


Abstract
Glutathione transferase ζ 1 (GSTZ1), also known as maleylacetoacetate isomerase, catalyzes the penultimate step of tyrosine catabolism and metabolizes several α-halocarboxylic acids, including dichloroacetic acid (DCA), an investigational drug used for lactic acidosis and, recently, solid tumors. Age-related differences have been suggested in DCA pharmacotoxicology, but no information is available on GSTZ1 ontogeny in humans. Here, we investigated the cytosolic GSTZ1 developmental expression pattern and the influence of haplotype on GSTZ1 activity with DCA by using human livers from donors between 10 weeks gestation and 74 years. GSTZ1 expression was very low in fetal livers (<2 pmol of GSTZ1/mg cytosol). The expression began to increase after birth in an age-dependent manner until age 7 years. GSTZ1 was then sustained at stable, yet variable, levels (median, 20.0 pmol/mg cytosol; range, 4.8–47.3 pmol/mg cytosol) until age 74 years. GSTZ1 activity with DCA was strongly associated with haplotype and expression level. Samples homozygous or heterozygous for GSTZ1A exhibited ∼3-fold higher DCA dechlorinating activity than samples carrying other alleles at a given level of expression. The correlations (r2) between activity and expression were 0.90 and 0.68, respectively, for GSTZ1A carriers (n = 11) and noncarriers (n = 61). GSTZ1 is expressed in mitochondria in addition to cytosol. The GSTZ1A allele exhibited similar effects in the mitochondrial fraction by conferring a higher activity with DCA. In summary, we report a neonatal onset and an age-related increase in GSTZ1 protein expression during human liver development. Haplotype influenced GSTZ1 activity with DCA but not protein expression.
Introduction
It has been recognized for some time that the safe and effective use of drugs in the pediatric population requires adjusting the dose to account for developmental differences in drug metabolism and elimination (Hines, 2008). Despite recent advances in our understanding of the ontogeny of human drug-metabolizing enzymes, there remain many gaps in our knowledge, especially of phase II enzymes.
Glutathione transferase ζ 1 (GSTZ1), also known as maleylacetoacetate isomerase (MAAI), is a member of the cytosolic GST gene family that encodes at least 17 functional proteins organized into seven families (Mannervik et al., 2005). Originally identified in the cytosol, GSTZ1 is also found in the mitochondria, where it is expressed to a lesser extent (Li et al., 2011b). GSTZ1 has no detectable activity with the prototypical xenobiotic GST substrate 1-chloro-2,4-dinitrobenzene but does recognize α-halocarboxylic acids, such as dichloroacetic and 2-chloropropionic acids, as substrates. GSTZ1 catalyzes the dehalogenation of dichloroacetate (DCA) to glyoxylate (Tong et al., 1998). As the penultimate enzyme of tyrosine/phenylalanine catabolism, GSTZ1 isomerizes maleylacetoacetate to fumarylacetoacetate (Fernández-Cañón et al., 1999). Studies with GSTZ1 knock-out mice showed that these mice survived unless challenged with a diet high in protein, tyrosine, or homogentisic acid (Fernández-Cañón et al., 2002).
GSTZ1-catalyzed DCA dechlorination is of particular interest because DCA exposure occurs both environmentally and therapeutically. Human populations are estimated to ingest 2 to 4 μg of DCA/kg body weight through consumption of drinking water, where DCA is considered an environmental contaminant produced during water chlorination (Mughal, 1992). Meanwhile, DCA has long been used as an investigational drug for the treatment of several metabolic disorders, including genetic mitochondrial diseases, and, recently, proliferative conditions, including cancer and pulmonary arterial hypertension (reviewed by Stacpoole, 2011). Patients treated with DCA have been reported to develop occasional asymptomatic hepatotoxicity and, more frequently, a reversible peripheral neuropathy. Although not yet elucidated, the mechanism of DCA neurotoxicity may be linked to its biotransformation. DCA is a mechanism-based inactivator of GSTZ1 (Anderson et al., 2002); consequently, it inhibits its own metabolism and exhibits markedly delayed plasma clearance after repeated doses. The physiological function of GSTZ1/MAAI in tyrosine catabolism is similarly reduced during DCA treatment, leading to the accumulation of several reactive tyrosine metabolites, such as maleylacetoacetate, maleylacetone, succinylacetoacetate, and succinylacetone, some of which may be responsible for DCA toxicity (Stacpoole, 2011).
Subject age recently emerged as an important determinant of DCA pharmacotoxicology. DCA-associated peripheral neuropathy occurred frequently in adult patients treated for the syndrome of mitochondrial myopathy, encephalomyopathy, lactic acidosis, and stroke-like episodes (Kaufmann et al., 2006) or for glioblastoma multiforme (Michelakis et al., 2010). In contrast, children with heterogeneous causes of congenital lactic acidosis showed no worsening of hepatotoxicity or neurotoxicity even after chronic DCA treatment (Stacpoole et al., 2006). Examination of DCA plasma kinetics revealed similar clearance and other indices between children (age, 2.2–7.1 years at entry; mean age ± S.D., 5.2 ± 1.8; n = 5) and older individuals (age, 14.0–33.9 at entry; mean age ± S.D., 23.6 ± 10.0; n = 4) after their initial doses, whereas a striking age-dependent decrease in DCA clearance was observed at the end of 6 months of treatment (Shroads et al., 2008).
To date, 17 nonsynonymous single nucleotide polymorphisms (SNPs) have been reported in the coding region of the human GSTZ1 gene (National Center for Biotechnology Information SNP database; accessed September 3, 2011). Of these, three SNPs at g.5696G>A (rs7975), g.5726G>A (rs7972), and g.6772C>T (rs1046428) have been shown to be functionally important, resulting in five major GSTZ1 haplotypes, i.e., Z1A, Z1B, Z1C, Z1D, and Z1F, with varying allelic frequencies (Table 1). Recently, the influence of GSTZ1 haplotype on DCA pharmacokinetics was examined in healthy adult subjects (Shroads et al., 2011). Individuals that carried at least one allele of Z1A, in general, showed faster clearance of DCA than individuals not carrying this allele after the first DCA dose. However, a more pronounced reduction in drug clearance was observed in GSTZ1A/1A and GSTZ1A/1D subjects after 5 days of DCA treatment compared with subjects who possessed other haplotypes. One person with very slow DCA clearance was found to possess a rare g.6822G>A, V99M mutation. In vitro studies using expressed recombinant human GSTZ1 revealed that the GSTZ1A (KRT) variant possessed different kinetic properties and rates of inactivation with DCA as substrate and inhibitor, respectively, compared with the GSTZ1C, 1D, and 1B variants (Tzeng et al., 2000).
Summary of the major GSTZ1 haplotypes and frequency of occurrence
Developmental changes and genetic variations in drug-metabolizing enzymes are recognized as major contributing factors to age-related and interindividual differences in both drug efficacy and drug toxicity. Several GST classes (GSTA, GSTM, and GSTP) exhibit distinct changes in hepatic expression during human development (Strange et al., 1989). In this study, we investigated the roles of subject age and GSTZ1 haplotype in determining the expression and activity of GSTZ1 with DCA in human liver samples.
Materials and Methods
Chemicals and Reagents.
The [1-14C]DCA (55 mCi/mmol, 99% purity) was purchased from American Radiolabeled Chemicals (St. Louis, MO). Unlabeled clinical-grade sodium DCA was purchased from TCI America (Portland, OR). To prepare a working solution of [1-14C]DCA, [1-14C]DCA was converted to the sodium salt by adding NaHCO3 and diluting with unlabeled sodium DCA to a concentration of 10 mCi/mmol. Rabbit polyclonal anti-human GSTZ1C antibody was produced using the intact recombinant human GSTZ1C as the antigen (Cocalico Biologicals, Reamstown, PA) (Guo et al., 2006). The antiserum was purified using the Pierce Protein A Antibody Purification kit (Thermo Fisher Scientific, Waltham, MA) before use in immunoblotting (Li et al., 2011b). All other chemicals were of American Chemical Society grade or higher and were purchased from commercial suppliers.
Human Liver Samples.
We studied 249 human liver samples from donors who succumbed at ages ranging from 10 weeks gestation to 74 years. Human liver cytosol from 61 prenatal and 167 postnatal samples was obtained from the Brain and Tissue Bank for Developmental Disorders, University of Maryland, Baltimore, and University of Miami and from the Laboratory for Developmental Biology, University of Washington. Most first-trimester and early second-trimester tissue samples were from pregnancy termination, whereas later samples were from preterm deliveries. In the former cases, the post-mortem interval (time between death and freezing of the tissue sample) was ≤1 h. For preterm deliveries, the median post-mortem interval was 9 h and ranged from 1 to 24 h. The cause of death was reported for most available postnatal tissue samples. Conditions that were likely to involve liver pathology were excluded from the tissue bank. The median post-mortem interval for postnatal samples was 18 h and ranged from 1 to 41 h. Donor characteristics, the preparation of subcellular fractions, and the isolation of DNA have been described previously (Koukouritaki et al., 2002; Stevens et al., 2008). The other 21 postnatal samples were deidentified human livers, of which 14 were obtained from Vanderbilt University (Nashville, TN) and 7 were collected during surgery at Shands Hospital at the University of Florida (Gainesville, FL). Liver cytosol and mitochondria fractions from these samples were prepared exactly as described by Li et al. (2011b). The cell nuclear pellet was used to isolate DNA for genotyping (QIAamp DNA Blood Mini kit; QIAGEN, Valencia, CA). Protein concentration was determined by the BCA Protein Assay (Thermo Fisher Scientific).
GSTZ1-Dependent DCA Dechlorination Activity.
GSTZ1-specific activity was measured with [1-14C]DCA as the substrate. Assay products were separated and detected using a high-performance liquid chromatography-coupled radioactivity flow detector as described previously (James et al., 1997). Human liver cytosol (0.2–0.6 mg) was incubated with 1 mM glutathione, 0.1 M HEPES-NaOH, pH 7.6, and a saturating concentration of [1-14C]DCA at 0.2 mM in an assay volume of 0.1 ml. The reaction was started by adding DCA, allowed to proceed for 15 min at 37°C with gentle shaking, and stopped by adding 0.1 ml of ice-cold methanol. All assays were performed in duplicate; substrate consumption did not exceed 15%: all reactions were linear under these conditions, and no inactivation by substrate was observed. GSTZ1-specific activity was expressed as nanomoles of glyoxylate formed per minute per milligram of protein. The limit of quantitation (LOQ) was 0.088 nmol. Samples with product formed below the LOQ were imputed a value equal to the LOQ divided by the square root of 2, i.e., 0.062 nmol (Armbruster and Pry, 2008). The samples with values below the LOQ are identified in Fig. 2.
Western Blot Analysis of GSTZ1 Expression.
Known amounts of protein (5–60 μg) were separated by SDS denaturing electrophoresis using 4 to 15% polyacrylamide gels (Ready Gel Tris-HCl Gel; Bio-Rad Laboratories, Hercules, CA) and subsequently transferred by electrophoresis onto polyvinylidene fluoride membranes (Millipore Corporation, Billerica, MA). The amount of cytosolic protein used was guided by the measured GSTZ1 activity (see above). After blocking with 5% nonfat dry milk, the membrane was incubated with rabbit polyclonal anti-hGSTZ1C-1C antibody diluted 1:5000 and with horseradish peroxidase-conjugated donkey anti-rabbit IgG antibody diluted 1:2000 (GE Healthcare, Chalfont St. Giles, UK). Protein signals were developed by ECL Plus Western Blotting detection reagents (Thermo Fisher Scientific) and visualized using Amersham Hyperfilm ECL (GE Healthcare). The chemiluminescent-derived signals were digitized by scanning, and the density of bands was analyzed by QuantiScan software (Biosoft, Cambridge, UK). A GSTZ1 standard curve was constructed using 0.023 to 0.733 pmol of purified recombinant human GSTZ1C (Guo et al., 2006) as shown in Supplemental Fig. 1. To quantify test samples, 5 μg of a designated human liver cytosol sample was used as a reference control on each blot, and the signal of each individual test sample was normalized against the reference control. The GSTZ1 content of individual samples was calculated by linear regression and expressed as picomoles of GSTZ1 per milligram of cytosolic or mitochondrial protein. Data were analyzed using Microsoft Office Excel 2007 (Microsoft, Redmond, WA). The LOQ was 0.023 pmol of GSTZ1. Samples with expression below the LOQ were imputed a value equal to the LOQ divided by the square root of 2, i.e., 0.016 pmol of GSTZ1.
Genotyping.
DNA from selected liver samples was subjected to polymerase chain reaction amplification followed by pyrosequencing (Langaee and Ronaghi, 2005), targeting three known GSTZ1 SNPs: rs7975 (g.5696G>A, E32K), rs7972 (g.5726G>A, G42R), and rs1046428 (g.6772C>T, T82M). Genotyping analysis was carried out using a PSQ HS 96 System (Biotage, Uppsala, Sweden). Haplotypes were inferred from the unphased data by computational methods (PHASE software, version 2.0.2) (Stephens et al., 2001). The Hardy-Weinberg equilibrium was calculated using a χ2 goodness-of-fit test with 1 degree of freedom.
Data Analysis.
For GSTZ1 expression and activity, age thresholds were determined by regression tree analysis using CART version 6 (Salford Systems, San Diego, CA). Least average deviation from the median was the function optimized to define the tree nodes; 80% of the sample was used to develop the tree, and 20% was used to test it. For GSTZ1 activity adjusted for liver weight, the thresholds of 2 months and 7 years were determined by visual inspection of the scatter plot analysis (data not shown). Outliers were defined as having specific content or activity that was 1.5-fold interquartile range away from the lower or upper quartile values. Statistical comparisons between age groups were performed using Kruskal-Wallis nonparametric tests, followed by stepwise step-down comparisons or all pairwised comparisons as specified in the figures (IBM SPSS Statistics 19; SPSS Inc., Chicago, IL). GSTZ1-adjusted activity was calculated by multiplying the activity (nanomoles of glyoxylate per minute per milligram of cytosol) with cytosolic protein yield (milligram of cytosol per gram of liver) and the ratio of liver to body weight (grams of liver per kilogram of body weight). The mean ± S.D. cytosolic protein yield was experimentally determined, using six adult livers, to be 71.0 ± 9.6 mg of cytosol/g liver. The average of the yield, 71.0 mg of cytosol/g liver, was used in calculations for all samples. A p value <0.05 was accepted as significant. Correlation analysis between GSTZ1 activity and expression included samples with specific contents above the LOQs and was estimated using Pearson's correlation (GraphPad Prism version 4.03; GraphPad Software Inc., San Diego, CA).
Results
GSTZ1 Haplotypes.
We genotyped 75 samples in the study. All genotype frequencies were in Hardy-Weinberg equilibrium. The frequencies of four common GSTZ1 haplotypes were as follows: wild-type haplotype GSTZ1C (EGT), 53%; Z1B (KGT), 20%; Z1D (EGM), 16%; Z1A (KRT), 10%. In addition, we identified one individual with the rare haplotype Z1F (KGM).
GSTZ1 Development in Human Liver Cytosol.
To quantitate GSTZ1 protein expression, we compared GSTZ1 immunoreactivity in individual liver cytosol samples with a standard curve constructed with purified recombinant human GSTZ1C protein (Supplemental Fig. 1). The activity of GSTZ1 was evaluated using the probe substrate DCA at a saturating concentration of 0.2 mM. Overall, GSTZ1 protein expression was low and exhibited no apparent changes during fetal development, but it increased with age after birth. Protein expression was paralleled by an increase in DCA dechlorination activity of the enzyme (Fig. 1). Sex and post-mortem interval did not influence the results.

GSTZ1 protein expression (A) and activity with DCA (B) as a function of age are shown. The breaks in the abscissa are set at birth and 1 year. The age of prenatal samples is shown as weeks of gestational age, whereas postnatal samples are shown as years. Samples from individuals that carried at least one allele of GSTZ1A (KRT) are shown as filled red triangles, and all the others are shown as open circles.
Age-dependent changes in GSTZ1 expression and activity were divided into five age brackets (Fig. 2, A and B), based on the regression tree analysis. Fetal livers exhibited undetectable to very low levels of cytosolic GSTZ1 expression (<2 pmol of GSTZ1/mg protein) and activity (<0.1 nmol of glyoxylate · min−1 · mg−1).

Box and whisker plots showing GSTZ1 protein expression (A) and activity with DCA (B) as a function of age, with nodes as indicated by regression tree analysis, are shown. The horizontal bar, box, and whiskers denote the median, interquartile range, and lowest to highest values of the group, respectively. Outliers (○) were defined as having specific contents 1.5-fold interquartile range away from the lower or upper quartiles. Each age group was statistically different (p < 0.05) from the other groups, except for GSTZ1 activity between the <0 group and the >0–2.5W group. Data were analyzed by the Kruskal-Wallis nonparametric test, followed by the stepwise step-down test. The number of samples having specific contents below the LOQs is shown below the abscissa (<LOQ N). W, week; Y, year.
Although depicted as a node because of the physiological importance of birth, the difference between fetal samples and those samples from donors in the immediate postnatal period (0–2.5 weeks) was very slight in either GSTZ1 activity or expression levels. Of the 34 immediate postnatal samples, 20 were born prematurely, 3 were full term, and others carried no information on gestational age. Inspection of the data did not suggest prematurity to be the cause of low GSTZ1 function in the immediate postnatal period. In contrast, a 3.5-fold increase in both median GSTZ1 activity and expression was observed between samples ≤2.5 weeks postnatal age and between samples greater than 2.5 weeks but ≤1.25 years. Median GSTZ1 activity and expression increased another 2-fold in samples greater than 1.25 years of age, but ≤7 years of age, and 1.5-fold in donor samples greater than 7 years of age. Because the samples were taken at different post-mortem intervals, we considered the possibility that post-mortem interval could influence the results. There was a poor correlation between post-mortem interval and either GSTZ1 expression levels (adjusted r2 = 0.0325) or GSTZ1 activity (adjusted r2 = 0.0342). As a further assessment of tissue quality, possible correlations were evaluated between GSTZ1 expression levels and previously determined expression levels of two other enzymes that exhibit substantial increases in expression in the perinatal period, i.e., SULT2A1 (Duanmu et al., 2006) and CYP2E1 (Johnsrud et al., 2003), and one enzyme that exhibits relatively constant levels of expression during development, i.e., CYP2C19 (Koukouritaki et al., 2004). In all instances, adjusted r2 values were less than 0.25. Visual inspection of the data also failed to reveal any tissue samples wherein low expression levels were consistently observed for multiple enzymes. Although realizing the limitations of conclusions drawn from such pathological specimens, the above analyses are consistent with the tissue samples used being of high quality.
Children are known to have a larger mass ratio of liver to body weight (Anderson and Meakin, 2002). The capacity of hepatic GSTZ1 activity per kilogram of body weight was calculated for each individual for which liver weight and body weight was available and reanalyzed by age (Fig. 3). The median adjusted activity of GSTZ1 in samples above 2 months to 18 years of age was 4- to 5-fold higher than that of samples ≤2 months postnatal age (p < 0.05). No statistical difference was shown between younger (>2 months to 7 years of age) and older (>7–18 years of age) children.

Box and whisker plot of GSTZ1-adjusted activity by age groups. The activity was adjusted to total hepatic GSTZ1 activity normalized to body weight as described under Materials and Methods. Samples with known liver and body weights were included in the analysis. The horizontal bar, box, and whisker denote the median, interquartile range, and lowest to highest values of the group, respectively. The outlier was more than 1.5-fold interquartile range higher than the upper quartile. The number of samples in each group is shown in parentheses. M, month; Y, year. Data were analyzed by the Kruskal-Wallis nonparametric test, followed by all pairwise comparisons. ***, p < 0.001; n.s., not significant.
Correlation Analysis of GSTZ1 Activity and Expression.
A correlation (r2) of 0.51 was obtained between GSTZ1 expression and activity with DCA when all postnatal samples with specific contents above the LOQs were analyzed. However, a few individuals clearly segregated from the main body by exhibiting 2- to 3-fold higher activity with DCA than others at similar levels of expression (Fig. 4). The ratio of activity/expression was used to select samples for genotyping. All samples that were greater than 1 S.D. from the mean of activity/expression were genotyped, as were 3 times that many samples with activity/expression ratios close to the mean. To avoid biases, genotyping further extended to all samples with available DNAs above age 3 years. In all, 72 postnatal samples were genotyped. Individuals with high activity relative to expression carried at least one GSTZ1A allele. Subsequent grouping of the genotyped individuals into GSTZ1A carriers or noncarriers improved the correlation of activity with expression to r2 = 0.90 and 0.68, respectively. Therefore, GSTZ1 activity with DCA is dependent on both the level of protein expression and the GSTZ1 haplotype.

Correlation analysis of GSTZ1 protein expression and activity with DCA. The r2 values were determined by Pearson correlation and were as follows: 0.51 for all samples (n = 181), 0.90 for GSTZ1A carriers (red triangles; n = 11; red regression line), 0.68 for GSTZ1A noncarriers (inverted green triangles; n = 61; green line), and 0.79 for the remaining not genotyped samples (open circle; n = 109; dashed black line). P values were <0.0001 for all three correlations. Postnatal samples with both activity and expression above the LOQs were included in the analysis. Sample selection criteria for GSTZ1 genotyping is described under Results.
Impact of GSTZ1A Variant.
Consistent with the correlation analysis, six of seven activity outliers in the >7- to 74-year bracket were GSTZ1A carriers and exhibited higher DCA dechlorinating activity (Fig. 2B). Differentiating all known GSTZ1A carriers in the scatter plots demonstrated a major impact of this allele on DCA-metabolizing activity, especially in samples from older age groups >7 years (Fig. 1B). The impact of this allele on young individuals (<7 years) is unclear based on the current study. No effect on protein expression was shown for this allele (Fig. 1A).
Influence of Haplotype on GSTZ1 Expression and Activity.
We further analyzed the role of each haplotype in determining GSTZ1 activity and expression. Individuals carrying at least one GSTZ1C (EGT) and/or GSTZ1B (KGT) alleles only were grouped together because of their lack of difference in activity or expression (analysis not shown). GSTZ1A (KRT) and GSTZ1D (EGM) carriers and GSTZ1A/Z1D heterozygotes were grouped separately based on a recent in vivo study indicating low activity for GSTZ1D and GSTZ1A carriers (Shroads et al., 2011) and our current findings of high in vitro activity associated with the GSTZ1A allele.
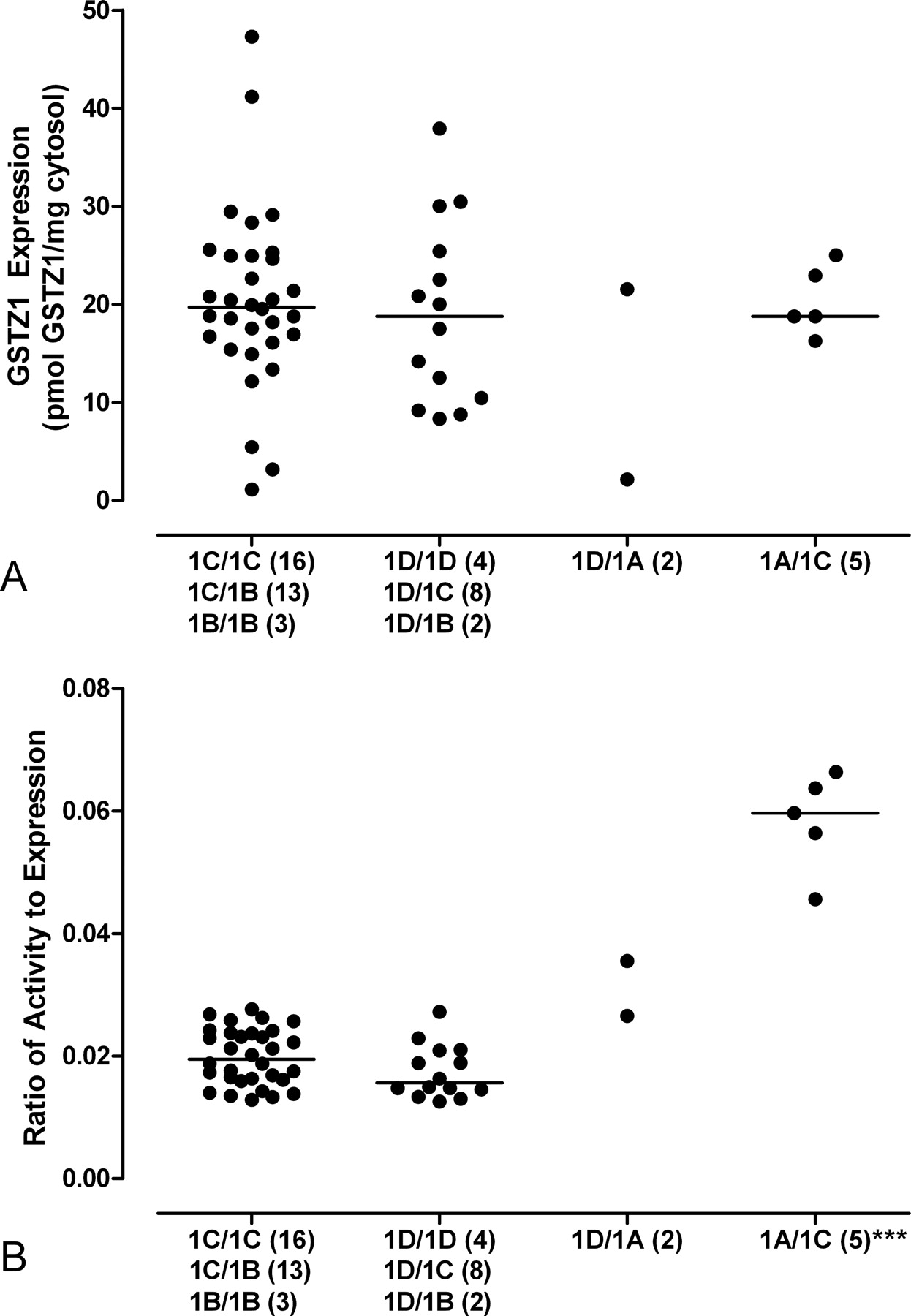
In liver cytosol of individuals 3 to 74 years of age, GSTZ1 haplotype had no effect on protein expression, as shown by the similar median values of different haplotype groups (Fig. 5A). On the other hand, the GSTZ1A allele when paired with the GSTZ1C allele clearly conferred a larger ratio of GSTZ1 activity to expression compared with other alleles, demonstrating higher DCA-metabolizing activity at a given level of protein expression (Fig. 5B). Two samples in our liver bank were homozygous for GSTZ1A; however, both were aged less than 1 month postnatal. These two GSTZ1A/1A homozygotes, like GSTZ1C/1A heterozygotes, also exhibited relatively large ratios of activity to expression (ratio, 2.9 and 3.8).

Influence of the GSTZ1 haplotype on protein expression (A) and the ratio of DCA-dechlorinating activity to protein expression (B) is shown. Samples of age 3 and above, with expression and activity above the LOQs, were included in the analysis. The activity/expression ratio of the 1A/1C group is significantly different from all the other groups (***, p < 0.001) as analyzed by one-way analysis of variance, followed by Bonferroni pairwise comparisons. The number of samples of each haplotype is given in parentheses. The horizontal bar denotes the median.
The two GSTZ1D/1A heterozygotes, although having similar ratios of activity to expression, exhibited distinct levels of expression: one was near the median values of other haplotypes, and the other was expressed at a very low level. In addition to the low-expression GSTZ1D/1A heterozygote, three individuals of GSTZ1B/1B, 1B/1C, and 1C/1C haplotype each possessed very low GSTZ1 content for their ages (5–11 years) (Fig. 5A). A rare haplotype, KRT/KGT, was identified in a 17-day-old child. GSTZ1 immunoreactivity for this individual was invisible on Western blot, and the enzymatic activity was identical to the blank; neither variable changed in response to assaying increased amounts of protein. Resequencing the GSTZ1 gene in these five low-expression individuals did not reveal the newly identified V99M SNP mutation.
Effect of Haplotype on Mitochondrial GSTZ1.
We recently reported the presence of GSTZ1 in liver mitochondria (Li et al., 2011b). Examination of GSTZ1 expression in cytosol and mitochondria showed a similar distribution of the enzyme between the two compartments, independent of the allelic differences (Table 2). As with cytosolic GSTZ1, mitochondrial GSTZ1 from GSTZ1A/1C heterozygotes exhibited ∼3-fold higher activity with DCA than GSTZ1A noncarriers at a given level of expression. Therefore, haplotype influenced mitochondrial and cytosolic GSTZ1 similarly for DCA-metabolizing activity but had no effect on protein expression or subcellular distribution of the enzyme.
GSTZ1 expression and activity with DCA in cytosol and mitochondria of individuals with various haplotypes
Within groups of GSTZ1A carriers or noncarriers, no statistical difference was observed between cytosol and mitochondria for the ratio of activity to expression.
Discussion
The safe and effective use of drugs in pregnancy and in pediatric populations will be aided by information on the activity and expression of drug-metabolizing enzymes in the fetus, infant, and child and the influence of genetic polymorphism. GSTZ1 is required for metabolism of DCA, which is used in pediatric and adult populations (Stacpoole, 2011), and the tyrosine catabolites maleylacetoacetate and maleylacetone (Cornett et al., 1999). An understanding of GSTZ1's ontogeny and other factors that can affect activity is important for selecting an appropriate dose of DCA or other substrates of this enzyme. GSTZ1 is known to be predominantly expressed in the liver (Lantum et al., 2002a). By studying human liver cytosols from 10 weeks of gestation to 74 years of life, we revealed age to be a major determinant of GSTZ1 protein expression and activity with DCA during human development. Similar to many other drug-metabolizing enzymes (e.g., GSTA, UGT1A1, CYP3A4, CYP1A2, and FMO3) studied to date (Hines, 2008), GSTZ1 was expressed minimally in the fetus and began to increase in the neonatal period. Neonatal onset of GSTZ1 expression was also observed at the mRNA level in mouse livers, although transcript levels reached adult values just 10 days after birth (Cui et al., 2010). Both differential transcription factor expression and epigenetic regulation have been suggested to contribute to the postnatal onset of drug-metabolizing enzymes. Indeed, postnatal enrichment of histone H3 lysine-4 dimethylation has been shown in the mouse Gstz1 gene (Cui et al., 2010).
The GSTZ1 haplotype frequencies in the current study are consistent with previous reports (Blackburn et al., 2001; Shroads et al., 2011). GSTZ1 haplotype had no effect on protein expression or subcellular distribution of the enzyme but had a major impact on enzymatic activity with DCA. Consistent with results obtained with the polymorphic variants of recombinant hGSTZ1 (Blackburn et al., 2001), the GSTZ1A allele, paired with or without the GSTZ1C allele, conferred higher DCA-metabolizing activity for a given level of expression in human liver samples. These findings were also consistent with the faster DCA clearance after an initial DCA dose observed in persons carrying the GSTZ1A allele, compared with those who lacked the allele (Shroads et al., 2011). However, the limited number of samples precluded determining whether the effect of the GSTZ1A allele was dominant over or additive to the effect of the GSTZ1C allele. For the same reason, it remains unknown whether GSTZ1A confers a similar effect on activity when paired with the GSTZ1D allele. Of note, although the GSTZ1A allele-associated high ratio of activity/expression was observed in individuals as early as 20 days postnatal, the high activity associated with this allele was not prominent in early life because of the low levels of protein expression in young GSTZ1A carriers. Because of the absence of GSTZ1A allele-bearing samples in our bank between the age period of 6 months and 7 years, we are unable to determine the age at which the high activity associated with bearers of the 1A allele becomes noticeable.
Unexplained by the current knowledge regarding GSTZ1 polymorphisms, several individuals in our liver bank exhibited very low GSTZ1 expression and activity for their ages. Potential contributing factors pertaining to liver quality were carefully examined and precluded. A novel V99M SNP (rs140540096) was recently identified in a KGM/KGT individual with an extraordinarily slow rate of DCA clearance (Shroads et al., 2011). However, none of the individuals with low GSTZ1 function in our study carried this mutation. L8P is another less studied SNP that was suggested to be very poorly expressed (Blackburn et al., 2001), but its occurrence has not yet been reported in any population. Regulatory polymorphism is also known to alter protein expression, but none of the SNPs in the GSTZ1 promoter region examined so far seemed to have a major effect on GSTZ1 expression (Fang et al., 2006). Therefore, it may be of interest to investigate genetic or environmental factors that may contribute to the low functionality of GSTZ1.
Human studies showed no differences in single-dose DCA pharmacokinetics between children younger than 7 years and adults (mean age, 24 years) (Shroads et al., 2008, 2011). The initial elimination half-life was 2.5 ± 0.4 h (mean ± S.D.) in children and 2.1 ± 1.5 h in adults, and other pharmacokinetic parameters were the same for both groups. Although the current study showed lower GSTZ1 activity and expression in young children less than 7 years, compared with those over age 7, the differences disappeared when the higher liver/body weight of children was considered. Thus, with adjustments for liver weight, the findings in this study are consistent with results of single-dose DCA pharmacokinetics.
This study was not designed to investigate GSTZ1 function after repeated DCA doses, which results in the inactivation of the enzyme. Therefore, it does not explain the differences in DCA clearance between children and adults upon chronic treatment (Shroads et al., 2008), or between individuals of GSTZ1A/Z1A and GSTZ1A/Z1D haplotypes relative to other haplotypes after repeated doses (Shroads et al., 2011). Our preliminary studies showed haplotype-dependent differences in DCA-induced inactivation of GSTZ1, an effect that was further modulated by chloride and other anions (Li et al., 2011a; W. Li and M. O. James, unpublished data). Therefore, we speculate that the differences in DCA pharmacokinetics after repeated exposure may be related to the rates of GSTZ1 inactivation by DCA and/or the turnover of the GSTZ1 enzyme.
Although factors influencing the inactivation of GSTZ1 are likely to play the most important role in determining biotransformation and elimination of DCA upon chronic dosing, not all substrates of GSTZ1 inactivate the enzyme. The endogenous substrate, maleylacetone, inhibits but does not inactivate GSTZ1 in the presence of physiological concentrations of GSH (Cornett et al., 1999; Lantum et al., 2002b). Xenobiotic substrates for GSTZ1 that form GSH conjugates, such as 2-chloropropionate, as well as those with α fluoro substituents, such as chlorofluoroacetic acid, do not inactivate GSTZ1 (Anderson et al., 1999). The developmental pattern of GSTZ1 shown in this study, considered in concert with the higher liver/body weight ratio of children, suggests that the human biotransformation of substrates that do not inactivate GSTZ1 will be low prenatally and in the immediate postnatal period but will be similar to adults in children over the age of 2 months.
GSTZ1/MAAI has the important physiological function of isomerizing maleylacetoacetate at the penultimate step of phenylalanine/tyrosine catabolism (Fernández-Cañón et al., 1999). In the fetal liver, this pathway is suggested to be rate limited by tyrosine aminotransferase, an enzyme that converts tyrosine to 4-hydroxylphenylpyruvate (Ohisalo et al., 1982). Like GSTZ1/MAAI, tyrosine aminotransferase possesses very low activity in the fetus and increases rapidly after birth (Delvalle and Greengard, 1977; Andersson et al., 1980; Ohisalo et al., 1982). This suggests that maleylacetoacetate exists at a very low concentration in fetal liver, so that slow nonenzymatic conversion to fumarylacetoacetate could be sufficient for its degradation (Fernández-Cañón et al., 2002). As with tyrosine aminotransferase, the rapid onset of GSTZ1/MAAI in neonates shown in this study may be regarded as a coordinated and adaptive response to the increased intake and consequent degradation of aromatic amino acids required for infant growth.
Developmental changes in overall GST activity have been studied using the common substrate 1-chloro-2,4-dinitrobenzene (Mulder et al., 1994; Raijmakers et al., 2001; Gallagher and Gardner, 2002). Both prenatal and postnatal livers possess good but variable activities for 1-chloro-2,4-dinitrobenzene, and a slight trend of increased activity was suggested with age. Detailed ontogenic information for specific GST members is available only from a study by Strange et al. (1989) on A, M, and P classes. The developmental pattern of Z1 is most similar to those of A1, A2, and M classes for a progressive increase with age, which is the reverse of that observed with the P class. The GSTZ1 enzyme is distinct from other classes in that it exhibits a near-baseline level in the fetus and a developmental onset beginning in the neonatal period. In adults, hepatic GSTZ1 content (median, 0.48 μg/mg cytosolic protein; range, 0.11–0.91; GSTZ1 molecular mass of 24 kD was used in conversion) is approximately 40-fold lower than that of GSTA1/A2, the most abundant cytosolic GSTs in postnatal livers with mean levels reported to be 16.5 μg/mg cytosolic protein by Strange et al. (1989) and 21.84 by Mulder et al. (1994).
In conclusion, we observed a neonatal onset and an age-related increase in GSTZ1 protein expression during human liver development. GSTZ1 activity with DCA is directly correlated to the level of protein expression and dependent on the GSTZ1 haplotype. The GSTZ1A (KRT) allele, paired with or without the GSTZ1C allele, confers ∼3-fold higher activity with DCA at a given level of protein expression than all other haplotypes that possess similar activities with DCA. The study provided in vitro explanations for the pharmacokinetics of a single dose of DCA and insights into GSTZ1/MAAI physiological function during human development.
Authorship Contributions
Participated in research design: Li, James, Hines, and Stacpoole.
Conducted experiments: Li and Gu.
Performed data analysis: Li, Gu, James, Hines, Simpson, and Langaee.
Wrote or contributed to the writing of the manuscript: Li, James, Hines, Langaee, and Stacpoole.
Acknowledgements
We thank staff members of the Center for Pharmacogenomics at the University of Florida for conducting GSTZ1 genotyping and Drs. F. Peter Guengerich (Vanderbilt University) and Chen Liu (University of Florida) for providing human livers.
Footnotes
This work was supported in part by the National Institutes of Health National Institute of Environmental Health Sciences [Grants ES007355, ES014617] (to P.W.S.); and the National Institutes of Health National Institute of General Medical Sciences [Grant GM081344] (to R.N.H.).
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
↵
 The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material.
The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material.-
ABBREVIATIONS:
- GSTZ1
- glutathione transferase ζ 1
- DCA
- dichloroacetate
- MAAI
- maleylacetoacetate isomerase
- SNP
- single nucleotide polymorphism
- LOQ
- limit of quantitation.

- Received July 5, 2011.
- Accepted October 25, 2011.
- Copyright © 2012 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}