


Abstract
YM976 is a novel and specific phosphodiesterase 4 inhibitor. In our previous report, we indicated that YM976 has less emetogenicity, a major adverse effect of PDE4 inhibitors, than rolipram. In the present study, we examined the antiasthmatic effects of YM976 in guinea pigs. YM976 orally administered exhibited inhibition of antigen-induced bronchoconstriction, airway plasma leakage, airway eosinophil infiltration, and airway hyperreactivity (AHR), with ED50values of 7.3, 5.7, 1.0, and 0.52 mg/kg, respectively. Rolipram also dose dependently suppressed these responses. Prednisolone suppressed eosinophil infiltration and AHR, whereas it failed to inhibit bronchoconstriction and plasma leakage. Theophylline moderately suppressed bronchoconstriction and edema, but neither eosinophil infiltration nor AHR. YM976 suppressed the peroxidase activity in the bronchoalveolar lavage fluid, and elevated the intracellular peroxidase activity and cAMP contents of infiltrated cells, suggesting that YM976 inhibited not only the infiltration but also the activation of leukocytes. In vitro studies revealed that YM976 potently suppressed eosinophil activation (EC30 = 83 nM), and exerted a little relaxation on LTD4-precontracted tracheal smooth muscle (EC50 = 370 nM). Rolipram exhibited a potent tracheal relaxation activity (EC50 = 50 nM). In vivo studies indicated that the inhibitory effect of YM976 on LTD4-induced bronchospasm was marginal even at 30 mg/kg p.o., although rolipram significantly inhibited the bronchospasm at the same dose. These results suggested that YM976, unlike rolipram, showed the inhibition of antigen-induced airway responses due to anti-inflammatory effects, but not to direct tracheal relaxation. In conclusion, YM976 may have potential therapeutic value in the treatment of asthma through its anti-inflammatory activities.
Bronchial asthma is characterized by reversible airway obstruction, inflammation, and airway hyperresponsiveness. One of the most important therapies for asthma is the inhibition of bronchoconstriction induced by antigen. Treatment with bronchodilators such as theophylline, which is a nonselective phosphodiesterase (PDE) inhibitor, is recommended in almost all cases of asthma.
Eosinophils, which are mainly related the inflammation of asthma, contain PDE4 isozyme, and inhibition of this enzyme is associated with the increase of the intracellular cAMP level and causes suppression of the activities of eosinophils (Giembycz and Dent, 1992). Tracheal smooth muscle cells contain PDE3 and PDE4 isozymes that have an important function as modulators of contractility (Bernareggi et al., 1999). Based on the putative role of cAMP in inflammatory cells and airway smooth muscle, PDE4 has been identified as a molecular target for novel antiasthmatic agents (Barnette, 1999).
YM976, 4-(3-chlorophenyl)-1,7-diethylpyrido[2,3-d]pyrimidin-2(1H)-one, is a novel, specific, and potent PDE4 inhibitor (Aoki et al., 2000b) with a chemical structure different from that of rolipram-like compounds. YM976 has a potent inhibitory activity for PDE4 (IC50 = 2.2 nM) as well as showing no inhibition against other PDE isozymes at 3 pM. Interestingly, YM976 showed anti-inflammatory activities in ferrets at doses causing none of the emesis that is a main adverse effect of PDE4 inhibitors (Aoki, 2000a). Thus, in YM976, the anti-inflammatory effects are dissociated from emetogenicity.
Antigen inhalation to the sensitized guinea pigs produced immediate bronchoconstriction, airway plasma leakage, airway hyperreactivity, and inflammatory cell infiltration. These events in guinea pigs have been used for the assessment of various candidates of antiasthmatic agents. In the present studies, whose aim was to throw light on the potential of YM976 as an antiasthmatic drug, we evaluated its effects on antigen-induced airway responses in guinea pigs. Additionally, the effect of YM976 on the activation of infiltrated leukocytes was examined in vivo. Its in vitro effects were also evaluated in guinea pig eosinophils and tracheal preparations. In some experiments, we compared the effects of YM976 with those of the other PDE4 inhibitors rolipram and RP73401, and those of the existing antiasthmatic agents prednisolone and theophylline.
Materials and Methods
Animals.
Male Hartley guinea pigs weighing 500 to 700 g were purchased from SLC (Hamamatsu, Japan) or Charles River Japan (Atsugi, Japan). The animals were maintained in ordinary animal cages in a constant 12-h light/dark cycle. Food and water were available ad libitum.
Chemicals.
YM976, (±)-rolipram, and RP73401 were synthesized by the Department of Chemistry, Yamanouchi Pharmaceutical Co. Ltd. (Tsukuba, Japan). Theophylline and prednisolone were purchased from Sigma Chemical Co. (St. Louis, MO). In in vitro experiments, all compounds were dissolved in dimethylsulfoxide (DMSO), and the final concentration of DMSO was less than 0.1%. In in vivo experiments, all compounds were suspended in 0.5% methylcellulose solution, and were administered orally at a volume of 3 ml/kg to animals that were fasted for 16 h. Animals in ovalbumin (OA) and saline groups received only the vehicle.
The reagents and chemicals used were pyrilamine maleate salt, histamine, acetylcholine (ACh), cytochrome c, superoxide dismutase, fMLP, OA grade V or VI, EDTA, Hanks' balanced salt solution without calcium chloride or magnesium sulfate, Krebs-Henseleit buffer, galamine, DMSO, and urethane were purchased from Sigma Chemical Co. Methylcellulose (TC-5E) was purchased from Shin-Etsu Chemical Co. (Tokyo, Japan), HEPES from Life Technologies (Rockville, MD), LTD4 from Funakoshi Co., Ltd. (Tokyo, Japan), diethyl ether and chloroform from Kanto Chemicals (Tokyo, Japan), aluminum hydroxide (Alum, Imject) from Pierce (Rockford, IL), heparin from Shimizu Seiyaku (Shimizu, Japan), pentobarbital sodium from Nacalai Tesque (Kyoto, Japan), and Diff-Quik from International Reagents Corp. (Kobe, Japan).
Sensitization of Guinea Pigs.
Male Hartley guinea pigs weighing 500 to 700 g (SLC) were actively sensitized by the intraperitoneal injection of OA (5 μg) and alum (1 mg) in saline (0.5 ml). Sensitizations were performed three times at 7- to 12-day intervals.
Antigen-Induced Bronchoconstriction in Anesthetized Sensitized Guinea Pigs.
Ten days after the final sensitization, the animals were anesthetized with urethane (1.2–1.5 g/kg i.p.). The trachea was cannulated to allow mechanical ventilation (Harvard 683 rodent ventilator; Harvard Bioscience, South Natick, MA) at 60 strokes/min with a stroke volume of 1 ml/100 g of body weight. The animals were secured in the plethysmograph box connected to the transducer and the ventilator just mentioned for measurement of the airway resistance. Bronchoconstriction was induced by 0.5% OA inhalation for 5 min with an ultrasonic nebulizer (NE-U12; Omron, Kyoto, Japan). Lung resistance (Rl) was monitored and calculated for 15 min after the initiation of OA inhalation with a pulmonary function analyzer (model P; Buxco Electronics Inc., Sharon, CT). Each compound was administered orally 30 min before the antigen challenge.
Antigen-Induced Plasma Leakage in Sensitized Guinea Pigs.
Ten to 14 days after the final sensitization, the animals were pretreated with an antihistamine agent, pyrilamine (2 mg/kg i.v.), to avoid anaphylactic shock. Ten minutes later, they were placed in an exposure chamber connected to the output of a glass nebulizer (Kinoshita, Tokyo, Japan) driven by compressed air at 100 ml/s. The nebulizer chamber was filled with 0.5% OA (control and compound-treated groups) or saline (saline group), and antigen inhalation was performed for 30 min. To evaluate the plasma leakage, Evans blue (40 mg/kg i.v.) was injected 5 min before the challenge. Each compound was administered orally 30 min before the antigen inhalation.
Thirty minutes after the end of inhalation, the animals were sacrificed by cervical dislocation and exsanguinated by cutting the cervical blood vessels. Bronchial tissue samples were taken and the Evans blue dye was extracted and quantified as previously described (Katayama et al., 1978).
Antigen-Induced Cell Infiltration into the Lungs in Sensitized Guinea Pigs.
The sensitized guinea pigs were pretreated with pyrilamine and challenged with antigen (0.5% OA) for 30 min by the same method as was used in the leakage experiments.
Twenty-four hours after the antigen challenge, the guinea pigs were sacrificed and the lungs were lavaged three times with 10 ml of saline containing heparin (1 unit/ml). The recovered lavage samples were divided into three fractions. One fraction was placed into boiled water for 2 min, and then cooled in ice and centrifuged at 10,000gfor 30 min at 4°C. The cAMP contents of the supernatant (pmol/106 cells) were measured using a cAMP enzyme immunoassay system (Amersham Pharmacia Biotech, Uppsala, Sweden). Another fraction was cooled on ice, and centrifuged at 200g for 10 min at 4°C. The supernatant peroxidase activity of BAL fluids and the intracellular peroxidase activity of infiltrated leukocytes were measured by the spectrophotometric reduction of cytochrome c. The data were indicated as the optical density value at 550 nM (O.D.550) value/106 cells.
The other fraction was used for counting the number of leukocytes. The total number of leukocytes was counted using a cell counter (Celltac-α; Nippon Kohden, Tokyo, Japan). At least 300 cells were differentiated according to standard morphological criteria (eosinophils, mononuclear cells, and neutrophils) using Cytospin preparations stained with Diff-Quik (International Reagents Corp.). In addition, basal cell accumulation was determined in saline-challenged animals. For each treatment group, results were expressed as the percentage inhibition of cell influx compared with that in the vehicle control group. Compounds were administered orally 30 min before and 3 h after the challenge.
Antigen-Induced AHR and Cell Infiltration in Sensitized Guinea Pigs.
The sensitization method described above could not elicit the apparent AHR in guinea pigs (data not shown), and so we carried out the evaluation using a different method of sensitization and challenge.
Male Hartley guinea pigs were actively sensitized by intraperitoneal injection of OA (10 μg) and alum (10 mg) in saline (0.5 ml). Seven days later, 5% OA inhalation for 5 min was performed three times at 7- to 14-day intervals. Seven days after the final inhalation, the animals were pretreated with pyrilamine (2 mg/kg i.v.), and 10 min later, they were challenged by the inhalation of 0.5% OA for 30 min by the same method as is described above. Twenty-four hours after the challenge, the animals were anesthetized with urethane and the trachea was cannulated to allow mechanical ventilation at 60 strokes/min with a stroke volume of 10 ml/kg of body weight. After galamine was injected (1 mg/kg i.v.) to eliminate spontaneous respiration and stabilize the ventilation, the airway reactivity was determined by subsequent inhalations of saline and each concentration of ACh (1, 3, 10, 30, and 100 mg/ml in saline) using a nebulizer (NE-U12; Omron). Intra-airway pressure (PIP; cm H2O) was monitored with a low-pressure transducer (TP-101; Nihon Kohden) connected to the sidearm of a tracheal cannula. Data were measured using carrier-amplitude amplifiers (AP-620G; Nihon Kohden) and recorded on a Mini-polygraph (RM-6100; Nihon Kohden) and on a Macintosh computer by a MacLab/8 software system (MKIII; AD Instruments, Castle Hill, Australia). The change in PIP induced by ACh inhalation was measured to represent the airway reactivity, and PIP at the basal level was given by the inhaled saline. Data were expressed as the concentrations at which PIP reached a value two times the basal level (log10PC200). Five minutes after the highest concentration (100 mg/ml) of ACh inhalation, the lungs were lavaged three times with 10 ml of saline containing heparin (1 unit/ml) through the cannula. The lavage fluid was centrifuged at 200g for 10 min at 4°C, and the cell pellet was resuspended in 1 ml of saline. The total leukocyte number was counted with Celltac-α, and the cell differentiation was determined by the method described above. Compounds were administered orally 30 min before and 3 h after the antigen challenge.
Superoxide Production from Isolated Guinea Pig Eosinophils.
Guinea pig eosinophils were isolated by a previously reported method (Souness et al., 1995) and the discontinuous density gradient method. Eosinophils (1 × 106/ml) were suspended in Hanks' balanced salt solution containing 20 mM HEPES, 0.1% gelatin, and 150 μM cytochrome c. Superoxide generation was initiated by the addition of fMLP (50 nM), and was measured by the spectrophotometric reduction of cytochrome c. Results were determined by the change in the absorbance at 550 nm over 40 min after the initiation.
LTD4-Induced Constriction of Isolated Bronchial Smooth Muscle from Guinea Pigs.
The method of isolating tracheal smooth muscle was essentially the same as that described previously (Baba et al., 1986). Guinea pig trachea was transferred into cold oxygenated Krebs' solution, and the connective tissue was carefully cleaned. The trachea was opened by a longitudinal cut through the cartilaginous region diametrically opposite the tracheal smooth muscle, and divided into four strips containing five or six cartilages. A strip was suspended vertically under 1-g tension in Krebs' solution (37°C, pH = 7.4) continuously gassed with 5% CO2and 95% O2 in a 10-ml organ bath. Carbachol (3 μM) was added to the bath, and after the contractile response had reached a plateau the tissues were washed four times over a 15-min period and allowed to equilibrate for a further 30 min.
Relaxation activity was assessed in tracheas precontracted with LTD4, which was determined as the concentration indicating the submaximal contraction from the preliminary experiment (data not shown). Following the equilibration period, the bronchial strip was contracted with 100 nM LTD4. After the contraction plateau was reached, the concentration-response curves for YM976 and the other compounds were established. After the maximal effect of each concentration was obtained, isoproterenol (1 μM) was added to determine the maximal relaxation response.
LTD4-Induced Bronchospasm in Anesthetized Guinea Pigs in Vivo.
Male Hartley guinea pigs were anesthetized with urethane. The trachea was cannulated to allow mechanical ventilation at 60 strokes/min with a stroke volume of 10 ml/kg of body weight. The changes in airway resistance were measured with the Buxco pulmonary function analyzer.
A cannula was tied into the right jugular vein to allow the injection of spasmogen solution. After the connection of the animals to ventilators, histamine (3 μg/kg) was injected intravenously twice with a 5-min interval to stabilize the responses. After the recovery to the basal level of airway resistance, LTD4 (0.3 μg/kg) was intravenously injected, and the change of Rl was monitored from 2 min before to 5 min after the LTD4injection. YM976 and the other compounds were orally administered 30 min before the first injection of histamine. Airway contraction was determined as the change from the basal value.
Data Analysis.
Data were expressed as the mean ± S.E. or means with 95% confidence limits. Statistical significance of differences between means of groups was determined by Dunnett's multiple range test or Student's t test, and probabilities of <0.05 were considered significant. Concentrations or doses causing 50 or 30% inhibition were determined by nonlinear curve fitting using a SAS system (SAS Institute Inc., Cary, NC).
Results
Effects on Antigen-Induced Bronchoconstriction.
Aerosolized OA caused an immediate bronchoconstriction, which peaked within 15 min in sensitized guinea pigs. Inhaled antigen increased Rl from 0.157 to 0.499 cm H2O/ml/s in the OA group (n = 7). On the other hand, in the saline group, Rl increased from 0.163 to 0.174 cm H2O/ml/s (n = 6). YM976 showed a dose-dependent inhibition of Rl increases, with an ED50 value of 7.3 mg/kg p.o. (Table 1; Fig.1). Rolipram and theophylline also dose dependently inhibited the bronchoconstriction with ED50 values of 7.7 and 67 mg/kg, respectively, whereas prednisolone did not cause suppression even at 100 mg/kg p.o. RP73401, one of the strongest PDE4 inhibitors, showed 68% inhibition at 1 mg/kg p.o.
Doses of YM976, rolipram, theophylline, and prednisolone effective against antigen-induced immediate bronchoconstriction, airway plasma leakage, and lung eosinophil infiltration in sensitized guinea pigs

Effects of YM976, rolipram, theophylline, prednisolone, and RP73401 on antigen-induced bronchoconstriction in sensitized guinea pigs. Each compound was administered orally 30 min before OA inhalation. Results are represented as the maximal Rl value during 15 min monitored after OA inhalation. Data are expressed as the mean ± S.E. of five to eight animals.###p < 0.001, significant difference between the saline and OA groups (Student's t test). *p < 0.05 and ***p < 0.001: significant differences between the OA group and the relevant compound treatment group (Dunnett's multiple range test).
Effects on Antigen-Induced Airway Plasma Leakage.
Antigen inhalation by the sensitized guinea pigs significantly increased airway plasma leakage, which was determined as the amount of dye (Evans blue) extravasation. In the saline group, the amount of dye was between 56 and 72 ng/mg tissue, and in the OA group, it was between 206 and 244 ng/mg tissue. YM976 dose dependently suppressed the increased extravasation of dye with an ED50 value of 5.7 mg/kg p.o. (Table 1; Fig. 2). Rolipram and theophylline also inhibited the extravasation, with oral ED50 values of 3.0 and 16 mg/kg, respectively. On the other hand, prednisolone failed to show significant inhibition up to 100 mg/kg p.o., and the maximal inhibition ratio was 46%.

Effects of YM976, rolipram, theophylline, and prednisolone on increases in antigen-induced microvascular permeability in sensitized guinea pigs. Each compound was administered orally 30 min before OA inhalation. Extravasation of Evans blue dye in intrapulmonary airways was measured. Data are expressed as the mean ± S.E. of five to seven animals. ##p < 0.01; and###p < 0.001, significant differences between the saline and OA groups (Student's t test). *p < 0.05, **p < 0.01, and ***p < 0.001, significant differences between the OA group and the relevant compound treatment group (Dunnett's multiple range test).
Effects on Antigen-Induced Cell Infiltration into the Lungs.
In the guinea pigs, OA inhalation caused increases in total leukocyte numbers in BAL fluid 24 h after the challenge. YM976 at oral doses of 0.3 to 10 mg/kg significantly and dose dependently inhibited eosinophil infiltration, with an ED50 of 1.0 mg/kg (Table 1; Fig. 3). Rolipram and prednisolone also dose dependently inhibited the cell infiltration, with oral ED50 values of 3.6 and 5.1 mg/kg, respectively. Theophylline did not achieve 50% suppression on the infiltration up to 100 mg/kg p.o.
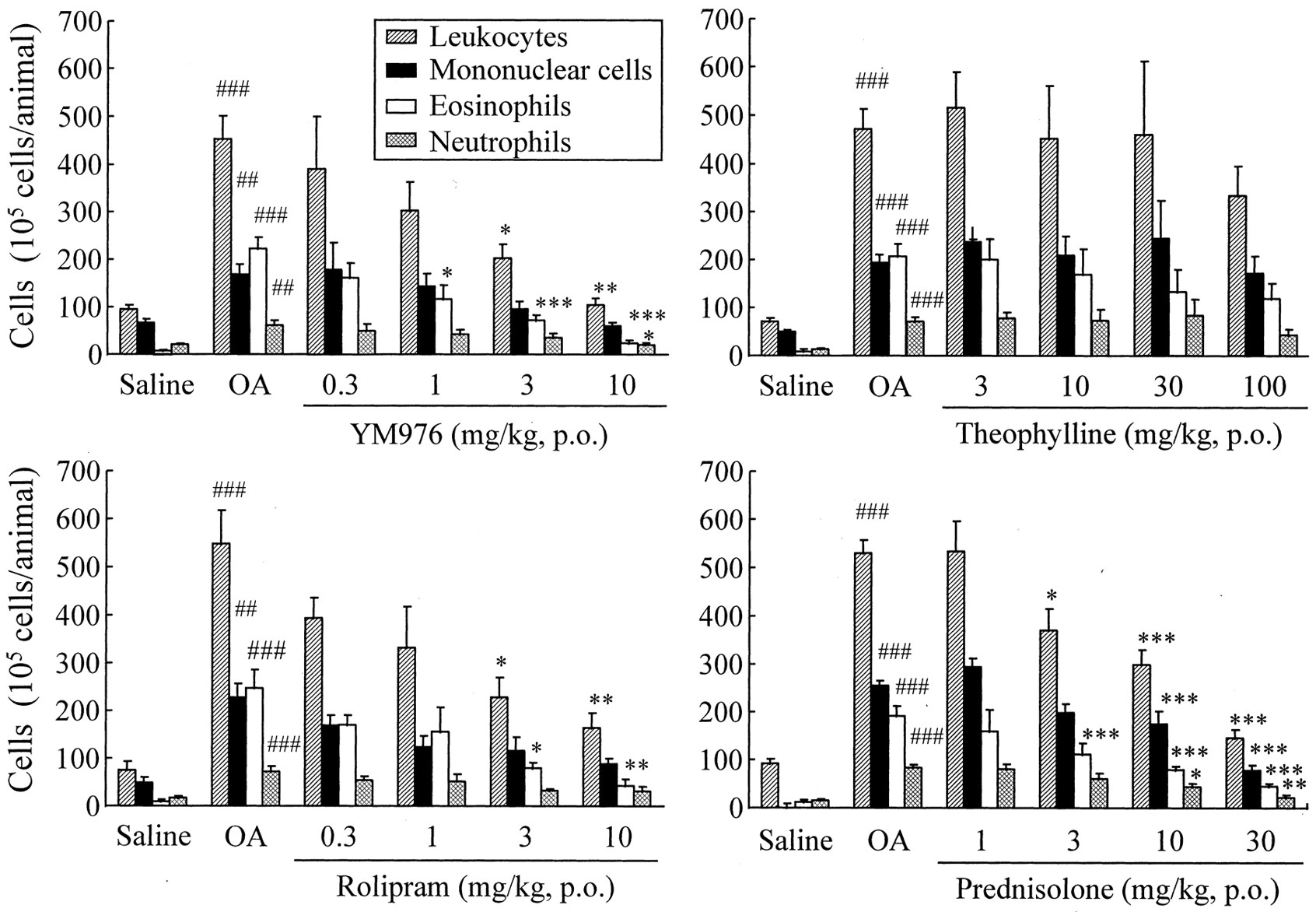
Effects of YM976, rolipram, theophylline, and prednisolone on antigen-induced inflammatory cell infiltration into the lungs in sensitized guinea pigs. Each compound was administered orally 30 min before and 3 h after antigen challenge. Data are expressed as the mean ± S.E. of five to seven animals.##p < 0.01 and###p < 0.001, significant differences between the saline and OA groups (Student's t test). *p < 0.05, **p < 0.01, and ***p < 0.001, significant differences between the OA group and the relevant compound treatment group (Dunnett's multiple range test).
In the same experiment, we measured the peroxidase activities and cAMP contents to examine the infiltrated inflammatory cell activation. YM976 inhibited the peroxidase activity in the supernatant of BAL fluid and elevated the intracellular peroxidase activity of the infiltrated leukocytes. YM976 also increased intracellular cAMP contents of the infiltrated cells (Fig. 4).

Effect of YM976 on peroxidase activity and intracellular cAMP content in antigen-induced cell infiltration experiments. Peroxidase activity was expressed as the reduction of cytochrome c (O.D. values at 550 nM). A, peroxidase activity in the supernatant of BAL fluid containing 106cells. B, intracellular peroxidase activity of the infiltrated cells. C, intracellular cAMP content of the infiltrated cells (106cells). Cell infiltration into the lungs was induced by antigen inhalation in sensitized guinea pigs. YM976 was administered orally 30 min before and 3 h after antigen challenge. Data are expressed as the mean ± S.E. of five to seven animals.###p < 0.001, difference between the saline and OA groups (Student's t test). *p < 0.05, significant difference between OA group and the relevant compound treatment group (Dunnett's multiple range test).
Antigen-Induced AHR and Cell Infiltration.
Sensitization with one intraperitoneal injection and three inhalations of antigen resulted in the induction of AHR to ACh. YM976 was orally administered 30 min before and 3 h after the final antigen inhalation, and it dose dependently and significantly shifted the PIP curve to the right between 1 and 10 mg/kg p.o. (Fig. 5). YM976 and rolipram dose dependently increased the PC200 value, with oral ED50 values of 0.52 and 2.7 mg/kg, respectively (Table 2). Prednisolone increased the PC200 value, but not to a statistically significant extent, with an ED50 value of 24 mg/kg p.o.. In the same experiment, YM976, rolipram, and prednisolone also dose dependently inhibited the eosinophil infiltration, with ED50 values of 1.8, 8.6, and 16 mg/kg, respectively (Table 2).

Effect of YM976 on antigen-induced airway hyperreactivity (A and B) and inflammatory cell infiltration (C). Sensitized guinea pigs were exposed to antigen inhalation for 30 min. Twenty-four hours later, airway reactivity was determined by subsequent inhalation of saline and various concentrations of ACh (1, 3, 10, 30, and 100 mg/ml) under anesthesia. Five minutes after 100 mg/ml ACh inhalation, the lungs were lavaged. YM976 was administered orally 30 min before and 3 h after inhalation. Data are expressed as the mean ± S.E. #p < 0.05,##p < 0.01, and###p < 0.001, significant difference between the saline and OA groups (Student's t test). *p < 0.05, **p < 0.01, and ***p < 0.001, significant differences between the OA group and the relevant compound treatment group (Dunnett's multiple range test, n = 9–10).
Doses of YM976, rolipram, theophylline, and prednisolone effective against antigen-induced airway hyperreactivity and inflammatory cell infiltration in guinea pigs
Inhibitory Effects on Superoxide Generation from Isolated Eosinophils.
Results were expressed as the concentrations that induced 30% inhibition (EC30), because it was reported that the superoxide generation from guinea pig eosinophils stimulated by fMLP and zymosan was inhibited by a maximum of 60% by various PDE4 inhibitors (Barnette et al., 1995). YM976, rolipram, and RP73401 dose dependently inhibited fMLP-induced superoxide generation, with EC30 values of 83, 500, and 7.6 nM, respectively (Table 3).
In vitro effects of YM976, rolipram, RP73401, and theophylline on superoxide generation from guinea pig eosinophils and relaxation activity on precontacted guinea pig tracheal smooth muscle
Relaxation Effects on LTD4-Induced Tracheal Contraction in Vitro.
LTD4 at 30 nM induced the contraction of tracheal smooth muscle in vitro. YM976 elicited concentration-dependent relaxation, with an EC50value of 370 nM (Fig. 6; Table 3). Rolipram and RP73401 showed potent relaxation effects, with EC50 values of 50 and 6.8 nM, respectively. Theophylline also relaxed the tracheal contraction, but the effect was much weaker (EC50 = 5000 nM) than that of the other compounds.

In vitro relaxation activities of YM976 (●), rolipram (■), RP73401 (○), and theophylline (▾). Relaxation activity was assessed in tracheas precontracted with LTD4. Following the equilibration period, the bronchial strip was contracted with 100 nM LTD4. After the contraction plateau was reached, the concentration-response curves for YM976 and the other compounds were established. After the maximal effect of each concentration was obtained, isoproterenol (1 pM) was added to determine the maximal relaxation response.
Effects on LTD4-Induced Bronchospasm in Vivo.
Intravenous LTD4 injection (0.3 μg/kg) produced an evident increase in Rl, which reached 0.256 cm H2O/ml/s. YM976 showed little suppression of bronchospasm induced by LTD4 whose inhibitory ratio is 28% at 30 mg/kg p.o. (Fig. 7). On the other hand, rolipram, theophylline, and RP73401 significantly inhibited the response at 30, 100, and 3 mg/kg p.o., respectively.

Dilatory effects on LTD4-induced bronchospasm in guinea pigs. LTD4 was injected intravenously at a dose of 0.3 pg/kg. The mean of the Rl basal values was 0.102 ± 0.003 cm H2O/ml/s, and the differences among the groups were not significant (p > 0.38, one-way ANOVA). All other compounds were administered orally 30 min before the first histamine injection. Data are expressed as mean ± S.E. of changes from the basal value of five to six animals. **p < 0.01 and ***p < 0.001, significant differences from the control group and the relevant compound treatment group (Dunnett's multiple range test). Theo, theophylline.
Discussion
Much evidence has appeared in the literature suggesting that PDE4 inhibitors might become potent novel therapeutic agents for bronchial asthma (Barnette, 1999). Indeed, the primary target disease of most PDE4 inhibitors under clinical development is bronchial asthma. Unfortunately, some existing PDE4 inhibitors tend to cause emesis at the same doses that cause anti-inflammatory effects (Heaslip and Evans, 1995), and the efficacy might be limited. A new PDE4 inhibitor is required that can provide therapeutic benefit without causing emesis. In our previous report (Aoki et al., 2000a), we indicated that YM976 was an orally active and selective PDE4 inhibitor with an apparent dissociation from emetic activity. In the present study, we evaluated the antiasthmatic effects of YM976 on some experimental guinea pig models in which symptoms were elicited by antigen inhalation.
YM976 suppressed antigen-induced immediate bronchoconstriction in a dose-dependent manner. To investigate the mechanism of action, the bronchodilator effect of YM976 was assessed in vitro and in vivo. YM976 exerted only a weak relaxation of precontracted smooth muscle (EC50 = 370 nM), although it effectively suppressed eosinophil superoxide generation (EC50= 83 nM), which could have induced the bronchoconstriction (Sagai et al., 1996; De Boer et al., 1998). Rolipram also inhibited antigen-induced bronchoconstriction. However, unlike YM976, rolipram showed a potent relaxation of precontracted bronchial smooth muscle in vitro (EC50 = 50 nM) compared with the inhibitory activity of superoxide generation (EC50 = 500 nM). Indeed, rolipram significantly inhibited LTD4-induced bronchospasm potently in guinea pigs at 30 mg/kg p.o., whereas YM976 showed slight inhibition for the bronchospasm. RP73401 also showed a potent in vivo inhibition of bronchoconstriction induced by antigen and LTD4(ED50 < 3 mg/kg p.o.) and a strong relaxation effect in vitro (EC50 = 6.8 nM). These results were supported by the report showing that rolipram and RP73401 relaxed guinea pig isolated tracheal preparations at a basal tone, and showed in vivo bronchodilator effects (Raeburn et al., 1994; Turner et al., 1996). PDE4 inhibitors were reported to inhibit the release of spasmogens, such as LTs (Heaslip et al., 1992; Howell et al., 1994) and thromboxane A2 (Uhlig et al., 1997). Indeed, YM976 and rolipram inhibited fMLP-induced LTC4/D4/E4release from human eosinophils with IC50 values of 3.9 and 12 nM, respectively (M. Aoki, unpublished data, 2000). It is most likely that YM976 suppresses antigen-induced bronchoconstriction through inhibition of the spasmogen release from the leukocytes rather than by any direct relaxation effect. On the other hand, rolipram and RP73401 may show the inhibition on the bronchoconstriction via the direct dilator activity as well as the inhibition of leukocytes.
Airway edema resulting from plasma leakage induced by antigen is an important inflammatory event in asthma (Persson, 1988). YM976 and rolipram dose dependently inhibited antigen-induced plasma leakage. Although the inhibitory mechanisms of YM976 have not been elucidated, rolipram is reported to inhibit platelet-activating factor-induced plasma leakage in guinea pigs at least partially by inhibiting contraction of the endothelial cells (Ortiz et al., 1993).
Accumulation in the lungs of inflammatory cells such as eosinophils is also known to be an important event in the development of bronchial asthma. The drugs that can inhibit inflammatory cell accumulation in the lungs are useful therapeutic agents for bronchial asthma. We assessed the effects of YM976 on the antigen-induced cell infiltration into the lungs in the sensitized guinea pigs. YM976 markedly decreased the eosinophil infiltration into the lungs 24 h after the challenge with an ED50 value of 1.0 mg/kg p.o. The precise mechanisms by which YM976 inhibits eosinophil infiltration are still unclear. However, the pharmacological actions of PDE4 inhibitors described in numerous studies could explain the inhibitory mechanisms of YM976. PDE4 inhibitors can act in several ways to reduce cell infiltration into the lungs: 1) suppressing eosinophil chemotaxis (Alves et al., 1996); 2) decreasing the survival of eosinophils (Hallsworth et al., 1996); 3) inhibiting the differentiation of eosinophil precursors such as IL-5 inhibition (Kaminuma et al., 1996); 4) suppressing other cell types such as monocytes, which can release mediators acting as chemoattractants for eosinophils (Nourshargh., 1993); 5) inhibiting the expression of adhesion molecules (Berends et al., 1997); and 6) inhibiting cytokine production and T-cell proliferation (Giembycz et al., 1996). Pulmonary vasculature and endothelium are other potential sites of action. The endothelial cells express PDE4 (Lugnier and Schini, 1990) and increased levels of cAMP reduce the expression of the adhesion molecules (Blease et al., 1998). YM976 inhibits tumor necrosis factor-α production (Aoki et al., 2000b), which can induce intercellular adhesion molecule-1 and endothelial-leukocyte adhesion molecule-1 expression (Wellicome et al., 1990). However, it is not immediately obvious which mechanism is the most likely to be of primary importance, since PDE4 has been identified in many kinds of inflammatory cells.
Recent clinical studies with humanized monoclonal antibodies to IL-5 demonstrated that blocking IL-5 had no effects on immediate or late responses to allergens, or on AHR, although it reduced eosinophil recruitment into the airways (Barnes, 1999). These results suggested that the inhibition of eosinophil infiltration itself was not sufficient to reduce asthma symptoms significantly, although eosinophils were strongly related to the development of asthma. The inhibition of functions of remaining eosinophils in the airway may be also necessary to exert antiasthmatic effects. Therefore, we examined the effect of YM976 on the activation of eosinophils in vivo and in vitro. In the cell infiltration experiment using guinea pigs, YM976 suppressed the peroxidase activity in the supernatants of BAL fluid derived from infiltrated leukocytes, including eosinophils. Simultaneously, YM976 increased the intracellular peroxidase activity of the infiltrated leukocytes. Since YM976 increased the cAMP contents of the infiltrated cells, it is likely that YM976 inhibited peroxidase release from leukocytes by the elevation of intracellular cAMP. As described above, an in vitro experiment indicated that YM976 suppressed the eosinophil activation. These results suggested that YM976 suppressed not only eosinophil infiltration into the lungs but also the activation of eosinophils in the airways.
YM976 inhibited the antigen-induced AHR to ACh in guinea pigs, with an ED50 value of 0.52 mg/kg p.o. Since YM976 had no effect on the bronchoconstriction induced by ACh, even at 10 mg/kg p.o. (data not shown), the inhibition of AHR was not due to antagonism toward ACh. At the same time, YM976 also suppressed the eosinophil infiltration into the lungs. AHR is induced by the activation of infiltrated eosinophils into the airway (Venge, 1990). Infiltrated and activated eosinophils can induce epithelial damage by the release of cytotoxic cationic proteins, such as eosinophil peroxidase or eosinophil cationic protein. Thus, the prevention of antigen-induced eosinophilic inflammation by YM976 may underlie the reduction in the development of AHR. Another possible action site for PDE4 inhibitors is the sensory nerves. Rolipram inhibited the excitatory noncholinergic neurotransmission in guinea pig bronchi (Undem et al., 1994), and augmented the nonadrenergic and noncholinergic relaxation of human isolated bronchus (Fernandes et al., 1994). These effects may be related to the inhibition of the release of bronchoconstrictor neuropeptides such as Substance P from the sensory nerve endings. This may also contribute to the anti-inflammatory effects of PDE4 inhibitors since Substance P is a chemotactic factor for eosinophils (Kudlacz and Kippenberg, 1994).
As summarized, YM976 prevented antigen-induced immediate bronchoconstriction, airway edema, eosinophil infiltration, and AHR in guinea pigs. Furthermore, YM976 also showed in vitro and in vivo inhibition on the activation of eosinophils. Since YM976, unlike rolipram and RP73401, had a weak direct bronchodilator effect, these antiasthmatic effects have contributed to anti-inflammatory effects such as the suppression of inflammatory cell functions. YM976 may, through its anti-inflammatory activity, have a potential therapeutic value in the treatment of bronchial asthma.
Footnotes
-
Send reprint requests to: Dr. Motonori Aoki, International Clinical Development Department, Yamanouchi Pharmaceutical Co., Ltd., 17-1 Hasune 3-Chome, Itabashi-ku, Tokyo, 174-8612, Japan. E-mail: aokim{at}yamanouchi.co.jp
- Abbreviations:
- PDE4
- phosphodiesterase type 4
- DMSO
- dimethyl sulfoxide
- OA
- ovalbumin
- ACh
- acetylcholine
- fMLP
- formyl-methionyl-leucyl-phenylalanine
- LTD4
- leukotriene D4
- Rl
- lung resistance
- AHR
- airway hyperreactivity
- BAL
- bronchoalveolar lavage
- PIP
- intra-airway pressure
- IL-5
- interleukin-5
- RP73401
- 3-cyclopentyloxy-N-(3,5-dichloro-4-pyridyl)-4-methoxybenzamide
- Received October 19, 2000.
- Accepted December 13, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}