


Abstract
Screening of porcine cerebellum cDNA library with porcine endothelinB (ETB) receptor cDNA revealed a novel ETB receptor cDNA that is distinctly different from the wild-type ETB receptor in length and the amino acid sequence at the C-terminal end. This sequence appears to represent alternate splicing of the carboxy terminal end of ETBreceptor, resulting in a polypeptide of 429 amino acids in length, which is 14 amino acids shorter than the wild-type porcine ETB receptor. Characterization of the wild-type and alternately spliced ETB receptors expressed in COS cells revealed that both receptors displayed very similar binding [apparent dissociation constant (Kd) and maximum binding (Bmax) for 125I-ET-1 were 71 pM and 1.6 pmol/mg protein for wild-type and 81 pM and 1.2 pmol/mg protein for splice variant ETB receptors] as well as functional properties. These data suggest that the differences in the amino acids at the C-terminal end had no effect on binding or functional coupling of these alternately spliced ETBreceptors.
Endothelins (ET), a family of peptide hormones, originally identified by Yanagisawa et al. (1988), exert a number of physiological effects, including potent vasoconstriction, transient vasodilation, proliferation, and neuroregulatory functions (Simonson and Dunn, 1990; Rubanyi and Parker Bothels, 1991). This family consists of three members, ET-1, ET-2, and ET-3, that are encoded by three separate genes (Inoue et al., 1989). Soon after the discovery of ET peptides, a family of 21 amino acid peptide toxins (sarafotoxins) that shared a high degree of homology to ET was identified (Kloog and Sokolovsky, 1989). The biological effects of these peptides are mediated by specific cell surface G protein-coupled receptor superfamily. Based on the binding profiles of ETs and sarafotoxins, three ET receptors (ETA, ETB, and ETC) have been cloned and characterized (Arai et al., 1990; Sakurai et al., 1990;Karne et al., 1993). Although ETA and ETB receptors have been cloned from a number of species, including human (Adachi et al., 1991; Lin et al., 1991; Saito et al., 1991; Elshourbagy et al., 1992, 1993), ETC receptors have been identified only inXenopus (Karne et al., 1993). ETBreceptors display similar affinities for ET-1, ET-2, ET-3, and sarafotoxin 6c (S6c), whereas ETA receptors display similar affinity for ET-1 and ET-2, 100-fold lower affinity for ET-3, and >1000-fold lower affinity for S6c. ETCreceptors have been shown to bind ET-3 with much higher affinity than ET-1 (Karne et al., 1993). In addition to ETC, another subtype of ET receptors (ETAX) has been cloned and characterized from Xenopus heart (Kumar et al., 1994). These receptors, although maintaining the pharmacological characteristics of ETA receptors for ET-3 and S6c, display extremely low affinity to BQ123, originally identified as an ETA-selective antagonist (Ihara et al., 1992). Also, a number of alternately spliced forms of ET receptors have been identified and characterized. Cheng et al. (1993) reported an alternately spliced form of ETB receptors from rat brain. Compared with the wild-type receptor, this receptor had four amino acid substitutions in the amino terminal end, and the 5′- and 3′-noncoding regions were different. The binding profiles for various agonists were the same for these two receptors, although no functional data were reported (Cheng et al., 1993). An alternately spliced form of human ETB receptors was reported by Elshourbagy et al. (1996) and Shyamala et al. (1994). Although the splice variant reported by Shyamala et al. (1994) had 10 additional amino acids in the second cytoplasmic domain, there were no differences in binding as well as functional parameters between the splice variant and wild-type receptors (Shyamala et al., 1994). However, the splice variant of human ETB receptors reported by Elshourbagy et al. (1996) showed significant differences in the last 52 amino acids at the carboxy terminal. Although the binding parameters between this alternately spliced form and wild-type ETBreceptors were the same, the splice variant of ETB receptors was functionally uncoupled (Elshourbagy et al., 1996). The data presented herein demonstrates the presence of an additional splice variant of porcine ETB receptor. This receptor shows significant differences in the last 32 amino acids at the carboxy terminal compared with wild-type porcine ETB receptors. Furthermore, the binding and functional characteristics of this receptor are identical with the wild-type ETBreceptor.
Experimental Procedures
Materials.
125I-ET-1 and125I-IRL-1620 (specific activities, 2200 Ci/mmol) were obtained from New England Nuclear (Boston, MA). Unlabeled ET-1, ET-3, and S6c were from American Peptides (Santa Clara, CA). All other reagents were of the highest grade available.
Construction and Screening of cDNA Libraries.
The porcine cerebellum cDNA library (Elshourbagy et al., 1992) in pcDNA vector was screened by hybridization to nitrocellulose replicates with32P-labeled porcine ETB-R cDNA coding sequence as a probe in 20% formamide, 5× SSC (SSC is 150 mM NaCl, 15 mM sodium citrate), 5× Denhardt's, 0.1% SDS, and 0.2 mg/ml Escherichia coli tRNA at 42°C (Elshourbagy et al., 1990). Filters were washed with 2× standard saline citrate, 0.1% SDS at 42°C. Several positive recombinant clones were isolated from the porcine cerebellum library and characterized. Preliminary sequence analysis showed that six of these clones encode the ETB-R clones, except for two clones that contain the same 5′-coding region of ETB-R and a divergent 3′-coding sequence.
Nucleotide Sequence Analysis.
The inserts of the porcine ETB splice variant receptor (SVR)560 were sequenced on both strands with a modification of the dideoxy chain termination method (Sanger et al., 1977) with the Sequenase II kit (US Biochemical Corp., Cleveland, OH). The Wisconsin Genetics Computer Group Software package (Devereux et al., 1984) was used to assemble composite sequences from the various fragments and for future sequence analysis.
RNA Blot Analysis.
For Northern analysis, poly(A)+ RNA was isolated from various porcine tissues with the guanidinum thiocyanate acid-phenol method (Chomczynski and Sacchi, 1987). One microgram of each RNA was fractionated on 1% agarose formaldehyde gels (Lehrach et al., 1977) and transferred to nitrocellulose membranes. Northern hybridizations were performed at 42°C in 50% formamide, 5% subacute sclerosing panencephalitis, 5× Denhardt's reagent, 0.1% SDS, and 100 μg/ml yeast tRNA (Elshourbagy et al., 1985). The blots were washed with 0.1× standard saline citrate, 0.1% SDS at 50°C and exposed to X-ray film for 4 days at −70°C. Autoradiograms were analyzed by quantitative scanning densitometry.
Expression of Porcine ETB Receptors in COS Cells.
Fragments containing the entire porcine ETB-SVR cDNA and ETB-R cDNA coding sequence was subcloned into the mammalian expression vector pRLDN (Elshourbagy et al., 1993). COS cells grown in 245 × 245-mm tissue culture plates were transfected with 75 μg of porcine ETB-SVR or porcine ETB-R cDNA and grown in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum for 2 days as previously described (Elshourbagy et al., 1992).
Membrane Preparation.
COS cells transfected with wild type as well as splice variant of porcine ETBreceptors were washed with Dulbecco's phosphate-buffered saline containing a protease inhibitor cocktail (5 mM EDTA, 0.5 mM phenylmethylsulfonyl fluoride, 5 μg/ml leupeptin, and 0.1 μg/ml aprotinin) and scraped in the same buffer. The membranes were prepared following the procedure of Elshourbagy et al. (1993). Protein was estimated by bicinchoninic acid method.
Radioligand-Binding Studies.
125I-ET-1 and 125I-IRL-1620 binding to COS cell membranes were performed following the procedure of Elshourbagy et al. (1993)with the following modifications. The assay volume was 50 μl and the protein used was 50 to 100 ng/tube. Each experiment was done three times with membranes prepared from different transfections. The data presented are from one representative experiment.
Phosphoinositide Turnover.
COS cells transfected with wild-type and splice variant porcine ETB receptor clones were treated with 1 μCi/ml myo [3H]inositol for 24 h in serum-free medium. At the end of the treatment, the medium was removed, cells were washed with Dulbecco's phosphate-buffered saline, and then exposed to indicated concentrations of agonist for 10 min at 37°C. The reactions were stopped with 10% trichloroacetic acid, and the inositol phosphates were separated with ion exchange chromatography following the procedure of Aiyar et al. (1986).
Desensitization with ET-1 or Pretreatment with Phorbol-12,13-Dibutyrate (PDBu).
COS cells transfected with wild-type and splice variant procine ETB receptor clones were treated with 100 nM PDBu or 0.3 nM ET-1 for 30 min at 37°C. At the end of treatment, the medium was removed, cells were washed with Dulbecco's phosphate-buffered saline, and processed for binding assays (membranes) or functinal assays (inositol phosphate measurements).
Results
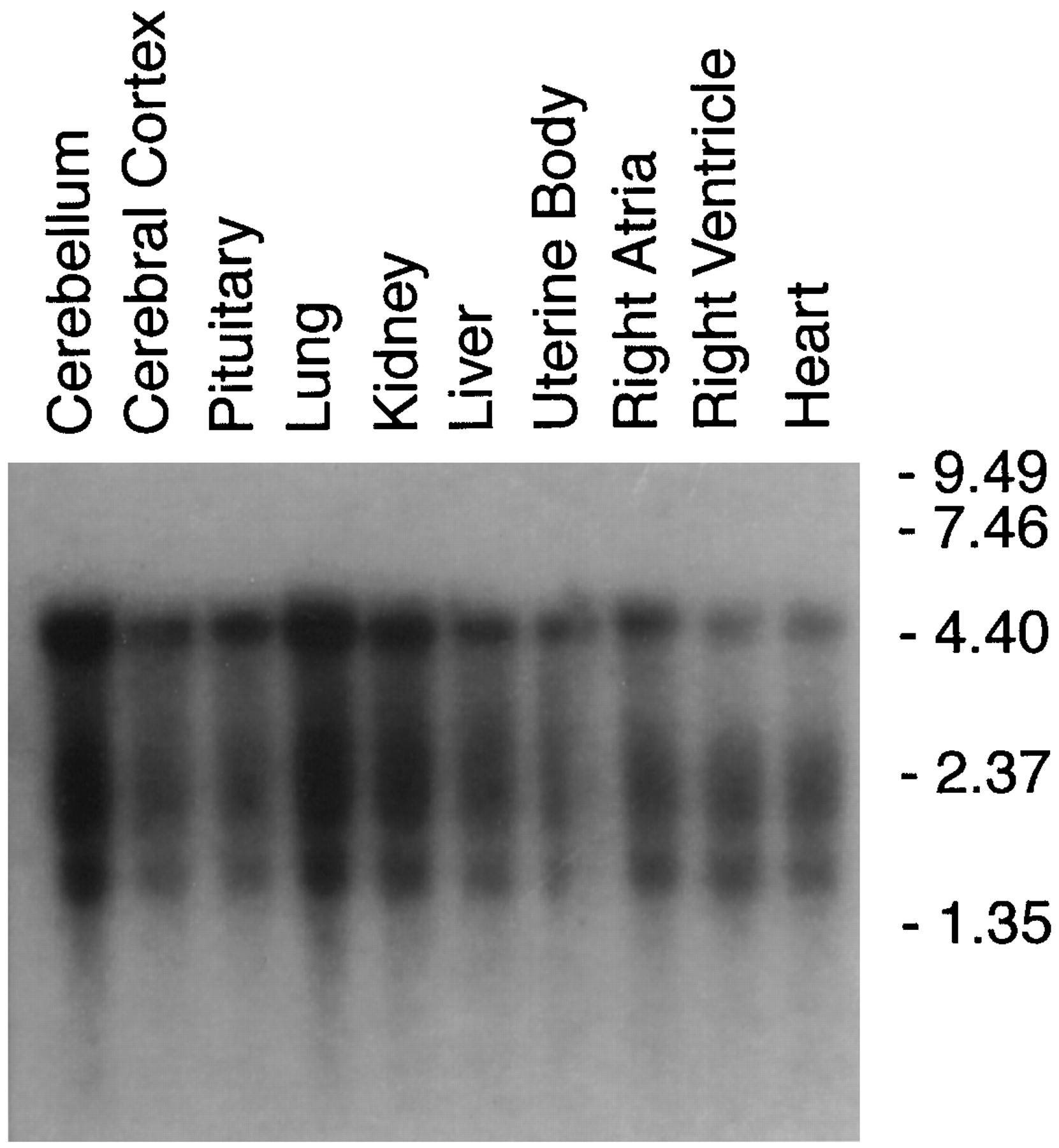
The deduced polypeptide of the alternately spliced form of porcine ETB (ETB-SVR) receptor consisted of 429 amino acid residues with a calculated molecular mass of ∼47.2 kDa (Fig. 1). This is different from the size of the wild-type porcine ETB receptor, which is 443 amino acids with a calculated molecular mass of 49 kDa (Fig. 1). Northern blot analysis and tissue distribution studies indicated the presence of two major bands (4.4 and 1.7 kilobases) corresponding to the full-length ETB receptor clones. These could have resulted from two polyadenylation signals that are 32 and 29 base pairs upstream of the polyadenylation sites of the two mRNA species. The additional band at 2.35 kilobases corresponds to the size of the splice variant ETB receptor. The three mRNA species appeared to be higher in the lung, kidney, cerebellum, and to a lesser extent in the cerebral cortex, pituitary, liver, uterine body, right atria, right ventricle, and the whole heart (Fig. 2).

Amino acid sequences of the porcine ETBreceptor cDNA clones. The recombinant plasmid clones of porcine ETB-R and ETB-SVR were sequenced by dideoxy method as explained in Experimental Procedures. Deduced amino acid residues are indicated beginning with the initiation methionine. The region identifying the positive TM as domains I–VII are underlined and numbered sequentially. The optimal alignment of the deduced amino acid sequences of ETB-R and ETB-SVR were made with Wisconsin program obtained fromDevereux et al. (1984).
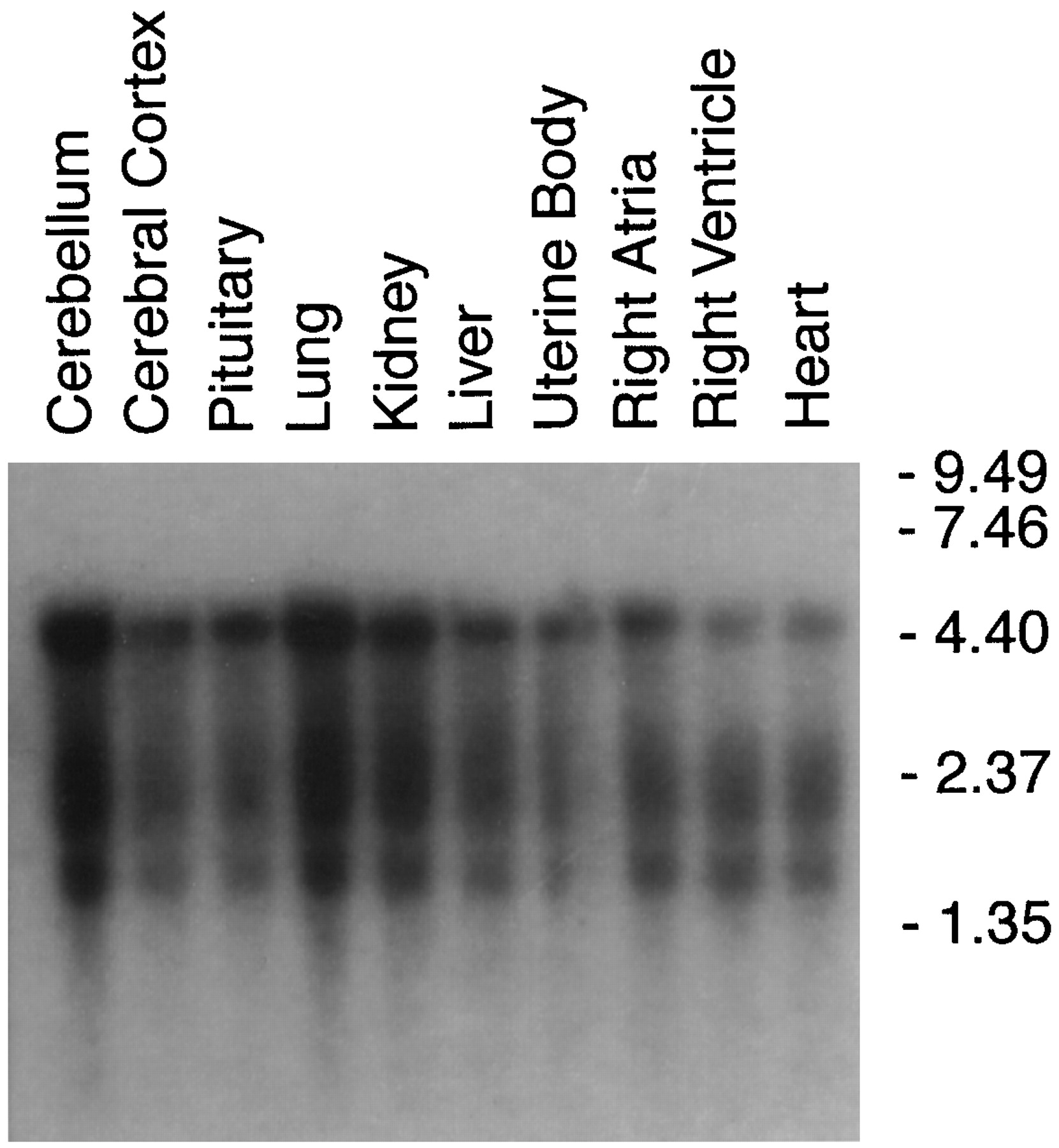
Size and tissue distribution of porcine ETB-R and ETB-SVR mRNA in various porcine tissues. Poly(A)+ RNA were prepared from the indicated tissues and then fractionated on a 1% agarose formaldehyde gel, blotted, and hybridized with the cloned 32P-labeled ETB-R cDNA. β-Actin was used as an internal standard for the amount of RNA loaded (data not shown).
Addition of increasing concentrations of125I-ET-1 or 125I-IRL-1620 (ETB-selective agonist) to membranes prepared from COS cells transfected with wild-type or splice variant porcine receptor clones resulted in specific, saturable, and high affinity binding as shown in Figs. 3 and4 (top and middle) for125I-ET-1 and125I-IRL-1620, respectively. The Scatchard transformation of the specific-binding data from saturation-binding experiments is presented in Figs. 3 and 4 (bottom). The apparent dissociation constant (Kd) and maximum binding (Bmax) for125I-ET-1 were 71 pM and 1.6 pmol/mg protein for wild-type and 81 pM and 1.2 pmol/mg protein for splice variant ETB receptors (Fig. 3; bottom). Under the same conditions, 125I-IRL-1620 gaveKd andBmax values of 124 pM and 1.2 pmol/mg protein, respectively, for wild-type and 158 pM and 1.1 pmol/mg protein, respectively, for splice variant ETBreceptors (Fig. 4; bottom). Competition-binding experiments with unlabeled ET-1, IRL-1620 [ETB-selective agonist (Takai et al., 1992; Nambi et al., 1994)], and RES701 [ETB-selective antagonist (Tanaka et al., 1994)] displayed identical binding profiles when tested against both receptors (Fig. 5). Untransfected or vector-transfected COS cells did not display any125I-ET-1 binding. No dissociation of bound125I-ET-1 was observed for wild-type and splice variant receptor (data not shown).
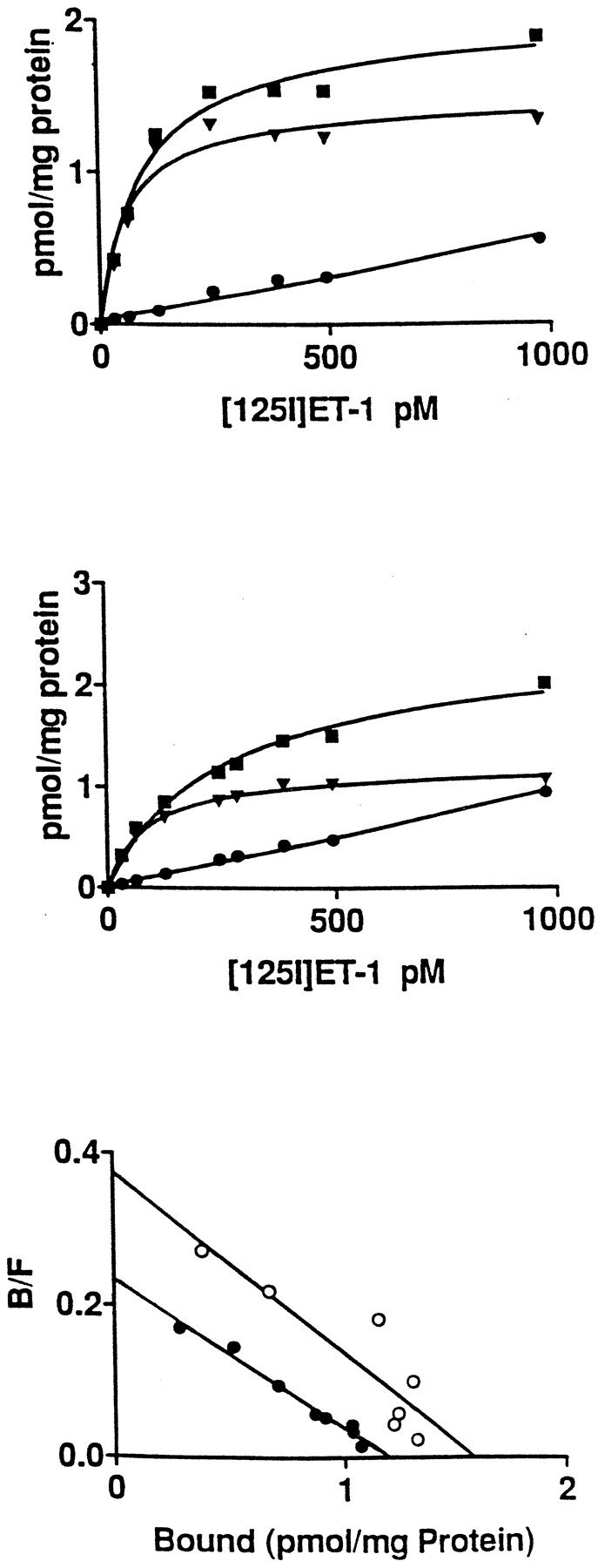
Saturation binding of 125I-ET-1 to membranes prepared from COS cells transfected with porcine ETB-R (top) and ETB-SVR (middle) clones. Increasing concentrations of 125I-ET-1 were added to membrane preparations in the absence (T, ▪) or presence (N, ●) of unlabeled ET-1 (1 μM). Bound and free ligands were separated as explained in Experimental Procedures. S (▾) represents specific binding. Bottom graph presents the Scatchard transformation of the specific-binding data obtained for porcine ETB-R (○) and ETB-SVR (●). The data presented are from one experiment that is representative of three experiments with different transfections.

Saturation binding of 125I-IRL-1620 to membranes prepared from COS cells transfected with porcine ETB-R (top) and ETB-SVR (middle) clones. Increasing concentrations of 125I-IRL-1620 were added to membrane preparations in the absence (T, ▪) or presence (N, ●) of unlabeled IRL-1620 (1 μM). Bound and free ligands were separated as explained in Experimental Procedures. S (▾) represents specific binding. Bottom graph presents the Scatchard transformation of the specific-binding data obtained for porcine ETB-R (○) and ETB-SVR (●). The data presented are from one experiment that is representative of three experiments with different transfections.

Competition of 125I-ET-1 binding to membranes prepared from COS cells transfected with porcine ETB-R (▪) and ETB-SVR (▴) by increasing concentrations of unlabeled ET-1 (top), IRL-1620 (middle), and RES701 (bottom). 125I-ET-1 (0.3 nM) was added to membranes in the absence and presence of increasing concentrations of unlabeled ligand and processed as explained in Experimental Procedures. Data presented are from one experiment that is representative of three experiments performed with different transfections.
Exposure of [3H]myoinositol-labeled COS cells expressing wild-type as well as splice variant porcine ETB receptors to increasing concentrations of ET-1 resulted in a concentration-dependent increase in the accumulation of inositol phosphates as shown in Fig.6. The concentration-response curves for ET-1 were identical for the two ETB receptors, indicating that the differences in the amino acids at the C-terminal end had no effect on the functional coupling of these receptors. ET-1 did not stimulate or inhibit inositol phosphates accumulation in untransfected or vector-transfected COS cells.
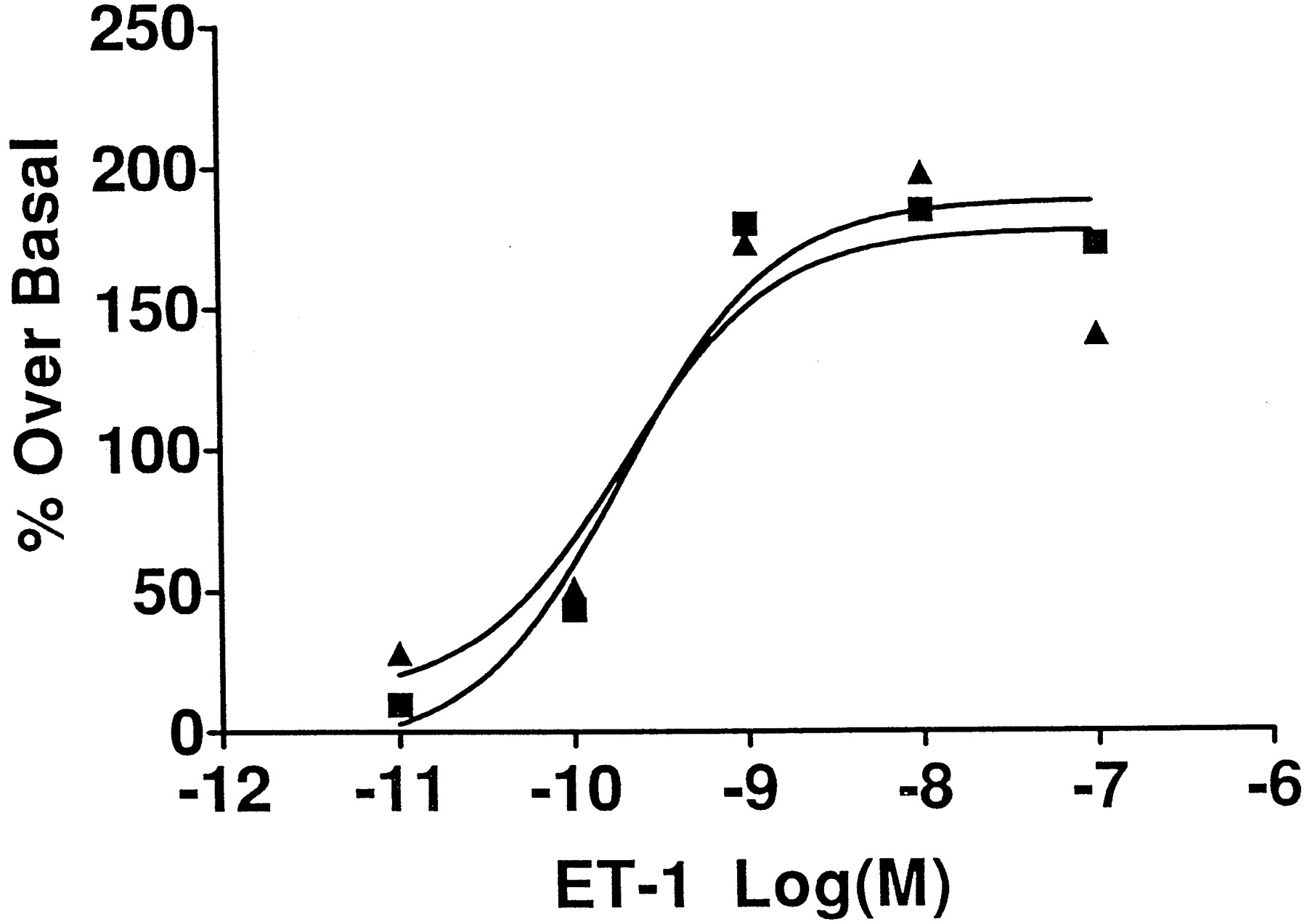
Concentration-response curves of ET-1 for the stimulation of inositol phosphates accumulation in COS cells transfected with ETB-R (▪) and ETB-SVR (▴). Transfected COS cells were pretreated with 1 μCi/ml [3H]myoinositol in inositol- and serum-free medium for 24 h. The cells were washed to remove excess [3H]myoinositol and challenged with increasing concentrations of ET-1 for 20 min at 37°C. Isolation and quantitation of inositol phosphates were done as explained in Experimental Procedures. The data presented are from one experiment that is representative of three experiments. The basal levels of inositol phosphates were 10,606 dpm/106 cells and 13,735 dpm/106 cells for wild-type and splice variant ETB receptor, respectively.
Experiments also were performed to test whether there were any differences in the regulation of these two receptors. Desensitization of these receptors by pretreating with ET-1 followed by rechallenge with ET-1 resulted in a 34.7% decrease in binding and a 35.3% decrease in inositol phosphates accumulation for wild-type and a 38% decrease in binding and a 41.6% decrease in inositol phosphates accumulation for splice variant receptor. There were no significant changes in Kd values for binding or EC50 values for function between control and desensitized receptors.
Similarly, pretreatment of these receptors with PDBu (an activator of protein kinase C) resulted in a 41.5 and 66.8% decrease in binding and inositol phosphates accumulation, respectively, for wild-type and a 28.4 and 36% decrease in binding and inositol phosphates accumulation, respectively, for splice variant receptors. Exposure of wild-type or splice variant ETB receptors to ET-1 had no effect on intracellular accumulation of cAMP (data not shown).
Discussion
The porcine ETB receptor cDNA, previously cloned in our laboratory (Elshourbagy et al., 1992) was used to probe porcine cerebellum cDNA library. Many positive clones were identified and nucleotide sequence analysis revealed that several of these positive clones encoded for ETB-R, except for two novel ETB-R clones that differed in the length and the amino acid sequence at the 3′-coding region. Sequence analysis of these clones revealed an identical sequence to the ETB receptor from the 5′-untranslated region through the putative seventh transmembrane domain (7TM). The sequence then diverged completely in the cytoplasmic domain and 3′-untranslated region. Several independent clones in our library were found to contain this sequence, which appears to represent an alternate splicing of the carboxy-terminal tail of the ETB receptor. Similar to the human ETB-SVR (Elshourbagy et al. 1996), the 3′-terminal tail of the porcine ETB-SVR clone was found to correspond to the extreme 3′-untranslated region of wild-type ETB-R. Analysis of the ETB-R genomic structure (Arai et al., 1993) revealed a similar splicing mechanism to that observed in the human ETB-R splice variant reported by Elshourbagy et al. (1996), namely, exons 1 through 6 are identical in ETB-SVR and the ETB-R in terms of their nucleotide sequence composition and their splice site. The carboxy-terminal tail of the procine and human ETB-SVR correspond to the extreme 3′-untranslated region of the ETB-R. However, the location of the splicing site of the porcine and the human are different; namely the putative splice site in the human and the porcine use an A(G/G)C and A(G/G)G only four nucleotides away from each other at the 3′-acceptor site. As a result, the 3′-coding region of the porcine ETB-SVR and the human ETB-SVR are different. Thus, two putative splice sites were identified at exon 7 in the porcine ETB receptor gene; the first splice site produced a 2925-bp exon that encodes the normal ETB-R, and the second splice site produced a 1151-bp exon that encodes the ETB-SVR.
Receptor subtypes can arise through divergent genes, e.g., ETA and ETB and in the case of intron-containing genes, additional variants within a subtype can arise by alternative RNA splicing. Recent studies have identified an ETB receptor variant that contains an additional 10 amino acids in the second cytoplasmic domain of the ETB receptor (Shyamala et al., 1994). This sequence was part of the ETB receptor intron that separates the second and third exons and therefore arises by alternative RNA splicing of a single gene. The identification of splice variants among the seven TM receptors has been increasing at a phenomenal rate following the initial observation of two variant forms for dopamine D2 receptor (Sibley and Monsma, 1992). Recent splice variants have been identified for thyrotropin-stimulating hormone (THR) (de la Pena et al., 1992), neurokinin receptors (Fong et al., 1992), prostaglandin EP3 receptor (Sugimoto el al., 1993; Namba et al., 1993), pituitary adenylyl cyclase-activating polypeptide (PACAP receptor) (Spengler et al., 1993), and monocyte chemoattractant protein (MCP-1) receptor (Charo et al., 1994). It is interesting to note that in all these examples, the cytoplasmic domain of the receptor is altered, suggesting that alternative RNA splicing may play a role in the generation of physiologically divergent receptor activity for the same ligand. However, there is no experimental data to support this.
Radioligand binding data indicated that these two receptors are expressed at the same level with similar affinities, suggesting that the modifications present at the carboxyl terminal of the receptor did not influence the binding of agonists or antagonists. This is not surprising because the binding of agonists and/or antagonists to 7TM G protein-coupled receptors is predicted to be at the extracellular and, in some cases, TM regions, whereas the functional coupling to the second messenger systems is predicted to be at the intracellular domain.
Contrary to what we have reported for human ETBreceptor splice variant (Elshourbagy et al., 1996), porcine ETBR and ETBSVR behaved very similarly in their functional response (stimulation of inositol phosphates accumulation). The critical requirements of palmitoylation and phosphorylation for signal transduction pathways are not clear. A careful comparison of the C-terminal region of wild-type and splice variant porcine ETB receptors reveals that the wild-type receptor contains 9 Ser, 5 Cys, 2 Tyr, and 0 Thr, whereas splice variant receptor contains 2 Ser, 2 Cys, and 2 Thr. In addition, the splice variant receptor is 14 amino acids shorter than the wild-type ETB receptor. The observation that wild-type as well as splice variant ETB receptors display very similar binding and functional properties suggests that neither the truncated C-terminal tail nor the changes in amino acids contribute to the binding or function of these receptors. Mutation of the Cys residues at the C-terminal end of a number of 7TM G protein-coupled receptors have resulted in different functional responses. Although mutation of the Cys residue at the C-terminal end of β-adrenergic (O'Dowd et al., 1989) and ETB(Koshimizu et al., 1995) receptors resulted in the loss of function of these mutated receptors, mutations of Cys residues of vasopressin V2 receptors (Sadeghi et al., 1997), α-2 adrenergic receptors (Kennedy and Limbird, 1993), dopamine D1 receptors (Jin et al., 1997), lutenizing hormone/human choriogonadotropin receptors (Kawate et al., 1997), and thyrotropin receptors (Kosugi and Mori, 1996) had no significant effect on the functional coupling of these receptors. These data clearly suggest that although palmitoylation of Cys residues is critical for certain receptors, it is not required for many other receptors for successful functional coupling. Koshimizu et al. (1995) reported the critical requirement of one Cys (amino acid 402) for functional coupling of human ETB receptors. This conclusion was based on the generation of a number of C-terminal mutants. It is possible that in the alternatively spliced porcine ETB receptors, this critical requirements of the Cys residue could have been satisfied by the Cys present in positions 409 or 410.
For a number of G protein-coupled receptors, the mechanism of desensitization appears to be phosphorylation of the receptors on serine and threonine residues by at least two classes of protein kinases. These are second messenger-activated kinases such as cAMP-dependent protein kinase or protein kinase C and a family of G protein-coupled receptor kinases (GRK). These kinases have been shown to preferentially phosphorylate residues in certain sequences. For example, cAMP-dependent protein kinase prefers RXS, RRXS, RXXS, or KRXXS, whereas protein kinase C prefers K/RXXS/T, S/TXK/R, K/RXS/T, or K/RXS/TXK/R. However, GRKs are classified as acidotropic kinases because of their preference for acidic amino acids proximal to phosphorylatable residues (EDST, EESSSS, EESV). Similar to several G protein-coupled receptors, ETA and ETB receptors each have cytoplasmic carboxy-terminal domains rich in serines and threonines. Because the homology between ETA and ETB receptors at the cytoplasmic tail is relatively low, it is possible that these receptors might be regulated differentially by GRKs. However, studies performed with cytoplasmic tail truncation mutants of ETA (Cyr et al., 1993) and ETB receptors (Koshimizu et al., 1995) suggest that agonist-mediated desensitization was unaffected by the removal of large portion of cytoplasmic tail of each receptor. The data presented in this report also suggest that there was no difference between wild-type and splice variant ETB receptor in ET-mediated desensitization or phorbol ester-mediated regulation, suggesting that the amino acid changes or C-terminal truncation of 14 amino acids did not have any effect on ET binding, function, or regulation.
Acknowledgments
We would like to thank Ganesh Sathe, Joyce Mao, and Stephanie Vanhorn for oligonucleotide synthesis and sequencing, and Sue Tirri and Maria McDevitt for excellent secretarial assistance.
Footnotes
-
Send reprint requests to: Ponnal Nambi, Ph.D., Cardiovascular Sciences, DuPont Pharmaceuticals Company, Experimental Station, E 400/3237, Wilmington, DE 19880-0400.
-
↵1 Present address: Cardiovascular Sciences, DuPont Pharmaceuticals Company, Experimental Station, E 400/3237, Wilmington, DE 19880-0400.
- Abbreviations:
- ET
- endothelin
- S6c
- sarafotoxin 6c
- ETB-SVR
- ETB splice variant receptor
- bp
- base pair
- PDBu
- phorbol-12,13-dibutyrate
- TM
- transmembrane
- GRK
- G protein-coupled receptor kinases
- Received December 22, 1998.
- Accepted August 11, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}