in the present study is likely to reflect both α4β2 and α6β2 subtype distributions. [125I]-α-conotoxin MII binding in mouse brain has also been shown to be high in the superficial layers of the superior colliculus, nigrostriatal pathways, optic tract, olivary pretectal, mediolateral and dorsolateral geniculate nuclei (Whiteaker et al, 2000) and therefore 5-[125I]-A-85380 in these areas could also reflect both α4β2 and α6β2 subtype distributions. 5-[125I]-A-85380 binding in the GP is largely absent, which is consistent with low [3H]nicotine binding in human brain tissue (Court et al, 2000a).
In the cerebellum, 5-[125I]-A-85380 binding appears more concentrated in the molecular layer than the granular layer, confirming previously reported [3H]cytisine (Aubert et al, 1992a) and [3H]epibatidine distribution (Court et al, 2000a); however, [3H]nicotine binding shows higher binding in the granular than the molecular layer (Court and Perry, 1995a). Moderate 5-[125I]-A-85380 binding was also present in the dentate nucleus. Immunohistochemical localization of nicotinic receptors in the cerebellum has shown a number of receptor subunits, including both α4 and α6, to be present in molecular and granule cell layer (Graham et al, 2002) and therefore 5-[125I]-A-85380 binding in the cerebellum may not be solely due to α4 but also to α6 subunit containing receptors. Cerebellar dysfunction has been implicated in attentional deficits. Nicotinic receptor agonists enhance attentional function and in autism α4 subunit abnormalities have been highlighted in the cerebellum (Lee et al, 2002) and therefore 5-I-A-85380 imaging may be useful in identifying abnormalities in this area in vivo.
In the occipital cortex, 5-[125I]-A-85380 binding appears to be concentrated in the visual cortex (Ba17) and the secondary visual area (Ba18), with highest levels in lower cortical layers, consistent with the specific action of acetylcholine in vision (Nobili and Sannita, 1997). Acetylcholine is thought to serve both sensory and cognitive processes involved in vision, and 5-[125I]-A-85380 binding in the LG nucleus may reflect the ‘gating’ of sensory information to the visual cortex.
Binding of 5-[125I]-A-85380 in white matter in the temporal, occipital, and cingulate cortices appears to course with cholinergic pathway previously observed using acetylcholinesterase histochemistry (Selden et al, 1998). Binding of 5-[125I]-A-85380 shown in the temporal cortex follows the course of the capsular division of the lateral cholinergic pathway. Fibers of this pathway run adjacent to the putamen in the medial aspect of the external capsule into the nucleus basalis of Meynert which then project down into the temporal cortex (Selden et al, 1998). Binding of 5-[125I]-A-85380 to fibers in the external capsule was not clearly seen due to moderate binding seen in the putamen and claustrum; however, binding to fibers running from the nucleus basalis of Meynert into the temporal cortex was clearly highlighted. The binding of 5-[125I]-A-85380 in the occipital lobe white matter correlates with lateral and medial cholinergic pathways merging posteriorly in the occipital lobe (Selden et al, 1998). 5-[125I]-A-85380 binding at the tip of the cingulate gyrus correlates with the medial cholinergic pathway supplying the cingulate gyrus (Selden et al, 1998). Streaks of intense [3H]nicotine white matter binding previously seen in foetal cerebellum and temporal lobe may also reflect the presence of nAChRs along cholinergic axons (Court et al, 1995b, 1997), probably visible in fetal tissue because quenching of [3H]nicotine is reduced due to lower myelination. It is reasonable to conclude that neuronal α4β2 (or possibly α6β2) receptors are located along cholinergic nerve fiber tracts possibly indicative of preassembled receptors being transported from the cell body along axons to terminals. A cholinergic fiber loss in cortical areas has previously been reported in AD brains compared to controls (Geula and Mesulam, 1996) and therefore 5-I-A-85380 may be useful in highlighting these deficits in vivo in humans. Further investigation into cortical areas in disease post-mortem tissue may show 5-I-A-85380 to be useful diagnostically, highlighting deficits in cholinergic pathways.
Disease Comparisons
Alzheimer's disease
Reports on the density of nAChRs in the striatum in AD are inconsistent with studies reporting both loss and preservation of agonist binding in the caudate and putamen (Shimohama et al, 1986; Perry et al, 1989; Rinne et al, 1991; Aubert et al, 1992a). In this study, posterior striatal levels were investigated and deficits in 5-[125I]-A-85380 binding in the caudate differentiated the AD group from controls. Deficits in striatal nAChRs do not appear, at least in some patients, to be associated with nigrostriatal dopaminergic deficits (Piggott et al, 1999) and are therefore more likely to be related to reduced striatal afferents from other sources, such as cortical areas subject to atrophy in AD and possibly thalamic areas. It may be that these striatal nAChRs deficits are responsible for disorders of movement observed in late stages of AD (Court et al, 2000a). Significantly reduced 5-[125I]-A-85380 binding in AD compared to controls was also found in the entorhinal cortex, consistent with reports of deficits in [3H]nicotine binding (Court et al, 2001) and [3H]epibatidine binding and α4 subunit protein expression in the neocortex in AD (Martin-Ruiz et al, 1999; Guan et al, 2000). A trend of lower 5-[125I]-A-85380 in thalamic nuclei was seen in AD; however, this did not reach statistical significance confirming earlier reports of no major deficits in thalamic areas in AD (Xuereb et al, 1990; Court et al, 2001).
Mild alzheimer's disease
There were no significant differences in 5-[125I]-A-85380 binding in mild AD compared to controls and also AD patients. This lack of change in binding in mild AD cases suggests that 5-[125I]-A-85380 may not be useful for early differential diagnosis. It must be noted, however, that only four mild AD cases were studied and investigation into areas where nAChRs are reported to be most markedly reduced in AD, such as temporal cortex, hippocampus, and entorhinal cortex, may still show 5-[125I]-A-85380 to be useful in this respect. In vivo 5-[125I]-A-85380 SPECT imaging, however, could potentially be useful in the assessment of effects of anticholinesterase therapy.
Lewy body dementia
Reduced 5-[125I]-A-85380 binding in both the caudate and putamen differentiated DLB from controls corroborating evidence of similar reductions in [3H]nicotine binding (Court et al, 2000b). In DLB, deficits in striatal M1 muscarinic acetylcholine receptors (mAChRs), measured using [3H]pirenzepine binding, have also been demonstrated (Piggott et al, 2003), highlighting a postsynaptic deficit, whereas 5-[125I]-A-85380 binding is likely to indicate a presynaptic deficit. In the SN, a region composed of dopaminergic nuclei projecting to the striatum, reduced 5-[125I]-A-85380 binding differentiated DLB from controls and also AD cases. The relative severity of reduced 5-[125I]-A-85380 binding seen in both the striatum and substantia nigra in DLB, comparable to that in PD, could potentially be a marker of early degeneration, possibly highlighting deficits in nigrostriatal inputs, as SN dopaminergic neuron loss is greater in PD than in DLB (Piggott et al, 1999), or deficits in other cortical inputs.
A reduction in 5-[125I]-A-85380 binding in the entorhinal cortex in DLB cases compared to controls confirms evidence of reductions in [3H]nicotine and [3H]epibatidine binding in this brain area in DLB (Perry et al, 1995; Martin-Ruiz et al, 2000). The relative severity of the reduction of [3H]epibatidine binding in the temporal cortex has been reported to be related to disturbances in consciousness (DOC) seen in some DLB patients, where patients with DOC show higher binding in the temporal cortex, including the entorhinal cortex, than patients showing no DOC (Ballard et al, 2002). Imaging with 5-[123I]-A-85380 could potentially highlight these differences in vivo, and clinical correlative studies in DLB are in progress.
Parkinson's disease
Deficits in 5-[125I]-A-85380 binding in both the caudate and the putamen differentiated PD from controls, as in DLB; however, lower 5-[125I]-A-85380 binding in the putamen also differentiated PD from AD cases. This result in PD is not unexpected due to the presence of presynaptic nAChRs on striatal dopaminergic terminals (Wonnacott, 1997). The loss of striatal 5-[125I]-A-85380 binding of 46–50% closely parallels previously reported [3H]nicotine reductions (Court et al, 2000b) and also the loss of nigrostriatal dopaminergic markers (Piggott et al, 1999). A study with 5-[125I]-A-85380 and [125I]epibatidine in control human and PD brains showed a similar decrease of 40–50% in binding of both radioligands in the striatum (Quik et al, 2002).
In the SN, reduced 5-[125I]-A-85380 binding also differentiated PD from controls and AD cases, highlighting the loss of dopaminergic nuclei projecting to the striatum. This finding is in agreement with evidence of reduced [3H]nicotine binding in the SN pars compacta and ventral tegmental area (Perry et al, 1995).
Reduced 5-[125I]-A-85380 binding in the entorhinal cortex also differentiates PD from controls, supporting a number of studies demonstrating a deficit in nAChRs with high-affinity agonists and ChAT activity in the neocortex in PD (Whitehouse et al, 1988; Rinne et al, 1991; Aubert et al, 1992b; Perry et al, 1993; Lange et al, 1993).
Thalamic 5-[125I]-A-85380 binding in the PD group tended to be lower than controls, with significance reached in the APr thalamic nucleus, the reticular nucleus, and also STH nucleus. Thalamic 5-[125I]-A-85380 binding in PD also tended to be lower than in DLB, with significance reached in the STH nucleus. Dopamine deficiency in PD causes the disinhibition and overactivity of the STH nucleus and its projections to the internal GP, which in turn overinhibits thalamocortical projections which is thought to be responsible for the appearance of symptoms such as akinesia, rigidity, and perhaps tremor (Henderson and Dunnett, 1998). A deficit in 5-[125I]-A-85380 binding in the STH nucleus in PD emphasizes the potential involvement of acetylcholine in the expression of these symptoms. Reduced 5-[125I]-A-85380 binding in the insular cortex also differentiated the PD group from the DLB group.
Vascular dementia: Although 5-[125I]-A-85380 binding in thalamic areas tended to be lower in a number of thalamic nuclei, no significant reductions were found for VaD in comparison with controls in any brain area investigated, supporting evidence of no reductions in either [3H]epibatidine binding or α4 nAChR subunit expression in the temporal cortex in VaD (Martin-Ruiz et al, 2000). Reduced 5-[125I]-A-85380 binding in striatal areas in AD, DLB, and PD and in the substantia nigra in DLB and PD differentiates these groups from VaD as well as controls. This could be valuable in differentiating VaD from primary degenerative dementia. It must be noted, however, that these differences were not seen between mild AD and VaD.
In summary, 5-[125I]-A-85380 binding reflects the previously reported distribution of α4β2 receptors in human brain and also differences of nAChRs seen with high-affinity agonists in age-related degenerative diseases. Significant differences were not only observed between dementia cases and controls but also between cases with different causes of dementia. Therefore, 5-[125I]-A-85380 is potentially a useful in vivo tool to aid specific diagnosis that is important in terms of treatment and management strategies.
References
Aubert I, Araujo DM, Cecyre D, Robitaille Y, Gauthier S, Quirion R (1992a). Comparative alterations of nicotinic and muscarinic binding sites in Alzheimer's and Parkinson's diseases. J Neurochem 58: 529–541.
Aubert I, Cecyre D, Gauthier S, Quirion R (1992b). Autoradiographic distribution of nicotinic receptor sites labeled with [3H]cytisine in the human brain. In: Clarke PBS, Quik M, Adlkofer F, Thurau K (eds). Effects on Biological Systems II.. Birkhauser Verlag: Basal. pp 363–369.
Ballard CG, Court JA, Piggott M, Johnson M, O'Brien J, McKeith I et al (2002). Disturbances of consciousness in dementia with Lewy bodies associated with alteration in nicotinic receptor binding in the temporal cortex. Conscious Cogn 11: 461–474.
Braak H, Braak E (1991). Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol (Berl) 82: 239–259.
Burghaus L, Schutz U, Krempel U, de Vos RA, Jansen Steur EN, Wevers A et al (2000). Quantitative assessment of nicotinic acetylcholine receptor proteins in the cerebral cortex of Alzheimer patients. Brain Res Mol Brain Res 76: 385–388.
Champtiaux N, Han ZY, Bessis A, Rossi FM, Zoli M, Marubio L et al (2002). Distribution and pharmacology of alpha 6-containing nicotinic acetylcholine receptors analyzed with mutant mice. J Neurosci 22: 1208–1217.
Court J, Martin-Ruiz C, Piggott M, Spurden D, Griffiths M, Perry E (2001). Nicotinic receptor abnormalities in Alzheimer's disease. Biol Psychiatry 49: 175–184.
Court JA, Lloyd S, Johnson M, Griffiths M, Birdsall NJ, Piggott MA et al (1997). Nicotinic and muscarinic cholinergic receptor binding in the human hippocampal formation during development and aging. Brain Res Dev Brain Res 101: 93–105.
Court JA, Lloyd S, Thomas N, Piggott MA, Marshall EF, Morris CM et al (1998). Dopamine and nicotinic receptor binding and the levels of dopamine and homovanillic acid in human brain related to tobacco use. Neuroscience 87: 63–78.
Court JA, Martin-Ruiz C, Graham A, Perry E (2000a). Nicotinic receptors in human brain: topography and pathology. J Chem Neuroanat 20: 281–298.
Court JA, Perry EK (1995a). Distribution of nicotinic receptors in the CNS. In: Stone TW (ed). CNS Neurotransmitters and Neuromodulators. CRC Press: London. pp 85–104.
Court JA, Perry EK, Spurden D, Griffiths M, Kerwin JM, Morris CM et al (1995b). The role of the cholinergic system in the development of the human cerebellum. Brain Res Dev Brain Res 90: 159–167.
Court JA, Piggott MA, Lloyd S, Cookson N, Ballard CG, McKeith IG et al (2000b). Nicotine binding in human striatum: elevation in schizophrenia and reductions in dementia with Lewy bodies, Parkinson's disease and Alzheimer's disease and in relation to neuroleptic medication. Neuroscience 98: 79–87.
Cross RB (1982). Demonstration of neurofibrillary tangles in paraffin sections: a quick and simple method using a modification of Palmgren's method. Med Lab Sci 39: 67–69.
Flores CM, Rogers SW, Pabreza LA, Wolfe BB, Kellar KJ (1992). A subtype of nicotinic cholinergic receptor in rat brain is composed of alpha 4 and beta 2 subunits and is up-regulated by chronic nicotine treatment. Mol Pharmacol 41: 31–37.
Geula C, Mesulam MM (1996). Systematic regional variations in the loss of cortical cholinergic fibers in Alzheimer's disease. Cereb Cortex 6: 165–177.
Grady SR, Meinerz NM, Cao J, Reynolds AM, Picciotto MR, Changeux JP et al (2001). Nicotinic agonists stimulate acetylcholine release from mouse interpeduncular nucleus: a function mediated by a different nAChR than dopamine release from striatum. J Neurochem 76: 258–268.
Graham A, Court JA, Martin-Ruiz CM, Jaros E, Perry R, Volsen SG et al (2002). Immunohistochemical localisation of nicotinic acetylcholine receptor subunits in human cerebellum. Neuroscience 113: 493–507.
Guan ZZ, Zhang X, Ravid R, Nordberg A (2000). Decreased protein levels of nicotinic receptor subunits in the hippocampus and temporal cortex of patients with Alzheimer's disease. J Neurochem 74: 237–243.
Henderson JM, Dunnett SB (1998). Targeting the subthalamic nucleus in the treatment of Parkinson's disease. Brain Res Bull 46: 467–474.
Holladay MW, Dart MJ, Lynch JK (1997). Neuronal nicotinic acetylcholine receptors as targets for drug discovery. J Med Chem 40: 4169–4194.
Horti AG, Koren AO, Lee KS, Mukhin AG, Vaupel DB, Kimes AS et al (1999). Radiosynthesis and preliminary evaluation of 5-[I-123/125]iodo3-(2(S)-azetidinylmethoxy)pyridine: a radioligand for nicotinic acetylcholine receptors. Nucl Med Biol 26: 175–182.
Kulak JM, Sum J, Musachio JL, McIntosh JM, Quik M (2002). 5-Iodo-A-85380 binds to alpha-conotoxin MII-sensitive nicotinic acetylcholine receptors (nAChRs) as well as alpha 4 beta 2*subtypes. J Neurochem 81: 403–406.
Lange KW, Wells FR, Jenner P, Marsden CD (1993). Altered muscarinic and nicotinic receptor densities in cortical and subcortical brain regions in Parkinson's disease. J Neurochem 60: 197–203.
Lee M, Martin-Ruiz C, Graham A, Court J, Jaros E, Perry R et al (2002). Nicotinic receptor abnormalities in the cerebellar cortex in autism. Brain 125: 1483–1495.
Lindstrom J, Anand R, Peng X, Gerzanich V, Wang F, Li Y (1995). Neuronal nicotinic receptor subtypes. Ann NY Acad Sci 757: 100–116.
Martin-Ruiz C, Court J, Lee M, Piggott M, Johnson M, Ballard C et al (2000). Nicotinic receptors in dementia of Alzheimer, Lewy body and vascular types. Acta Neurol Scand Suppl 176: 34–41.
Martin-Ruiz CM, Court J, Molnar E, Lee M, Gotti C, Mamalaki A et al (1999). Alpha 4 but not alpha 3 and alpha 7 nicotinic acetylcholine receptor subunits are lost from the temporal cortex in Alzheimer' s disease. J Neurochem 73: 1635–1640.
Marubio LM, Mar Arroyo-Jimenez M, Cordero-Erausquin M, Lena C, Le Novere N, de Kerchove dA et al (1999). Reduced antinociception in mice lacking neuronal nicotinic receptor subunits. Nature 398: 805–810.
Marutle A, Warpman U, Bogdanovic N, Nordberg A (1998). Regional distribution of subtypes of nicotinic receptors in human brain and effect of aging studied by (+/−)-[H-3]epibatidine. Brain Res 801: 143–149.
McKeith IG, Galasko D, Kosaka K, Perry EK, Dickson DW, Hansen LA et al (1996). Consensus guidelines for the clinical and pathologic diagnosis of dementia with Lewy bodies (DLB): report of the consortium on DLB international workshop. Neurology 47: 1113–1124.
Mukhin AG, Gundisch D, Horti AG, Koren AO, Tamagnan G, Kimes AS et al (2000). 5-Iodo-A-85380, an alpha 4 beta 2 subtype-selective ligand for nicotinic acetylcholine receptors. Mol Pharmacol 57: 642–649.
Musachio JL, Scheffel U, Finley PA, Zhan YG, Mochizuki T, Wagner HN et al (1998). 5-[I-125/123]iodo-3(2(S)-azetidinylmethoxy)pyridine, a radioiodinated analog of A-85380 for in vivo studies of central nicotinic acetylcholine receptors. Life Sci 62: L351–L357.
Nobili L, Sannita WG (1997). Cholinergic modulation, visual function and Alzheimer's dementia. Vis Res 37: 3559–3571.
Pabreza LA, Dhawan S, Kellar KJ (1991). [3H]cytisine binding to nicotinic cholinergic receptors in brain. Mol Pharmacol 39: 9–12.
Paterson D, Nordberg A (2000). Neuronal nicotinic receptors in the human brain. Prog Neurobiol 61: 75–111.
Perry EK, Irving D, Kerwin JM, McKeith IG, Thompson P, Collerton D et al (1993). Cholinergic transmitter and neurotrophic activities in Lewy body dementia: similarity to Parkinson's and distinction from Alzheimer disease. Alzheimer Dis Assoc Disord 7: 69–79.
Perry EK, Morris CM, Court JA, Cheng A, Fairbairn AF, McKeith IG et al (1995). Alteration in nicotine binding sites in Parkinson's disease, Lewy body dementia and Alzheimer's disease: possible index of early neuropathology. Neuroscience 64: 385–395.
Perry EK, Smith CJ, Court JA, Perry RH (1990a). Cholinergic nicotinic and muscarinic receptors in dementia of Alzheimer, Parkinson and Lewy body types. J Neural Transm Park Dis Dement Sect 2: 149–158.
Perry EK, Smith CJ, Perry RH, Johnson M, Fairbairn AF (1989). Nicotine [3H]-nicotine receptor binding in human brain: characterization and involvement in cholinergic neuropathlogy. Neurosci Res Commun 5: 117–124.
Perry RH, Irving D, Blessed G, Fairbairn A, Perry EK (1990b). Senile dementia of Lewy body type. A clinically and neuropathologically distinct form of Lewy body dementia in the elderly. J Neurol Sci 95: 119–139.
Picciotto MR, Zoli M, Lena C, Bessis A, Lallemand Y, LeNovere N et al (1995). Abnormal avoidance learning in mice lacking functional high-affinity nicotine receptor in the brain. Nature 374: 65–67.
Piggott MA, Marshall EF, Thomas N, Lloyd S, Court JA, Jaros E et al (1999). Striatal dopaminergic markers in dementia with Lewy bodies, Alzheimer's and Parkinson's diseases: rostrocaudal distribution. Brain 122(Part 8): 1449–1468.
Piggott MA, Owens J, O'Brien J, Colloby S, Fenwick J, Wyper D et al (2003). Muscarinic receptors in basal ganglia in dementia with Lewy bodies, Parkinson's disease and Alzheimer's disease. J Chem Neuroanat 25: 161–173.
Quik M, Okihara M, Forno L, Musachio JL (2002). The in vivo imaging ligand iodo-A-85380 and radiolabeled epibatidine bind to similar nicotinic receptor sites in human control and Parkinson disease brains in vitro, programme no. 887.12. 2002 Abstract Viewer/Itinerary Planner. Society for Neuroscience: Washington, DC. Online.
Rinne JO, Myllykyla T, Lonnberg P, Marjamaki P (1991). A postmortem study of brain nicotinic receptors in Parkinson's and Alzheimer's disease. Brain Res 547: 167–170.
Selden NR, Gitelman DR, Salamon-Murayama N, Parrish TB, Mesulam MM (1998). Trajectories of cholinergic pathways within the cerebral hemispheres of the human brain. Brain 121: 2249–2257.
Shimohama S, Taniguchi T, Fujiwara M, Kameyama M (1986). Changes in nicotinic and muscarinic cholinergic receptors in Alzheimer-type dementia. J Neurochem 46: 288–293.
Sihver W, Gillberg PG, Nordberg A (1998). Laminar distribution of nicotinic receptor subtypes in human cerebral cortex as determined by [H-3](−)nicotine, [H-3]cytisine and [H-3]epibatidine in vitro autoradiography. Neuroscience 85: 1121–1133.
Spurden DP, Court JA, Lloyd S, Oakley A, Perry R, Pearson C et al (1997). Nicotinic receptor distribution in the human thalamus: autoradiographical localization of [3H]nicotine and [125I] alpha-bungarotoxin binding. J Chem Neuroanat 13: 105–113.
Warpman U, Nordberg A (1995). Epibatidine and ABT 418 reveal selective losses of alpha 4 beta 2 nicotinic receptors in Alzheimer brains. Neuroreport 6: 2419–2423.
Wevers A, Monteggia L, Nowacki S, Bloch W, Schutz U, Lindstrom J et al (1999). Expression of nicotinic acetylcholine receptor subunits in the cerebral cortex in Alzheimer's disease: histotopographical correlation with amyloid plaques and hyperphosphorylated-tau protein. Eur J Neurosci 11: 2551–2565.
Whiteaker P, McIntosh JM, Luo S, Collins AC, Marks MJ (2000). 125I-alpha-conotoxin MII identifies a novel nicotinic acetylcholine receptor population in mouse brain. Mol Pharmacol 57: 913–925.
Whitehouse PJ, Martino AM, Wagster MV, Price DL, Mayeux R, Atack JR et al (1988). Reductions in [3H]nicotinic acetylcholine binding in Alzheimer's disease and Parkinson's disease: an autoradiographic study. Neurology 38: 720–723.
Wonnacott S (1997). Presynaptic nicotinic ACh receptors. Trends Neurosci 20: 92–98.
Xuereb JH, Perry EK, Candy JM, Bonham JR, Perry RH, Marshall E (1990). Parameters of cholinergic neurotransmission in the thalamus in Parkinson's disease and Alzheimer's disease. J Neurol Sci 99: 185–197.
Zoghbi SS, Tamagnan G, Baldwin MF, Al Tikriti MS, Amici L, Seibyl JP et al (2001). Measurement of plasma metabolites of (S)-5-[123I]iodo-3-(2azetidinylmethoxy)pyridine (5-IA-85380), a nicotinic acetylcholine receptor imaging agent, in nonhuman primates. Nucl Med Biol 28: 91–96.
Zoli M, Lena C, Picciotto MR, Changeux JP (1998). Identification of four classes of brain nicotinic receptors using beta2 mutant mice. J Neurosci 18: 4461–4472.
Acknowledgements
Sally Pimlott is in receipt of a studentship award from Amersham Health. Dr Deborah Dewar, Wellcome Surgical Institute, Glasgow, provided the use of an MCID image analysis system.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pimlott, S., Piggott, M., Owens, J. et al. Nicotinic Acetylcholine Receptor Distribution in Alzheimer's Disease, Dementia with Lewy Bodies, Parkinson's Disease, and Vascular Dementia: In Vitro Binding Study Using 5-[125I]-A-85380. Neuropsychopharmacol 29, 108–116 (2004). https://doi.org/10.1038/sj.npp.1300302
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1300302
Keywords
This article is cited by
-
The effects of different acetylcholinesterase inhibitors on EEG patterns in patients with Alzheimer’s disease: A systematic review
Neurological Sciences (2024)
-
Neurotransmitter and receptor systems in the subthalamic nucleus
Brain Structure and Function (2023)
-
Behavioral changes after nicotine challenge are associated with α7 nicotinic acetylcholine receptor-stimulated glutamate release in the rat dorsal striatum
Scientific Reports (2017)
-
Muscarinic receptor binding changes in postmortem Parkinson’s disease
Journal of Neural Transmission (2017)
-
Imaging α4β2 Nicotinic Acetylcholine Receptors (nAChRs) in Baboons with [18F]XTRA, a Radioligand with Improved Specific Binding in Extra-Thalamic Regions
Molecular Imaging and Biology (2017)
