


Abstract
Trypsin and mast cell tryptase cleave within the extracellular N terminus of proteinase-activated receptor-2 (PAR-2), exposing a tethered ligand (SLIGRL) that binds and activates the cleaved receptor. We examined the neuronal expression of PAR-2 and its role in intestinal ion transport. Short-circuit current elevations in response to trypsin or the receptor-activating peptide SLIGRL-NH2 were measured in sheets of mucosa-submucosa from porcine ileum. SLIGRL-NH2 or trypsin rapidly elevated short-circuit current after their contraluminal application with respective 50% effective concentrations of 184 and 769 nM. Their actions were attenuated after contraluminal administration of the neuronal conduction blocker saxitoxin (0.1 μM); the cyclooxygenase inhibitor indomethacin (10 μM); or the Na+/K+/Cl− cotransport inhibitor furosemide (10 μM), but not by atropine (0.1 μM), a muscarinic cholinergic antagonist. In addition, soybean trypsin inhibitor (5 μg/ml) reduced mucosal responses to trypsin. The δ-opioid agonist [d-Pen2,5]-enkephalin (0.1 μM) inhibited trypsin action, an effect that was prevented by naltrindole (0.1 μM), a δ-opioid antagonist. PAR-2 immunofluorescence was localized in the mucosa using a receptor-specific antibody. PAR-2-like immunoreactivity was detected in myenteric and submucosal neurons, nerve fibers innervating ileal smooth muscle and mucosa, and in enteroendocrine cells. Some neurons coexpressed PAR-2- and choline acetyltransferase-like immunoreactivity. These results indicate that PAR-2 is expressed on cholinergic and noncholinergic submucosal neurons in porcine ileum. PAR-2 agonists stimulate active anion secretion by a neurogenic mechanism that is modulated by prostanoids and opioids. These receptors may have a potentially important role in intestinal neuroimmunomodulation.
Because of its direct contact with the external environment and its vast surface area, the mammalian gastrointestinal (GI) tract is vulnerable to colonization by many pathogens. To combat enteric pathogens such asSalmonella, the GI tract has evolved several nonspecific defenses that are mediated by mast cell products. Activated mast cells release inflammatory mediators that include proteinases, eicosanoids, histamine, and cytokines (Marone et al., 1998). Through its dilution of luminal pathogens and facilitation of the movement of secretory IgA, defensins, mucins, and other protective factors to the surface of the intestinal epithelium, active transepithelial anion secretion is an important component of primary mucosal defense. It can be evoked by several classes of inflammatory mediators acting at receptors located on enteric neurons (Perdue and McKay, 1994). There is a particularly strong association between mucosal mast cells and enteric secretomotor neurons in the neuroimmune modulation of intestinal ion transport (Berin et al., 1999).
Proteinase-activated receptors (PARs) are members of a newly discovered subfamily of G-protein-coupled receptors that play important roles in responses to injury, including inflammation and repair (Déry et al., 1998; Hollenberg, 1999). Proteases cleave within the extracellular N-terminal tails of PARs to expose tethered ligand domains that bind to and activate the cleaved receptors. Thrombin cleaves and activates PAR-1, PAR-3, and PAR-4, which mediate platelet aggregation and the inflammatory and proliferative effects of thrombin in multiple tissues (Déry et al., 1998; Hollenberg, 1999). In the mouse and rat, trypsin and mast cell tryptase cleave PAR-2 to expose the tethered ligand sequence SLIGRL-NH2. PAR-2 is highly expressed in the GI tract where it has been detected in enterocytes (Kong et al., 1997), myocytes (Corvera et al., 1997), and enteric neurons (Corvera et al., 1999). Trypsin, tryptase, and analogs of the tethered ligand (i.e., activating peptides) stimulate eicosanoid secretion in intestinal epithelial cells (Kong et al., 1997), inhibit colonic motility (Corvera et al., 1997), and excite enteric neurons (Corvera et al., 1999) and spinal afferent neurons (Steinhoff et al., 2000).
In the rat jejunum, trypsin and SLIGRL-NH2, but not thrombin, have been found to stimulate active ion transport (Vergnolle et al., 1998). It has not been determined whether these effects extend to other animal species or intestinal segments. Moreover, the mechanisms underlying PAR-2-mediated intestinal secretion have not been documented. Because it can be activated by mast cell tryptase, PAR-2 may be involved in intestinal inflammatory reactions to infection or tissue damage. Therefore, in this investigation we tested the hypothesis that PAR-2 is expressed on enteric neurons that mediate active anion secretion. Moreover, we examined the involvement of prostanoids in the effects associated with PAR-2 activation. Finally, because δ-opioid receptors associated with secretomotor neural pathways inhibit active secretion in the porcine intestinal mucosa (Quito and Brown, 1991), the ability of a δ-opioid receptor agonist to modify PAR-2-mediated intestinal secretion was investigated.
Materials and Methods
Chemicals and Drugs.
Trypsin (TPCK treated) was obtained from Worthington Biochemical Corp. (Freehold, NJ). Peptidase inhibitors (captopril, amastatin, and phosphoramidon), atropine, indomethacin, furosemide, soybean trypsin inhibitor, and carbamylcholine chloride (carbachol) were obtained from Sigma Chemical Co. (St. Louis, MO). Saxitoxin and naltrindole were purchased from Research Biochemicals International (Natick, MA); [d-Pen2,5]-enkephalin (DPDPE) was purchased from Peninsula Laboratories (Belmont, CA). The PAR-2 agonist SLIGRL-NH2 and its reversed peptide analog LRGILS-NH2 were synthesized by solid phase methods and purified by HPLC.
Animals.
Yorkshire pigs of each sex, 6 to 8 weeks old, were obtained from the University of Minnesota Research Animal Resources and University of Minnesota Agricultural Experiment Station (Rosemount, MN) breeding facilities. Pigs received nonmedicated pig feed ad libitum and were not fasted before sacrifice. The pigs were initially sedated with an i.m. injection of tiletamine hydrochloride-zolazepam (Telazol, 8 mg/kg; Fort Dodge Laboratories, Fort Dodge, IA), in combination with xylazine (8 mg/kg). The animals were euthanized by barbiturate overdose (Beuthanasia-D, 30 mg/lb i.v.; Schering-Plough Animal Health Corp., Kenilworth, NJ) in accordance with approved University of Minnesota Animal Care Committee protocols. A midline laparotomy was performed, and a 20- to 30-cm segment of ileum was obtained that extended from the ileocecal junction to 10 cm oral to the termination of the ileocecal ligament. Each intestinal segment was excised along its antimesenteric aspect, intestinal contents removed, and the tissue was placed in ice-cold oxygenated physiological salt solution modified to approximate the composition of porcine extracellular fluid (Chandan et al., 1991).
Measurement of Transepithelial Ion Transport.
The circular and longitudinal muscle layers were removed by blunt dissection, and sheets of ileal mucosa with attached submucosa were mounted in Ussing chambers (flux area = 1 or 2 cm2) under short-circuit conditions. The tissues were continuously bathed at 39°C (porcine core temperature) in oxygenated physiological salt solution with 10 mM mannitol or d-glucose added to the luminal or contraluminal sides of the tissue, respectively. Short-circuit current (Isc) and open-circuit transmural potential difference were measured across mucosal sheets, and tissue conductance (Gt) was calculated from these electrical parameters by Ohm's law as previously described (Chandan et al., 1991). Tissues were incubated for 25 to 35 min until the baseline Isc stabilized, and drugs and other substances were then added to either the luminal or contraluminal side of each sheet. Changes in Isc and potential difference were measured after drug administration. At the end of each experiment after Isc returned to baseline values, 10 mM glucose was added to the luminal aspect of each tissue, and subsequent changes in mucosal Isc were measured in response to glucose-coupled sodium absorption to assess tissue viability.
In determinations of concentration-effect relationships with SLIGRL-NH2 or its reversed analog LRGILS-NH2, tissues were pretreated with a cocktail of protease inhibitors containing 10 μM captopril, 100 μM amastatin, and 1 μM phosphoramidon. The cocktail was added to the contraluminal side 5 min before peptide addition. In some experiments, 5 μg/ml soybean trypsin inhibitor (SBTI), 0.1μM saxitoxin, 10 μM indomethacin, or 10 μM furosemide was added to the contraluminal side 5 min before trypsin or SLIGRL-NH2 addition. In other experiments, tissues were pretreated contraluminally with 100 nM DPDPE for 5 min before trypsin addition; in some cases, the δ-opioid antagonist naltrindole was present in the contraluminal bathing medium 5 min before DPDPE was added.
Localization of PAR-2 Immunoreactivity.
To localize PAR-2 immunoreactivity, an antiserum to PAR-2-B5, raised in rabbits to a peptide fragment of rat PAR-2 (30GPNSKGR↓SLIGRLDT46P-YGGC; arrow denotes trypsin cleavage site) and conjugated to keyhole limpet hemocyanin (Saifeddine et al., 1996) was generously provided by Dr. Morley D. Hollenberg (University of Calgary, Canada). To identify cholinergic neurons, an antibody to choline acetyltransferase (ChAT) was purchased from Chemicon International, Inc. (Temecula, CA). Protein gene product 9.5 (PGP9.5; Chemicon International, Inc.) was used as a general neuronal marker. Donkey anti-rabbit IgG-indocarbocyanine- and donkey anti-goat IgG-fluorescein isothiocyanate-conjugated secondary antibodies were purchased from Jackson Immunoresearch Laboratories, Inc. (West Grove, PA).
Ileal segments from three pigs were isolated and immersed in ice-cold 2% paraformaldehyde in PBS at pH 7.4 for 2 h. All tissues were cryoprotected in increasing (10–30%) concentrations of sucrose in PBS, embedded in Tissue Tek O.C.T. compound (Baxter Healthcare Corp., McGaw Park, IL), and frozen. Coronal sections of ileum (15 μm in thickness) were thaw-mounted onto Superfrost-plus slides (Fisher Scientific, Pittsburgh, PA) and stored at −20°C until use.
A standard immunofluorescence staining protocol was followed. Briefly, tissue sections were rehydrated in PBS (pH 7.4) for 15 min, and then incubated in 0.4% Triton X-100 (Sigma Chemical Co.) and 2% BSA (Sigma Chemical Co.) in PBS for 30 min to block nonspecific binding. Sections were simultaneously incubated in antibodies to PAR-2-B5 (1:250 dilution) and ChAT (1:30 dilution) in 0.4% Triton X-100 and 2% BSA overnight at 4°C. After three rinses in PBS, sections were further incubated with appropriate secondary antibodies (donkey anti-rabbit indocarbocyanine-conjugated IgG at 1:400 dilution; or donkey anti-goat fluorescein isothiocyanate-conjugated IgG at 1:40 dilution) in PBS for 1 h in the dark. After three rinses in PBS for 15 min, coverslips were mounted with Vectashield (Vector Laboratories, Burlingame, CA), and the edges were sealed with nail polish. Antibody to PGP9.5 (1:150 dilution, overnight incubation) was used as general neuronal marker to confirm the neuronal morphology.
Controls consisted of omission of primary antibody from the staining protocol, replacing the primary antibody with another unrelated primary antibody or preabsorbing the primary antibodies against the antigen that resulted in complete absence of specific immunoreactivity. Omission of PAR-2 antibody resulted in absence of specific immunoreactivity pattern. ChAT immunoreactivity was substantially reduced when anti-ChAT antiserum was preincubated with 2 μg/ml of ChAT antigen; however, a nonspecific punctate staining pattern persisted.
Sections were scanned using a Bio-Rad confocal laser scanning microscope (CLSM, model 1000) that was attached to a Nikon fluorescence microscope. Images were obtained using Comos software (version 6.05.8; Comos Bio-Rad, Hercules, CA) and further processed using NIH Image (version 1.59) and Adobe Photoshop (version 4.0) software.
Data Analysis.
Data are expressed as mean ± S.E. of peak changes in Isc relative to baseline values occurring in response to drug administration. Tissues from at least three pigs were used in all experiments, with the exception of experiments with LRGILS-NH2, which used three tissues from two pigs. Determinations of agonist concentration-effect relationships through nonlinear regression methods and statistical analyses of trypsin and SLIGRL-NH2 data were performed using the PRISM computer software program (GraphPad, San Diego, CA). Comparisons between a single control mean and treatment mean were made with an unpaired, two-tailed Student's t test; comparisons of a control mean with multiple treatment means were made by ANOVA followed by Tukey's test. In all cases, the limit for statistical significance was set at P < .05.
Results
Effects of Trypsin and the PAR-2-Activating Peptide SLIGRL-NH2 on Transepithelial Conductance and Isc.
After their contraluminal addition at single concentrations, trypsin and SLIGRL-NH2 produced rapid increases in Isc. Contraluminally applied LRGILS-NH2, however, did not significantly affect either Isc or Gt (Fig.1). When added in cumulatively increasing contraluminal concentrations, trypsin and SLIGRL-NH2 were equieffective in elevating Isc (Fig. 2). Maximal mucosal responses to cumulative additions of trypsin or SLIGRL-NH2were not significantly different from those produced by the administration of these substances at single concentrations (based on comparisons of responses to 1.0 μM trypsin or 0.1 μM SLIGRL-NH2; one-way ANOVA, P > .05). Relative to its effects after contraluminal addition, luminally applied trypsin was less potent and effective in elevating Isc (Fig.2).

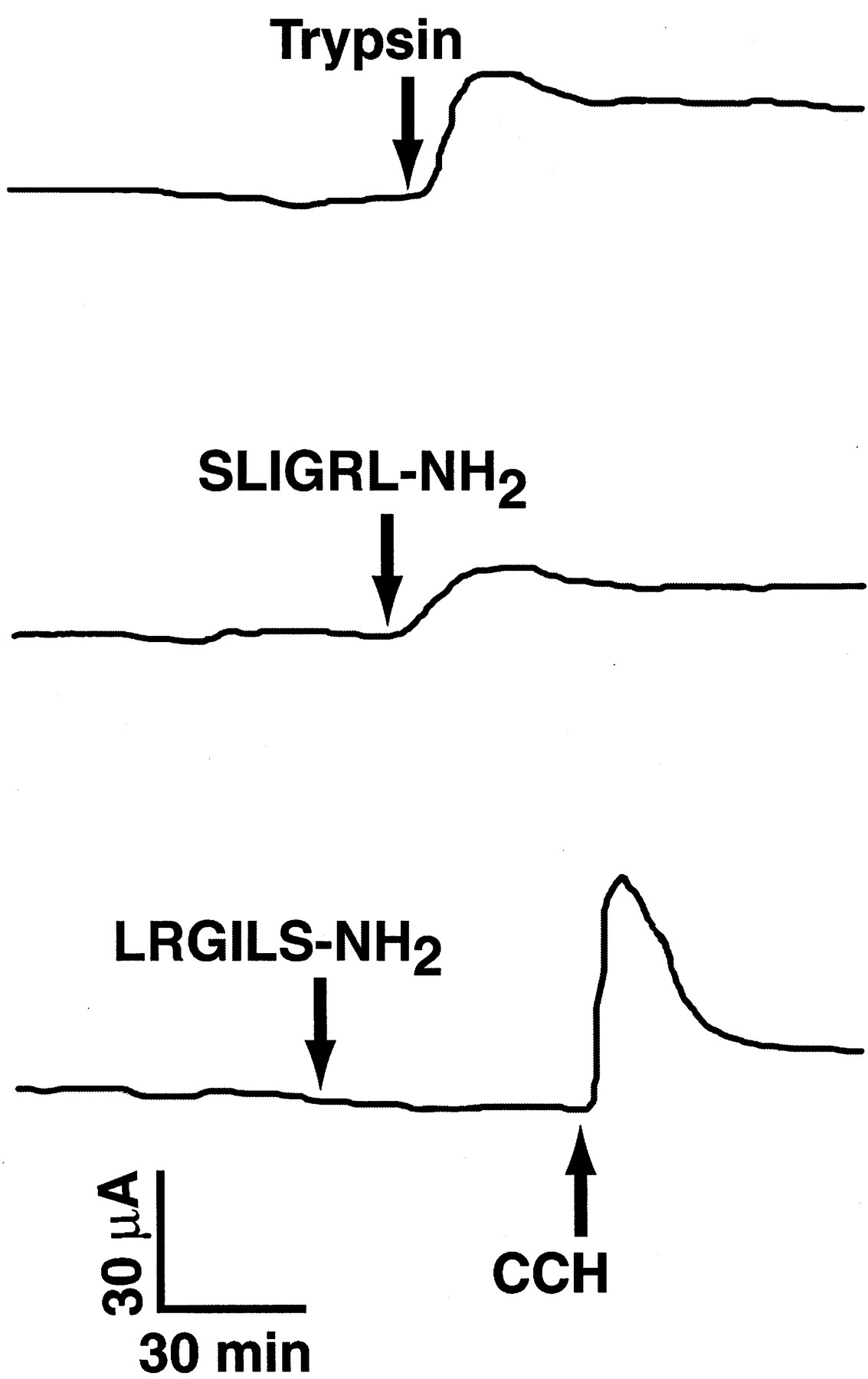
Chart records depicting the effects of trypsin (1.0 μM), the PAR-2-activating peptide SLIGRL-NH2(0.1 μM), and the reverse peptide analog LRGILS-NH2 (0.1 μM) on Isc after their contraluminal addition to porcine ileal mucosal sheets. Total flux area of chambers was 1 cm2. The records are representative of mucosal responses in 7 tissues from 6 pigs for trypsin, 21 tissues from 7 pigs for SLIGRL-NH2, and 3 tissues from 2 pigs for LRGILS-NH2. As an additional control, 10 μM carbachol (CCH) was added to the LRGILS-NH2-treated tissue to confirm tissue viability. Average respective changes in Isc and Gt produced by these substances were as follows: trypsin = 36 ± 6 μA cm−2 and −3 ± 1 mS · cm−2; SLIGRL-NH2= 13 ± 3 μA cm−2 and −6 ± 3 mS · cm−2; and LRGILS-NH2 = 0 ± 1 μA/cm−2 and 2 ± 1 mS · cm−2. mS, millisiemens.

Cumulative concentration-effect relationships for trypsin and SLIGRL-NH2 in porcine ileal mucosa. Trypsin was added to either the contraluminal (●) or luminal (○) aspects of mucosal sheets in increasing concentrations; SLIGRL-NH2 (▪) was only administered contraluminally. Each data point represents the mean ± S.E. of responses in 16 tissues from 8 pigs for contraluminal trypsin, 7 tissues from 3 pigs for luminal trypsin, and 14 tissues from 7 pigs for contraluminal SLIGRL-NH2. The mean 50% effective concentrations for contraluminal SLIGRL-NH2 and trypsin were 184 ± 67 and 769 ± 272 nM (P = .053, two-tailed unpaired t test).
Mucosal responses to the contraluminal addition of trypsin at 10 μM were reduced in tissues pretreated with 5 μg/ml SBTI added to the contraluminal bathing medium (mean ΔIsc after trypsin in absence and presence of SBTI = 33 ± 5 and 10 ± 6 μA/cm2, respectively; one tissue from each of three to four pigs, P = .01).
Mechanism of Trypsin and SLIGRL-NH2 Action.
To elucidate the mechanisms underlying the effects of trypsin and SLIGRL-NH2 on Isc, tissues were pretreated with Na+/K+/Cl− cotransport blocker furosemide, the neuronal conduction blocker saxitoxin, or the cyclooxygenase inhibitor indomethacin; these substances were added to the contraluminal aspect of mucosal sheets (Fig.3). Pretreatment of the tissues with 10 μM furosemide before trypsin or SLIGRL-NH2addition significantly decreased subsequent mucosal responses to trypsin by 82% and SLIGRL-NH2 by 51%. Saxitoxin at 0.1 μM reduced subsequent mucosal responses to trypsin by 84% and SLIGRL-NH2 by 80%. Indomethacin at 10 μM significantly decreased subsequent mucosal responses to trypsin by 97% and SLIGRL-NH2 by 49% (Fig. 3).

Effects of inhibitors on mucosal responses to contraluminal addition of 1 μM trypsin (top) or SLIGRL-NH2 (bottom). Tissues were contraluminally pretreated with furosemide (10 μM) saxitoxin (0.1 μM), or indomethacin (10 μM) for 5 min before the contraluminal addition of trypsin or SLIGRL-NH2. Dimethyl sulfoxide, the vehicle for indomethacin and furosemide, was without effect (data not shown). Each column represents the mean ± S.E. of Isc elevations in 6 to 11 tissues from 5 to 6 pigs. *P < .05 relative to agonist effect in tissue controls untreated with blockers, Tukey's test.
The effects of trypsin or SLIGRL-NH2 on Isc remained unaffected in tissues pretreated with the muscarinic cholinergic antagonist atropine at a contraluminal concentration of 0.1 μM. However, secretory responses to 10 μM carbachol, a cholinergic agonist, were absent in these tissues (Fig.4).

Effect of atropine on trypsin action. Representative chart tracings depict the lack of effect of atropine (0.1 μM, contraluminal addition) on mucosal Isc responses to the contraluminal addition of 1 μM trypsin (top). The effect of trypsin in the absence of atropine is shown (bottom) for comparison. Carbachol (CCH, 10 μM, contraluminal addition) was included at the end of each experiment to confirm blockade of muscarinic cholinergic receptors. Similar results were obtained with SLIGRL-NH2. Mean changes in Isc for trypsin alone and after atropine were 15 ± 2 and 15 ± 4 μA · cm−2, respectively (n = 7–8 tissues from three pigs). Mucosal responses to 1 μM SLIGRL-NH2 alone and in the presence of atropine were 9 ±1 and 8 ± 2 μA · cm−2, respectively (n= 6–8 tissues from four pigs).
The selective δ-opioid receptor agonist DPDPE had minimal effects on baseline Isc after its contraluminal addition at 0.1 μM, but it reduced mucosal responses to trypsin by 66%. This inhibitory action of DPDPE was not produced in tissues pretreated with an equimolar concentration of the selective δ-opioid receptor antagonist naltrindole (Fig. 5).

Inhibition of trypsin action by the δ-opioid agonist DPDPE and prevention of this effect by the δ-opioid antagonist naltrindole (NTI). Tissues were contraluminally pretreated with DPDPE (0.1 μM) 5 min before the contraluminal addition of trypsin. Some tissues were contraluminally pretreated with naltrindole (0.1 μM) 5 min before DPDPE addition (NTI/DPDPE). Each column represents the mean ± S.E. of Isc elevations in 10 to 16 tissues from 9 pigs. *P < .05 relative to trypsin effect in control tissues untreated with DPDPE or naltrindole, Tukey's test.
Immunohistochemical Localization of PAR-2 Immunoreactivity.
PAR-2 immunoreactivity was localized in neurons within the myenteric plexus and the external and internal submucosal plexuses. These neurons exhibited a relatively diffuse pattern of immunoreactivity compared with neurons displaying PGP 9.5 immunoreactivity. In all preparations, however, PAR-2-immunoreactive neurons and fibers also expressed PGP 9.5 immunoreactivity. Intense immunoreactivity for PAR-2 was observed in several neurons, although some neurons displayed relatively less intense PAR-2 immunoreactivity. Immunoreactivity for the cholinergic marker ChAT was highly colocalized with PAR-2 immunoreactivity in myenteric and submucosal neurons (Fig.6). In contrast, fibers immunoreactive for PAR-2 that innervated circular smooth muscle did not express ChAT immunoreactivity. In the ileal epithelium, intense PAR-2 immunoreactive cells were observed scattered in the villus epithelium; these had the appearance of open-type enteroendocrine cells and did not display ChAT immunoreactivity (Fig. 7A). Relatively thin and sometimes varicose PAR-2-immunoreactive nerve fibers were localized at the base of the villous epithelium (Fig. 7B). In comparison, ChAT-immunoreactive fibers were mostly situated in the midregion of villi; some of the thicker fibers also displayed PAR-2 immunoreactivity.
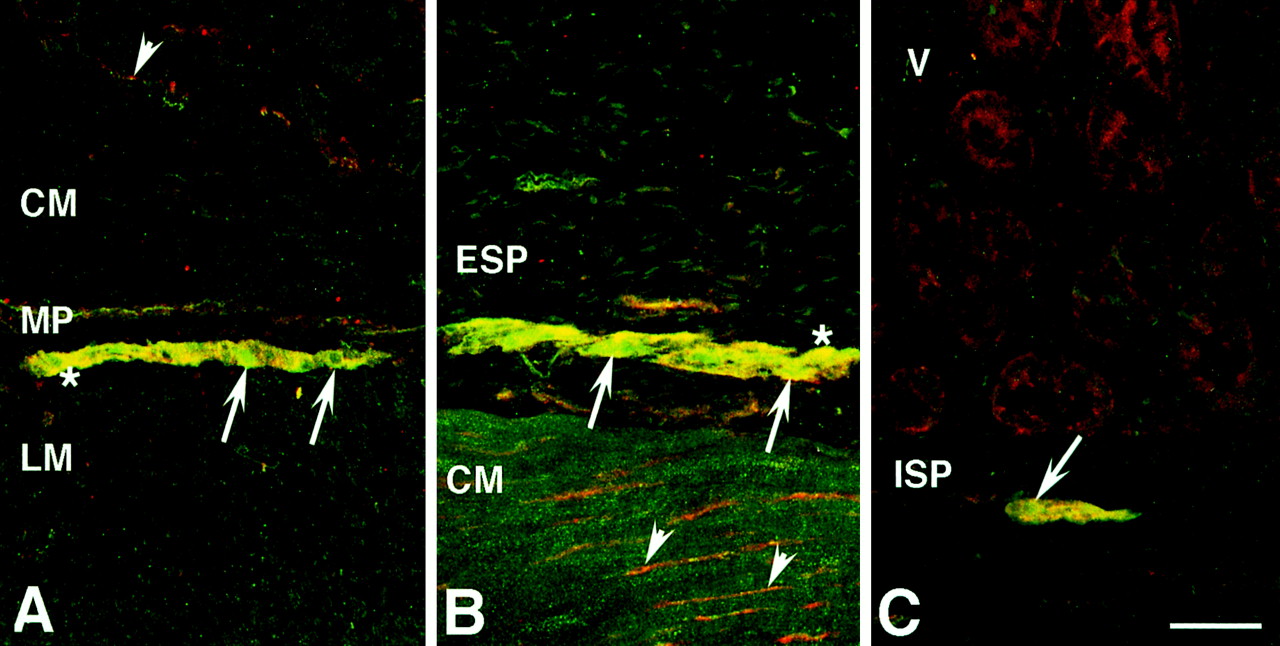
Localization of PAR-2 and ChAT immunoreactivities in porcine ileum. A, intense PAR-2 and ChAT immunoreactivities are colocalized in neurons (∗, yellow) of the myenteric plexus. Due to the diffuse nature of the immunoreactivity, relatively weak PAR-2 immunoreactivity is observed in some neurons (arrows). PAR-2-immunoreactive fibers (arrowhead, red) that are not colocalized with ChAT-immunoreactive fibers are seen in the circular muscle. B, cluster of large neurons in the ganglionated external submucosal plexus that exhibits colocalization (yellow hue) of PAR-2 (red) and ChAT (green) immunoreactivities. PAR-2 and ChAT immunoreactivities are highly colocalized (∗); however, weak PAR-2 immunoreactivity in the presence of ChAT immunoreactivity is also seen, which imparts a slightly greenish hue in some neurons (arrows). PAR-2-immunoreactive fibers (arrowheads) are seen in the circular muscle. C, colocalization of PAR-2 and ChAT immunoreactivities in neurons (arrow) of the external submucosal plexus. CM, circular muscle; ESP, external submucosal plexus; ISP, internal submucosal plexus; LM, longitudinal muscle; MP, myenteric plexus; V, villi. Scale bars in A to C, 50 μm.

Localization of PAR-2 and ChAT immunoreactivities in the porcine ileal submucosa. A, intense PAR-2 immunoreactivity is observed in enteroendocrine-like cells in the epithelium (arrow). ChAT immunoreactivity is not expressed by these cells. B, PAR-2-immunoreacive fibers are visualized at the base of the villous epithelium (arrows, red). Note that a separate population of ChAT-immunoreactive (arrowheads, green) fibers is seen in the midregion of the villi. PAR-2 and ChAT immunoreactivities are colocalized in a small population of thicker fibers (double arrow, yellow). V, villus. Scale bars in A and B, 50 μm.
Discussion
Our results show that application of the PAR-2 agonist SLIGRL-NH2 and trypsin to the contraluminal side of the porcine ileum produces increases in Isc that are due to anion secretion because furosemide, an inhibitor of basolateral chloride loading into epithelial cells, inhibited these effects. PAR-2 agonists stimulate active anion transport by a neurogenic mechanism because saxitoxin also inhibited this stimulation. Moreover, a large proportion of submucosal neurons and nerve fibers that were situated in proximity to the basolateral membrane of enterocytes expressed immunoreactive PAR-2. Mast cells containing tryptase are closely associated with nerve fibers in the normal and inflamed human intestine (Berin et al., 1999). Thus, our results support the hypothesis that tryptase, which is released from mast cells when they degranulate, cleaves PAR-2 on submucosal neurons to trigger the release of unidentified neurotransmitters that stimulate fluid and electrolyte secretion from enterocytes. This novel mechanism of regulation of anion secretion by proteases and their receptors may serve to protect the intestinal mucosa from pathogen invasion.
Trypsin and the PAR-2-specific-activating peptide SLIGRL-NH2 increased Isc after their contraluminal addition to ileal mucosa sheets with potencies that were at least an order of magnitude higher than reported for their actions in other intestinal and nonintestinal preparations in vitro (Nystedt et al., 1994; Santulli et al., 1995; Bohm et al., 1996a,b; Corvera et al., 1997; Vergnolle et al., 1998). Over the course of the entire investigation, however, mucosal responses to SLIGRL-NH2 tended to display some interanimal variability (cf. Figs. 2 and 3). Its actions were nevertheless specific because a peptide bearing a reversed amino acid sequence (LRGILS-NH2) was inactive. Although trypsin has been found to be more potent than SLIGRL-NH2 in several previous reports in vitro (Nystedt et al., 1994; Santulli et al., 1995; Bohm et al., 1996a,b; Corvera et al., 1997), its potency was not significantly different from that of SLIGRL-NH2 in the porcine ileal mucosa, a finding that may suggest that the activating sequence or ligand-binding domains of the porcine PAR-2 differ from that characterized in rodents or that a second subtype of SLIGRL-NH2-preferring PAR-2 is expressed in the intestine, as previously hypothesized (Vergnolle et al., 1998). Mucosal responses to trypsin or SLIGRL-NH2 at a fixed concentration did not differ after the contraluminal addition of these agonists at single or increasing cumulative concentrations. Although the effects of SLIGRL-NH2 were measured in porcine ileal mucosa pretreated with a protease inhibitor cocktail, a recent report suggests that SLIGRL-NH2 is not susceptible to rapid protease degradation. Its ability to alter ion transport in rat jejunum remained unaltered in the presence or absence of protease inhibitors (Vergnolle et al., 1998).
The Isc elevations occurring in response to the PAR-2 agonist SLIGRL-NH2 or by trypsin may be attributable to active anion secretion because the effects of trypsin and, to a lesser extent, SLIGRL-NH2 were markedly reduced by furosemide, an inhibitor of basolateral chloride loading in enterocytes. This finding is in agreement with that of Vergnolle et al. (1998) who reported that mucosal responses to SLIGRL-NH2 were attenuated in rat jejunal mucosal sheets bathed in chloride-free buffer. The furosemide-resistant portion of the Isc response to the PAR-2 agonists might be attributable to bicarbonate secretion or furosemide-insensitive chloride transport. In contrast to this previous report, the prosecretory actions of trypsin and SLIGRL-NH2 in porcine ileum were inhibited by saxitoxin, a neuronal conduction blocker. This result, in combination with our immunohistochemical data, supports the hypothesis that PAR-2 is present on submucosal neurons that release prosecretory neurotransmitter substances. The neural circuits expressing PAR-2 do not appear to contain muscarinic cholinergic synapses because mucosal responses to SLIGRL-NH2 or trypsin were unaltered by atropine, administered at a contraluminal concentration that was sufficient to antagonize the secretory actions of the cholinergic agonist carbachol (Chandan et al., 1991). It is possible that PAR-2 activation does not result in release of acetylcholine from submucosal neurons or that the neurotransmitter acts on postsynaptic nicotinic, rather than muscarinic, cholinergic receptors. The actions of trypsin were nearly abolished by the cyclooxygenase inhibitor indomethacin; those to SLIGRL-NH2 were less sensitive to this drug. The different sensitivities of trypsin and SLIGRL-NH2 to both indomethacin and furosemide may be a manifestation of an additional, prosecretory prostanoid component in trypsin action. Eicosanoids released by cells in the lamina propria play an important role in intestinal inflammation (Eberhart and Dubois, 1995). Some prostanoids may indirectly evoke mucosal secretion through interactions with enteric neurons (Bern et al., 1989). The prostanoid(s) involved in the PAR-2-mediated mucosal responses to trypsin should be identified in future investigations. With respect to the blocking agents used in these experiments, it should be noted that the apparent differences in the magnitude of their inhibitory effects on tissue responses to trypsin and SLIGRL may be the consequence of the use of these PAR-2 agonists at single concentrations. Although it would be desirable to assess the effects of blockers over the entire effective concentration ranges for each agonist, this approach was impractical with the present paradigm.
Opiates such as morphine and loperamide are effective in alleviating secretory diarrheas through their actions on enteric neurons that modulate intestinal motility and transepithelial ion transport (Brown, 1994). δ-Opioid receptors predominate in the porcine ileum (Brown et al., 1999). They are expressed on submucosal neurons and mediate the antisecretory effects of opiates (Quito and Brown, 1991; Brown et al., 1998). In this study, we addressed the hypothesis that submucosal opioid receptors inhibit the enteric neural pathways that trigger active anion secretion and are activated by inflammatory mediators. DPDPE, an agonist highly selective for δ-opioid receptors (Mosberg et al., 1983), significantly attenuated the secretory effect of trypsin. Furthermore, the inhibitory action of DPDPE was reduced by the highly selective δ-opioid receptor blocker naltrindole (Portoghese et al., 1988), a result indicating that it was mediated by δ-opioid receptors.
In rat jejunum, luminally applied trypsin is effective in stimulating mucosal transport (Kong et al., 1997). In porcine ileum, however, trypsin was more potent and effective in increasing Isc after its contraluminal addition to porcine mucosal sheets. Moreover, the colocalization of PAR-2 immunoreactivity with the neuronal marker PGP9.5 within the submucosal plexus supports our functional observations that PAR-2 is present on enteric neurons. However, the apparent localization of PAR-2 immunoreactivity in enteroendocrine cells suggests that this receptor may regulate the release of GI hormones under the influence of luminal trypsin. The present results clearly indicate that the cellular expression of PAR-2 in the small intestine may vary according to the species or intestinal segment examined.
Human intestinal mast cells are concentrated in the intestinal submucosa, where they secrete tryptase (Aldenborg and Enerbäck, 1994). Trypsin and tryptase have a similar catalytic activity, and neuronal PAR-2 may be in juxtaposition to mast cells in the ileal submucosa to permit their activation by tryptase (Molino et al., 1997). Indeed, nearly 90% of mast cells in the human intestine are within 2 μm of neurons, and recent evidence indicates that there are direct interactions between nerves and mast cells (Suzuki et al., 1999). Neuronal PAR-2 may function in intestinal host defense by mediating active anion secretion that could provide the means to rid the intestinal mucosa of potentially pathogenic microorganisms. Opiates, particularly those interacting with δ-opioid receptors, may be clinically beneficial in alleviating diarrheas associated with PAR-2 activation and other intestinal inflammatory states.
Acknowledgment
We thank Dr. Morley D. Hollenberg for the generous gift of anti-PAR-2 antiserum.
Footnotes
-
Send reprint requests to: David R. Brown, Ph.D., Department of Veterinary PathoBiology, University of Minnesota, 1988 Fitch Ave., St. Paul, MN55108-6010. E-mail: brown013{at}tc.umn.edu
-
↵1 This study was funded in part by National Institutes of Health Grant DA-10200 (to D.R.B.) and NIH Grants DK-57840, DK-39957, and DK-43207, and an award from the Crohn's and Colitis Foundation of America (to N.W.B). Salary support for B.T.G. and A.K-N. was provided by Alcohol, Drug Abuse, and Mental Health Administration/National Institute on Drug Abuse Psychoneuroimmunology and Substance Abuse training Grant T32 DA07239.
- Abbreviations:
- GI
- gastrointestinal
- PAR
- proteinase-activated receptor
- DPDPE
- [d-Pen2,5]-enkephalin
- Isc
- short-circuit current
- Gt
- tissue conductance
- SBTI
- soybean trypsin inhibitor
- ChAT
- choline acetyltransferase
- PGP9.5
- protein gene product 9.5
- Received April 11, 2000.
- Accepted June 9, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}