


Abstract
SIB-1765F ([±]-5-ethynyl-3-(1-methyl-2-pyrrolidinyl)pyridine fumarate) is a novel nicotinic acetylcholine receptor (NAChR) agonist displaying a different in vitro pharmacological profile than nicotine and epibatidine, suggestive of NAChR subtype selectivity. Our study describes the effects of SIB-1765F on locomotor activity in rats, which were compared to those observed for nicotine and epibatidine. The three NAChR agonists decreased or increased locomotor activity in rats naive or habituated to the test apparatus, respectively. The transient reduction in locomotor activity induced by SIB-1765F was quantitatively similar to those induced by nicotine and epibatidine but, unlike the effects of nicotine and epibatidine, was not blocked by the NAChR antagonists mecamylamine and dihydro-β-erythroidine, suggesting different mechanisms of action. Furthermore, SIB-1765F produced a larger and longer-lasting increase in locomotor activity when administered to rats familiar with the test apparatus. Mecamylamine and dihydro-β-erythroidine but not hexamethonium blocked the increase in locomotor activity induced by SIB-1765F, suggesting that SIB-1765F elicits this effect predominantly through the activation of central NAChR. The SIB-1765F-induced increase in locomotor activity was also attenuated by selective D1 and D2 dopamine receptor antagonists, implying that this increase in locomotor activity is mediated through the activation of dopamine receptors subsequent to the release of dopamine. Based on these results, SIB-1765F appears to have a different locomotor activity profile than nicotine and epibatidine.
Conflicting reports have appeared in the literature showing stimulant or depressant effects of nicotine on locomotor behavior in rats. This discrepancy may be explained by differences in age/weight, housing conditions, time of day or strain; but, most importantly, may be attributed to the environmental experience of the rats before testing. Indeed, acute administration of nicotine to rats naive to the test apparatus reduces the high levels of exploratory locomotor activity seen upon exposure to this novel environment. This effect seems to be related primarily to side effects induced by nicotine and may be followed by a slight increase in locomotor activity as the rats recover and habituate to the test apparatus (Stolerman et al., 1973; Clarke and Kumar, 1983). Repeated administration of nicotine produces a rapid tolerance to the depressant locomotor effects seen in this setting (Morrison and Stephenson, 1972; Stolerman et al., 1973, 1974). Alternatively, when the basal level of locomotor activity is low due to habituation of the rats to the test apparatus, acute administration of nicotine increases locomotor activity after an insignificant depressant effect (O’Neill et al., 1991; Benwell and Balfour, 1992). With low basal activity, sensitization to the stimulant effect of nicotine develops rapidly after repeated administration of the compound (Clarke and Kumar, 1983; Ksir et al., 1987; Benwell and Balfour, 1992). Thus, nicotine may decrease or increase locomotor activity, depending on previous exposure of the animals to the drug and/or to the test apparatus.
Interestingly, other NAChR agonists produce different effects on locomotor activity. For instance, lobeline, anatoxin and isoarecolone decrease locomotor activity in rats not habituated to the test environment, but fail to increase locomotion in conditions where nicotine produces a stimulant effect (Reavill et al., 1990;Stolerman et al., 1992, 1995; Whiteaker et al., 1995). The different effects of NAChR agonists on locomotor activity may be the result of differential effects on DA release (Mirza et al., 1996). The locomotor stimulant effect of nicotine is predominantly mediated through action on the mesolimbic DA system, at the level of the cell bodies in the ventral tegmentum area (Reavill and Stolerman, 1990; Museo and Wise, 1990; Leikola-Pelho and Jackson, 1992) and/or at the level of DA terminal fields in the nucleus accumbens (Imperato et al., 1986, Damsma et al., 1989;Welzl et al., 1990). The striatum also appears to be involved in nicotine-induced locomotor activity (Richardson and Tizabi, 1994), as nicotine has been shown to release DA in the striatum (Damsmaet al., 1988). Differences in the effects of NAChR agonists on locomotor activity may also be related to the presence of various NAChR subtypes in the central nervous system. Molecular and electrophysiological studies suggest that NAChR subtypes are structurally and functionally diverse (for review, see Patrick and Luetje, 1993). Thus, differences in locomotor effects may reflect different actions of NAChR agonists at distinct subpopulations of NAChR or differential activities at the same receptor(s) (i.e., partial agonist vs full agonist). The unraveling of the pharmacology of NAChR has been hampered by the limited number of subtype-selective NAChR agonists and antagonists.
SIB-1765F is a novel NAChR agonist with an in vitropharmacological profile suggestive of NAChR subtype selectivity (Lloydet al., 1995, Sacaan et al., accompanying manuscript). SIB-1765F displaces [3H]-cytisine binding to rat cortical membrane with high affinity but has a lower affinity than nicotine at muscarinic cholinergic and α-bungarotoxin binding sites. SIB-1765F is moderately more effective than nicotine at releasing DA from rat striatal and olfactory tubercle slices in vitro(Rao et al., 1995; Sacaan et al., accompanying manuscript) and is as effective as nicotine at releasing [3H]-NE from rat thalamic and cortical slices. However, unlike nicotine, SIB-1765F only slightly stimulates [3H]-NE release from rat hipoccampal slices. Because the release of striatal DA and hippocampal NE appears to be regulated by distinct NAChR (Sacaan et al., 1995; Clarke and Reuben, 1996), differential effects of SIB-1765F on striatal DA and hippocampal NE release suggest that SIB-1765F may preferentially activate specific neuronal NAChR subtypes as compared to nicotine (Lloyd et al., 1995; Sacaan et al., accompanying manuscript).
Our experiments have been carried out to determine whether the different pharmacological profile of SIB-1765F, as compared to nicotine, translates into a different locomotor profile. SIB-1765F was also compared to epibatidine, a potent NAChR agonist that has recently been demonstrated to increase locomotor activity in rats (Sacaanet al., 1996). The three NAChR agonists were administered to rats that were naive or habituated to the test environment to compare the locomotor effects of each compound. In addition, the mechanisms by which these NAChR agonists produced their effects on locomotor activity were assessed using the peripherally active NAChR antagonist hexamethonium, the centrally active noncompetitive NAChR antagonist mecamylamine (Martin et al., 1993; Varanda et al., 1985) and the centrally active competitive NAChR antagonist DHβE (Williams and Robinson, 1984; Valera et al., 1992). Finally, the role of the dopaminergic system in the stimulant effects induced by nicotine and SIB-1765F was investigated using the D1 and D2 dopamine receptor antagonists SCH 23390 and eticlopride, respectively.
Materials and Methods
Animals.
Male Sprague-Dawley rats (200–350 g) (Harlan, San Diego, CA) were housed four per cage and maintained in a humidity- (50–55%) and temperature- (22–24°C) controlled facility on a 12 hr:12 hr light/dark cycle (lights on at 6:30 a.m.) with free access to food (Harlan-Teklad 4% rat diet 7001) and water. Rats were allowed a 1-wk period of habituation to the animal room before testing. The animals were handled once during this period.
Compounds.
Mecamylamine hydrochloride, (-)-nicotine hydrogen tartrate and hexamethonium bromide were obtained from Sigma Chemical Co. (St. Louis, MO). DHβE, (±)-epibatidine di-hydrochloride, S-(-)-eticlopride hydrochloride and R(+)-SCH 23390 hydrochloride were purchased from Research Biochemicals International (RBI, Natick, MA). SIB-1765F was synthesized at SIBIA as per methods previously described (Cosford et al., 1996). Nicotine and SIB-1765F were dissolved in saline and pH was adjusted to 7.0 by addition of 10N NaOH. DHβE, epibatidine, eticlopride, mecamylamine, hexamethonium and SCH 23390 were dissolved in saline. Doses of epibatidine and nicotine are expressed in terms of their free base concentrations. All compounds were administered s.c. into the dorsal neck region in a volume of 1 ml/kg. Saline was used as control.
Locomotor activity.
Locomotor activity was assessed in photocell activity cages (San Diego Instruments, San Diego, CA). Each cage consisted of a standard plastic rodent cage (24 × 45.5 cm) surrounded by a stainless steel frame. Four infrared photocell beams were located across the long axis of the frame, raised 3.2 cm above the floor and spaced 9 cm apart. The number of cage crosses (crossovers,i.e., consecutive interruptions of one beam followed immediately by interruption of an adjacent beam) was recorded and used as a measure of locomotion. The number of crossovers was recorded by a computer system during consecutive 5-min intervals.
Decreases in locomotor activity were evaluated in rats naive to the test apparatus (hereafter referred to as naive rats). Rats received injections with test compound or vehicle and were placed in the photocell activity cages 5 min after injection. Locomotor activity was recorded for a 30-min period.
Increases in locomotor activity were evaluated in rats previously habituated to the test apparatus (hereafter referred to as habituated rats). Rats were habituated to the photocell activity cages for 180 min, 24 hr before testing. This habituation period was required to overcome the potentially stressful nature of the test apparatus and, therefore, reduce the basal levels of activity of the rats. On the test day, the rats were placed in the photocell activity cages for a habituation period of 90 min, after which they were removed, injected with the test compound and were returned to the cages to be monitored for 120 to 360 min.
In studies evaluating the effects of NAChR antagonists on NAChR agonists-induced decreases in locomotor activity, naive rats were administered with antagonists 15 min before the injection of NAChR agonists. After administration of NAChR agonists, locomotor activity was recorded for 30 min.
In studies evaluating the effects of antagonists on NAChR agonists-induced increases in locomotor activity, habituated rats were administered with nicotinic or dopaminergic receptor antagonists 15 or 30 min before the injection of NAChR agonists, respectively. After administration of NAChR agonists, activity was recorded for 90 min. Nicotinic and dopaminergic receptor antagonists were also tested alone in a novel environment to determine potential intrinsic sedative or depressant effects. Naive rats received injections in their home cages and were placed in the photocell activity cages 15 min after injection of NAChR antagonists and 30 min after injection of DA receptor antagonists. Locomotor activity was recorded for 30 min.
These studies were based on independent subject design, with each subject used only once. Twelve rats (individually caged) were tested at one time. Testing was carried out between 8:00 a.m. and 5:30 p.m. each day (light cycle).
Statistics.
Results were analyzed by Student’s ttest or by one- and two-factor analyses of variance, with repeated measure methods where appropriate (SigmaStat Software, Jandel, San Rafael, CA). Dunnett’s test and Newman-Keul’s test were used forpost hoc comparisons. P < .05 was considered significant.
Results
Effects of NAChR agonists on locomotor activity in naive rats.
When administered to naive rats, the three NAChR agonists, SIB-1765F, nicotine and epibatidine, decreased locomotor activity as illustrated by a decrease in the number of crossovers (fig. 1, A-C, left panels). SIB-1765F and epibatidine reduced locomotor activity in a dose-related manner [SIB-1765F: F(4, 25) = 5.53, P = .0025; epibatidine: F(4, 25) = 18.0, P < .0001] whereas nicotine decreased locomotor activity in a non-dose-related manner during the first 10 min of the session [F(4, 25) = 6.60, P = .0009] (fig.1).

Decreases in locomotor activity induced by SIB-1765F (A), nicotine (B) and epibatidine (C) in rats not previously exposed to the test apparatus (naive rats). Left panel, Dose-related effects presented as total crossovers over a 10-min period beginning 5 min after administration of the compounds (mean ± S.E.M.,n = 6/group). *P < .05 vsvehicle-treated group, Dunnett’s test. Right panel, Time course of effects presented as crossovers taken at 5-min intervals beginning 5 min after the administration of the compounds (n = 6/group). Each point represents mean ± S.E.M. *P < .05vs vehicle-treated group, Newman-Keul’s test.
Figure 1 (right panels) also shows the time course for the effect of each NAChR agonist over the entire session (30 min). For clarity, only one dose of each compound is shown and compared to its respective vehicle. There was a significant decline in activity for each vehicle-treated group as the animals became habituated to the test environment. SIB-1765F at a dose of 20 mg/kg produced a depressant effect that decreased over time [SIB-1765F × time interaction: F(5, 50) = 16. 78, P < .0001] (fig. 1A, right panel). This transient decrease in locomotor activity was followed by a small increase in locomotor activity. However, this increase in activity was not significantly different from the activity of vehicle-treated animals.
Nicotine and epibatidine produced similar transient decreases in locomotor activity followed by mild increases in locomotor activity [nicotine × time interaction: F(5, 50) = 11.62, P < .0001; epibatidine × time interaction: F(5, 50) = 39.42, P < .0001] (fig. 1, B-C, right panels). The magnitudes of the depressant effects were similar for the three NAChR agonists.
Effects of NAChR antagonists on NAChR agonists-induced decreases in locomotor activity in naive rats.
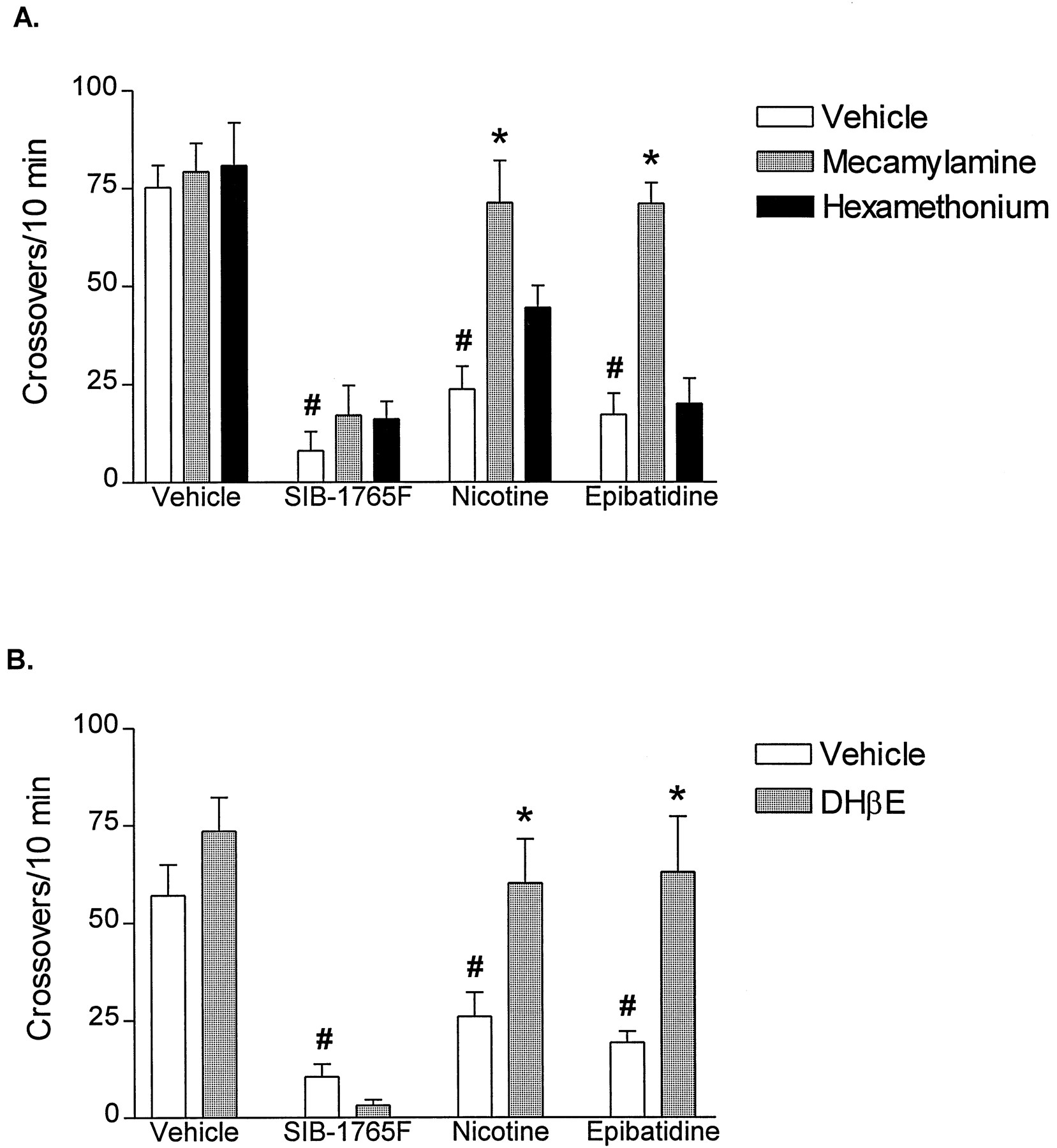
Figure 2 shows the effects of mecamylamine, hexamethonium and DHβE on the decrease in locomotor activity induced by a single dose of each NAChR agonist in naive rats. The NAChR antagonists had no significant effect on locomotor activity when administered alone (fig. 2A-B). Mecamylamine (3 mg/kg) blocked the effects induced by nicotine (0.4 mg/kg) [interaction nicotine × mecamylamine: F(1, 20) = 8.10, P < .009] and epibatidine (3 μg/kg) [interaction epibatidine × mecamylamine: F(1,20) = 17.3, P < .0005] but did not attenuate the effect induced by SIB-1765F (20 mg/kg) [interaction SIB-1765F × mecamylamine: F(1,20) = .14, P = .71] (fig. 2A). Similarly, DHβE (6 mg/kg) blocked the locomotor depressant effects induced by nicotine and epibatidine but failed to attenuate the locomotor depressant effect induced by SIB-1765F (fig.2B). Hexamethonium (10 mg/kg) had no effect on the decrease in locomotor activity induced by nicotine, epibatidine or SIB-1765F (fig.2A).

Effects of mecamylamine (3 mg/kg) (A), hexamethonium (10 mg/kg) (A) and DHβE (6 mg/kg) (B) on NAChR-agonists induced decreases in locomotor activity in naive rats. The NAChR antagonists were administered 15 min prior to SIB-1765F (20 mg/kg), nicotine (0.4 mg/kg) and epibatidine (3 μg/kg). Data are represented as mean total crossovers over a 10-min period beginning 5 min after the injection of the agonists (mean ± S.E.M., n = 6). #P < .05 vs vehicle/vehicle-treated group, *P < .05 vs vehicle/NAChR agonist-treated group, Newman-Keul’s test.
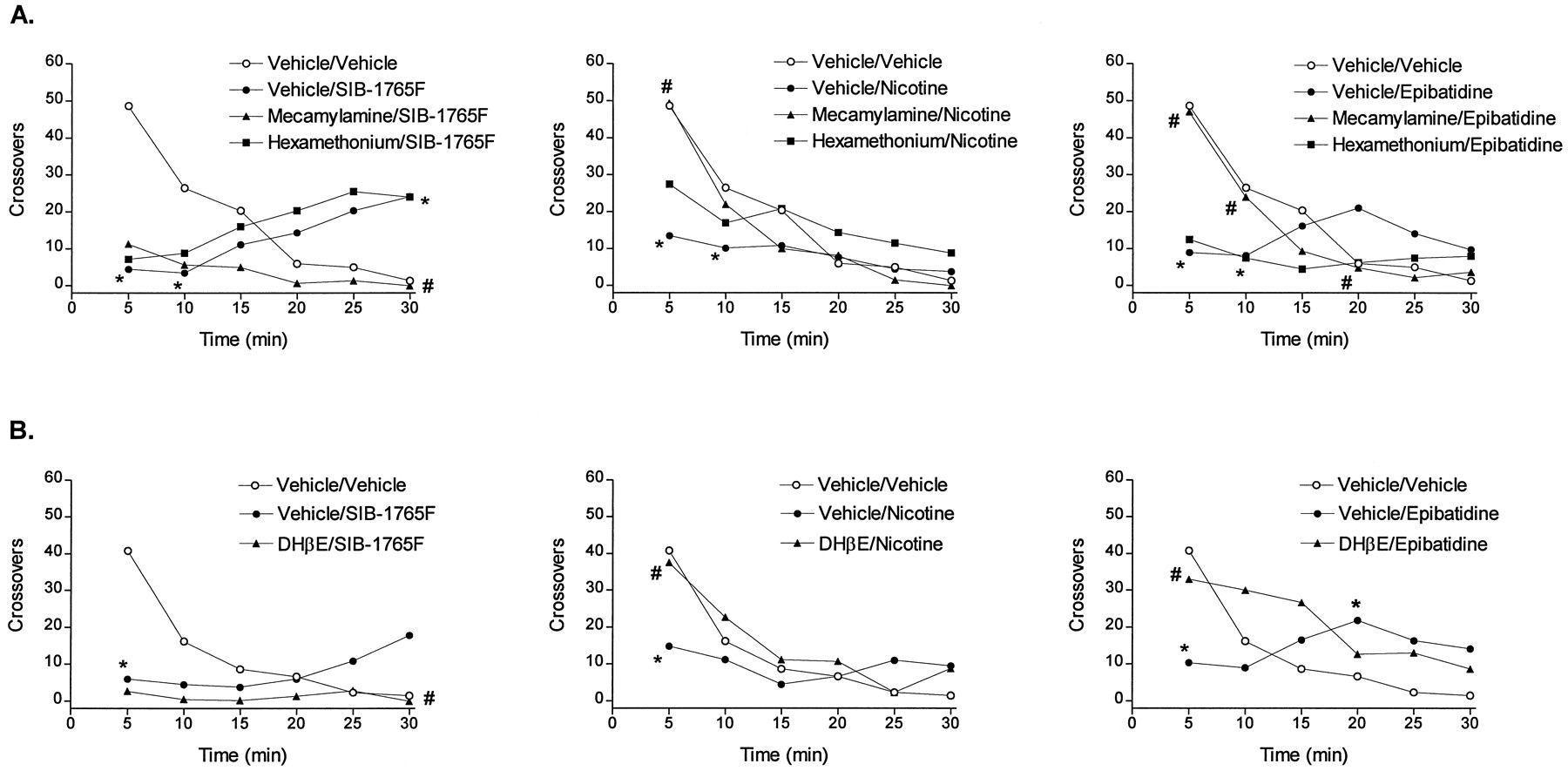
Analysis of the time courses of the effects of mecamylamine and DHβE over a period of 30 min revealed that these centrally acting NAChR antagonists failed to block the decrease in locomotor activity induced by the administration of a 20-mg/kg dose of SIB-1765F, but did block the mild increase in locomotor activity that followed (fig.3, A-B, left panels). Under identical conditions, the decreases in locomotor activity induced by nicotine and epibatidine were both attenuated by mecamylamine and DHβE and were insensitive to hexamethonium (fig. 3, A-B, center and right panels).

Time courses of the antagonism by mecamylamine (3 mg/kg) and hexamethonium (10 mg/kg) (A) and DHβE (6 mg/kg) (B) of the locomotor effects induced by NAChR agonists in naive rats. Data are presented as average crossovers taken at 5-min intervals beginning 5 min after the administration of SIB-1765F (20 mg/kg), nicotine (0.4 mg/kg) and epibatidine (3 μg/kg) (n = 6/group). Doses used were chosen on the basis of equal efficacy. S.E.M. were not shown for the clarity of the graphs. *P < .05 vsvehicle/vehicle-treated groups, #P < .05 vsvehicle/NAChR agonist, Newman Keul’s test.
Effects of NAChR agonists on locomotor activity in habituated rats.
Figure 4 shows the effects of the three NAChR agonists after administration to rats habituated to the photocell activity cages. There was a significant difference between the levels of activity of vehicle-treated naive and habituated rats over the first 30 min of exposure to the cages (compare fig. 1A vs fig. 4A: right panels). Habituated rats had a statistically significant lower level of activity than naive rats (total crossovers/30 min: naive rats: 86.2 ± 9.4, habituated rats: 31.5 ± 5.7; P < .05 Student’s t test). In habituated rats, s.c. administration of SIB-1765F increased the number of crossovers in a dose-related manner [F(5, 41) = 20.4, P < .0001], with the maximum effect occurring with a dose of 40 mg/kg (P < .05) (fig. 4A, left panel). Locomotor activity after administration of the 40 mg/kg dose of SIB-1765F was nearly 15 times greater than that after administration of vehicle, over the 120-min period.

Increases in locomotor activity induced by SIB-1765F (A), nicotine (B) and epibatidine (C) in rats habituated to the test cages. Left panel, Dose-related effects presented as total crossovers over a 120-min period beginning immediately after the administration of the compounds (mean ± S.E.M.,n = 7–8/group). *P < .05 vsvehicle-treated group, Dunnett’s test. Right panel, Time courses of effects presented as crossovers taken at 5-min intervals beginning immediately after the administration of the compounds (n = 7–8/group). Each point represents mean ± S.E.M. The indications of statistical significance have been omitted to enhance visual clarity.
Nicotine also increased locomotor activity in a dose-dependent manner [F(5,40) = 4.28, P < .003] but the maximum effect was never more than 3.5 times that of vehicle (fig. 4B, left panel). Epibatidine slightly increased locomotor activity [F(5,40) = 3.58, P < .009], but the total number of crossovers produced by the most effective dose (3 μg/kg) was less than twice that of vehicle (fig.4C, left panel).
Figure 4 also shows the time courses for the effects of the NAChR agonists on locomotor activity. A decline in crossovers for both vehicle- and drug-treated rats was observed as the session proceeded. In the study of SIB-1765F (20 mg/kg), there was a main effect of the compound [F(1, 14) = 281.5, P < .0001], a main effect of time [F(23, 322) = 2.98, P < .0001] and a significant interaction between the two factors [F(23, 322) = 2.14, P = .0021] (fig. 4, A-C, right panels). The increase in activity induced by SIB-1765F at doses of 20 and 40 mg/kg was observed within 5 to 15 min and lasted for 2.2 and 3.5 hr, respectively (fig. 5). The increase in locomotor activity induced by nicotine at its maximally effective dose (0.4 mg/kg) was observed within 5 min and lasted no more than 80 min after administration [nicotine effect: F(1, 14) = 20.42, P < .0006; time effect: F(23, 299) = 8.23, P < .0001; and no interaction nicotine × time: F(23, 299) = 1.11, P = .33] (fig. 4B, right panel). Epibatidine, at a dose of 3 μg/kg, increased locomotor activity within 20 min post-injection and the effect lasted for less than 50 min [epibatidine effect: F(1, 15) = 5.40, P = .03; time effect: F(23, 345) = 5.58, P < .0001; epibatidine × time interaction: F(23, 345) = 1.66, P = .03] (fig. 4C, right panel).

Duration of the locomotor activity induced by SIB-1765F in rats habituated to the test cages. Data are presented as average of crossovers taken at 5-min intervals beginning immediately after the administration of the compound (n = 8/group). S.E.M. were not shown for the clarity of the graph.
Effects of NAChR antagonists on NAChR agonists-induced increases in locomotor activity in habituated rats.
Pretreatment with mecamylamine (3 mg/kg) or DHβE (6 mg/kg) significantly attenuated the locomotor activity induced by SIB-1765F (20 mg/kg) [mecamylamine × SIB-1765F interaction: F(1,20) = 9.85, P = .005; DHβE × SIB-1765F interaction: F(1,20) = 6.63, P = .01] (fig.6A, left and right panels). In contrast, hexamethonium (10 mg/kg) had no effect on locomotor activity induced by SIB-1765F [SIB-1765F × hexamethonium interaction: F(1,20) = .46, P = .50] (fig. 6A, center panel). The locomotor activity induced by nicotine (0.4 mg/kg) was also blocked by mecamylamine and DHβE as shown by a significant agonist and antagonist interaction [nicotine × mecamylamine: F(1,28) = 20.6, P < .0001 nicotine × DHβE: F(1,20) = 6.34, P = .02] and was insensitive to hexamethonium [nicotine × hexamethonium: F(1,27) = 1.69, P = .20] (fig. 6B). Epibatidine- (3 μg/kg) induced locomotor activity was blocked by mecamylamine [epibatidine × mecamylamine: F(1,20) = 36.0, P < .0001] but was insensitive to DHβE [epibatidine × DHβE: F(1,34) = .04, P = .85] or hexamethonium [epibatidine × hexamethonium: F(1,20) = 2.81, P = .11] (fig. 6C).

Effects of mecamylamine, hexamethonium and DHβE on the locomotor activity induced by SIB-1765F (20 mg/kg) (A), nicotine (0.4 mg/kg) (B) and epibatidine (3 μg/kg) (C) in rats habituated to the test cages. The antagonists were administered 15 min before the NAChR agonists. Data are represented as mean total counts taken over a 90-min period after the injection of vehicle (white bar) or NAChR agonists (gray bar) (mean ± S.E.M., n = 6–9). #P < .05 vs vehicle/vehicle-treated group, *P < .05 vs vehicle/NAChR agonist-treated group, Newman-Keul’s test.
Effect of selective D1 and D2 DA receptor antagonists on NAChR agonists-induced locomotor activity in habituated rats.
To determine if the locomotor activity induced by SIB-1765F and nicotine were mediated through the activation of the dopaminergic system, the selective D1 receptor antagonist SCH 23390 and the selective D2 receptor antagonist eticlopride were administered before the test compounds.
SCH 23390 reduced SIB-1765F-induced locomotor activity in a dose-dependent manner [SCH 23390 × SIB-1765F interaction: F(3,40) = 13.7, P < .0001] (fig. 7A, left panel). SCH 23390 also attenuated the locomotor activity induced by nicotine [SCH 23390 × nicotine interaction: F(3,40) = 4.14, P = .01] (fig. 7B, left panel). SCH 23390 did not have a significant intrinsic effect on activity when administered to habituated rats; however, it reduced spontaneous locomotor activity in experimentally naive rats in a dose-dependent manner [SCH 23390: F(3,20) = 9.82, P = .0003] (fig. 8A, left panel). Analysis of the time course showed a main effect of SCH 23390 [F(3,20) = 6.36, P < .003], a main effect of time [F(5,143) = 47.93, P < .0001] and a significant interaction between the two factors [F(15,143) = 5.66, P < .0001] (fig. 8A, right panel).

Effects of SCH 23390 and eticlopride on the locomotor activity induced by SIB-1765F (20 mg/kg) (A) and nicotine (0.4 mg/kg) (B) in rats habituated to the test cages. SCH 23390 and eticlopride were administered 30 min before the NAChR agonists. Data are presented as crossovers taken over a 90-min period after the injection of saline (white bar) or NAChR agonists (gray bar) (mean ± S.E.M., n = 6). #P < .05 vsvehicle/vehicle-treated group, *P < .05 vsvehicle/NAChR agonist-treated group, Newman-Keul’s test.
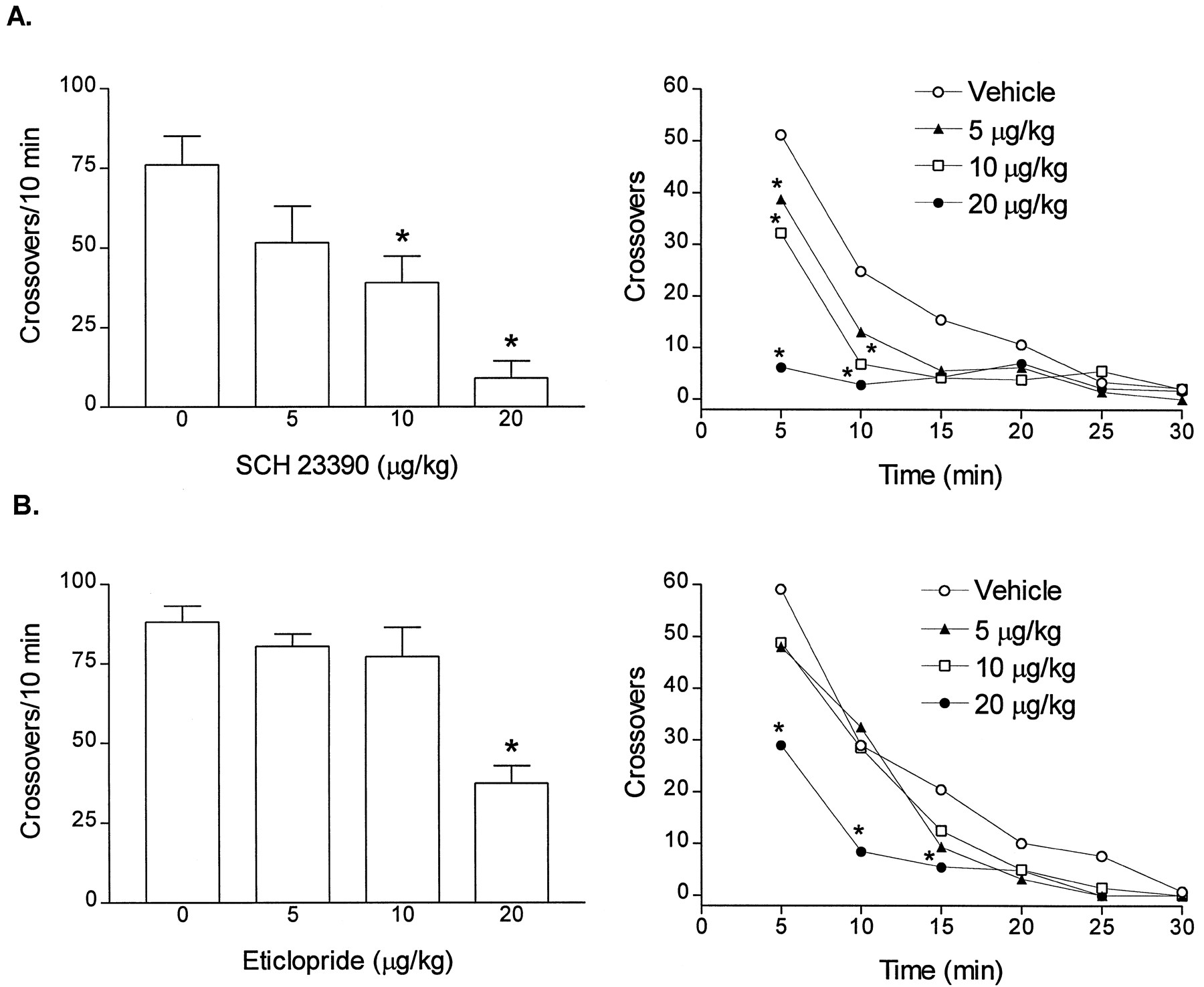
Effects of SCH 23390 (A) and eticlopride (B) on locomotor activity of naive rats. Left panel, Dose-related effects presented as total crossovers taken over a 10-min period beginning 30 min after the administration of the compounds (mean ± S.E.M.,n = 6/group). *P < .05 vsvehicle-treated group, Dunnett’s test. Right panel, Time course of effects presented as average crossovers taken at 5-min intervals beginning 30 min after the administration of the compounds (n = 6/group). S.E.M. were not shown for the clarity of the graph. *P < .05 vs vehicle-treated group.
The selective D2 receptor antagonist eticlopride also produced a significant reduction in the effect of SIB-1765F on locomotor activity [eticlopride × SIB-1765F interaction: F(3,40) = 14.9, P < .0001] (fig. 7A, right panel). Analysis of the time course showed a main effect of eticlopride [F(3,20) = 12.63, P < .0001], a main effect of time [F(5,143) alone = 112.68, P < .0001], and a significant interaction between the two factors [F(15,143) = 2.95, P < .0007] (fig. 8B, right panel). Low doses of eticlopride (5–10 μg/kg) that had no significant effect on spontaneous activity when tested in naive rats (fig. 8B) did attenuate SIB-1765F-induced locomotor activity (fig. 7A: right panel). Eticlopride at a 20-μg/kg dose also attenuated the effect of nicotine on locomotor activity although there was no significant interaction between the drugs [eticlopride effect: F(3,40) = 2.79, P = .05; nicotine effect: F(1,40) = 23.76, P < .0001; eticlopride × nicotine interaction: F(3,40) = 2.07, P = .12] (fig. 7B, right panel).
Discussion
Depressant or stimulant locomotor effects induced by NAChR agonists can be revealed by exposing rats to a novel or a familiar testing environment. Acute s.c. administration of nicotine, epibatidine or the novel NAChR agonist SIB-1765F decreased locomotor activity in experimentally naive rats and increased locomotor activity in rats habituated to the test apparatus. The decreases in locomotor activity induced by these three NAChR agonists were quantitatively similar. In contrast, the three NAChR agonists differ considerably in their stimulatory effects, with SIB-1765F producing a greater increase in locomotor activity than nicotine or epibatidine.
The decreases in locomotor activity observed after administration of SIB-1765F, nicotine and epibatidine in experimentally naive rats are similar to those observed previously with other NAChR agonists (Reavillet al., 1990; Garcha et al., 1993; Stolermanet al., 1995). These effects have a short duration (<15 min).
The noncompetitive NAChR antagonist mecamylamine (Varanda et al., 1985; Martin et al., 1993) and the competitive NAChR antagonist DHβE (Reavill et al., 1988, Williams and Robinson, 1984; Valera et al., 1992, Damaj et al., 1995) completely reversed the locomotor depressant effects induced by nicotine and epibatidine, implying that these effects were mediated through the activation of NAChR. The peripherally active NAChR antagonist hexamethonium failed to block the decrease in locomotor activity induced by nicotine or epibatidine. Thus, the decreases in locomotor activity induced by nicotine and epibatidine appear to be mediated through the activation of central and not peripheral NAChR. Interestingly, hexamethonium, mecamylamine and DHβE failed to block the decrease in locomotor activity produced by SIB-1765F in naive rats. This suggests that SIB-1765F acts on NAChR subtype(s) insensitive to mecamylamine and DHβE or reduces locomotor activity by a nonnicotinic mechanism. Mecamylamine also failed to block the decrease in locomotor activity induced by the NAChR agonists lobeline and anatoxin in naive rats (Stolerman et al., 1992, 1995). Therefore, the depressant effect induced by SIB-1765F in experimentally naive rats is similar in magnitude to those induced by nicotine and epibatidine, but appears to be mediated through a different mechanism.
The locomotor stimulatory effects induced by SIB-1765F and nicotine in habituated rats were blocked by mecamylamine and DHβE but not by hexamethonium, indicating that these NAChR agonists elicit their stimulant effects predominantly through the activation of central NAChR. This contrasts with the lack of effect of mecamylamine and DHβE on SIB-1765F-induced decrease in locomotor activity in experimentally naive rats, and suggests that the stimulant and depressant effects of SIB-1765F are likely to be mediated through different mechanisms. Moreover, epibatidine-induced increase in locomotor activity was attenuated by mecamylamine but not by DHβE whereas epibatidine-induced decrease in locomotor activity was attenuated by both centrally active NAChR antagonists. These data suggest that the depressant and stimulant effects of epibatidine are also likely to be mediated through different mechanisms.
NAChR agonists are known to increase locomotor activity through an interaction with the dopaminergic system (Reavill and Stolerman, 1990;Museo and Wise, 1990; Imperato et al., 1986). Therefore, the effect of DA antagonists on the increases in locomotor activity induced by nicotine and SIB-1765F were evaluated. SCH 23390, a selective D1 DA receptor antagonist, produced a dose-dependent decrease in spontaneous locomotor activity in naive animals, but did not affect locomotor activity in habituated animals. These results contradict a previous report from O’Neill et al. (1991) showing that SCH 23390 at doses five times higher than those used in our study failed to attenuate spontaneous locomotor activity in naive rats. In habituated rats, SCH 23390 attenuated SIB-1765F induced locomotor activity at doses that did not alter spontaneous locomotor activity in naive rats, suggesting that D1 receptor activation may in part contribute to the stimulant effect of SIB-1765F. Previously, SCH 23390 was shown to reduce the locomotor stimulant effects induced by nicotine and epibatidine (O’Neill et al., 1991; Sacaan et al., 1996). The D2 DA receptor antagonist eticlopride also attenuated the increase in locomotor activity induced by SIB-1765F at doses (5 and 10 μg/kg) that did not significantly reduce spontaneous activity in naive rats. Nicotine-induced locomotor activity was blocked by a 20-μg/kg dose of eticlopride that also attenuated spontaneous activity in naive rats. Nicotine-induced locomotor activity is known to be sensitive to other D2 antagonists such as raclopride (O’Neillet al., 1991) and haloperidol (Arnold et al., 1995). We recently reported that the stimulant effect of epibatidine was also attenuated by eticlopride (Sacaan et al., 1996). These data provide additional evidence that the acute increases in locomotor activity induced by NAChR agonists are mediated, at least in part, through the activation of both D1 and D2 DA receptors subsequent to the release of DA.
SIB-1765F had substantial effects on locomotor activity in habituated rats as compared to nicotine or epibatidine. Over a period of 90 min, a maximally effective dose of SIB-1765F was approximately four times more efficacious and lasted two to three times longer than nicotine or epibatidine in increasing locomotor activity. In vitrostudies have shown that SIB-1765F releases [3H]-DA from rat striatal and olfactory tubercle slices, regions containing terminal fields of the nigrostriatal and mesolimbic dopaminergic pathways, respectively (Rao et al., 1995; Sacaan et al., accompanying manuscript). In these assays, SIB-1765F tends to be more effective than nicotine at releasing [3H]-DA. Microdialysis studies also indicate that SIB-1765F is considerably more efficacious than nicotine at releasing DA from both rat striatum and nucleus accumbens in vivo (Menzaghi et al., 1995; Sacaan et al., accompanying manuscript). Furthermore, SIB-1765F is more efficacious than nicotine and epibatidine at inducing ipsilateral turning in unilaterally 6-OHDA-lesioned rats (Lloydet al., 1995; Cosford et al., 1996). Thus, the greater effect of SIB-1765F on DA release in vivo, as compared to nicotine, may be, at least in part, responsible for a greater increase in locomotor activity. In addition, activation of different NAChR subtypes and differences in pharmacokinetics may also contribute to these differences in profiles.
In summary, these results demonstrate that the novel NAChR agonist SIB-1765F both increased and decreased spontaneous locomotor activity in rats depending on the experimental paradigm. These effects can be demonstrated by testing animals in a novel or a familiar environment. Similar effects were seen with the NAChR agonists nicotine and epibatidine. Interestingly, the decreases in locomotor activity induced by these three nicotinic agonists were similar in magnitude but appeared to be mediated through different mechanisms. Furthermore, our data suggest that the stimulant and depressant effects induced by SIB-1765F or epibatidine are likely to be mediated through different mechanisms. SIB-1765F produced a larger and longer-lasting increase in locomotor activity. This effect is dependent on D1 and D2 DA receptor activation subsequent to DA release.
Footnotes
-
Send reprint requests to: Dr. Frédérique Menzaghi, SIBIA Neurosciences, Inc., 505 Coast Boulevard South, Suite 300, La Jolla, CA 92307-4641.
- Abbreviations:
- DA
- dopamine
- DHβE
- dihydro-β-erythroidine
- NAChR
- nicotinic acetylcholine receptor(s)
- NE
- norepinephrine
- SIB-1765F ([±]-5-ethynyl-3-(1-methyl-2-pyrrolidinyl)pyridine fumarate).
- Received April 26, 1996.
- Accepted August 8, 1996.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}