


Abstract
Peripheral analgesic effects of opioids are pronounced under inflammatory conditions, e.g., arthritis; however, little is known about adaptive changes of μ opioid receptor binding and G protein coupling in the peripheral versus central nervous system. The present study investigated the effects of inflammation on μ opioid receptor (MOP receptor) binding and G protein coupling of supraspinal, spinal, and peripheral MOP receptors. In addition, MOP receptors were identified in immunohistochemical experiments in dorsal root ganglia (DRG) of inflamed and noninflamed rats. The number of MOP receptor binding sites decreased from hypothalamus (HT) > spinal cord (SC) > DRG. Unilateral Freund's complete adjuvant inflammation of one hindpaw induced a significant up-regulation of MOP receptor sites only in DRG but not in HT or SC. This up-regulation was time-dependent, restricted to the inflamed side, and showed a peak at 24 h. The full-agonist [d-Ala2,N-MePhe4,Gly5-ol]-enkephalin (DAMGO) induced MOP receptor G protein coupling with decreasing efficacies (Emax) from HT > SC > DRG. Inflammation resulted in significant increases in MOP receptor G protein coupling only in membranes of DRG, but not in HT, SC, or DRG on the contralateral side of inflammation. This suggests that changes in MOP receptor levels are not related to systemically released mediators. These findings show that inflammation causes changes in MOP receptor binding and G protein coupling after DAMGO stimulation selectively in primary afferent neurons but did not cause any adaptive changes of MOP receptor in HT or SC.
Opioid receptors are G protein-coupled receptors that are activated by opioid peptides and alkaloids. Both classes of agonists mediate their analgesic effects in the central nervous system (CNS) and in the peripheral nervous system (Raynor et al., 1994; Binder et al., 2001). G proteins are heterotrimers of α, β, and γ subunits, and μ opioid receptors (MOP receptor) (International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification, 2003) produce biological responses by selectively activating G proteins of the pertussis toxin-sensitive Gi/Go family (Forse, 2000; Kenakin, 2002). After ligand binding at the receptor, G proteins can be activated by coupling. Interaction of G protein with an agonist-stimulated receptor leads to the replacement of GDP with GTP and dissociation of the α subunit from the remaining β,γ dimer (Neer, 1995). The α subunit and the βγ subunits influence various effectors, including inhibition of adenylyl cyclase, stimulation of potassium channel conductance, and inhibition of calcium channel conductance (Forse, 2000). All of these can result in diminished pain sensations (Woolf and Salter, 2000). The peripheral analgesic effects of opioids are elicited by activation of MOP receptor on primary afferent neurons (DRG). This is best described under local inflammatory conditions and has been shown in clinical (Likar et al., 1998) and experimental (Stein et al., 1989) studies. Intrathecal administration of opioids is widely used in clinical routine and shows significant MOP receptor-mediated analgesic effects at the first synaptic relay in the transmission of nociceptive messages (Julius and Basbaum, 2001). The current report investigates differences in MOP receptor binding and signaling at peripheral, spinal, or supraspinal neurons in an animal model of intraplantar Freund's complete adjuvant (FCA)-induced inflammation. This study investigates whether 1) opioids bind with the same affinity at MOP receptor in hypothalamus (HT), spinal cord (SC) or primary afferent neurons (DRG); 2) painful inflammation can cause a significant increase in MOP receptor binding sites in HT, SC, or DRG; 3) inflammation induces changes in G protein coupling in membranes of HT, SC, or DRG. These studies help to differentiate central versus peripheral alterations of MOP receptor binding and signaling after a painful inflammation.
Materials and Methods
Drugs. Guanosine-5′-O-(3-[35S]thio)-triphosphate ([35S]GTPγS) (1250 Ci/mmol) was purchased from PerkinElmer Life Sciences, Boston, MA). [3H]-[d-Ala2,N-MePhe4,Gly5-ol]enkephalin) (56 Ci/mmol) ([3H]DAMGO) was purchased from Amersham Biosciences UK, Ltd. (Little Chalfont, Buckinghamshire, UK). DAMGO and naloxone were purchased from Sigma (Taufkirchen, Germany). Scintillation fluid was obtained from PerkinElmer Life Sciences (Turku, Finland). Antibodies for immunohistochemistry were obtained from Vector Laboratories (Burlingame, CA). Synthetic peptide for MOP receptor was obtained from Gramsch Laboratories (Schwabhausen, Germany). Polystyrene-xylene was provided by Merck (Darmstadt, Germany). Tissue Tek compound (OCT) was provided by Miles (Bayer Corp., Elkhart, IN). Anesthesia was performed with halothane from Willy Rüsch GmbH (Böblingen, Germany). FCA was obtained from Calbiochem (San Diego, CA).
Subjects. Experiments were performed in male Wistar rats (180–200 g) individually housed in cages lined with sawdust, with free access to rat chow and water in a laboratory equipped with a 12/12-h light/dark cycle. Room temperature and relative humidity were maintained at 22 ± 0.5°C and 60%, respectively. The animal protocol was approved by the committee of the Landesamt für Arbeitsschutz, Gesundheit und Technische Sicherheit Berlin and the guidelines on ethical standards for investigations of experimental pain in animals were followed (Zimmermann, 1983).
Induction of Inflammation. For induction of inflammation, 0.15 ml of FCA was administered into the plantar surface of the right hind limb of rats under brief halothane anesthesia. A description of the time course and magnitude of the inflammatory reaction is given elsewhere (Stein et al., 1988). The inflammation remained confined to the inoculated paw throughout the observation period. The maximum observation period of FCA inflammation was 96 h.
Membrane Preparations. Rats were killed by halothane anesthesia after saline treatment (control animals) or treatment with FCA for 24 or 96 h, and HT, lumbal SC (L3–L5), and DRG (L3–L5) were removed. To obtain enough tissue for each DRG binding experiment, tissue from 10 euthanized rats was pooled for one binding curve ([3H]DAMGO or [35S]GTPγS). This procedure was performed in inflamed and noninflamed (control) animals. Contralateral DRG were examined to proof unilateral alterations and not used for control experiments. The tissue was placed immediately on ice in cold assay buffer (50 mM Tris-HCl, 1 mM EGTA, pH 7.4). Membrane preparations were made by pooling tissue from 10 rats. Tissue was homogenized with a Polytron homogenizer (Kinematica AG, Littau, Switzerland) and centrifuged at 42,000g at 4°C for 20 min. The pellet was resuspended in assay buffer followed by a 10-min incubation at 37°C to degrade endogenous ligands. The homogenate was centrifuged again at 42,000g and resuspended in assay buffer. Membranes were aliquoted and stored at –80°C for at least 30 min.
MOP Receptor Binding. Appropriate concentrations of cell membranes (50–100 μg) were prepared as described above and incubated in assay buffer with varying concentrations (0.02–3 nM) of the MOP receptor agonist [3H]DAMGO. In all opioid binding experiments, nonspecific binding, which typically represented 15 to 35% of total binding, was defined with naloxone (10 μM). Only specific binding is reported. Membranes were incubated in a final volume of 1 ml for 1 h at 30°C in assay buffer. Filters were soaked in 0.1% (w/v) polyethyleneimine solution for 30 min before using. Bound and free ligand were separated by rapid filtration under vacuum through Whatman GF/B glass fiber filters, followed by four washes with cold buffer (50 mM Tris-HCl, pH 7.4). Bound radioactivity was determined by liquid scintillation spectrophotometry at 70% counting efficiency for 3H after overnight extraction of the filters in 3 ml of scintillation fluid.
Measurement of DAMGO-Induced G Protein Binding at MOP Receptor. Membranes were thawed, homogenized, and centrifuged at 42,000g for 10 min. Membranes were incubated in [35S]GTPγS assay buffer containing 50 mM Tris-HCl, pH 7.4, 5 mM MgCl2, 0.2 mM EGTA, 100 mM NaCl, and 1 mM dithiothreitol. The buffer composition was similar to that of Newman-Tancredi et al. (1997). Concentration-effect curves were generated by incubating the appropriate concentration of membranes (30–50 μg) and varying concentrations of DAMGO (10–12–10–4 M), with 50 μM GDP and 0.05 nM [35S]GTPγS in a total volume of 800 μl. Basal binding was assessed in the absence of agonist, and nonspecific binding was measured in the presence of 10 μM unlabeled GTPγS. The reaction mixture was incubated for 2 h at 30°C. Bound and free [35S]GTPγS were separated by vacuum filtration through GF/B filters and quantified by liquid scintillation counting.
[35S]GTPγS Saturation Binding at MOP Receptor. In the presence (10 μM) or absence of DAMGO, saturation analysis of DAMGO-stimulated [35S]GTPγS binding to membranes was performed. Membranes were incubated with varying concentrations of [35S]GTPγS (0.05–2 nM) and 50 μM GDP in assay buffer for 2 h at 30°C. Unstimulated [35S]GTPγS binding was subtracted from agonist stimulated binding at each measurement point. The incubations for all experiments were terminated by filtration under vacuum through Whatman GF/B glass fiber filters, followed by four washes with cold buffer (50 mM Tris-HCl, pH 7.4). Bound radioactivity was determined by liquid scintillation spectrophotometry after extraction overnight in scintillation fluid.
Immunohistochemistry in DRG. Five rats per group (inflamed and noninflamed) were deeply anesthetized with halothane 4 days after saline or FCA treatment and transcardially perfused with 60 ml of warm saline, followed by 300 ml of 4% (w/v) paraformaldehyde with 0.2% (v/v) picric acid in 0.16 M phosphate buffer solution (pH 6.9). The ipsilateral L5 DRG were removed, postfixed in the same fixatives for 90 min, and then placed in 15% (w/v) sucrose solution at 4°C overnight. The tissue was embedded in Tissue Tek compound (OCT; Bayer Corp.), frozen, cut in 14-μm sections, and the sections were incubated overnight with anti-MOP receptor (1:1000) (kindly provided by Drs. S. Schulz and V. Höllt, Department of Pharmacology and Toxicology, Otto-von-Guericke University, Magdeburg, Germany). The appropriate biotinylated secondary antibody (Gramsch Laboratories-Biotechnology, Schwabhausen, Germany) was added (Mousa et al., 2002). Finally, the sections were washed and stained with 3′,3′-diaminobenzidine tetrahydrochloride containing 0.01% H2O2 in 0.05 M Tris-buffered saline (pH 7.6) for 3 to 5 min. After the enzyme reaction, the sections were washed in tap water, mounted onto gelatin-coated slides, dehydrated in alcohol, cleared in xylene, and mounted in polystyrene-xylene.
The method of quantification for DRG staining has been described previously (Mousa et al., 2001). Briefly, the total number of MOP receptor containing neurons was counted by an observer blinded to the experimental protocol. The total number of MOP receptor containing neurons divided by the total number of neurons in each DRG section was used to calculate the percentage of MOP receptor immunoreactive neurons. The cell body diameter was measured with the nucleus in the focal plane and was estimated from the average length and width determined with a calibrated micrometer. The diameters of 30 immunoreactive neurons with nucleus were measured in each animal.
Data Analysis. All ligand binding and [35S]GTPγS binding data are reported as mean ± standard error values of at least four experiments, which were each performed in duplicate. [3H]DAMGO ligand binding experiments and [35S]GTPγS saturation binding experiments were fitted to a one-site binding hyperbola using GraphPad Prism (GraphPad Software Inc., San Diego, CA) to determine Kd and Bmax values. Nonspecific binding was subtracted from all [3H]DAMGO and [35S]GTPγS binding data. EC50 values in [35S]GTPγS binding assays were determined by use of GraphPad Prism. Efficacy (Emax) is defined as the maximum percentage of stimulation by DAMGO, as determined by nonlinear regression analysis of concentration-effect curves. Statistical differences between animals with and without FCA inflammation were determined by the nonpaired Student's t test or analysis of variance (ANOVA) with posthoc comparison using Tukey test (Sigma Stat 2.03; SPSS Science, Chicago, IL).
Results
Binding Affinities and Opioid Receptor Numbers in Membranes of HT, SC, and DRG. Saturation binding of [3H]DAMGO displaced by naloxone showed similar binding affinities (Kd) in membrane preparations of HT, SC, and DRG (Table 1). The number of MOP receptor binding sites (Bmax) decreased from HT > SC > DRG. No differences in Kd and Bmax were detectable for HT and SC in animals with and without FCA inflammation (Table 1; Fig. 1, A and B). However, the number of opioid receptors (Bmax) increased significantly (p < 0.05; ANOVA) in DRG membranes at 24 and 96 h of FCA inflammation (Table 1; Fig. 1C). No change in Bmax was detectable in DRG membranes on the contralateral side of inflammation (29 ± 3.1 fmol/mg protein). Earlier data showed that opioid full agonists (e.g., fentanyl) produce dose-dependent elevations of paw pressure threshold in animals with FCA inflammation; however, much lower elevations of paw pressure threshold in animals without FCA inflammation (Antonijevic et al., 1995). Our biochemical data support better analgesic effects in inflammatory painful conditions.
Bmax and Kd values from saturation binding experiments of [3H]DAMGO in HT, SC, and DRG membranes of animals with (24 and 96 h FCA) and without (control) inflammation
Membranes were incubated with varying concentrations of [3H]DAMGO as described under Materials and Methods. Data are mean values of at least four independent experiments, obtained from nonlinear regression analysis of binding saturation curves.

Determination of MOP receptor was performed with [3H]DAMGO binding experiments in HT (A), SC (B), and DRG (C) membranes of animals with (24 and 96 h) and without FCA inflammation. Nonspecific binding was determined with 10 μM naloxone. Data shown are means of at least four representative experiments.
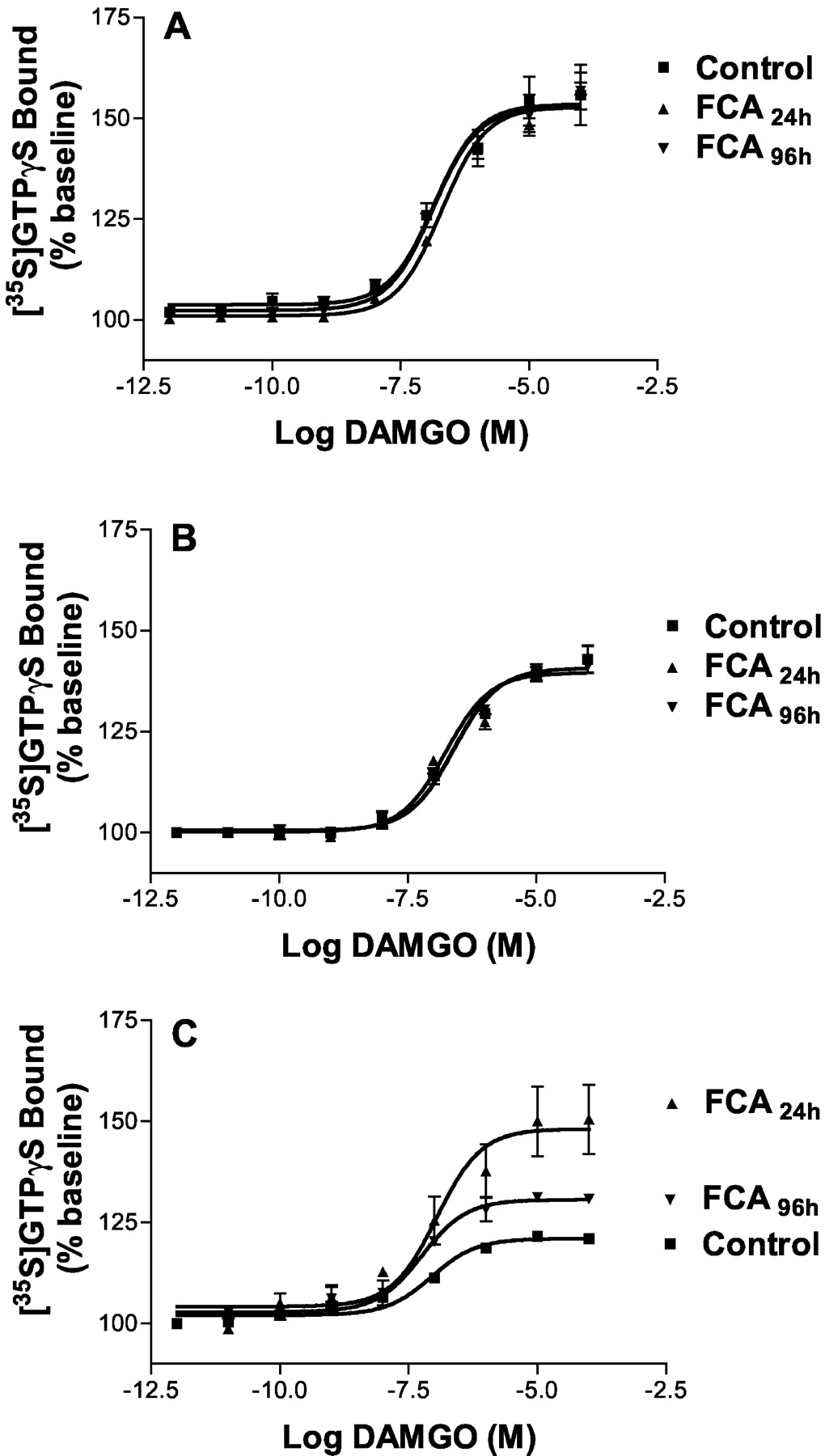
Potencies and Efficacies of DAMGO-Stimulated [35S]GTPγS Binding in Membranes of HT, SC, and DRG. [35S]GTPγS binding stimulated by increasing DAMGO concentrations revealed no significant differences in EC50 and Emax values at different time intervals of FCA inflammation for HT and SC and no significant differences in EC50 for DRG (Table 2; Fig. 2, A and B). However, after 24 and 96 h of FCA inflammation the efficacy (Emax) of DAMGO-induced G protein coupling in DRG membranes was significantly increased compared with controls (p < 0.05; ANOVA) (Table 2; Fig. 2C). A significant increase in the basal levels of DAMGO induced [35S]GTPγS binding in DRG of animals without FCA inflammation (3740 ± 173) compared with animals with 24 (6478 ± 415 cpm) and 96 (8250 ± 794 cpm) h of FCA inflammation was detectable (p < 0.05; ANOVA). However, basal [35S]GTPγS did not change in HT nor SC (data not shown).
Intrinsic efficacies (Emax) and potencies (EC50) of DAMGO-induced [35S]GTPγS binding in membranes of HT, SC, and DRG in animals without (control) and with (FCA 24 h and FCA 96 h) inflammation
Data are mean values ± S.E.M. of at least four independent experiments as described under Materials and Methods. Emax values are percentage of stimulation over basal (100%).

Stimulation of [35S]GTPγS binding to HT (A), SC (B), and DRG (C) membranes of animals without and with (24- and 96-h FCA) inflammation. Concentration-response curves were determined for DAMGO as described under Materials and Methods. Nonspecific binding was determined using 10 μM cold GTPγS and was subtracted from each data set. Basal [35S]GTPγS binding in the absence of added drugs was 4000 to 6000 cpm in all groups. Each value represents the mean ± S.E.M. of at least four independent experiments performed in duplicate.
[35S]GTPγS Saturation Binding Experiments. Saturation analysis of agonist-stimulated [35S]GTPγS binding was used to determine the apparent affinity of [35S]GTPγS for the activated G protein, and the number of G proteins activated. [35S]GTPγS saturation binding revealed high apparent affinities for G proteins at MOP receptor in HT, SC, and DRG after DAMGO (10 μM) stimulation (Table 3). No significant differences were detectable between apparent Kd Gprotein in membranes of HT, SC, and DRG in animals with and without FCA inflammation (Table 3; Fig. 3). Animals with FCA inflammation showed a significant increase in DAMGO stimulated apparent Bmax G protein in DRG membranes, but not in membranes of HT and SC (Table 3; Fig. 3). The relative amplification factor (amount of G protein bound/number of opioid receptors in membrane fractions) was calculated according to Selley et al. (1998). The number of G proteins activated by one MOP receptor in noninflamed tissue was three in HT, 13 in SC, and nine in DRG and was not significantly different between animals with and without FCA inflammation.
Characterization of G protein binding to MOP receptor at HT, SC, and DRG
Apparent affinity (Kd Gprotein) and apparent number (Bmax Gprotein) of G proteins for net agonist-stimulated [35S]GTPγS binding in DRG membranes of animals without (control) and with (FCA) inflammation after DAMGO stimulation. Membranes were incubated with varying concentrations of [35S]GTPγS as described under Materials and Methods. Data are mean apparent Bmax and apparent Kd values ± S.E.M., obtained from at least four independent experiments.

Saturation analysis of DAMGO-stimulated [35S]GTPγS binding to HT (A), SC (B), and DRG (C) membranes of animals with (24- and 96-h FCA) and without inflammation. Saturation binding of [35S]GTPγS was performed in the absence and presence of 10 μM DAMGO in HT (A), SC (B), and DRG (C) membranes.
Immunohistochemistry. Representative DRG sections of animals with and without FCA inflammation are shown in Fig. 4. Immunohistochemical visualization showed a significant increase of MOP immunoreactive neurons in DRG of rats with 96 h of FCA inflammation (25.8 ± 1.5%) compared with rats without FCA inflammation (16.8 ± 0.6%) (Fig. 4). This represents a 53% relative increase during inflammation. Cell characterization showed that mainly cells of small-to-medium diameter (20–50 μm) stained positive for MOP receptor.

Brightfield micrographs showing representative MOP receptor positive neurons in L5 DRG of rats without FCA inflammation (A) and in DRG of rats with FCA inflammation (B). MOP receptor positive neurons were significantly higher in inflamed (B) compared with noninflamed (A) sections. MOR-IR is mainly seen in small DRG neurons. Scale bar, 20 μm.
Discussion
In this report, we investigated the effect of a pathological, local FCA inflammation on MOP receptor binding and signaling in the central compared with the peripheral nervous system. The major finding of this study is that MOP receptor binding and G protein coupling is differentially altered in HT, SC, and DRG after FCA inflammation. Although MOP receptor binding remains unaffected in HT and SC, it is significantly increased in ipsilateral DRG after FCA hindpaw inflammation. The contralateral side is not affected by FCA inflammation. In addition, MOP receptor G protein coupling significantly increased in DRG neurons of animals with FCA inflammation. Consistently, our immunohistochemistry experiments show a rise in the number of small- but not large-diameter MOP receptor-positive DRG neurons. These results suggest that FCA inflammation of the hindpaw affects predominantly neurons in DRG associated with inflammation. This indicates that differences are not related to the systemic release of mediators of inflammation but are restricted to neurons that innervate painful inflamed tissue.
MOP receptor are localized and expressed in the central nervous system and in peripheral sensory neurons. The number of MOP receptor (Bmax) in DRG membranes in animals without inflammation is 22-fold lower than in HT and 4-fold lower than in SC, indicating that the density of MOP receptor decreases from HT > SC > DRG. The experiments were performed in rat hypothalamus because it was shown that this brain region is enriched with MOP receptors (Sim et al., 1995; Maher et al., 2000) and is involved in pain transmission (Basbaum and Fields, 1984). The affinity of DAMGO to MOP receptors is not different in HT, SC, and DRG, which supports previous studies showing that MOP receptors bind DAMGO in all isolated membrane preparations equally well (Maher et al., 2000).
FCA-induced inflammation of hindpaws does not alter MOP receptor binding sites in hypothalamus and spinal cord. In line with these findings, Spetea et al. (2002) reported no differences in binding affinities and Bmax from nonarthritic and arthritic rats in striatum, hypothalamus, hippocampus, frontal cortex, and lumbar spinal cord. Autoradiography experiments did not show differences in the binding of DAMGO, [d-Ala2,d-Leu5]-enkephalin, or bremazocine at μ, δ, or κ opioid receptors in lumbar spinal cord sections after FCA inflammation (Millan et al., 1988). However, in immunohistochemistry experiments by Goff et al. (1998) and Mousa et al. (2002), a significant increase (38 and 34%) of MOP receptor in lumbar sections of spinal cord after FCA-induced inflammation of the hindpaw was detectable. The discrepancy between immunohistochemistry and ligand binding might be explained by differences in the sensitivity of both techniques.
However, in animals with FCA inflammation, we found changes in the number of MOP receptor binding sites selectively on ipsilateral but not contralateral DRG membranes, reflecting adaptive changes in response to sustained inflammation. Bmax analysis showed a large increase of MOP receptor binding sites 24 h after inflammation in lumbar DRG sections. As shown previously, 96 h after inflammation Bmax was still significantly higher compared with control animals (Zöllner et al., 2003). However, the current study shows that the number of MOP receptor binding sites is lower compared with 24-h FCA inflammation. These results suggest that intraplantar application of FCA leads to a selective increase in MOP receptor expression in DRG neurons innervating painful inflamed tissue. This was confirmed in our immunohistochemical experiments with MOP receptor binding sites after FCA inflammation. An increase in MOP receptor density was detectable and predominantly localized in small diameter neurons. This indicates that MOP receptor immunoreactivity is mainly restricted to nociceptive neurons (Ji et al., 1995; Mousa et al., 2001) and is in agreement with a report by Silbert et al. (2003), who have shown that small nociceptors express MOP receptor mRNA at quantitatively higher levels. In a previous study using a ribonuclease protection assay, μ opioid receptor mRNA did not increase during FCA inflammation (Schäfer et al., 1995). However, changes in receptor protein were not examined. Current investigations using quantitative real-time polymerase chain reaction technology show that mRNA for μ opioid receptors might increase at early time points of FCA inflammation. This supports the hypothesis that an increase in the efficacy of opioid analgesia during inflammation might be related in part to an up-regulation of MOP receptor expression in small size nociceptors. For human neuroblastoma cells it was shown recently that an increase in MOP receptor mRNA might be related to mediators of inflammation (e.g., interleukin-4, tumor necrosis factor) (Kraus et al., 2001). In addition, nerve growth factor is also thought to be critical for altered gene transcription and protein synthesis in DRG neurons (Lindsay and Harmar, 1989; Alvares and Fitzgerald, 1999).
The functional status of MOP receptor can be measured in concentration-effect curves for agonist-stimulated [35S]GTPγS binding (Audinot et al., 2002). We performed a set of experiments to determine this initial stage of G protein activation to reveal possible changes in MOP receptor-mediated G protein-coupling of DRG in animals with and without FCA inflammation. As predicted from classical receptor theory (Selley et al., 1998), the relationship between receptor occupancy and G protein activation by MOP receptor depends on receptor density. Consistently, in our studies the absolute magnitude of the G protein stimulation (efficacy Emax) was higher in isolated membranes with higher receptor density (Emax: HT > SC > DRG). In HT and SC, no significant differences in efficacy were measurable after induction of inflammation. This supports our MOP receptor binding data showing that a local inflammation does not affect binding and signaling of opioids in the CNS. An important finding of the present study is that the efficacy of DAMGO-stimulated G protein activation increased significantly, time dependently, and selectively in DRG membrane preparations of animals with FCA inflammation but not in membrane preparations of the CNS. Maximal stimulation of [35S]GTPγS binding by DAMGO in DRG increased 24 and 96 h after inflammation only on the ipsilateral side. An increase in G protein coupling during inflammation might explain why in a number of studies the application of exogenous opioids in behavioral experiments of peripheral antinociception are enhanced under inflammatory conditions (Schäfer et al., 1995; Stein et al., 2001). The basal levels of [35S]GTPγS binding are higher in DRG membranes of animals with 24 and 96 h of inflammation. This increase is not detectable in HT nor SC. It might indicate constitutive activity in DRG membranes of inflamed animals, which could be related to permanent stimulation through mediators of inflammation (e.g., prostaglandins).
Some studies have found that GTPγS binding to membrane preparations is reversible and could reach equilibrium under certain conditions (Hilf et al., 1992; Breivogel et al., 1998). The saturation analysis of agonist-stimulated [35S]GTPγS binding in our experiments was performed in the presence of GDP in a competitive manner. Therefore, the affinity of [35S]GTPγS for the activated G protein and the number of G proteins activated are only apparent values. The apparent Bmax Gprotein (app. Bmax) and the apparent Kd Gprotein (app. Kd Gprotein) do not represent the exact maximal number of G proteins or affinity of G proteins to the receptor. However, relative comparisons between tissues are possible as described previously in detail (Newman-Tancredi et al., 1999; Selley et al., 2001). We found that DAMGO occupied MOP receptor receptors bind G proteins with high affinity in HT, SC, and DRG in animals with and without FCA inflammation. However, significant differences were detectable in the number of G proteins activated. The system with the highest receptor density (HT) showed the highest number of G proteins activated. In comparison, a saturated concentration of DAMGO in SC or DRG sections could activate only 80 or 16%, respectively, compared with G protein activation in HT. A significant increase in the number of activated G proteins was detected in DRG membranes after 24 or 96 h of FCA-induced inflammation. This increase could underlie the enhanced antinociception produced by treatment with opioids in inflammatory conditions as shown in previous behavioral experiments (Stein et al., 1989).
The relative amplification factor represents the extent of G protein amplification between treatment (FCA) and control tissue. This factor depends on the relative expression levels of all forms of G protein and might vary in tissue after FCA treatment; however, we have not investigated changes in G protein content between control and FCA-treated animals. The number of G proteins activated by one MOP receptor was three in the HT, 13 in the SC, and nine in the DRG. Relative amplification factors for DRG, HT, and SC have not significantly changed between inflamed and noninflamed animals. The amplification factor for HT is different from a previous publication by Maher et al. (2000), in which the amplification factor for HT was reported with 19. However, the specific assay conditions were different (e.g., GDP concentration), which might explain differences in the relative amplification factors.
In conclusion, painful inflammation leads to differential alterations of MOP receptor expression in brain, spinal cord, and DRG. It is associated with an up-regulation of MOP receptor mainly in small sized primary afferent neurons, but not in the CNS. This suggests that a locally applied inflammation can affect neurons that are innervating the region of inflammation. This has functional relevance because the efficacy of MOP receptor agonists in G protein coupling is enhanced in primary afferent fibers. Our findings indicate that clinical treatment of patients with opioids in inflammatory diseases is different from patients with noninflammatory painful conditions. The known increased analgesic efficacy of locally applied opioids could be explained by a selectively increased numbers as well as G protein coupling efficacy of MOP receptor in primary afferent neurons.
Acknowledgments
We thank Drs. Stefan Schulz and Volker Höllt for the gifts of the MOP receptor antibody.
Footnotes
-
This work was supported by “Klinische Forschergruppe Grant from the Deutsche Forschungsgemeinschaft (DFG) KFO 100/1, International Anesthesia Research Society, and Graduiertenkolleg 276/2: Signalerkennung und-umsetzung”.
-
DOI: 10.1124/jpet.103.057257.
-
ABBREVIATIONS: CNS, central nervous system; MOP, μ opioid receptor; DRG, dorsal root ganglion; FCA, Freund's complete adjuvant; HT, hypothalamus; SC, spinal cord, spinal cord; GTPγS, guanosine-5′-O-(γ-thio)-triphosphate; DAMGO, [d-Ala2,N-MePhe4,Gly5-ol]enkephalin; ANOVA, analysis of variance.
-
↵1 These authors contributed equally to this work.
- Received July 22, 2003.
- Accepted October 14, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}