


Abstract
The glucuronide conjugate of methyl 1-(3,4-dimethoxyphenyl)-3-(3-ethylvaleryl)-4-hydroxy-6,7,8-trimethoxy-2-naphthoate (S-8921; S-8921G) is a 6000-fold more potent inhibitor of an ileal apical sodium-dependent bile acid transporter (SLC10A2) than S-8921 and is responsible for the hypocholesterolemic effect of S-8921 in rats. Because S-8921G is formed in the intestine and liver, the present study investigated the transporters involved in the secretion of S-8921G that govern its exposure to the target site and thereby play an important role in its pharmacological action. Organic anion transporting polypeptide (OATP) 1B1- and OATP1B3-expressing cells exhibited saturable accumulation of S-8921G with Km values (micromolar) of 1.9. The uptake of [14C]S-8921G by human cryopreserved hepatocytes was saturable and sodium-independent. Comparison of protein expression between the cDNA transfectants and hepatocytes suggests that the contribution of OATP1B1, OATP1B3, and Na+-taurocholate cotransporting polypeptide to the hepatic uptake of S-8921G is 63, 35, and 2.6%, respectively. The basal-to-apical transport of S-8921G was enhanced in Madin-Darby canine kidney cells expressing both OATP1B1 and multidrug resistance-associated protein (MRP) 2. In Mrp2-deficient mutant rats [Eisai hyperbilirubinemic rats (EHBR)], the biliary excretion clearance based on the plasma concentration was 20% of the normal value, whereas the pharmacokinetic parameters did not show any significant change in Bcrp-/- mice. Furthermore, the secretion clearance of S-8921G to the mucosal side was also significantly lower in everted jejunum sacs from EHBR (9.18 and 20.8 μl/min/g tissue). These results suggest that MRP2 is responsible for the secretion of S-8921G to the intestinal lumen and bile and that OATP1B1 and OATP1B3 account for the hepatic uptake. These transporters deliver S-8921G to the target site of its pharmacological action.
Hyperlipidemia, especially hypercholesterolemia, is a major risk factor for atherosclerosis, leading to coronary heart disease. In clinical trials, reducing serum low-density lipoprotein (LDL) cholesterol has been shown to reduce the incidence of coronary heart disease and to reverse atherosclerotic lesions in hypercholesterolemic patients (Shepherd et al., 1995). The hypocholesterolemic agents most commonly used are the HMG-CoA reductase inhibitors (so-called statins). Statins have a potent hypocholesterolemic action and few side effects, resulting in good patient compliance. As alternative pharmacological targets for the hypocholesterolemic effect, there is growing interest in inhibition of the intestinal absorption of cholesterol and bile acids. S-8921 is a novel inhibitor of an ileal apical sodium-dependent bile acid transporter (ASBT/SLC10A2) and inhibits the absorption of bile acids from the ileum. The bile acid pool is tightly regulated; thus, inhibition of bile acid reabsorption via ASBT in the ileum by S-8921 would lead to an increase in the biosynthesis of bile acids from cholesterol. This is partly ascribed to the increase in the uptake of plasma LDL cholesterol through the up-regulation of LDL receptors as a result of partial depletion of hepatic cholesterol (Packard and Shepherd, 1982). In addition, S-8921 up-regulates hepatic LDL receptor expression and cholesterol 7α-hydroxylase in rabbits (Higaki et al., 1998). These events result in a reduction in the plasma cholesterol.
Initially, the reduction in plasma cholesterol by S-8921 was ascribed to the direct inhibition of ASBT by S-8921 following p.o. administration. Later, it became evident that the glucuronide conjugate of S-8921 (S-8921G), rather than S-8921 itself, is responsible for the pharmacological action of S-8921. Indeed, S-8921G is a 6000-fold more potent inhibitor of ASBT, and repeated p.o. administration of S-8921 has no significant effect in Gunn rats, which are hereditarily deficient in all UDP-glucuronosyltransferase (UGT) 1 isoforms because of the mutation in the common region of UGT1 isoforms, whereas S-8921G significantly reduces serum non–high-density lipoprotein cholesterol levels (Sakamoto et al., 2007). S-8921 is a substrate of UGT1 isoforms, particularly UGT1A1, UGT1A3, UGT1A8, UGT1A9, and UGT1A10, which are expressed in the liver and intestine (Sakamoto et al., 2007). After p.o. administration of [14C]S-8921 to rats, most of the radioactivity is excreted into the bile as the glucuronide (Yamaguchi et al., 1998). S-8921G is produced in the intestine and then excreted to the mucosal side. Both S-8921G and S-8921 are also transferred to the portal vein following administration of S-8921 to the intestinal lumen and finally excreted into the bile (Sakamoto et al., 2007). Therefore, transporters involved in the biliary excretion and intestinal efflux will play a significant role in delivering S-8921G to the target site. Previously, it was shown that the hepatic uptake of S-8921G by freshly isolated rat hepatocytes was saturable and mediated by sodium-dependent and -independent mechanisms (Sakamoto et al., 2007).
The purpose of the present study is to identify the transporters involved in the hepatobiliary transport and intestinal efflux of S-8921G. In the sinusoidal membrane of human hepatocytes, Na+-taurocholate cotransporting polypeptide (NTCP/SLC10A1) is responsible for the sodium-dependent uptake of bile acids (Hagenbuch and Meier, 1994), and organic anion transporting polypeptides (OATP/SLCO), such as OATP1B1 and OATP1B3, play major roles in the sodium-independent uptake of a variety of amphipathic organic anions (Hagenbuch and Meier, 2003). We examined the uptake of S-8921G by OATP1B1, OATP1B3, and NTCP, and their contribution was evaluated by comparing the protein expression between the expression system and human cryopreserved hepatocytes. Multidrug resistance-associated protein 2 (MRP2/ABCC2) is a well known transporter involved in the biliary excretion of many glucuronide conjugates (Kusuhara and Sugiyama, 2001). In addition, Mrp2 is highly expressed in the duodenum, and its expression decreases from the proximal to the distal region of the small intestine (Maher et al., 2005). Mrp2 also mediates the mucosal efflux of the glucuronide conjugates of xenobiotics into the lumen (Adachi et al., 2005). To identify S-8921G as Mrp2 substrate, transcellular transport of S-8921G was examined in Madin-Darby canine kidney (MDCK) II cells expressing both OATP1B1 and MRP2. In addition, involvement of Mrp2 in the biliary excretion and intestinal efflux was examined using Mrp2-deficient mutant rats [Eisai hyperbilirubinemic rats (EHBR)]. Because it has been shown that breast cancer resistance protein (Bcrp/Abcg2) plays a more significant role in the biliary excretion of some glucuronide conjugates than Mrp2 in mice (Zamek-Gliszczynski et al., 2006b), the biliary excretion of S-8921G was also examined in Bcrp-/- mice.
Materials and Methods
Materials. S-8921 (99.8%), S-8921G (99.7%), and [14C]S-8921G (1.64 MBq/mg) were synthesized at Shionogi Research Laboratories (Shionogi & Co., Ltd., Japan). [3H]Estrone sulfate ([3H]E-sul; 2.1 TBq/mmol), [3H]taurocholic acid ([3H]TCA; 44 GBq/mmol), and [3H]estradiol-17β-d-glucuronide ([3H]E217βG; 1.7 TBq/mmol) were purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). [3H]Cholecystokinin (CCK)-8 (3.63 TBq/mmol) was purchased from GE Healthcare (Little Chalfont, Buckinghamshire, UK). Unlabeled E-sul, TCA, E217βG, and CCK-8 were purchased from Sigma-Aldrich (St. Louis, MO). Tween 80, dimethylacetamide, and saline for injection were purchased from Sigma-Aldrich, Wako Pure Chemicals (Osaka, Japan), and Otsuka Pharmaceutical (Tokyo, Japan), respectively. Human cryopreserved hepatocytes were obtained from Charles River Japan (Yokohama, Japan). Other chemicals were of analytical or reagent grade.
Animals. All the procedures for the animal experiments were approved by the Institutional Animal Care and Use Committee. Male EHBR and male Sprague-Dawley (SD) rats were purchased at 7 weeks of age from Japan SLC (Shizuoka, Japan). The Bcrp-/- mice used in this study had been established previously (Jonker et al., 2002). Male Bcrp-/- and wild-type FVB mice (11–12 weeks of age) were used in the present study. The animals were housed in a room under controlled light (on from 8 AM to 8 PM), temperature (23 ± 3°C), and humidity (30–70%). The rats were allowed free access to tap water and solid laboratory food.
Transport Study Using Transporter Expression Systems. Human OATP1B1-, OATP1B3-, NTCP-, or vector-transfected human embryonic kidney (HEK) 293 cells used in this study were established previously (Hirano et al., 2004; Maeda et al., 2006). The transport study by human OATP1B1-, OATP1B3-, and NTCP-expressing HEK293 cells was carried out as described previously (Hirano et al., 2004; Maeda et al., 2006). Briefly, cells were seeded in 12-well plates at a density of 1.5 × 105 cells/well and cultured with 5 mM sodium butyrate for 24 h. Uptake was initiated by adding Krebs-Henseleit buffer containing radiolabeled and unlabeled substrates after cells had been washed twice and preincubated with Krebs-Henseleit buffer at 37°C for 15 min. The Krebs-Henseleit buffer consisted of 118 mM NaCl, 23.8 mM NaHCO3, 4.83 mM KCl, 0.955 mM KH2PO4, 1.20 mM MgSO4, 12.5 mM HEPES, 5 mM glucose, and 1.53 mM CaCl2 adjusted to pH 7.4. The uptake was terminated at a designated time by adding ice-cold Krebs-Henseleit buffer after removal of the incubation buffer. Then, cells were washed twice with 1 ml of ice-cold Krebs-Henseleit buffer and solubilized in 500 μl of 0.2 N NaOH. Aliquots (800 μl) were transferred to counting vials after adding 500 μl of 0.2 N HCl. The radioactivity associated with the cells and incubation buffer was measured in a liquid scintillation counter (Tri-Carb 3100TR; PerkinElmer Life and Analytical Sciences) after adding 10 ml of scintillation fluid (Pico-Fluor 40; PerkinElmer Life and Analytical Sciences). The remaining 10 μl of cell lysate was used to determine the protein concentration with a Wako protein assay kit (Wako Pure Chemicals) with bovine serum albumin as a standard.
MDCK II cells expressing human OATP1B1, MRP2, and both OATP1B1 and MRP2 had been established previously (Sasaki et al., 2002). The transcellular transport study by transfected MDCK cells was carried out as described previously. Briefly, the transfected MDCK cells were seeded in 24-well plates at a density of 1.4 × 105 cells/well and cultured with 5 mM sodium butyrate for 24 h. Cells were then washed three times and preincubated with Krebs-Henseleit buffer at 37°C for 30 min. The experiments were initiated by replacing the medium on either the apical or the basal side of the cell monolayer; the complete medium contained [14C]S-8921G (1 μM). The cells were incubated at 37°C, and aliquots of medium were taken from each compartment at 30, 60, and 120 min. Radioactivity in an aliquot (100 μl) of medium was measured with a liquid scintillation counter (LS-6000SC; Beckman Coulter, Fullerton, CA) after addition of 5 ml of scintillation fluid (Clear-sol; Nacalai Tesque, Kyoto, Japan). At the end of the experiments, the cells were washed three times with ice-cold transport buffer and solubilized in 450 μl of 0.2 N NaOH. After addition of 450 μl of 0.2 N HCl, an aliquot (800 μl) was transferred to a scintillation vial, and aliquots of cell lysate (20 μl) were used to determine protein concentrations by the method of Lowry et al. (1951) with bovine serum albumin as a standard.
The uptake was expressed as the uptake volume (microliter per milligram of protein), given as the amount of radioactivity associated with the cells (disintegrations per minute per milligram of protein) divided by its concentration in the incubation medium (disintegrations per minute per microliter). The specific uptake was obtained by subtracting the uptake into vector-transfected cells from that into cDNA-transfected cells. Kinetic parameters were obtained using the following equation:  where v is the uptake velocity of the substrate (picomole per minute per milligram of protein), S is the substrate concentration in the medium (micromolar), Km is the Michaelis constant (micromolar), Vmax is the maximum uptake rate (picomoles per minute per milligram of protein), and Pdif is the nonsaturable uptake clearance (microliters per minute per milligram of protein). Fitting was performed by the nonlinear least-squares method using WinNonlin (Pharsight, Mountain View, CA). The input data were weighted as the reciprocals of the observed values.
where v is the uptake velocity of the substrate (picomole per minute per milligram of protein), S is the substrate concentration in the medium (micromolar), Km is the Michaelis constant (micromolar), Vmax is the maximum uptake rate (picomoles per minute per milligram of protein), and Pdif is the nonsaturable uptake clearance (microliters per minute per milligram of protein). Fitting was performed by the nonlinear least-squares method using WinNonlin (Pharsight, Mountain View, CA). The input data were weighted as the reciprocals of the observed values.
Uptake Study Using Human Cryopreserved Hepatocytes. The uptake of S-8921G by human cryopreserved hepatocytes was determined as described previously (Shitara et al., 2003). In this experiment, we selected three batches of human cryopreserved hepatocytes (lot ETR, UYII, and PFQI) purchased from In Vitro Technologies (Baltimore, MD) (lot ETR) and from the Research Institute for Liver Disease (Shanghai, China) (lot UYII and PFQI). Just before the study, the hepatocytes were thawed at 37°C, quickly suspended in 10 ml of ice-cold Krebs-Henseleit buffer, and centrifuged (50g) for 2 min at 4°C, followed by removal of the supernatant. This procedure was repeated once more to remove cryopreservation buffer, and then the cells were resuspended in the same buffer to give a cell density of 1.0 × 106 viable cells/ml for the uptake study. The number of viable cells was determined by the trypan blue exclusion test. In this study, the viability of human hepatocytes was 51 to 78%. To measure the uptake in the absence of Na+, sodium chloride and sodium bicarbonate in Krebs-Henseleit buffer were replaced with choline chloride and choline bicarbonate, respectively. Before the uptake studies, the cell suspensions were prewarmed in an incubator at 37°C for 3 min. The uptake studies were initiated by adding an equal volume of buffer containing labeled and unlabeled substrates to the cell suspension. After incubation at 37°C for 0.5, 2, or 5 min, the reaction was terminated by separating the cells from the substrate solution. For this purpose, an aliquot (70 or 80 μl) of incubation mixture was collected and placed in a centrifuge tube (450 μl) containing 50 μl of 2 N NaOH under a layer of 100 μl of oil (density, 1.015, a mixture of silicone oil and mineral oil; Sigma-Aldrich), and then the sample tube was centrifuged for 15 s using a tabletop centrifuge (16,000g, Eppendorf Micro Centrifuge 5415; Eppendorf North America, New York, NY). During this process, hepatocytes passed through the oil layer into the alkaline solution. After an overnight incubation in alkali to dissolve the hepatocytes, the centrifuge tube was cut, and each compartment was transferred to a scintillation vial. The compartment containing the dissolved cells was neutralized with 50 μl of 2 N HCl and mixed with 10 ml of scintillation mixture (Pico-Fluor 40), and the radioactivity was measured in a liquid scintillation counter (Tri-Carb 3100TR).
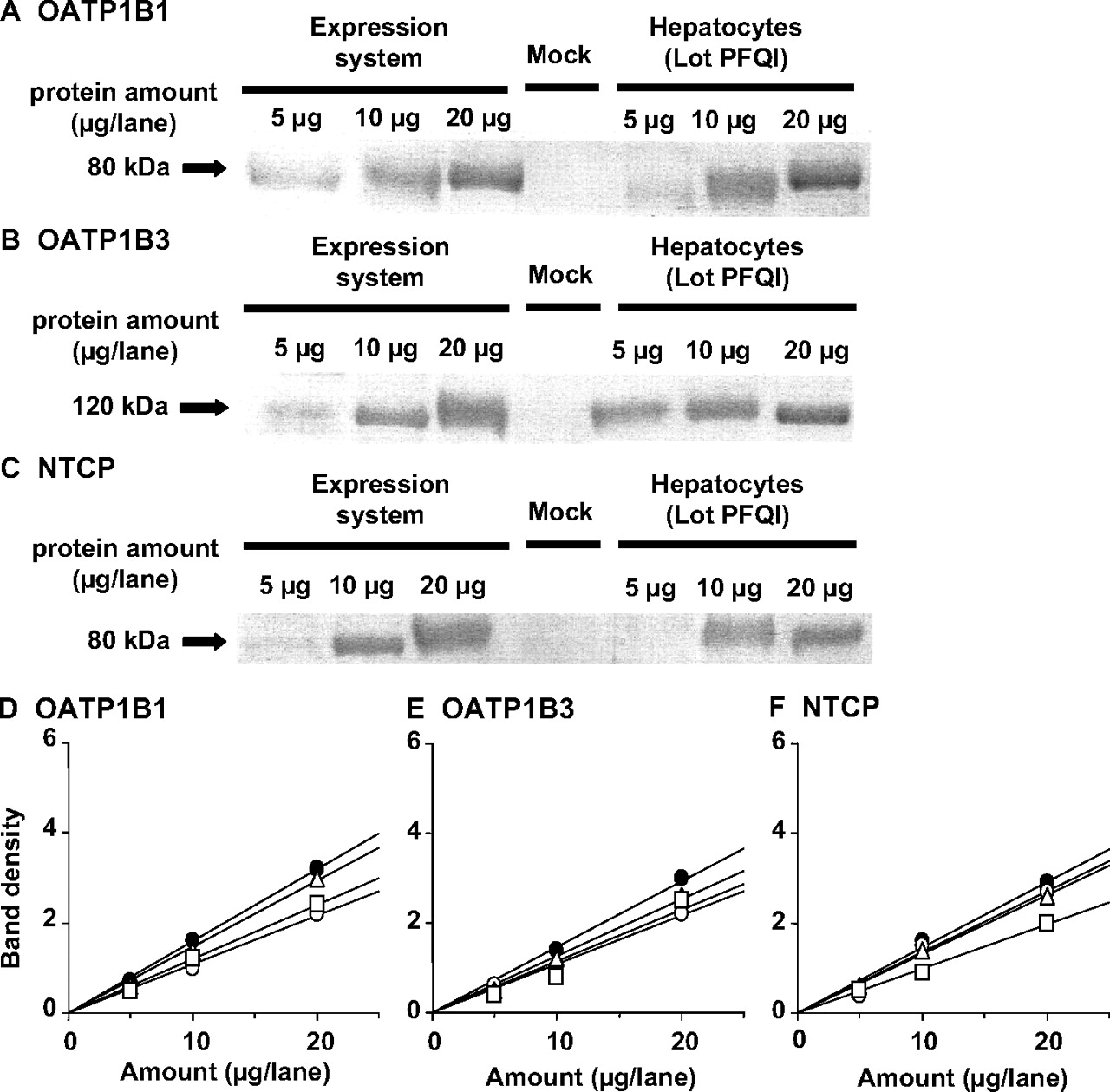
Western Blotting. Anti-OATP1B1 was purchased from PROGEN Biotechnik GmbH (Heidelberg, Germany). As shown in previous reports, anti-OATP1B3 serum was raised in rabbits against a synthetic peptide consisting of the 15 carboxyl-terminal amino acids of OATP1B3 coupled to keyhole limpet hemocyanin at its N terminus via an additional cysteine (Konig et al., 2000). Anti-NTCP serum was raised in rabbits against a synthetic peptide consisting of the 15 carboxyl-terminal amino acids of NTCP at its C terminus. Western blot analysis was performed as described previously (Hirano et al., 2004). Briefly, crude membrane fractions prepared from human hepatocytes and transporter-expressing HEK293 cells were diluted with 3× Red loading buffer (Bio-Labs, Hertfordshire, UK) and loaded onto a 7% SDS-polyacrylamide gel with a 4.4% stacking gel. Proteins were electroblotted onto a polyvinylidene difluoride membrane (Pall, East Hills, NY) using a blotter (Trans-blot; Bio-Rad Laboratories, Hercules, CA) at 15 V for 1 h. The membrane was blocked with Tris-buffered saline containing 0.05% Tween 20 (TBS-T) and 5% skimmed milk for 1 h at room temperature. After washing with TBS-T, the membrane was incubated with anti-OATP1B1 serum (dilution 1:500), anti-OATP1B3 serum (dilution 1:1000), or anti-NTCP serum (dilution 1:500). The membrane was incubated with a horseradish peroxidase-labeled anti-rabbit IgG antibody (Wako Pure Chemicals) diluted 1:1000 in TBS-T for 1 h at room temperature followed by washing with TBS-T. The band was detected, and its intensity was quantified using an image analyzer (LAS-1000 Plus, Fuji Film, Tokyo, Japan).
Contribution of OATP1B1, OATP1B3, and NTCTP to the Net Hepatic Uptake. The contribution of OATP1B1, OATP1B3, and NTCP to the overall uptake by human hepatocytes was estimated according to the method reported previously (Hirano et al., 2004). The protein expression of OATP1B1, OATP1B3, and NTCP was determined in crude membrane from transfected cells and hepatocytes by Western blot analyses. R values were defined for OATP1B1, OATP1B3, and NTCP by the following equation:  where slope represents the slope of the relationship between the applied amount and band density (Fig. 3), and yield is the correction factor, allowing conversion of the protein expression per unit protein of crude membrane to whole cells, and represents the amount of crude membrane protein obtained from 1 mg of whole cell protein for cDNA expression systems or from 106 cells for hepatocytes. The subscripts h, OATP1B1, OATP1B3, and NTCP represent hepatocytes and corresponding cDNA transfectants, respectively. We were able to obtain 137, 102, and 141 μg of protein in crude membrane from 1 mg of whole cell protein in OATP1B1-, OATP1B3-, and NTCP-expressing cells, respectively, whereas 92, 154, and 111 μg of protein in crude membrane were obtained from 106 hepatocytes of lot ETR, UYII, and PFQI, respectively. The OATP1B1-, OATP1B3-, and NTCP-mediated uptake of S-8921G in hepatocytes (CLh,OATP1B1, CLh,OATP1B3, CLh,NTCP) was estimated by multiplying the intrinsic transport activity in cDNA-transfected cells by the R value (eq. 3):
where slope represents the slope of the relationship between the applied amount and band density (Fig. 3), and yield is the correction factor, allowing conversion of the protein expression per unit protein of crude membrane to whole cells, and represents the amount of crude membrane protein obtained from 1 mg of whole cell protein for cDNA expression systems or from 106 cells for hepatocytes. The subscripts h, OATP1B1, OATP1B3, and NTCP represent hepatocytes and corresponding cDNA transfectants, respectively. We were able to obtain 137, 102, and 141 μg of protein in crude membrane from 1 mg of whole cell protein in OATP1B1-, OATP1B3-, and NTCP-expressing cells, respectively, whereas 92, 154, and 111 μg of protein in crude membrane were obtained from 106 hepatocytes of lot ETR, UYII, and PFQI, respectively. The OATP1B1-, OATP1B3-, and NTCP-mediated uptake of S-8921G in hepatocytes (CLh,OATP1B1, CLh,OATP1B3, CLh,NTCP) was estimated by multiplying the intrinsic transport activity in cDNA-transfected cells by the R value (eq. 3):  where CLh presents the uptake clearance mediated by the corresponding transporters in cryopreserved hepatocytes, and CL represents the Vmax/Km value determined in OATP1B1-, OATP1B3-, and NTCP-expressing cells. The contribution of OATP1B1, OATP1B3, and NTCP is defined with reference to the net uptake, namely, the sum of CLh,OATP1B1, CLh,OATP1B3, and CLh,NTCP.
where CLh presents the uptake clearance mediated by the corresponding transporters in cryopreserved hepatocytes, and CL represents the Vmax/Km value determined in OATP1B1-, OATP1B3-, and NTCP-expressing cells. The contribution of OATP1B1, OATP1B3, and NTCP is defined with reference to the net uptake, namely, the sum of CLh,OATP1B1, CLh,OATP1B3, and CLh,NTCP.
Biliary Excretion Study of [14C]S-8921G in Rats. Male EHBR (8–9 weeks of age, body weight of 424–487 g) and male SD rats (8–9 weeks of age, body weight of 373–391 g) were used for these experiments. All the cannulations were performed under ether anesthesia on the dosing day. A polyethylene tube (PE-50; BD Biosciences, San Jose, CA) was inserted into the femoral artery and vein for blood sampling and administration of [14C]S-8921G, respectively. A polyethylene tube (PE-10; Becton Dickinson) was inserted into the common bile duct toward the liver for bile sampling. After cannulation, the animals were housed individually in Bollman cages. After complete recovery from the ether anesthesia, [14C]S-8921G, dissolved in Tween 80, dimethylacetamide, and saline (10:15:75, by volume), was administered i.v. to the rats at a dose of 1 mg/kg. At designated times after dosing, blood and bile were collected. The blood was immediately centrifuged at 16,000g for 5 min at room temperature to obtain plasma. An aliquot (20 μl) of each plasma sample was weighed into a counting vial and mixed with 100 μl of distilled water and 10 ml of scintillation fluid (Hionic Fluor; PerkinElmer Life and Analytical Sciences). The total weight of each collected bile sample was measured, and an aliquot (25–100 μl) was weighed into a counting vial and mixed with 10 ml of Hionic Fluor. The radioactivity was measured in a liquid scintillation counter (LS-6000SC).
Biliary Excretion Study of [14C]S-8921G in Bcrp-/- Mice. Male Bcrp-/- and wild-type FVB mice weighing 27 to 31 g were used for these experiments. All the cannulations were performed under urethane anesthesia on the day of dosing. Under urethane anesthesia (1.4 g/kg), the bile duct was cannulated with a polyethylene catheter (SP-8) for bile collection. After administration of [14C]S-8921G at a dose of 1 mg/kg (vehicle, Tween 80/dimethylacetamide/saline; 10:15:75, by volume), blood samples were collected from the jugular vein. Bile was collected at 30-min intervals throughout the experiments. The blood was immediately centrifuged at 16,000g for 3 min at room temperature to obtain plasma. An aliquot (25 μl) of each plasma sample was weighed into a counting vial and mixed with 200 μl of distilled water and 10 ml of Pico-Fluor 40. An aliquot (10 μl) of each bile sample was weighed into a counting vial and mixed with 10 ml of Pico-Fluor 40. The radioactivity was measured in a liquid scintillation counter (Tri-Carb 3100TR).

Saturable uptake of [14C]S-8921G by OATP1B1-, OATP1B3-, and NTCP-expressing HEK293 cells. A, the uptake of [14C]S-8921G (2 μM) by HEK293 cells expressing human OATP1B1, OATP1B3, or NTCP was determined at 37°C. Open and closed circles represent the uptake in vector-transfected and transporter-expressing cells, respectively. B, the concentration dependence of the uptake of [14C]S-8921G by HEK293 cells expressing human OATP1B1 and OATP1B3 is shown as Eadie-Hofstee plots. The uptake of [14C]S-8921G was measured between 0.5 and 50 μM for 2 min at 37°C. The uptake of [14C]S-8921G by NTCP-expressing HEK293 cells at tracer (2 μM) and excess (50 μM) concentration was determined for 2 min at 37°C. Transporter-mediated uptake was calculated by subtracting the uptake into vector-transfected cells from that into transporter-expressing cells. Each data point and vertical bar represent the mean ± S.D. of three experiments.
Pharmacokinetic Analysis. Pharmacokinetic analysis was performed using WinNonlin (Pharsight) software based on a noncompartment model with uniform weighting. The total and biliary excretion clearances (CLtot,p and CLbile,p, respectively) of [14C]S-8921G were calculated from the following equation (eq. 4):  where fe,bile is the fraction of the cumulative biliary excretion of radioactivity within 240 min for rats or 120 min for mice after dosing, and AUC is the area under the plasma concentration-time curve from zero to infinity in rats or to 120 min in mice. AUC was obtained by the linear trapezoidal rule.
where fe,bile is the fraction of the cumulative biliary excretion of radioactivity within 240 min for rats or 120 min for mice after dosing, and AUC is the area under the plasma concentration-time curve from zero to infinity in rats or to 120 min in mice. AUC was obtained by the linear trapezoidal rule.
Everted Sac Study. SD rats and EHBR were anesthetized with ether and sacrificed by exsanguination from the femoral artery. Immediately after sacrifice, the jejunum (approximately 5 cm away from the ligament of Treitz) was dissected. The segment was ligated at one end and then everted. The open end of the everted sac was ligated after the insertion of a polyethylene tube (PE-50; Natsume, Tokyo, Japan). The length of the prepared everted sac was 5 cm. Krebs-Ringer buffer (KRB, pH 6.4, 500 μl) was added to the serosal side of the everted sac through the cannula followed by incubation in KRB for 10 min at 37°C. Then, the sac was transferred to KRB containing 10 μM S-8921 and incubated at 37°C. Aliquots (500 μl) were collected from the mucosal solution at 0, 10, 20, 40, and 60 min after incubation. At the last sampling point, the serosal solution was collected via the cannula. The sacs were washed with ice-cold PBS, weighed, and homogenized with a 9-fold volume of PBS to obtain 10% tissue homogenates. The amounts of S-8921G secreted to the mucosal side were calculated by multiplying the concentrations of S-8921G by the volume of the mucosal solution. The secretion clearance to the mucosal side (CLmucosal) was calculated by dividing the efflux rate of S-8921G to the mucosal side by the intestinal concentration of S-8921G at 60 min.
For the measurement of S-8921G, an aliquot (100 μl) of sample was added to 220 μl of acetonitrile followed by centrifugation at 16,000g for 3 min at room temperature. The supernatant (20 μl) was subjected to high-performance liquid chromatography analysis under the following chromatographic conditions: pump model LC-10AD, UV detector model SPD-10AVvp (λ = 260 nm), controller model SCL-10Avp (Shimadzu, Kyoto, Japan); column, Cosmosil 5C18 AR-II (150 × 3.0 mm i.d.; Nacalai Tesque); mobile phase A, 0.1% acetate in water and mobile phase B, 0.1% acetate in acetonitrile (A/B = 70:30–15:85, v/v); and flow rate, 0.4 ml/min.
Statistical Analysis. Statistical analysis for significant differences was performed by the Welch test using the SAS program (SAS Institute, Cary, NC). A probability <0.05 was considered to be statistically significant.
Results
Uptake of [14C]S-8921G by OATP1B1-, OATP1B3-, and NTCP-Expressing HEK293 Cells. [3H]E-sul (0.1 μM), [3H]CCK-8 (0.1 μM), and [3H]TCA (1 μM) were used as a positive control for OATP1B1-, OATP1B3-, and NTCP-expressing HEK293 cells. The uptake of [3H]E-sul, [3H]CCK-8, and [3H]TCA was similar to that in the published literature (Hirano et al., 2004; Maeda et al., 2006) (data not shown). The uptake of [14C]S-8921G by OATP1B1-, OATP1B3-, and NTCP-expressing HEK293 cells was significantly greater than that in vector-transfected cells (Fig. 1A). The specific uptake of [14C]S-8921G consists of one saturable component (Fig. 1B). The kinetic parameters are summarized in Table 1.
Kinetic parameters of the uptake of [14C]S-8921G by OATP1B1-, OATP1B3-, and NTCP-expressing HEK293 cells Data shown in Fig. 1B were used to determine the parameters calculated by nonlinear regression analysis.
Uptake of [14C]S-8921G by Human Cryopreserved Hepatocytes. [3H]E217βG(1 μM) and [3H]TCA (1 μM) were used as a positive control. The uptake of [3H]E217βG and [3H]TCA was observed in all the batches of human hepatocytes (lot ETR, UYII, and PFQI). The uptake clearance of [3H]E217βG was 13.6 ± 4.8 μl/min/106 cells (lot ETR, 8.11; UYII, 15.6; PFQI, 17.0 μl/min/106 cells). The uptake clearance of [3H]TCA in the presence and absence of Na+ was 7.67 ± 2.80 μl/min/106 cells (lot ETR, 9.66; UYII, 8.88; PFQI, 4.47 μl/min/106 cells) and 3.89 ± 3.17 μl/min/106 cells (lot ETR, 5.83; UYII, 5.61; PFQI, 0.235 μl/min/106 cells), respectively.
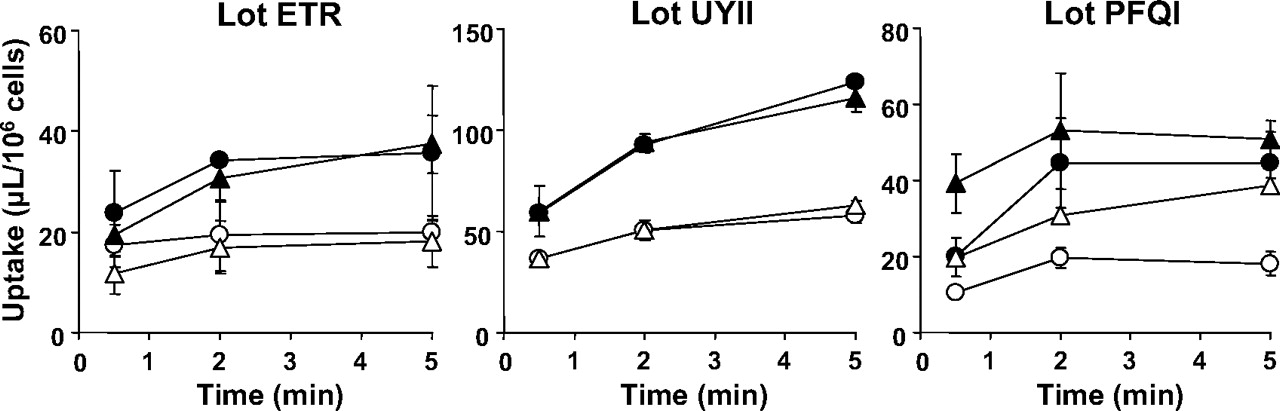
The uptake of [14C]S-8921G (5 μM) was determined in three batches of human cryopreserved hepatocytes (lot ETR, UYII, and PFQI). The uptake was significantly reduced in the presence of an excess (500 μM) of unlabeled S-8921G in all the batches of hepatocytes. The uptake of [14C]S-8921G by human hepatocytes was determined in the presence and absence of Na+. Absence of Na+ did not affect the uptake of [14C]S-8921G in all the batches of human hepatocytes (Fig. 2).
Contribution of OATP1B1, OATP1B3, and NTCP to the Hepatic Uptake of S-8921G. The protein expression of OATP1B1, OATP1B3, and NTCP was determined in crude membrane from transfected cells and hepatocytes by Western blot analyses. The molecular weights of OATP1B1, OATP1B3, and NTCP in the human hepatocytes were almost the same as those in the corresponding cDNA transfectants (Fig. 3). No expression of OATP1B1, OATP1B3, and NTCP was observed in vector-transfected HEK293 cells (Fig. 3). A linear relationship was observed between the applied amount of crude membrane obtained from transporter-expressing cells and human hepatocytes (Fig. 3). Comparison of the predicted clearances suggests the contribution of OATP1B1, OATP1B3, and NTCP to be 60.6 to 64.6, 33.0 to 36.2, and 2.1 to 3.2%, respectively (Table 2).
Uptake clearances of [14C]S-8921G by human hepatocytes and contribution of OATP1B1, OATP1B3, and NTCP to the hepatic uptake of [14C]S-8921G Uptake clearances (CLhep) were calculated by subtracting CL (2–0.5 min) in the presence of excess unlabeled substrate from that in the presence of tracer substrate. Values in parentheses represent the ratio of the OATP1B1-, OATP1B3-, or NTCP-mediated uptake clearance relative to the sum of the estimated clearance mediated by OATP1B1, OATP1B3, and NTCP.
Transcellular Transport of [14C]S-8921G across OATP1B1-, MRP2-, and OATP1B1/MRP2-Expressing MDCK Monolayers. [3H]E217βG(1 μM) was used as a positive control. Its basal-to-apical flux was significantly greater in human OATP1B1/MRP2-MDCK cells than that in the opposite direction (data not shown). In the control cells, human OATP1B1-, and human MRP2-MDCK cells, the basal-to-apical flux of [14C]S-8921G was slightly higher than the apical-to-basal flux (Fig. 4). On the other hand, in the human OATP1B1/MRP2-MDCK cells, [14C]S-8921G was significantly transported in the basal-to-apical direction (Fig. 4). These results show that S-8921G is a substrate of MRP2.
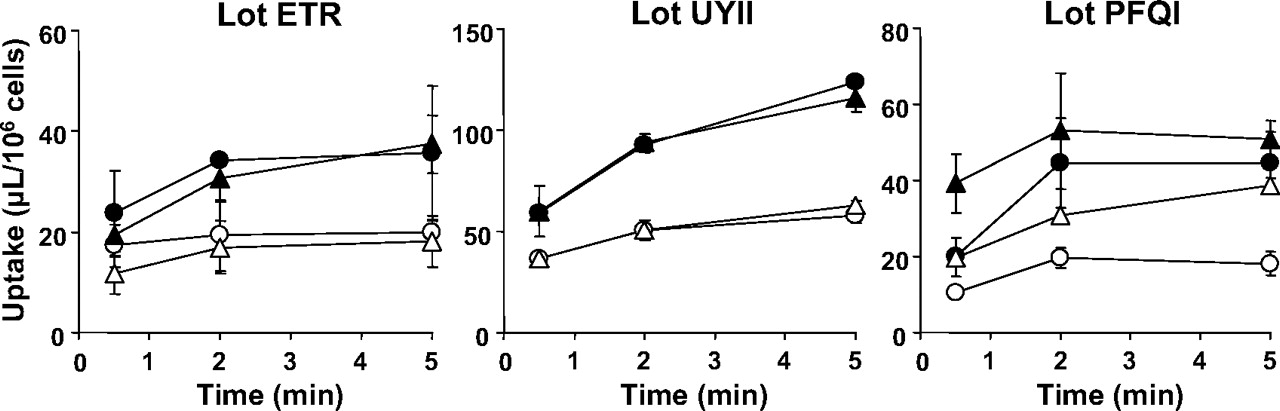
Time-profiles of uptake of [14C]S-8921G by human hepatocytes (lot ETR, UYII, and PFQI). The uptake of [14C]S-8921G for 0.5, 2, and 5 min was determined at two concentrations (closed symbol, 5 μM; open symbol, 500 μM) at 37°C. Circles and triangles indicate the [14C]S-8921G uptake in the presence and absence of Na+, respectively. Each point and vertical bar represent the mean ± S.D. (n = 3).
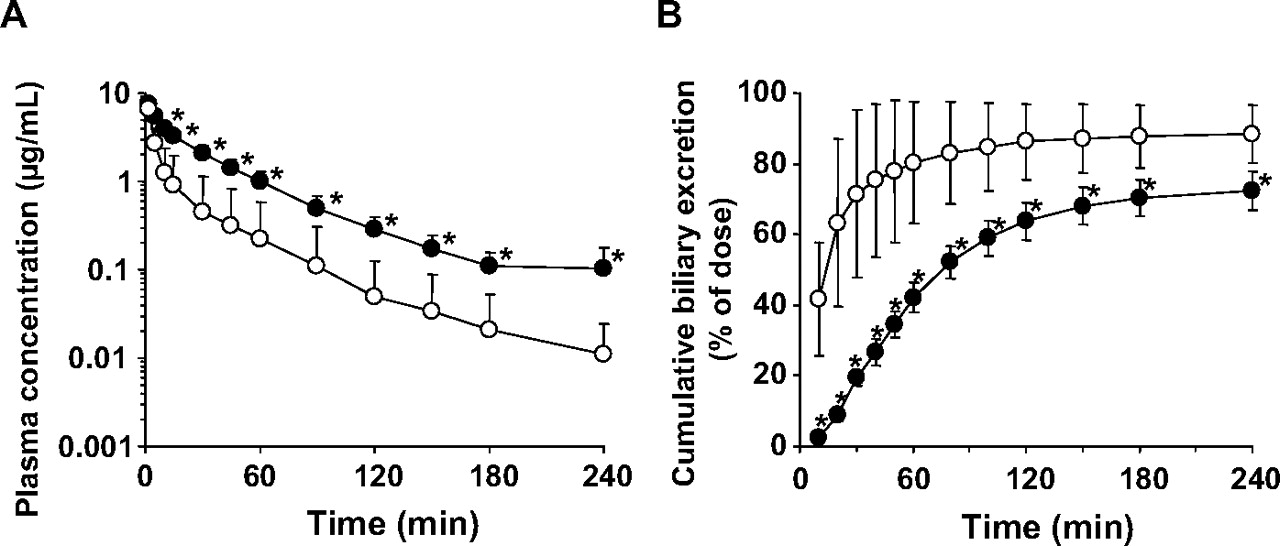
Biliary Excretion after i.v. Administration of [14C]S-8921G in EHBR. Pharmacokinetic analysis following i.v. administration of [14C]S-8921G at 1 mg/kg was carried out in SD rats and EHBR. The kinetic parameters are summarized in Table 3. Most of the radioactivity in plasma specimens at 240 min after dosing of [14C]S-8921G was associated with the unchanged form (data not shown). In both SD rats and EHBR, biliary excretion is the major elimination pathway of [14C]S-8921G (Fig. 5). The plasma concentrations of [14C]S-8921G were higher in EHBR than in SD rats (Fig. 5), resulting in a 3.8-fold lower total body clearance (Table 3). The biliary excretion of [14C]S-8921G was delayed in EHBR, although the cumulative amount excreted into the bile was similar. The biliary excretion clearance of [14C]S-8921G based on the plasma concentrations was 5-fold lower in EHBR than in SD rats (Table 3). The cumulative urinary excretion of radioactivity up to 240 min after dosing in EHBR (16.9 ± 6.3% of dose) was higher than that in SD rats (5.55 ± 3.95% of dose). Total recoveries of radioactivity at 240 min after dosing were similar in SD rats (89.2 ± 4.0% of dose) and EHBR (94.1 ± 4.4% of dose). The concentration of radioactivity in the liver at 240 min after dosing in EHBR (0.286 ± 0.101 μg equivalent of S-8921G/g liver) was 5-fold higher than that in SD rats (0.055 ± 0.064 μg equivalent of S-8921G/g liver).
Pharmacokinetic parameters of radioactivity after i.v. administration of [14C]S-8921G in SD rats and EHBR The pharmacokinetic parameters were calculated by WinNonlin using the plasma concentrations shown in Fig. 5. Data represent the mean ± S.D. of four rats.
Biliary Excretion after i.v. Administration of [14C]S-8921G in Bcrp-/- Mice. The time-profiles of the plasma concentrations and biliary excretion of [14C]S-8921G were unchanged in Bcrp-/- mice (Fig. 6). The biliary excretion was also the predominant elimination pathway of S-8921G in mice (Bcrp-/- mice, 93.9 ± 2.6% of dose; wild-type mice, 92.0 ± 6.4% of dose). The biliary excretion clearance based on the plasma concentrations was 13.6 ± 4.5 and 8.44 ± 2.10 ml/min/kg in Bcrp-/- and wild-type mice, respectively, and was not significantly different (Table 4).
Pharmacokinetic parameters of radioactivity after i.v. administration of [14C]S-8921G in wild-type and Bcrp–/– mice The pharmacokinetic parameters were calculated by WinNonlin using the plasma concentrations shown in Fig. 6. Data represent the mean ± S.D. of three mice.
Efflux of S-8921G into the Intestinal Lumen in EHBR. The efflux of S-8921G was investigated in everted jejunum sacs (Fig. 7). After incubation of S-8921 in everted jejunum sacs, S-8921G was detected both in the mucosal solution and tissue. The mucosal efflux rate of S-8921G in EHBR was reduced compared with that in SD rats, and the tissue concentration of S-8921G was 1.3-fold greater in the jejunum everted sacs of EHBR, resulting in a 2-fold reduction in the CLmucosal of S-8921G in EHBR (SD rats, 20.8 ± 3.7; EHBR, 9.18 ± 1.99 μl/min/g tissue).
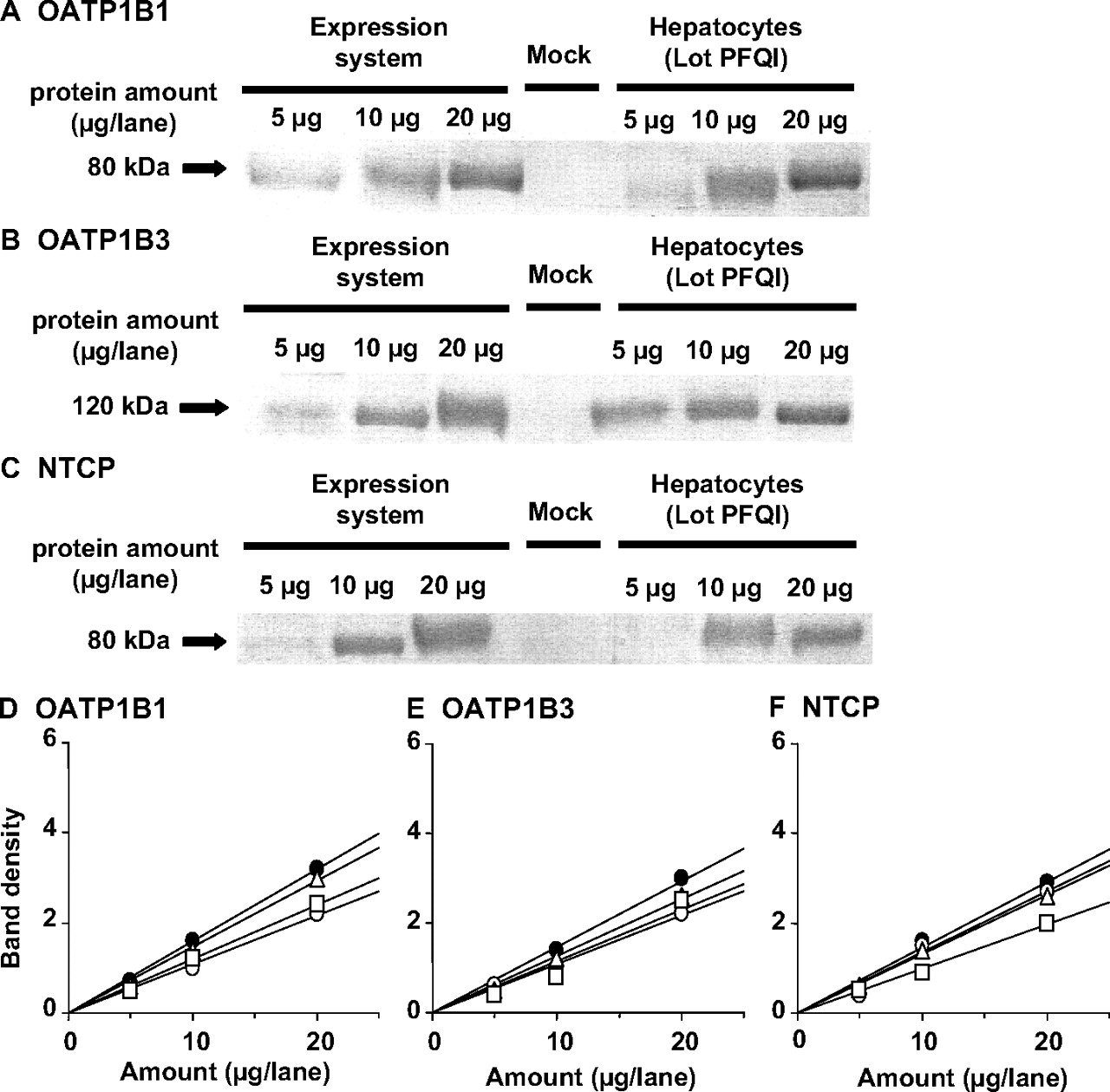
Western blot analysis of OATP1B1, OATP1B3, and NTCP. Crude membrane fractions (5, 10, and 20 μg) prepared from OATP1B1-, OATP1B3-, and NTCP-expressing HEK293 cells and human hepatocytes (lot PFQI) were loaded and separated by SDS-polyacrylamide gel electrophoresis (7% separating gel). OATP1B1 (A), OATP1B3 (B), and NTCP (C) were detected by preimmune antisera raised against the carboxyl terminus of human OATP1B1, OATP1B3, and NTCP, respectively. Comparison of the relative expression levels of OATP1B1 (D), OATP1B3 (E), and NTCP (F) between transfectants and human hepatocytes is shown. The x-axis and y-axis represent the amount of crude membrane obtained from transfectants and human hepatocytes and the intensity of the specific band in Western blot analysis, respectively. Closed circles, open circles, open squares, and open triangles indicate the band density of the transporter expressing HEK293 cells, lot ETR, lot UYII, and lot PFQI of human hepatocytes, respectively.
Discussion
Glucuronidation plays a key role in the pharmacological action of S-8921, converting S-8921 to a 6000-fold more potent inhibitor. Because glucuronidation of S-8921 occurs in the small intestine and liver, secretion into the intestinal lumen directly or indirectly through biliary excretion governs exposure of S-8921G to the target site. In the present study, the transporters involved in the hepatobiliary and efflux transport in the small intestine were examined.
In vitro transport studies using HEK293 cells expressing hepatic uptake transporters identified S-8921G as a substrate of OATP1B1, OATP1B3, and NTCP with similar Km values, although the transport activity by NTCP was markedly lower than that by OATPs (Fig. 1). Saturable uptake of [14C]S-8921G was observed in human hepatocytes (Fig. 2). Unlike rat hepatocytes (Sakamoto et al., 2007), the uptake of S-8921G by human cryopreserved hepatocytes did not show sodium dependence. Furthermore, according to the method reported by Hirano et al. (2004), the contribution of OATP1B1, OATP1B3, and NTCP to the net hepatic uptake of S-8921G was evaluated by comparing protein expression in cDNA transfectants and cryopreserved human hepatocytes. Although the sum of the predicted clearances extrapolated from cDNA transfectants was greater than the observed values in the cryopreserved hepatocytes, this method allows estimation of the contribution of each transporter involved in the hepatic uptake process (Hirano et al., 2004). As summarized in Table 2, both OATP1B1 and OATP1B3 are mainly involved in the hepatic uptake, making an almost identical contribution, whereas NTCP makes only a marginal contribution. The small contribution of NTCP is consistent with the absence of sodium dependence in the uptake of S-8921G by human cryopreserved hepatocytes. The mechanism underlying the species difference in the sodium dependence between rat and human hepatocytes remains unknown. NCTP may exhibit a species difference in terms of the transport activity of S-8921G. Such a species difference has been reported for rosuvastatin. It should be noted that, in this case, human NTCP accepts rosuvastatin as a substrate, whereas rat Ntcp does not (Ho et al., 2006).

Time-profiles of transcellular transport of [14C]S-8921G across OATP1B1-, MRP2-, and OATP1B1/MRP2-expressing MDCK monolayers. The time-profiles of transcellular transport of [14C]S-8921G (1 μM) across the monolayers of vector-transfected (A), OATP1B1- (B), MRP2- (C), and OATP1B1/MRP2-expressing (D) MDCK cells. Open and closed circles represent the transcellular transport in the apical-to-basal and basal-to-apical directions, respectively. Each point and vertical bar represent the mean ± S.D. of four experiments.
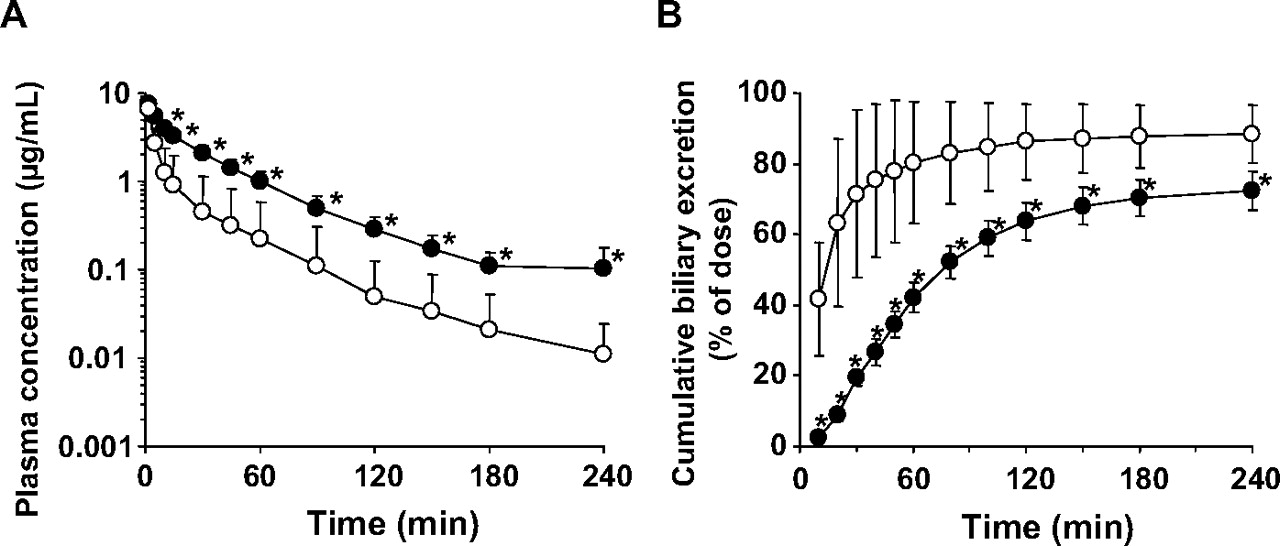
Plasma concentration and cumulative biliary excretion of radioactivity after i.v. administration of [14C]S-8921G in SD rats and EHBR. The plasma concentration (A) and biliary excretion (B) of radioactivity after i.v. administration of [14C]S-8921G (1 mg/kg) were determined in SD rats (open circle) and EHBR (closed circle). Each symbol and vertical bar represent the mean ± S.D. of four rats. *, p < 0.05; the statically significant differences between SD rats and EHBR.

Plasma concentration and cumulative biliary excretion of radioactivity after i.v. administration of [14C]S-8921G in wild-type and Bcrp-/- mice. The plasma concentration (A) and biliary excretion (B) of radioactivity after i.v. administration of [14C]S-8921G (1 mg/kg) were determined in wild-type mice (open circle) and Bcrp-/- mice (closed circle). Each symbol and vertical bar represent the mean ± S.D. of three mice.
To obtain an insight into the canalicular efflux process, the transcellular transport was performed using MDCK cells expressing both OATP1B1 and MRP2. The basal-to-apical transport of S-8921G was observed in the double transfectants (Fig. 4), indicating that S-8921G is a substrate of MRP2. The role of MRP2 in the biliary excretion of S-8921G was further investigated in Mrp2-deficient mutant rats (EHBR). In EHBR, both systemic elimination and biliary excretion of S-8921G were significantly delayed, although the recovery of S-8921G in the bile was only slightly reduced. The total body and biliary excretion clearances of S-8921G based on the plasma concentrations were significantly reduced in EHBR, whereas the radioactivities associated with liver specimens at 240 min after dosing were greater in EHBR than in SD rats. Therefore, the reduction in the biliary clearance based on plasma concentration will be ascribed to the impaired biliary excretion mediated by Mrp2. In addition, because MRP3 expression is markedly up-regulated in EHBR (Akita et al., 2001) and it mediates the basolateral efflux of glucuronide conjugates in the mouse liver (Zelcer et al., 2005, 2006; Zamek-Gliszczynski et al., 2006a), the greater sinusoidal efflux by Mrp3 can be part of mechanisms to account for the increase in the AUC of S-8921G in EHBR. The recovery of S-8921G in the bile for 240 min was almost identical between SD rats and EHBR, and this is ascribed to the minimal contribution of the renal clearance to the net systemic elimination (Table 3).
The CLbile,p of S-8921G was markedly decreased in EHBR but not impaired. Whether this is mediated by other transporters remains unclear. Because Zamek-Gliszczynski et al. (2006b) reported that Bcrp mediates biliary excretion of some glucuronide conjugates together with Mrp2 in mice, Bcrp is one of the candidate transporters. Therefore, the pharmacokinetic profiles of S-8921G were compared between wild-type and Bcrp-/- mice. It was found that neither the plasma concentration time-profile nor the biliary excretion of S-8921G was changed. Therefore, Bcrp makes a limited contribution to the biliary excretion of S-8921G, if any, at least in mice (Table 3), although the contribution of Bcrp remains to be evaluated in rat liver. The fact that S-8921G is a substrate of NTCP, bile salt exporting pump (ABCB11) is another candidate transporter. Bile salt exporting pump mediates biliary excretion of bile acids, but it accepts nonbile acids, such as pravastatin, as a substrate (Hirano et al., 2005). Further studies, particularly in vitro transport studies using gene expression system, are necessary to elucidate the biliary excretion mechanisms of S-8921G in rat liver.
If the urinary excretion clearance is very low, a certain amount is excreted in the bile, enough even when the biliary excretion clearance in EHBR is reduced. In the case of S-8921G, because the urinary excretion clearance is very low and the biliary excretion clearance in EHBR is reduced to approximately 20% of control, we consider that MRP2 plays an important role in the biliary excretion of S-8921G. However, also in such a case, it is reported that the biliary excretion in Bcrp-/- mice decreases. So, we have performed the study in Bcrp-/- mice, and the result shows that the contribution of Bcrp is very small.
In addition to the liver, Mrp2 is also expressed in the brush-border membrane of the small intestine (Mottino et al., 2000; Rost et al., 2002). In particular, it has been reported that S-8921G is formed in intestinal epithelial cells followed by secretion into the intestinal lumen (Sakamoto et al., 2007). To examine the involvement of Mrp2, the mucosal efflux rate of S-8921G was compared in everted jejunum sacs from SD and EHBR. After incubation of S-8921 with everted jejunum sacs, the efflux rate of S-8921G was significantly reduced in EHBR with the tissue concentrations being similar (Fig. 6). The CLmucosal of S-8921G, representing an intrinsic efflux activity across the brush-border membrane, was 44% of the control value in EHBR. Therefore, Mrp2 is also involved in the intestinal secretion of S-8921G. When the efflux to the luminal side is the major sequestration pathway from inside the cells, the tissue concentrations should be increased in the everted jejunum sacs from EHBR. Therefore, the unaltered tissue concentration suggests that an alternative route, namely, basolateral efflux, will make a greater contribution than the luminal efflux. This is consistent with a previous report where the cumulative amount of S-8921G recovered in the bile was 5-fold greater than that in the lumen 6 h after S-8921 was applied directly to the loop of the jejunum in rats that had undergone bile duct cannulation (Sakamoto et al., 2007). The transporter responsible for the basolateral efflux remains unknown. Recently, Mrp3 has been shown to play a significant role in the sinusoidal efflux of glucuronide conjugates (Shoji et al., 2004; Zelcer et al., 2006). Because Mrp3 is also expressed in the basolateral membrane of the intestinal epithelial cells (Rost et al., 2002), Mrp3 may play a role in the absorption of S-8921G into the blood circulation in the small intestine.

Mucosal efflux of S-8921G in the everted jejunum sac. The everted sacs from SD rats (closed circle) and EHBR (open circle) were incubated with S-8921 (10 μM) at 37°C. A, the mucosal efflux rates of S-8921G were determined at designated times. B, tissue concentrations of S-8921G were determined after a 60-min incubation. Data are represented as mean ± S.D. of four determinations using four everted sacs independently prepared from four rats. *, p < 0.05; statically significant differences between SD rats and EHBR.
The present study has identified the transporters involved in the hepatobiliary transport and intestinal efflux of S-8921G. OATP1B1 and MRP2 play important roles in the hepatobiliary transport of S-8921G absorbed into the portal vein, and MRP2 is also involved in the efflux of S-8921G into the intestinal lumen. These transporters will efficiently deliver S-8921G to the lumen of the small intestine, the target site of its pharmacological action. Their importance in the cholesterol-lowering effect of S-8921 will be examined in future studies. Considering that the biliary excretion of S-8921G was delayed in EHBR with only a slight reduction in the cumulative biliary excretion, functional impairment of Mrp2 alone might not significantly influence the cholesterol-lowering effect of S-8921. Further studies are necessary to elucidate the importance of transporters in the pharmacological action of S-8921.
In conclusion, it was found that S-8921G is a substrate of OATP1B1, OATP1B3, NTCP, and MRP2. Both OATP1B1 and OATP1B3 mediate the hepatic uptake of S-8921G, and MRP2 plays significant roles in the biliary excretion across the canalicular membrane and secretion into the intestinal lumen.
Acknowledgments
We thank Dr. Alfred H. Schinkel (The Netherlands Cancer Institute, Amsterdam, The Netherlands) for supplying Bcrp knockout mice and Dr. Piet Borst (The Netherlands Cancer Institute) for providing the parent and MRP2-MDCK cells.
Footnotes
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.108.020511.
-
ABBREVIATIONS: LDL, low-density lipoprotein; S-8921, methyl 1-(3,4-dimethoxyphenyl)-3-(3-ethylvaleryl)-4-hydroxy-6,7,8-trimethoxy-2-naphthoate; ASBT, apical sodium-dependent bile acid transporter; S-8921G, glucuronide conjugate of S-8921; UGT, UDP-glucuronosyltransferase; NTCP, Na+-taurocholate cotransporting polypeptide; OATP, organic anion transporting polypeptides; MRP, multidrug resistance-associated protein; MDCK, Madin-Darby canine kidney; EHBR, Eisai hyperbilirubinemic rats; BCRP, breast cancer resistance protein; E-sul, estrone sulfate; TCA, taurocholic acid; E217βG, estradiol-17β-d-glucuronide; CCK, cholecystokinin; SD, Sprague-Dawley; HEK, human embryonic kidney; TBS-T, Tris-buffered saline/Tween 20; AUC, area under the plasma concentration-time curve; KRB, Krebs-Ringer buffer.
- Received January 17, 2008.
- Accepted May 8, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}