


Abstract
By means of the expression of two chimeric receptors, α2/M3 and M3/α2, in which the carboxy-terminal receptor portions, containing transmembrane domains VI and VII, were exchanged between the α2C-adrenergic and the M3 muscarinic receptor, it has been shown that G protein-coupled receptors are able to interact functionally with each other at the molecular level to form (hetero)dimers. In the present study, we tested the hypothesis that interaction between two different muscarinic receptor subtypes can lead to the formation of a heterodimeric muscarinic receptor with a new pharmacological profile. Initially, muscarinic M2 or M3 wild-type receptors were expressed together with gene fragments originating from M3 or M2 receptors, respectively. Antagonist binding, performed with pirenzepine and tripitramine, revealed the presence of two populations of binding sites: one represents the wild-type M2 or M3receptors, the other the heterodimeric M2/M3receptor. In another set of experiments, we constructed a point mutant M2 receptor M2 (Asn404→Ser), in which asparagine 404 was replaced by serine. Although this receptor alone did not show any binding forN-[3H]methylscopolamine (up to 2 nM), when cotransfected with M3, it resulted in the rescue of a high-affinity binding for tripitramine. These findings demonstrate that M2 and M3 muscarinic receptor subtypes can cross-interact with each other and form a new pharmacological heterodimeric receptor.
G protein-coupled receptors are transmembrane proteins that mediate a variety of signaling processes, such as neurotransmission, hormonal response, olfaction, and light transduction. Muscarinic receptors are members of this family and molecular cloning has revealed the existence of five different subtypes (Kubo et al., 1986; Bonner et al., 1987,1988; Peralta et al., 1987) that show a high degree of sequence homology but differ in their ligand binding and functional properties, as well as in their tissue distribution (Hulme et al., 1990). They are predicted to be composed of seven hydrophobic transmembrane domains (TMD; I-VII) connected by alternating cytoplasmic and extracellular loops, an extracellular amino-terminal domain, and an intracellular carboxy-terminal segment. Although they are generally considered closely packed structures, an increasing amount of evidence indicates that they can behave structurally in a fashion analogous to multiple subunit receptors. In previous experiments, Maggio et al. (1993a)showed that when truncated M2 or M3 receptors (containing TMDs I-V, named M2- or M3-trunc) were coexpressed in African green monkey kidney (COS-7) cells with gene fragments coding for the corresponding carboxy-terminal receptor portions (containing TMDs VI and VII, named M2- or M3-tail), muscarinic receptors with ligand-binding properties and functional activity similar to the wild-type receptors were obtained. Because the separate transfection of each fragment individually was not able to show any binding, it was supposed that the two receptor domains interact with each other to form a macromolecular complex with the same characteristics as the wild-type receptor. Similar results have been obtained with bacteriorhodopsin (Kahn and Engelman, 1992), β2-adrenergic (Kobilka et al., 1988), rhodopsin (Ridge et al., 1995), V2 vasopressin (Schöneberg et al., 1996), and D2 dopamine receptors (Barbier et al., 1996). Furthermore, experiments performed with the M3muscarinic receptor have shown that it can be split not only at the level of the third cytoplasmic loop (i3 loop), but also in the second intra- and third extracytoplasmic loops, and that it can retain its binding properties (Schöneberg et al., 1995).
As all muscarinic receptors have a long i3 loop that connects TMDs V to VI, it has been speculated that wild-type receptors can actually express the domains constituted by the amino- (containing TMDs I–V) and carboxyl-terminal receptor portions (containing TMDs VI and VII) as separate units capable of interacting with each other. Furthermore, this interaction was thought to occur not only intra- but also intermolecularly. This was demonstrated by creating two chimeric receptor molecules, α2/M3and M3/α2, in which the carboxy-terminal receptor portions (including TMDs VI and VII) were exchanged between the α2C-adrenergic and the M3 muscarinic receptors (Maggio et al., 1993b). Although transfection of the two chimeric constructs alone into COS-7 cells did not result in any detectable binding activity, coexpression of the two mutant receptors resulted in a significant number of specific-binding sites for the muscarinic ligandN-[3H]methylscopolamine ([3H]NM) and the adrenergic ligand [3H]rauwolscine. Other experiments performed with functionally impaired muscarinic receptors confirmed these findings (Maggio et al., 1993b).
In a later article, Maggio et al. (1996) demonstrated that receptor interaction is prevented by the shortening of the i3 loop. Chimeric α2/M3 and M3/α2 receptors with the deletion of a large portion of the i3 loop were no longer able to cross-interact and bind M3 muscarinic and α2-adrenergic receptor ligands when coexpressed in the same cells. Furthermore, although the activity of a functionally impaired M3 muscarinic receptor mutant in which 16 amino acids of the amino-terminal part of the i3 loop were substituted with the corresponding M2 sequence [M3/M2(16aa)] was rescued by coexpression with a healthy M3-trunc fragment bearing the correct sequence, the deletion of 196 amino acids from the i3 loop of M3/M2(16aa), although leaving intact the binding characteristics of the receptor, completely prevented the functional rescue operated by M3-trunc. Experiments performed with a short form of the wild-type M3 muscarinic receptor (M3-short), in which 196 amino acids of the i3 loop had been removed, demonstrated that the large deletion of the i3 loop leaves unvaried the binding characteristics of the receptor, the phosphatidyl inositol hydrolysis activity, and the ability of the receptor to internalize.
The property of receptors to cross-interact at the molecular level and the ability of cotransfected fragments to form a macromolecular receptor complex capable of binding with high-affinity muscarinic ligands leads to the hypothesis that when different subtypes of muscarinic receptors are coexpressed in the same cells, they might interact at the molecular level and form a new pharmacological receptor. In agreement with this view, the cotransfection of M2-trunc with M3-tail results in a fragmented chimeric M2-trunc/M3-tail receptor capable of binding muscarinic ligands with a high affinity (the other chimeric combination, M3-trunc + M2-tail did not show any binding; Maggio et al., 1993a). Barbier et al. (1998) studied the antagonist binding characteristics of the chimeric M2-trunc/M3-tail receptor with the purpose of identifying compounds capable of discriminating between this chimera and the wild-type M2 and M3 muscarinic receptors. The pharmacological profile of this fragmented chimeric receptor was clearly different from that of M2 and M3 wild-type receptors; whereas most of the compounds tested showed an intermediate affinity between M2 and M3, tripitramine, and pirenzepine showed a peculiar pharmacological profile. Tripitramine, a selective antagonist for the M2 receptor (Maggio et al., 1994), also displayed a high-affinity binding for the chimeric M2-trunc/M3-tail receptor that was more than 100-fold higher than that found for the M3 muscarinic receptor. Pirenzepine, on the other hand, had a 12- and 3-fold higher affinity for the chimeric receptor than for the M2 and M3wild-type receptors, respectively. In the present work, using the unique pharmacological profile of M2-trunc/M3-tail chimeric receptor, we provide evidence that the exchange of the amino- and carboxy-terminal receptor domains between the wild-type M2 and M3 receptors leads to the formation of a heterodimeric M2-trunc/M3-tail muscarinic receptor.
Materials and Methods
Preparation of Mutant Receptor Constructs.
HM2pcD and RM3pcD (Bonner et al., 1987), two mammalian expression vectors containing the entire coding sequence of the human M2 and the rat M3 muscarinic receptors, were used to construct the various fragmented (M2-trunc and M3-tail) and mutant [M3-short and M2 (Asn404 → Ser)] muscarinic receptor genes.
pcDM2-Trunc and pcDM3-Tail.
The construction of these fragmented receptors has been described previously (Maggio et al., 1993a). The encoded M2-trunc receptor contains an in-frame stop codon after the amino acid codon Ser-283 of the human M2 sequence, whereas M3-tail codes for the 202 carboxy-terminal amino acids (from Leu-388 to Leu-589) of the rat M3 muscarinic receptor (Fig. 1). When transfected together to allow an efficient coexpression of the two muscarinic receptor fragments, a plasmid with the two transcriptional units (pcDM2-trunc/M3-tail) was used. The pcDM2-trunc/M3-tail plasmid contains the simian virus 40 early region promoter, the M3-tail coding sequence, and the segment carrying the simian virus 40 late region polyadenylation signal from pcDM3-tail cloned into the blunt-ended SalI site of the pcDM2-trunc.

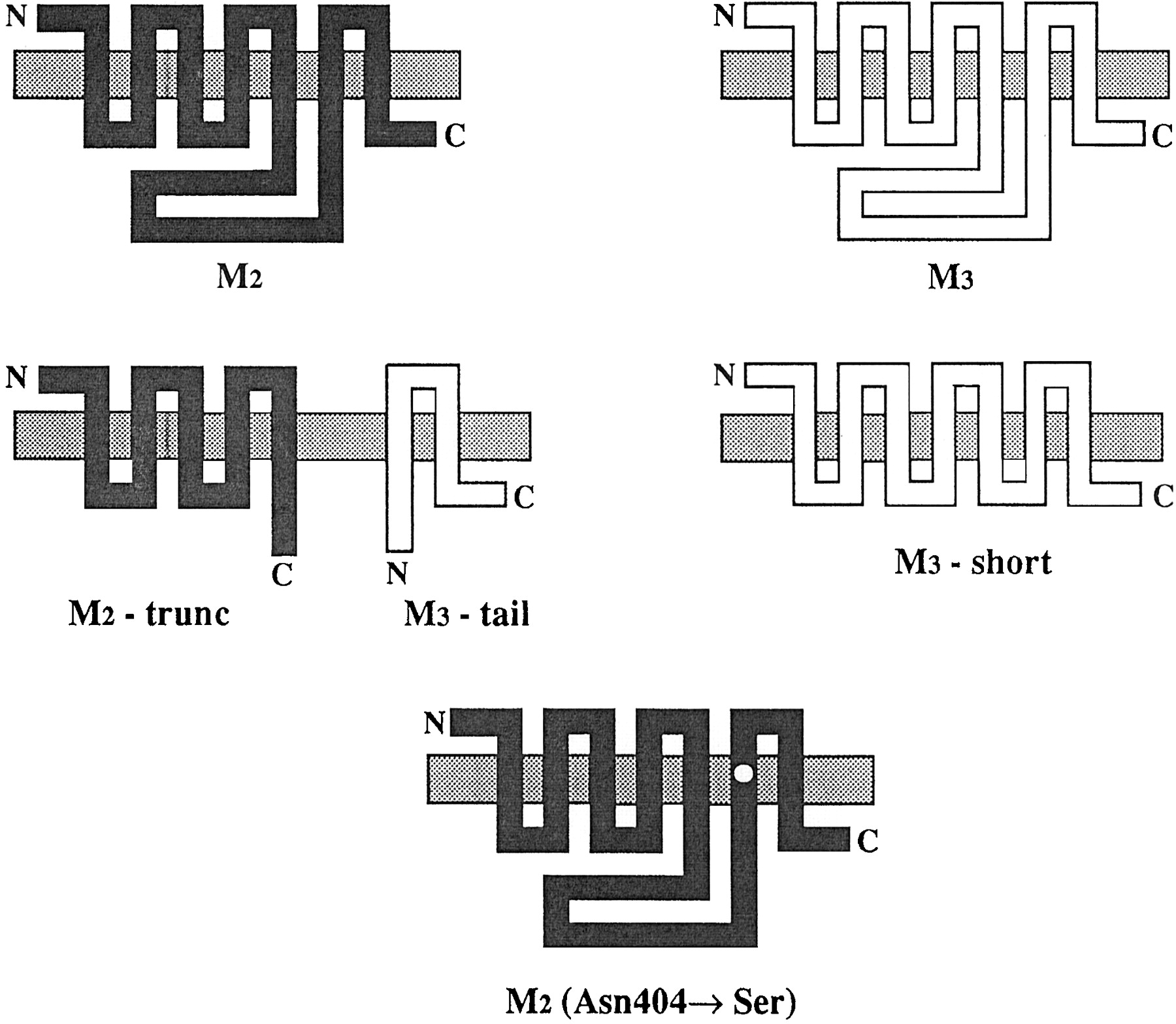
Schematic representation of the wild-type human M2 and rat M3, the fragments M2-trunc and M3-tail, and the mutants M3-short and M2 (Asn404 → Ser) muscarinic receptors. The truncated fragment, M2-trunc, contains the amino-terminal domain, the first five hydrophobic transmembrane regions and the initial portion (56 amino acids) of the i3 loop of the wild-type muscarinic M2 receptor. The M3-tail fragment contains the final portion of the i3 loop (105 amino acids), the last two hydrophobic transmembrane regions, and the carboxy-terminal segment of the wild-type M3 muscarinic receptor. The short construct (M3-short) represents a receptor in which 196 amino acids of the i3 loop have been deleted; the remaining i3 loop is 43 amino acids long. The point mutant M2 (Asn404 → Ser) has the asparagine 404 replaced with serine.
pcDM3-Short.
The construction of this mutant receptor has been described previously (Maggio et al., 1996). The encoded M3-short receptor has a deletion of 196 amino acids in the i3 loop, from Ala-274 to Lys-469 (Fig. 1).
pcDM2(Asn404 → Ser).
A 1.8-kilobase ApaI-HindIII and a 4.1-kilobaseDraIII-HindIII fragments were removed from HM2pcD and ligated together with a piece of DNA obtained by cutting with ApaI and DraIII, a fragment derived from the polymerase chain reaction of two partially complementary oligonucleotides (46- and 47-mer). The 47-mer oligonucleotide carried a point mutation that transforms the amino acid codon 404 of the HM2 sequence from asparagine to serine (AAT → AGC; Fig. 1). The integrity of the coding sequences was confirmed by dideoxy-DNA sequencing of the regions derived from the synthetic oligonucleotides and by restriction endonuclease analysis.
Transient Expression of Mutant Receptors.
COS-7 cells were incubated at 37°C in a humidified atmosphere (5% CO2) and grown in Dulbecco’s modified Eagle’s medium supplemented with 10% (v/v) fetal bovine serum, 2% (v/v) l-glutamine 200 mM, 1% (v/v) penicillin (10,000 U/ml) and streptomycin (10 mg/ml) solution, and 1% (v/v) minimal essential medium nonessential amino acid solution. The cells were seeded at a density of ∼1.5 × 106 cells/100-mm dish and 24 h later transiently transfected with the various receptor constructs (a total of 4 μg of plasmid DNA/dish) by a DEAE-dextran method (Cullen, 1987).
Radioligand-Binding Assays.
COS-7 cells were harvested 72 h after transfection. Cells were washed twice with PBS, scraped into ice-cold binding buffer (25 mM sodium phosphate containing 5 mM magnesium chloride at pH 7.3), and homogenized for 30 s using a polytron (setting 5). Membranes were pelleted at 15,000gfor 30 min at 4°C. The pellet was resuspended in 5 ml of ice-cold binding buffer and the membranes were rehomogenized. Saturation experiments were performed with 7 different [3H]NM concentrations (12.5–800 pM). Inhibition experiments were carried out with 16 different concentrations of the cold ligand (muscarinic agonist or antagonist), against an [3H]NM concentration of 200 pM. Atropine (1 μM) was used to define nonspecific binding. Incubation was at room temperature for 3 h. The bound ligand was separated on glass fiber filters (Whatman GF/B) with a Brandel cell harvester. The filters were washed three times with 4 ml ice-cold binding buffer. Filters were transferred to vials and counted on a liquid scintillation counter. Results are the mean ± S.E. of at least four experiments, each performed in duplicate.
Statistical Analysis.
Kd andBmax values of [3H]NM were determined in direct saturation experiments, whereas IC50values for all other compounds were calculated in competition curves fitted to one- or two-site binding models using the iterative, nonlinear, least-squares regression analysis of the Kaleidagraph software running on a Macintosh computer. To determine whether the data were best fitted by a one- or two-site model, the residual sums of squares were compared by Scheffé’s F test.
Results
Cotransfection of M2-trunc with M3-tail (see Fig. 1) results in the formation of a chimeric receptor with a high-affinity binding for [3H]NM. Table 1shows the affinity constants obtained by Barbier et al. (1998) for tripitramine, pirenzepine, methoctramine,para-fluoro-hexahydrosiladifenidol, and [3H]NM at M2, M3, and M2-trunc/M3-tail chimeric receptors. These compounds were used in our work to explore the possibility that cross-interaction between the wild-type M2 and M3 receptors could result in the formation of the heterodimeric M2-trunc/M3-tail receptor. We initially expressed the wild-type M2 receptor together with the M3-tail fragment in COS-7 cells. Then we tested pirenzepine in a displacement curve against [3H]NM. The idea was to see whether the M3-tail fragment (which by itself does not bind muscarinic ligands) was able to recruit the amino-terminal receptor domain of the muscarinic M2 receptor in the formation of the chimeric receptor. As pirenzepine has a higher affinity for the chimeric M2-trunc/M3-tail receptor than for the M2 receptor (Table 1), the displacement curve should have been shallow and best fitted by a two-site model. In agreement with our anticipation, the displacement curve we obtained with pirenzepine had a Hill coefficient of 0.79 ± 0.02 (significantly lower than unity), and the curve was best fitted by a two-site model (Fig. 2a). Thirty-four percent of receptors were in the high-affinity state, indicating that a fair amount of receptors were recruited in the chimeric receptor form. The IC50s that we calculated were very close to those found with pirenzepine for the wild-type M2 receptor and the coexpressed M2-trunc and M3-tail receptor fragments (compare Tables 1 and 2). In a similar experiment in which pirenzepine was tested on membranes obtained from cells transfected separately with M2 and M3-tail and then pooled together, we obtained a Hill coefficient not far from unity (1.01 ± 0.01; Fig. 2a) and an IC50 equal to that of the wild-type M2 receptor (Table2), indicating that the cotransfection of M2 and M3-tail in the same cells was the condition to observe two affinities.
Binding affinities of wild-type muscarinic M2 and M3receptors and the fragmented chimeric M2-trunc/M3-tail receptor for five different muscarinic receptor antagonists

Displacement of specific [3H]NM binding to cotransfected M2 and M3 wild-type and mutant muscarinic receptors by pirenzepine and tripitramine. a, pirenzepine inhibition curve (●) in COS-7 cells cotransfected with the M2 wild-type receptor and the M3-tail fragment. The curve was best fitted by a two-site model (p < .05, Scheffé’s F test); the two populations of binding sites showed a 14-fold difference in affinity. b, tripitramine inhibition curve (●) for the cotransfected M3 wild-type receptor and M2-trunc fragment. Also, this graph was best fitted by a two-site model (p < .05, Scheffé’s F test); the two populations of binding sites showed a 181-fold difference in affinity. c, tripitramine inhibition curve (●) for the cotransfected M3-short receptor and M2-trunc fragment. In this case, a single population of binding sites was identified by the fitting. d, tripitramine inhibition curve (●) for the cotransfected M2 (Asn404 → Ser) and M3 wild-type receptor. The curve was best fitted by a two-site model (p < .05, Scheffé’s F test); the two populations of binding sites showed a 346-fold difference in affinity. a, b, and d, ○, represent curves obtained in membranes from COS-7 cells separately transfected with M2 and M3-tail, M3and M2-trunc, and M2 (Asn404 → Ser) and M3, respectively. c, ○, curve obtained with the M3-short receptor transfected alone.
Antagonist binding parameters of wild-type, fragmented, and mutant muscarinic receptors coexpressed in COS-7 cells
[3H]NM binding to the M2receptor was not affected by the presence of the M3-tail fragment. The saturation curve was best fitted by a one-site model and the Kd value was very similar to that of the M2 receptor transfected alone (Table 3).
[3H]NM binding parameters of wild-type, fragmented, and mutant muscarinic receptors coexpressed in COS-7 cells
In another set of experiments, we cotransfected the wild-type M3 receptor with the M2-trunc receptor fragment. Then we tested tripitramine in a displacement curve against [3H]NM. As in the previous experiment, the idea was to see whether the M2-trunc fragment could recruit the carboxy-terminal domain of M3 to form the chimeric receptor. Due to the big difference in affinity between the M3 and the chimeric receptor, the inhibition curve obtained with tripitramine was clearly biphasic (Fig. 2b). The curve was best fitted by a two-site model formula and the affinities found were very close to those of the M3wild-type receptor and the fragmented M2-trunc/M3-tail chimeric receptor (compare Tables 1 and 2). Similar experiments performed with tripitramine on membranes obtained from COS-7 cells separately transfected with M3 and M2-trunc and then pooled together gave an inhibition curve indicative of a single population of binding sites (Hill coefficient 0.98 ± 0.03; Fig. 2b) with the affinity of the muscarinic M3 receptor (Table 2).
As observed with the previous cotransfection, [3H]NM binding to the M3receptor was not affected by the presence of the M2-trunc fragment. The saturation curve was best fitted by a one-site model and the Kd value was very similar to that of the M3 receptor (Table 3).
Using COS-7 cells cotransfected with M3-tail + M2 and M2-trunc + M3, we tested two other compounds:para-fluoro-hexahydrosiladifenidol and methoctramine. The data obtained with these two compounds in both cotransfections did not fit with a two-site model at the level of p < .05 by Scheffe’s F test, probably because they do not differentiate powerfully between M2-trunc/M3-tail on the one hand and M2 or M3 on the other (Table 2).
As mentioned in the introduction, receptor interaction is prevented by shortening of the i3 loop. We cotransfected a short form of the wild-type M3 muscarinic receptor (M3-short, Fig. 1) in which 196 amino acid of the i3 loop had been removed, with the M2-trunc receptor fragment, and then we tested tripitramine in a displacement curve against [3H]NM. Although we obtained a biphasic curve (see above) with the cotransfection of the muscarinic M3 wild-type receptor with the M2-trunc fragment, in this case we observed a monophasic curve, the expression of a single population of binding sites (the Hill coefficient was not significantly different from unity, Fig. 2C). The affinity of tripitramine was equal to that found when the M3-short receptor was transfected alone (Table2). These data clearly confirm that the recruitment of the carboxyl terminal receptor portion of M3 by the M2-trunc fragment (a fundamental requirement for the formation of the chimeric receptor) directly depends on the length of the i3 loop. [3H]NM binding parameters for the cotransfection tested above are reported in Table 3.
In the previous experiment we described interactions between receptors as a whole and receptor fragments. We did not test the two wild-type M2 and M3 receptors together because we do not have yet a ligand fully selective for the hybrid M2-trunc/M3-tail receptor. For this reason, the possibility remains that when the two wild-type M2 and M3receptors are coexpressed together, the steric hindrance of the proteins could prevent receptor interaction. To address this issue, we constructed a point mutant M2 receptor in which the asparagine 404 in the TMD VI was replaced by serine: M2 (Asn404 → Ser; Fig. 1). This amino acid (conserved in all five muscarinic receptors) has been demonstrated to drastically reduce antagonist binding (Blüml et al., 1994). In line with this finding, [3H]NM up to 2 nM was unable to bind M2 (Asn404 → Ser); nevertheless, when this receptor was transfected with the M3-tail receptor fragment, [3H]NM specific binding was detected, indicating the correct insertion of the receptor into the membrane; furthermore, the pirenzepine inhibition curve was best fitted by a one-site model (Hill coefficient = 1.05 ± 0.07) and the IC50 value was 135 ± 12 nM. Then we transfected M2 (Asn404 → Ser) together with the wild-type M3 receptor and we tested tripitramine in a displacement experiment against [3H]NM. As can be seen in Fig. 2d, this cotransfection resulted in the appearance of a high-affinity binding for tripitramine. The best ratio between M2 (Asn404 → Ser) and M3in terms of plasmid DNA amount was 3.3 and 0.7 μg, respectively. With this ratio of plasmid DNA we had 17% of the total binding in the high-affinity state, whereas the rest was in a low-affinity state. These results were not observed on membranes obtained from COS-7 cells separately transfected with M2 (Asn404 → Ser) and M3 and then pooled together (Fig. 2d). We performed this experiment also with a reduced amount of plasmid DNA to have a more physiological level of expression. Plasmid DNA was reduced to 0.94 and 0.2 μg, respectively for M2 (Asn404 → Ser) and M3receptors (the difference was made up with 2.86 μg of empty vector). In this condition, the total amount of [3H]NM binding calculated in saturation experiments decreased from 173 ± 36 to 36 ± 7 fmol/mg of proteins; theKd value was similar in both experiments and was similar to the value obtained with the muscarinic M3 receptor transfected alone (Table 3). Also, with a lower amount of receptors expressed in the membrane, tripitramine displacement experiments resulted in the appearance of high-affinity binding sites (Table 2).
We can conclude that a molecular interaction occurs between M2 (Asn404 → Ser) and M3when they are together in the same cells, and that a receptor with a high affinity for tripitramine is formed. Because M2 (Asn404 → Ser) and M2wild-type have only one amino acid difference, they probably have the same steric impediment, and consequently steric hindrance does not seem to play against the formation of the heterodimeric M2-trunc/M3-tail receptor.
Discussion
In the last few years, an increasing number of articles have been published that support the existence of G protein-coupled receptor dimers. Receptors that show some evidence of dimerization are muscarinic M2 and M3(Potter et al., 1991; Maggio et al., 1993b, 1996; Wreggett and Wells, 1995), β2-adrenergic (Hebert et al., 1996,1998), neurokinin NK-1 and -2 (Huang et al., 1994, 1995), adenosine A1 (Ciruela et al., 1995), dopamine D2 (Ng et al., 1996), angiotensin II (Monnot et al., 1996), δ-opioid (Cvejic and Devi, 1997), metabotropic glutamate mGluR5 (Romano et al., 1996), Ca2+-sensing (Bai et al., 1998, 1999), and γ-aminobutyric acid (GABA)B (Kaupmann et al., 1998; Jones et al., 1998; White et al., 1998). There is currently much debate on the role of dimers in G protein-coupled receptor, but three recent works on GABAB receptors (Kaupmann et al., 1998; Jones et al., 1998; White et al., 1998) have shed light on the functional significance of this phenomenon. Neither GABAB receptor 1 (R1) nor GABABR2, when expressed individually, activates inwardly rectifying K+ channels; however, the combination of GABABR1 and GABABR2 confer robust stimulation of K+ channel activity. This indicates that heteromeric assembly of the two GABAB receptor subunits is essential to confer function and suggests that intermolecular interaction could be of general importance for G protein-coupled receptor activity. The molecular mechanism underlying dimerization is not known, but recently Gouldson et al. (1997, 1998), using computer simulations, have proposed domain swapping in G protein-coupled receptor dimerization. Domain swapping is a very efficient method of forming dimers, because the interactions within the monomer are reused in the dimer.
Domain swapping has been proposed to be the base of α2/M3 and M3/α2 chimeric receptors heterodimerization. This phenomenon implies that fragment exchange occurs between the two chimeric receptors and that the trunc and tail portions of the α2 and M3receptors (even though they are present in separate proteins) have the ability to recognize each other and reconstitute the wild-type receptor complex. A long i3 loop allows considerable spatial freedom for the fragments belonging to the α2 and M3 receptors located in the two chimeric α2/M3 and M3/α2 receptors (Maggio et al., 1996). As all muscarinic receptors have a long i3 loop, it is reasonable to think that fragment exchange can occur also between the wild-type muscarinic receptors. If this proves to be true, interaction between two different muscarinic receptor subtypes could lead to the spontaneous formation of a new heterodimeric receptor.
It has previously been found that the amino-terminal fragment of the M2 receptor (M2-trunc) is able to interact with the carboxy-terminal part of the M3 receptor (M3-tail) to form a fragmented chimeric receptor (Maggio et al., 1993a). A pharmacological screening with muscarinic antagonists identified several compounds with high affinity for this fragmented M2-trunc/M3-tail receptor hybrid (Barbier et al., 1998). In this study, two of these compounds, pirenzepine and tripitramine, were used to show that interaction occurs between the M2 and the M3muscarinic receptors. Displacement experiments performed with these two compounds against [3H]NM indicated that the wild-type M2 and M3receptors are able to complex with the M3-tail and M2-trunc fragments, respectively. These receptor fragments are able to recruit a fraction of the wild-type M2 and M3 muscarinic receptors in the formation of the chimeric M2-trunc/M3-tail receptor. A definitive experiment using both M2 and M3 wild-type receptors in the same preparation could not be performed in our work because of the lack of a ligand fully selective for the chimeric M2-trunc/M3-tail receptor. For this reason, the use of a point mutant M2(Asn404 → Ser) was required to demonstrate that two entire receptors are still able to interact with each other. The M2 (Asn404 → Ser) receptor does not bind [3H]NM up to 2 nM, but binding can be rescued by cotransfection with the M3-tail fragment, indicating that this receptor is correctly inserted into the plasma membrane. Cotransfection of this mutant M2(Asn404 → Ser) with the wild-type M3 receptor results in the appearance of a high-affinity binding for tripitramine because of the interaction of the trunc part of M2 (Asn404 → Ser) with the tail part of M3. This experiment rules out the possibility that steric hindrance can interfere with receptor cross-interaction, and strengthens the concept that wild-type muscarinic receptors are able to interact. The possibility that the M3-tail fragment or the full-length M3 could serve only as chaperone to target the M2 (Asn404 → Ser) receptor to the cell surface and that the high-affinity binding for tripitramine in the cotransfection of M2 (Asn404 → Ser) with the wild-type M3 receptor could be caused by the rescue of M2 binding and not by the interaction of the trunc part of M2 (Asn404 → Ser) with the tail part of M3 is unlikely. Although the inhibition curve obtained with pirenzepine in the cotransfection of M3-tail with M2 was best fitted by a two-site model, the inhibition curve obtained with the cotransfection of M3-tail with M2 (Asn404 → Ser) was best fitted by a one-site model; the affinity found was similar to the high-affinity binding obtained with the cotransfection of M3-tail with M2 and there was no evidence of a low-affinity binding component, indicative of the rescue of M2binding. Nevertheless, it is possible that for the efficient targeting and the correct folding of the M2 (Asn404 → Ser) receptor in the plasma membrane, the presence of M3-tail or the full-length M3 receptor is required.
The comparison of these data and those obtained with the chimeric α2/M3 and M3/α2 receptors deserves comment. In the latter case, there is no interaction between the amino- and carboxy-terminal receptor parts inside the chimeric α2/M3 and M3/α2 receptors because they originate from two different receptor types; therefore, all the fragments are in some way forced to cross-interact. In the case of the wild-type M2 and M3receptors, the intramolecular interaction of the amino- and carboxy-terminal parts inside the receptors competes with the intermolecular interaction with the foreign fragments. For example, in the cotransfection of M2 with M3-tail, the trunc part of M2 will have a far greater probability of hitting the tail part of M2 than the foreign M3-tail fragment. As we calculated in our experiments that up to 34% of the total binding could be found in the hybrid receptor form, it is likely that the relative affinity between the trunc part of M2 and the M3-tail receptor fragment is high enough to overcome this disadvantage. Furthermore, it is possible that ligands, depending on their relative affinities for the hybrid and the wild-type receptors, can favor the intra- or the intermolecular interaction.
In our experiments, the existence of heterodimers is entirely based on pharmacological properties of coexpressed receptors and no biochemical evidence of their existence were presented. For the moment, we do not know the functional implications that the interaction between M2 and M3 receptors may have, and additional experiments will be necessary to reveal the biological significance of this phenomenon. As coexpression of different subtypes of muscarinic receptors in the same cells occurs in several tissues (Weiner et al., 1990; Fukamauchi et al., 1993; Eglen et al., 1994), it is possible that heterodimeric receptors could be formed in vivo. Based on the high structural homology found among all G protein-coupled receptors, our findings should be of general importance for the entire class of integral membrane proteins.
Acknowledgments
We thank R. Packham for his help in restyling the English of the manuscript.
Footnotes
-
Send reprint requests to: Roberto Maggio, Dipartimento di Neuroscienze, Sezione Farmacologia, University of Pisa, Via Roma 55-56126 Pisa, Italy. E-mail:r.maggio{at}drugs.med.unipi.it
- Abbreviations:
- TMD
- transmembrane domain
- COS-7
- African green monkey kidney
- [3H]NM
- N-[3H]methylscopolamine
- i3 loop
- third cytoplasmic loop
- Received January 20, 1999.
- Accepted May 4, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}