


Abstract
Benztropine (BZT) analogs inhibit dopamine uptake but are less effective than cocaine in producing behavioral effects predicting abuse liability. The present study compared reinforcing effects of intravenous BZT analogs with those of standard monoamine uptake inhibitors and the effects of their oral pretreatment on cocaine self-administration. Responding of rats was maintained by cocaine [0.032–1.0 mg/kg/injection (inj)] or food reinforcement under fixed-ratio five-response schedules. Maximal rates of responding were maintained by 0.32 mg/kg/inj cocaine or substituted methylphenidate, with lower rates maintained at lower and higher doses. The N-methyl BZT analog, AHN 1-055 (3α-[bis(4′-fluorophenyl)methoxy]-tropane), also maintained responding (0.1 mg/kg/inj), although maximal rates were less than those with cocaine. Responding was not maintained above vehicle levels by the N-allyl, AHN 2-005 (N-allyl-3α-[bis(4′-fluorophenyl)methoxy]-tropane), and N-butyl, JHW 007 [N-(n-butyl)-3α-[bis(4′-fluorophenyl)methoxy]-tropane], BZT analogs, and it was not maintained with nisoxetine or citalopram. Presession treatment with methylphenidate (3.2–32 mg/kg) dose-dependently shifted the cocaine self-administration dose-effect curve leftward, whereas nisoxetine and citalopram effects were not significant. An intermediate dose of AHN 1-055 (32 mg/kg) increased responding maintained by low cocaine doses and decreased responding maintained by higher doses. A higher dose of AHN 1-055 completely suppressed cocaine-maintained responding. Both AHN 2-005 and JHW 007 dose-dependently (10–32 mg/kg) decreased cocaine self-administration, shifting its dose-effect curve down. Decreases in cocaine-maintained responding occurred at doses of methylphenidate and BZT analogs that left food-maintained responding unchanged. During a component in which injections were not available, methylphenidate and AHN 1-055, but not AHN 2-005 or JHW 007, increased response rates. These findings further support the low abuse liability of BZT analogs and their potential development as medications for cocaine abuse.
Drug abuse is a problem worldwide, with concomitant medical, social, and economic burdens at the individual and societal level. Drug self-administration procedures using laboratory animals have been used to assess the liability for abuse of drugs, to understand the behavioral and neurobiological mechanisms underlying drug reinforcement, and to discover potential medical treatments for drug abusers. Such treatments have been difficult to obtain with cocaine and other psychomotor stimulants (Vocci and Ling, 2005).
Cocaine inhibits the uptake of monoamine neurotransmitters (Taylor and Ho, 1978), although most studies support actions at the DAT as the primary biological mechanism underlying its abuse-related effects. For example, DAT affinities of various monoamine uptake inhibitors are correlated with their potencies in drug self-administration procedures, whereas there is no such correlation for the SERT or NET (Ritz et al., 1987; Bergman et al., 1989). These and other results suggesting the DAT as the primary target for cocaine's abuse liability also suggest it as a target for medications to treat cocaine abuse (Carroll et al., 2006).
Preclinical features considered desirable in potential treatments for cocaine abuse include effectiveness in modifying cocaine self-administration and a selectivity in doing so (e.g., Mello and Negus, 1996). In addition, reduced cocaine-like effects, in particular reduced reinforcing effects, should limit the treatment's own potential for abuse. Although many of the DAT inhibitors have behavioral effects resembling those of cocaine (Bergman et al., 1989; Spealman et al., 1989), several studies have pointed to what may be important differences among some DAT inhibitors. For example, various dopamine uptake inhibitors stimulated locomotor activity to different extents at comparable DAT binding potencies (Vaugeois et al., 1993). Moreover, Carroll et al. (2004) reported on an extensive series of cocaine analogs finding substantial differences in their maximal stimulation of locomotor activity; several were identified that did not stimulate locomotor activity and one that did not fully substitute for cocaine in rats trained to discriminate cocaine from saline. The authors suggested that the lack of full cocaine-like behavioral effects of these DAT inhibitors was due to their other actions. However, previous studies reported “atypical” DAT inhibitors, including analogs of BZT, that have pharmacological profiles unlike that of cocaine (e.g., Newman et al., 1995; Katz et al., 1999), that more recently were suggested to be due to different molecular interactions at the DAT (e.g., Beuming et al., 2008; Loland et al., 2008).
Among BZT analogs previously studied are several N-substituted analogs with high affinity for the DAT (11.8–29.9 nM), greater than 50-fold selectivity for the DAT over SERT and NET, and varying degrees of cocaine-like effects (Agoston et al., 1997; Katz et al., 2004). In addition, these compounds vary with regard to their selectivity for the DAT over muscarinic M1 receptors. A para-F-substituted BZT analog, AHN 1-055, has comparable DAT and M1 affinity and produces locomotor stimulant effects in mice, although to a lesser extent than cocaine. AHN 1-055 also substitutes for cocaine in rats trained to discriminate cocaine from saline after a 90-min delay (Katz et al., 1999). The N-allyl (AHN 2-005) and N-butyl (JHW 007) analogs of AHN 1-055 were 6- and 16-fold selective for the DAT over muscarinic M1 receptors, produced less stimulation of locomotor activity, and did not fully substitute in rats trained to discriminate cocaine at any time points examined (Katz et al., 2004). Furthermore, none of the three compounds produced robust place conditioning in rats compared with that of cocaine, even when accommodation was made for a delayed onset of action (Li et al., 2005). Finally, JHW 007 antagonized the locomotor stimulant effects of cocaine in mice (Desai et al., 2005a), whereas other DAT inhibitors typically shift the cocaine dose-effect curve leftward.
Because the effects of the above-mentioned N-substituted BZT-analogs represent varying degrees of similarity to the effects of cocaine, their self-administration was compared with several standard monoamine uptake inhibitors as an indication of their liability for abuse. In addition, the potential of these drugs for development as treatments for cocaine abuse was assessed by examining their effects on the subsequent self-administration of cocaine. Those effects were compared with effects on responding maintained by food reinforcement to assess the specificity of their actions for cocaine self-administration. We report here that the N-substituted BZT analogs either were not self-administered or were self-administered less than cocaine. In addition, the drugs decreased cocaine self-administration without affecting food-maintained behavior. These preclinical findings suggest the possibility for development of N-substituted BZT analogs as medical treatments for cocaine abuse, which themselves have low abuse liability.
Materials and Methods
Subjects. Male Sprague-Dawley rats were obtained from Charles River Laboratories, Inc. (Wilmington, MA) and acclimated to the laboratory for a minimum of 1 week. During this time, food (Scored Bacon Lover Treats; Bio-Serv, Frenchtown, NJ) and tap water were freely available. After the acclimation period, weights of rats were maintained at approximately 320 g by adjusting their daily food ration. The humidity- and temperature-controlled colony room was maintained on a 12-/12-h light/dark cycle, with lights on at 7:00 AM. The animals were cared for in accordance with the Guide for Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, 1996), the National Institute on Drug Abuse Intramural Research Program Animal Program, which is fully accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International.
Apparatus. Drug self-administration studies were conducted in experimental chambers (modified ENV-203; MED Associates, St. Albans, VT) that measured 25.5 × 32.05 × 26.5 cm and were placed within sound-attenuating cubicles equipped with a fan for ventilation. On the front wall of each chamber were two response levers, 4.0 cm above the grid floor and 5.0 cm from the midline. Downward displacement of the levers by a 20-g weight produced an audible click of a relay mounted behind the front wall of the chamber and were counted as responses. Three light-emitting diodes (LEDs) were located in a row above each lever. A receptacle for the delivery of food pellets was mounted on the midline of the front wall between the two levers and 2.0 cm above the floor. A syringe infusion pump (model 22; Harvard Apparatus Inc., Holliston, MA) mounted above each chamber delivered injections of specified volumes and durations from a 10-ml syringe. The syringe was connected by Tygon tubing to a single-channel fluid swivel (375 Series Single Channel Swivels; Instech Laboratories, Inc., Plymouth Meeting, PA), which was mounted on a balance arm above the chamber. Tygon tubing from the swivel to the subject's catheter was protected by a surrounding spring and completed the connection to the subject.
Procedures. Subjects were initially trained to lever-press the right lever with food reinforcement (45-mg food pellets; Bio-Serv) under a fixed-ratio (FR) five-response schedule of reinforcement. Each fifth response produced a food pellet (FR five-response schedule), which was followed by a 20-s time-out period during which responses had no scheduled consequences (other than feedback clicks). Sessions lasted for 20 min or until 30 food pellets were delivered. A total of 18 subjects were used in the study, with a minimum of six at each tested dose.
When subjects obtained 30 food pellets within each of three consecutive sessions, they were assigned to one of two groups. One group continued with food reinforcement, whereas subjects in the other group were then implanted in the right external jugular vein with a chronic indwelling catheter that exited at the midscapular region of the animal's back. Implantation of catheters was performed under anesthesia mixture of ketamine (60.0 mg/kg i.p.) and xylazine (12.0 mg/kg i.p.). Catheters were infused daily with 0.1 ml of a sterile saline solution containing heparin (30.0 IU/ml) and penicillin G potassium (250,000 IU/ml) to minimize the formation of clots, fibroids, and infection. All animals were allowed to recover from surgery for approximately 7 days before they were given access to cocaine.
Cocaine self-administration training was conducted in 2-h daily sessions until criteria for stable cocaine self-administration behavior were met. During these sessions, illumination of the LEDs above the right lever signaled the availability of injections. Completion of an FR 5 turned off the LEDs and activated the infusion pump for 0.2 s. A 20-s time-out period, during which LEDs were off and responding had no scheduled consequences, started with the injection, after which the LEDs were again illuminated, and responding had scheduled consequences. Responses on the left lever at all times and on the right lever during time-out periods were recorded but had no scheduled consequences throughout the study; these data are not presented. The criteria for acquisition of cocaine self-administration consisted of three consecutive sessions with at least 30 reinforced responses during the 2-h session.
To assess a full range of cocaine doses in a single session, the final phase of training consisted of separating the session into five sequential 20-min components, each preceded by a 2-min time-out period. The components were identical with the exception that infusion volumes and durations were adjusted to deliver cocaine injections in an ascending order in the five sequential components as follows: no injection (also referred to as extinction because responses had no scheduled consequences), 0.03, 0.10, 0.32, and 1.0 mg/kg/inj. Infusion volumes and durations were, respectively, 0, 5.6, 18.0, 56.0, and 180 μl and 0, 0.32, 1.0, 3.2, and 10.0 s, based on a body weight of 0.32 kg. A sample injection of cocaine at the corresponding dose occurred independently of responding at the end of the time-out period that preceded each component. Training continued until: 1) a minimum of 5.0 mg/kg cocaine was self-administered within a session with less than 20% variation in the total number of cocaine injections compared with the previous session, 2) the dose of cocaine that maintained maximal response rates varied by no more than one-half log unit over two consecutive test sessions, and 3) maximal response rates were at least 5-fold higher than response rates maintained during extinction. The effects of substitution of other drugs for cocaine (methylphenidate, citalopram, nisoxetine, AHN 1-055, AHN 2-005, JHW 007) or their presession treatments on cocaine self-administration were separated by a minimum of 48 h and were conducted only if during the most recent session, the dose of cocaine that maintained peak response rates was obtained in the third, fourth, or fifth component. All of the tests were conducted with a mixed order of drugs and doses.
Subjects studied under the FR 5 schedule of food reinforcement were trained until they obtained 30 food pellets within three consecutive sessions. Because the response rates maintained were higher than those maintained by cocaine and because the conditions of the experiment including the control response rates can influence the effects of drug treatments, the schedule of food reinforcement was modified such that it resembled that used in studies of cocaine self-administration. Under this modified procedure, the experimental session was separated into five sequential 20-min components, each preceded by a 2-min time-out period. During the first of the five components, responses had no scheduled consequences, whereas in the subsequent four components, each fifth response produced a food pellet. To approximate the response rates maintained by cocaine, subjects were given their daily ration of food (Harlan Rodent Chow, 15 g; Harlan, Indianapolis, IN) 60 min before sessions. Effects of prior treatment with AHN 1-055, AHN 2-005, JHW 007, and methylphenidate on the response rates in the fourth of the components (corresponding to the one in which cocaine was self-administered at its maximal rate) were assessed as described above.
Drugs. The drugs used in the present study were as follows: (-)-cocaine hydrochloride (Sigma-Aldrich, St. Louis, MO), methylphenidate (National Institute of Drug Abuse), nisoxetine (Sigma-Aldrich), and citalopram (Sigma-Aldrich). Self-administration of the test drugs was assessed with intravenous delivery of injections. Presession treatments of the test drugs were delivered orally by gavage in a volume no greater than 3.5 ml using feeding needles and gentle hand restraint of subjects. To habituate subjects to the gavage procedure, saline was administered before daily sessions until subjects tolerated the procedure with minimal struggling. Methylphenidate, nisoxetine, and citalopram were administered at 60 min before sessions, whereas AHN 1-055, AHN 2-005, and JHW 007 were administered at 180, 240, and 300 min before sessions, respectively. All N-substituted BZT analogs were synthesized in the Medicinal Chemistry Section (Intramural Research Program, National Institute on Drug Abuse, Baltimore, MD) according to published procedures (AHN 1-055, Newman et al., 1995; AHN 2-005 and JHW 007, Agoston et al., 1997). All drug solutions were prepared fresh daily in 0.9% NaCl and administered intravenously (self-administration) or orally (presession treatments).
Data Analysis. Response rates were determined by dividing responses by elapsed time in each component, excluding time-out periods. Total drug intake in each component was determined as milligrams per kilogram by multiplying the number of injections by the dose per injection. The significance of effects on response rate or total intake was assessed by ANOVA, with Bonferroni t test for post hoc analyses as appropriate. To determine whether there was a difference in effects of cocaine compared with saline self-administration, a two-way, repeated-measures ANOVA was used. A one-way, repeated-measures ANOVA was used to assess the effects of successive components in the substitution for cocaine of the test drugs. A two-way ANOVA was used to assess the effects of presession treatments of the test drugs on cocaine self-administration and for the comparison of effects of test drugs on responding maintained by cocaine or food reinforcement. For the latter comparison, response rates maintained by 0.32 mg/kg/inj cocaine (peak responses under control conditions) or food reinforcement in the fourth of the five components were directly compared. For all analyses, the criterion for significance was set at p < 0.05.
Results
Performances maintained by cocaine were similar to performances under FR schedules maintained by conventional reinforcers and cocaine injections; a brief pause was followed by a sequence of five responses made in rapid sequence producing the injection (Fig. 1, top). There were few, if any, responses on the inactive lever (vertical marks on the line below cumulative curve) or during the 2-min time-out periods between successive increments in dose (lower event line displaced upward). In the extinction component, no injections were delivered, and response rates were low and emitted in a pattern resembling the FR pattern. As dose of cocaine increased, response rates increased, and pauses decreased. The highest rate of responding was obtained in the fourth component, in which injections of 0.32 mg/kg/inj were available. When saline injections were available in the second through fifth components (Fig. 1, second panel, curves above 0.0), responses were never emitted at rates greater than those maintained in extinction (first component).
The average response rates were an inverted U-shaped function of cocaine dose, with a maximum of 0.33 ± 0.15 responses/s at 0.32 mg/kg/inj, which was significantly greater than the rates of approximately 0.03 responses/s occurring in extinction or with vehicle injections (Fig. 2A, top). Two-way repeated measures ANOVA indicated a significant effect of component (dose or no injection) on response rate (F4,20 = 4.06; p = 0.014) but no effect of treatment (cocaine or saline) condition (F1,20 = 3.62; p = 0.115). However, there was an interaction of treatment and component (F4,20 = 4.321; p = 0.011), indicative of the reinforcing effect of cocaine. Post hoc comparisons indicated that 0.32 mg/kg/inj maintained response rates greater than those obtained in extinction (t = 4.681; p < 0.001) and greater than those obtained when saline injections were available (t = 3.989, p < 0.001). With saline injections (Fig. 2A, open symbols), response rates were low (less than 0.03 responses/s), and post hoc comparisons indicated no significant effect of successive components.

Representative cumulative records of performances maintained by intravenous cocaine under the fixed-ratio five-response schedule and those obtained when different drugs or saline were available for self-administration. Ordinates, cumulative responses. Abscissae, time. Each experimental session started with a 2-min time-out period during which all lights were off, and responses had no schedule consequences (lower event line up). After the time out (lower event line down), lights above the levers were illuminated, although responses had no scheduled consequences (extinction) for 20 min, followed by another 2-min time out. When the lights were again illuminated, each fifth response turned off the LEDs and activated the infusion pump for 0.2 s (diagonal marks on the cumulative record). Vertical marks on the line below the cumulative curve indicate responses on the left (inactive) lever. A 20-s time-out period during which lights were off and responding had no scheduled consequences followed each injection, after which the LEDs were again illuminated, and responding had scheduled consequences. During each 20-min period of drug availability, the injection volumes were adjusted to deliver doses, as indicated on the figure, in an ascending order. The cumulative curve reset to the baseline at the end of the 20-min component.
Total saline intake (in milliliters) increased through successive components as injection volume increased (Fig. 2A, bottom, open symbols). A two-way ANOVA indicated a significant effect of cocaine versus saline (F1,20 = 33.7; p = 0.002), component (F4,20 = 46.4; p = 0.001), and their interaction (F4,20 = 45.6; p = 0.001).
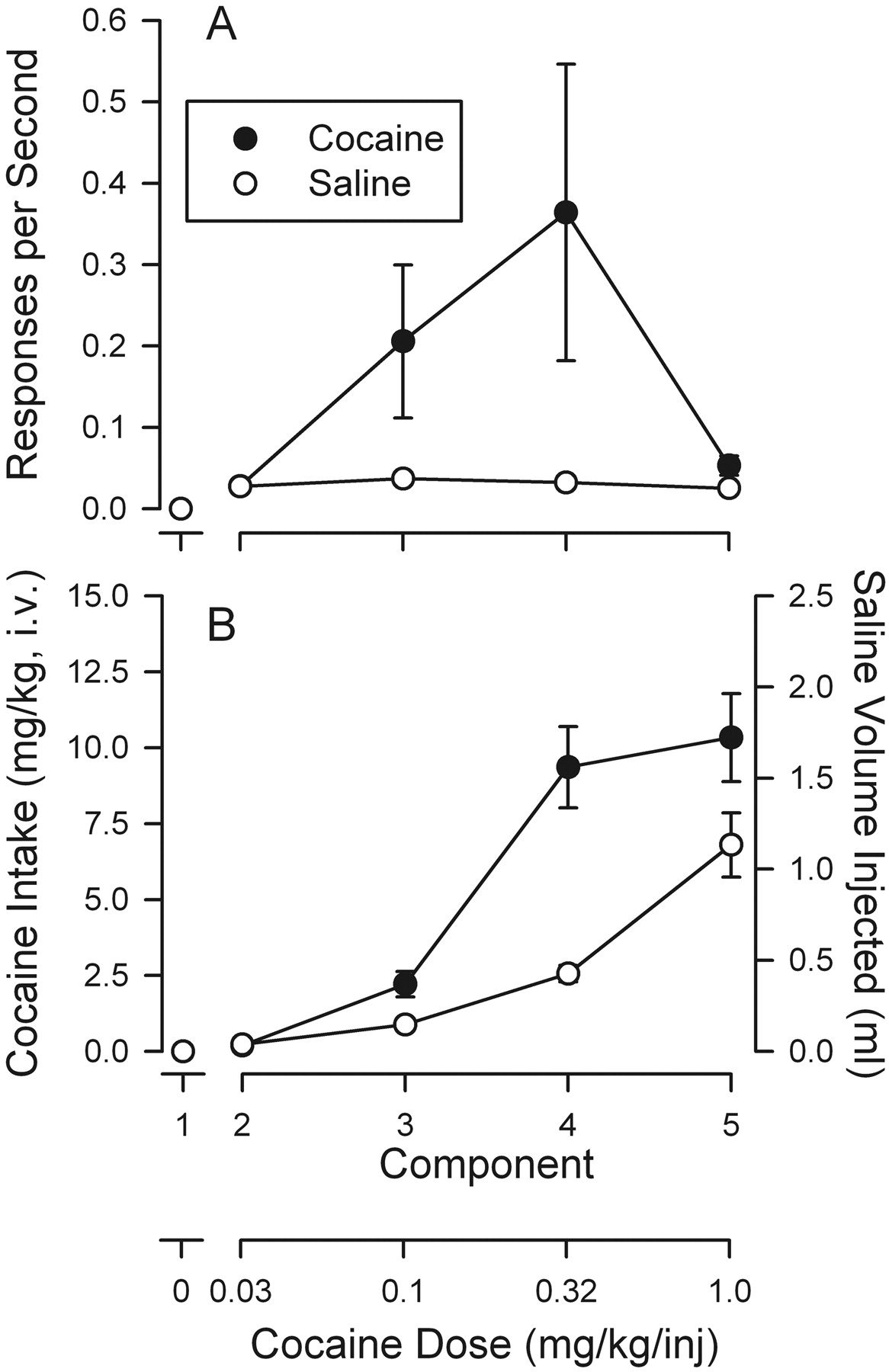
Substitution of cocaine or saline in rats trained to self-administer cocaine. Top ordinates, responses per second. Bottom ordinates, total intake, in milligrams per kilogram (drug) or milliliters (saline). Abscissae, injection dose in milligrams per kilogram. Each point represents the mean ± S.E.M. (n = 6–11).
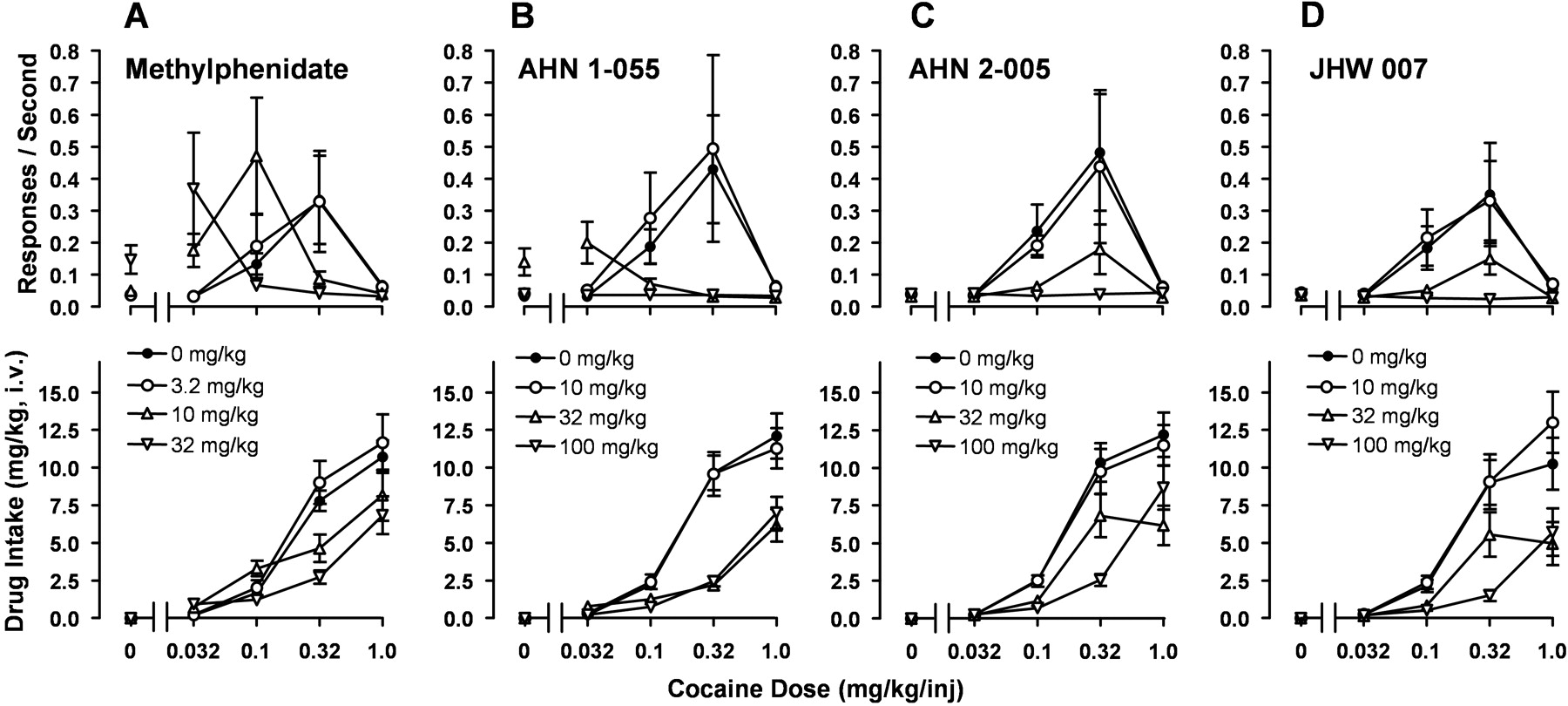
Performances maintained by methylphenidate were similar to those maintained by cocaine (Fig. 1, third panel). Methylphenidate maintained maximal response rates comparable with those maintained by cocaine and was similar in potency (Fig. 3A). The highest rate of responding was maintained by 0.32 mg/kg/inj, and lower rates were maintained at higher and lower doses. Response rates (F4,20 = 4.774; p = 0.007) and total intake (F4,20 = 52.98; p < 0.001) were significantly affected by dose, and post hoc tests indicated that rates maintained by 0.32 mg/kg/inj were significantly greater than those in extinction (t = 3.618; p = 0.017) and those maintained by 0.032 (t = 3.626; p = 0.17) and 1.0 (t = 3.415; p = 0.027) mg/kg/inj.
Significant effects of component with citalopram and nisoxetine were also obtained (F4,20 = 7.401; p =0.001; F4,20 = 3.357; p = 0.029, respectively). However, post hoc tests indicated no significant differences between any dose/injection compared with the no-injection component, indicating that neither served as a reinforcer.
AHN 1-055, at a dose of 0.1 mg/kg/inj, maintained rates of responding that were significantly higher than the rate maintained when no injections were available (Fig. 3C, top, open circles). Effects of AHN 1-055 dose on response rate were significant (F4,20 = 10.153, p < 0.001), with post hoc tests indicating response rates at 0.1 mg/kg/inj significantly greater than those obtained without injections (t = 4.991; p < 0.001) and those obtained at 0.032 (t = 5.049; p < 0.001) and 1.0 mg/kg/inj (t = 4.936; p < 0.001). A representative performance shows that at 0.1 mg/kg dose/inj, response rates were approximately twice those obtained at the lower dose or during extinction (Fig. 1, fourth panel, compare third with first or second components); however, responding ceased after approximately 20 injections. Increasing the dose 3-fold (0.32 mg/kg/inj) decreased response rates.

Substitution of monoamine uptake inhibitors and BZT analogs in rats trained to self-administer cocaine. A, cocaine (filled circles) and methylphenidate (open circles). B, cocaine (filled circles) and citalopram (open circles) or nisoxetine (triangles). C, cocaine (filled circles) and AHN 1-055 (open circles), AHN 2-005 (triangles up), or JHW 007 (triangles down). All other details are as in Fig. 2.
AHN 2-005 did not maintain response rates significantly greater than those occurring in extinction (Fig. 3C, upward triangles). Effects of AHN 2-005 dose on response rate were significant (F4,20 = 3.493, p = 0.026); however, post hoc tests indicated that the significance was due to the lower response rates at 1.0 mg/kg/inj compared with those obtained without injections (t = 3.411; p = 0.028). A representative performance with this compound (Fig. 1, panel 5) was characterized by relatively long pauses and cessation of responding well before drug availability ended.
For JHW 007, there was a significant effect of component (F4,20 = 4.237, p = 0.012), with post hoc tests indicating that response rates at 0.1 mg/kg/inj were greater than those obtained without injections (t = 3.436; p = 0.026). However, as shown in Figs. 1 (panel 6) and 3 (C, downward triangles), response rates at that dose were relatively low (0.07 responses/s at 0.1 mg/kg/inj) and comparable with those obtained with no injections. As with AHN 2-005, a representative performance (Fig. 1; panel 6) shows relatively long pauses and a cessation of responding before drug availability ended.
Effects of presession treatment with methylphenidate and N-substituted BZT analogs on cocaine self-administration. Each point represents the mean ± S.E.M. (n = 6–10). Methylphenidate, AHN 1-055, AHN 2-005, or JHW 007 were administered orally at 60, 180, 240, or 300 min before sessions, respectively. All other details are as in Fig. 2.
Presession treatments with methylphenidate shifted the cocaine self-administration dose-effect curve leftward, without affecting maximum response rate (Fig. 4A, top). Doses of 10 and 32 mg/kg methylphenidate shifted the dose-effect curve approximately 3- and 10-fold to the left, respectively (Fig. 4A). Representative performances show that with methylphenidate pretreatment, there was often a cessation in responding toward the latter part of the component (Fig. 5, compare first and second panels). In addition, at the highest dose, methylphenidate pretreatment increased response rates during extinction (Fig. 4A, top, triangle pointed down). A two-way ANOVA indicated a significant effect of cocaine dose (F4,120 = 3.575; p = 0.009), a nonsignificant effect of presession treatment (F3,120 = 0.309; p = 0.819), and a significant interaction of the two (F12,120 = 3.232; p < 0.001). The higher two doses of methylphenidate decreased cocaine intake (Fig. 4A, bottom).
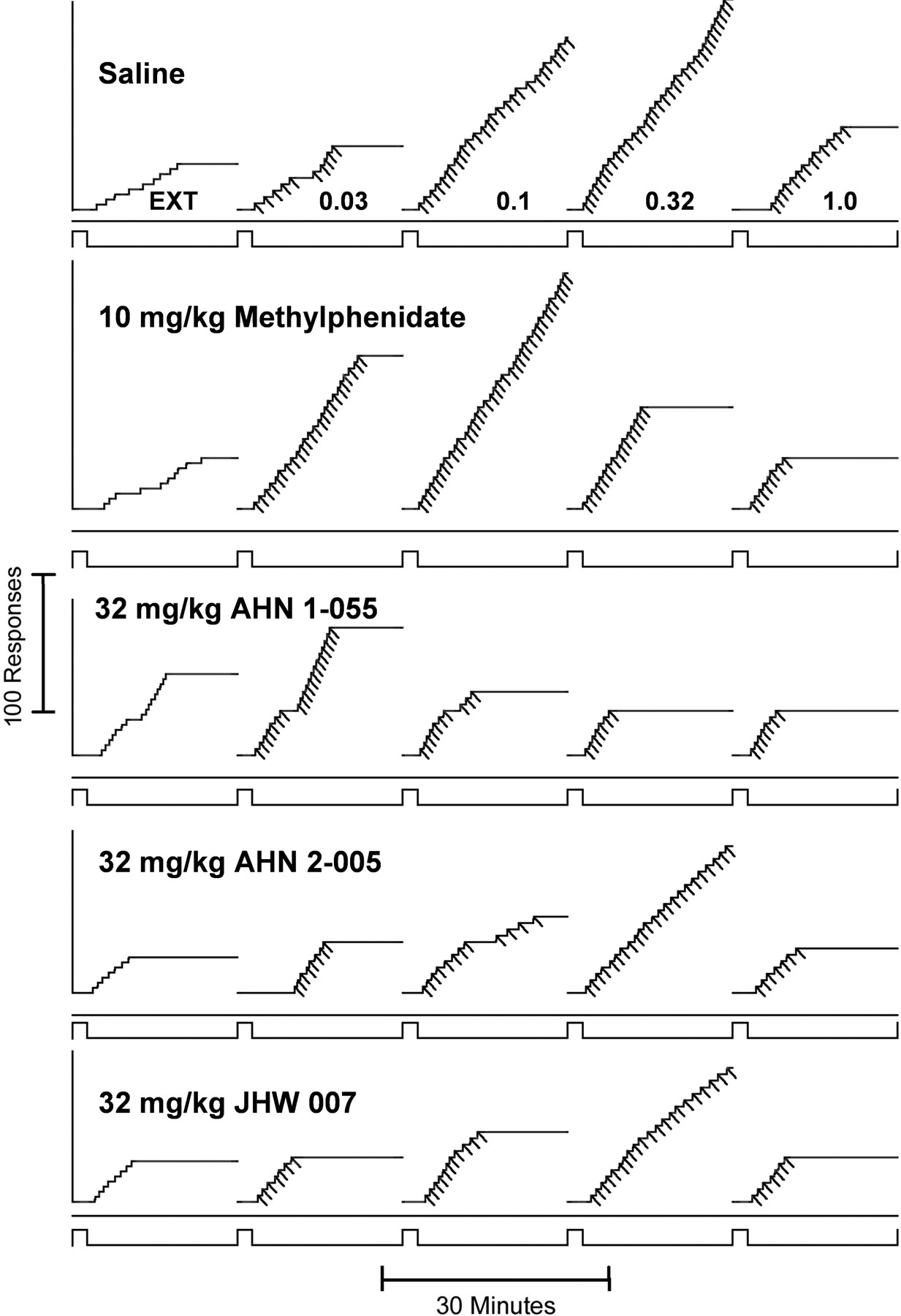
Representative cumulative records showing effects of saline or selected doses of methylphenidate, AHN 1-055, AHN 2-005, and JHW 007 presession treatments on performances maintained by intravenous cocaine under the fixed-ratio five-response schedule. Each panel shows a different presession treatment drug; all other aspects are as described for Fig. 1.
In contrast to the effects of methylphenidate, neither the norepinephrine uptake inhibitor, nisoxetine, nor the serotonin uptake inhibitor, citalopram, substantially altered cocaine self-administration. Two-way ANOVA of the effects of citalopram on response rates indicated a significant effect of cocaine dose (F4,140 = 11.392; p < 0.001) but no effect of presession treatment (F4,140 = 1.283; p = 0.280) or their interaction (F16,140 = 0.598; p = 0.881). Likewise, a two-way ANOVA of the effects of nisoxetine on response rates indicated a significant effect of cocaine dose (F4,140 = 13.947; p < 0.001) but no effect of presession treatment (F4,140 = 0.660; p = 0.621) or their interaction (F16,140 = 0.263; p = 0.998). Table 1 shows the effects of these two compounds at the 0.32 mg/kg/inj dose of cocaine that maintained the highest rates of responding. Post hoc comparisons indicated that the highest dose of citalopram significantly decreased response rates compared with no treatment (t = 3.426; p = 0.008). In contrast, none of the effects of nisoxetine was significant.
Effects of presession treatments with citalopram or nisoxetine on cocaine self-administration
Presession treatment with the lowest (10 mg/kg) dose of AHN 1-055 had no effects on response rates or cocaine intake, whereas 32 mg/kg increased response rates at the lowest dose of cocaine and those occurring in extinction (Fig. 4B). That dose and a higher dose of AHN 1-055 decreased the normally high rate of responding maintained by 0.32 mg/kg/inj cocaine (Fig. 4B, top; compare upward triangles with filled circles). The highest dose generally shifted the cocaine dose-effect curve downward, with no dose of cocaine maintaining responding at levels above those maintained in extinction (Fig. 4B, downward triangles). The two highest doses of AHN 1-055 decreased cocaine intake (Fig. 4B, bottom). A two-way ANOVA of effects on response rates indicated a significant effect of cocaine dose (F4,120 = 3.194; p = 0.016), an effect of presession treatment, and the interaction that approached significance (F3,120 = 2.414 = 0.070; F12,120 = 1.811; p = 0.054, respectively). Post hoc tests indicated that the effects of 32 (t = 3.144; p = 0.013) and 100 (t = 3.108; p < 0.014) mg/kg AHN 1-055 significantly decreased response rates maintained by the 0.32 mg/kg/inj dose of cocaine. A representative performance shows that 32.0 mg/kg AHN 1-055 increased response rates maintained by 0.03 mg/kg/inj cocaine (Fig. 5, compare top and third panels) and that the pattern of responding was characteristic of fixed-ratio schedules but most often ceased before the end of each component.
AHN 2-005 dose-dependently decreased the maximal response rates maintained by cocaine and decreased cocaine intake (Fig. 4C). At the highest dose of AHN 2-005, no dose of cocaine maintained responding at levels above those maintained in extinction (Fig. 4C; compare downward triangles with filled circles). The ANOVA of effects of AHN 2-005 on response rates indicated a significant effect of cocaine dose (F4,110 = 7.578; p < 0.001), presession treatment (F3,110 = 3.627 = 0.015), and a nonsignificant interaction (F12,110 = 1.441; p = 0.158). Post hoc tests indicated that the effects of 32 (t = 2.866; p = 0.03) and 100 (t = 4.199; p < 0.001) mg/kg AHN 2-005 significantly decreased response rates maintained by the 0.32 mg/kg/inj dose of cocaine. The representative performance (Fig. 5; panel 4) shows that AHN 2-005 increased the pauses before initiation of responding and that responding often ceased before the end of the component (compare fourth with first panel).
The effects of JHW 007 resembled those of AHN 2-005, with dose-dependent decreases in the maximal effects of cocaine (Fig. 4D). At 32 mg/kg, the maximal effect of cocaine was significantly decreased, and at 100 mg/kg, JHW 007 decreased responding at all doses of cocaine (Fig. 4D; compare triangles with filled circles) to levels maintained in extinction (point above 0). The two highest doses of JHW 007 also decreased cocaine intake (Fig. 4D, bottom). The ANOVA indicated a significant effect of cocaine dose (F4,105 = 9.519; p < 0.001), presession treatment (F3,105 = 6.027; p < 0.001), and a significant interaction of the two (F12,105 = 1.959; p = 0.035). Post hoc tests indicated that the effects of 32 (t = 3.242; p = 0.01) and 100 (t = 4.990; p < 0.001) mg/kg AHN 2-005 significantly decreased response rates maintained by the 0.32 mg/kg/inj dose of cocaine. As with AHN 2-005, performances after treatment with the intermediate dose of JHW 007 were characterized by increased lengths of pauses and cessation of responding before the end of the component (Fig. 5, compare fourth panel with bottom).

Effects of presession treatments with monoamine uptake inhibitors on responding maintained by cocaine injection or food presentation. Ordinates, response rates as percentage of control response rates (sessions before drug tests), which averaged 0.64 (±0.14) and 0.44 (±0.15) responses/s, respectively, for food- and cocaine-maintained responding. Abscissae, milligrams per kilogram of test compounds administered per os, log scale. Closed circles and open squares, behavior maintained by cocaine (0.32 mg/kg/inj i.v.) and food, respectively. AHN 1-055, AHN 2-005, JHW 007, and methylphenidate were administered 180, 300, 240, and 60 min before sessions. Responding was from the fourth 20-min component of the session (see Materials and Methods). Each point represents the mean ± S.E.M. of from six to nine subjects.
A comparison of the effects of the BZT analogs and methylphenidate on responding maintained by cocaine and food reinforcement was conducted at the 0.32 mg/kg/inj dose of cocaine that maintained maximal response rates (Fig. 6). Rates of responding maintained by food reinforcement in the fourth component of the session averaged 0.64 responses/s, whereas those maintained by 0.32 mg/kg/inj cocaine (also in the fourth component) averaged 0.44 responses/s. These values were not significantly different (t15 = 0.926; p = 0.369). All four compounds decreased rates of responding maintained by cocaine (Fig. 6, filled symbols) at doses below those that altered responding maintained by food reinforcement. Two-way ANOVA indicated a significant effect of the reinforcer for both AHN 2-005 (F1,18 = 6.390; p = 0.026) and methylphenidate (F1,17 = 10.104; p = 0.007), whereas the reinforcer was not significant for AHN 1-055 (F1,18 = 1.055; p = 0.323) and approached significance for JHW 007 (F1,19 = 4.715; p = 0.052). Nonetheless, post hoc comparisons indicated significance of reinforcer for each of these two drugs at doses of 32 mg/kg. Two-way ANOVA also indicated a significant effect of reinforcer for citalopram (F1,40 = 15.279; p < 0.001) but not nisoxetine (F1,40 = 3.109; p = 0.085). Post hoc comparisons indicated that the significance for reinforcer obtained with citalopram was due to the increase in responding maintained by food reinforcement at the 100 mg/kg dose. AHN 1-055 had approximately a 3-fold greater potency in its effects on responding maintained by cocaine compared with food-reinforced responding, whereas with the other drugs, the potency difference was greater but could not be reliably estimated.
Discussion
In the present study, cocaine and methylphenidate were equipotent and maintained comparable rates of self-administration. Compared with its actions at the DAT, methylphenidate has 6-fold lower NET affinity and even lower SERT affinity (Gatley et al., 1996). In contrast, cocaine has comparable DAT and SERT affinities and lower NET affinity (Ritz et al., 1987). As previously reported for primates, the selective NET inhibitor, nisoxetine (Woolverton, 1987), was not self-administered. Furthermore, citalopram was not self-administered, as reported with other selective SERT inhibitors (e.g., Howell and Byrd, 1995). Thus, consistent with previous studies, the present results suggest that reinforcing effects of methylphenidate and cocaine are mediated by the DAT.
Despite DAT affinity, none of the BZT analogs presently studied maintained self-administration with rates of responding as high as those maintained by cocaine. As with AHN 1-055 in this study, a previous study using primates (Woolverton et al., 2000) found rates of responding maintained by 3′-Cl-BZT and 4′-Cl-BZT lower than those maintained by cocaine but greater than those maintained by vehicle. In addition, neither BZT itself (Woolverton et al., 2000) nor 3′-,4″-diCl-BZT (Woolverton et al., 2001) maintained responding significantly above vehicle levels, as with AHN 2-005 and JHW 007 in the present study. Thus, under some conditions, reinforcing effects can be obtained with some BZT analogs (and cocaine-like discriminative stimulus effects and place conditioning as described above). The conditions that promote self-administration of AHN 1-055 are not entirely clear and may or may not involve the brief opportunities of subjects to self-administer in the present study in contrast to a more prolonged exposure, as is often studied. Nonetheless, several BZT analogs consistently yield results indicating decreased cocaine-like effects in several behavioral procedures indicative of abuse liability.
Differences in the self-administration of BZT analogs and cocaine could be due to pharmacokinetic differences among the drugs. Lile et al. (2000) suggested that the impact of uptake and distribution on binding association kinetics is a critical factor in reinforcing effectiveness of DAT inhibitors. In their study, 2β-propanoyl-3β-(4-tolyl)-tropane, a DAT inhibitor with reduced reinforcing effectiveness in primates, had a lower apparent DAT association rate compared with cocaine in rats, presumably because of slower uptake and distribution (see also Wee et al., 2006; Kimmel et al., 2008). However, as also noted (Woolverton et al., 2002; Kimmel et al., 2007), actions at other sites may obscure relationships between reinforcing effectiveness and rate of DAT occupancy. Regarding the present findings, the differences among BZT analogs as reinforcers may be more clearly related to rate of occupancy because these compounds are relatively selective for the DAT among monoamine uptake sites (Katz et al., 2004).
Pharmacokinetic studies have found BZT analogs in brain shortly after injection, with only small differences in central nervous system permeability compared with cocaine (Raje et al., 2003; Othman et al., 2007). Despite this rapid central nervous system availability, Tanda et al. (2005) showed slower increases in extracellular dopamine produced by 4-Cl-BZT compared with cocaine. The disparity between onset of effects and relatively similar distributions to brain may be due to rates of association to the DAT. Desai et al. (2005a,b) showed substantially reduced apparent association rates of several BZT analogs, including those presently studied, compared with cocaine. Studies of the binding of [3H]JHW 007 in vitro indicate that at least some of its slower apparent association rate compared with that of cocaine, is due to a slow “on rate” (Kopajtic et al., 2006). Differences in binding kinetics may be related to dynamics at the site of action that differ among between cocaine and BZT analogs (e.g., Beuming et al., 2008; Loland et al., 2008). We recently suggested that several BZT analogs preferentially bind to inward-facing conformations of the DAT, whereas cocaine analogs preferentially bind to outward-facing conformations (Loland et al., 2008), and this difference in conformational preference predicted cocaine-like behavioral effects. It is possible that conformational preferences of BZT analogs affect their association kinetics, which in turn contributes to differences in their in vivo pharmacology.
One question that has been considered important is whether presession treatments decrease cocaine-reinforced responding selectively. In the present study, methylphenidate and the BZT analogs selectively decreased cocaine-compared with food-reinforced responding. Likewise, Glowa et al. (1995) reported that GBR 12909 decreased responding maintained by cocaine in rhesus monkeys at doses that had little or no effect on food-maintained responding. However, a previous study (Skjoldager et al., 1993) in primates failed to obtain a substantial effect of similar doses of GBR 12909 on responding maintained by cocaine injection, and a study in rats (Barrett et al., 2004) did not find the same degree of selectivity. One difference between some of the studies is the method of drug delivery. Glowa et al. (1995) delivered GBR 12909 by an intravenous infusion that in some, but not all, experiments was 15 min in duration, whereas its duration in the other two studies was presumably shorter. Slow infusions of GBR 12909 probably created a slow apparent association rate similar to the slow apparent association of the present BZT analogs, which may have contributed to the selective effects obtained.
In the study by Barrett et al. (2004), low response rates when responses were not reinforced (extinction), or when responding was maintained by low concentrations of liquid food, were increased by d-amphetamine and GBR 12909. Likewise, in the present study, increases in responding during extinction were obtained with methylphenidate and AHN 1-055 but not with AHN 2-005 or JHW 007. Increases in extinction of responding previously maintained by drug injections have been referred to as “reinstatement,” and it has been suggested that the increases indicate that the drug treatment will predispose a recovering addict to relapse. If such increases in responding are indications of potential relapse, the present results suggest differences among drugs acting at the DAT and a low potential for this effect with AHN 2-005 and JHW 007.
Several other studies of DAT inhibitors have found that cocaine analogs decreased responding maintained by cocaine at some dose (e.g., Carroll et al., 2006), although selective effects on responding reinforced with cocaine are not always obtained (Dworkin et al., 1998; Howell et al., 2000). It is interesting that the DAT occupancy produced by ED50 doses for decreasing cocaine self-administration seems to differ among cocaine analogs (Lindsey et al., 2004; Howell et al., 2007), which may contribute to differences in their effects on cocaine self-administration. Among BZT analogs, there are also differences in the locomotor stimulant effects produced by comparable amounts of DAT occupancy (Desai et al., 2005a). Variations in locomotor stimulation resulting from comparable binding also have been reported for a number of other dopamine uptake inhibitors (Rothman et al., 1992; Vaugeois et al., 1993). Differences in occupancy producing in vivo effects may be a reflection of how dopamine uptake inhibitors interact with the DAT (e.g., Loland et al., 2008), which in turn may ultimately translate to differences that affect the drug's abuse liability and may be exploited for the development of medications to treat cocaine abuse.
Methylphenidate, when administered before cocaine self-administration, produced dose-related leftward shifts in the cocaine dose-effect curve. Similar results have been reported for other indirect dopamine agonists, including the DAT inhibitor, GBR 12909, and d-amphetamine (Barrett et al., 2004). Schenk (2002) also reported that GBR 12909 and the cocaine analog, WIN 35,428, shifted the cocaine self-administration dose-effect curve to the left, although in that study, maximal response rates maintained by cocaine were also decreased. Mello and Negus (1996) suggested that medications that simply alter the potency of the abused drug will be less than satisfactory, whereas those that shift the dose-effect curve downward will have the most general therapeutic utility. In the present study, doses of methylphenidate and the BZT analogs each produced decreases in responding reinforced with cocaine at doses that minimally affected food-reinforced responding. However, both AHN 2-005 and JHW 007 dose-dependently shifted the cocaine dose-effect curve downward, whereas methylphenidate shifted the curve leftward. Because a leftward shift may represent increasing reinforcing potency of cocaine, it would seem to be the least desirable of changes and one that would not translate into an effective medical treatment. Clinical trials of methylphenidate have not demonstrated success in treating cocaine abusers (Grabowski et al., 2004). Whether the different manner by which BZT analogs alter the cocaine dose-effect curve is translated into clinical efficacy awaits further studies.
In summary, the BZT analogs were selective in decreasing cocaine- versus food-maintained responding, and in contrast to the selectivity produced by methylphenidate, the BZT analogs shifted the cocaine self-administration dose-effect curve downward. In addition, the BZT analogs have been reported to be less effective than cocaine in producing several cocaine-like effects (e.g., Katz et al., 2004) and seem to have lower abuse liability than cocaine in self-administration studies. Moreover, two of the BZT analogs failed to increase (reinstate) extinguished responding previously maintained by cocaine in the present study. These findings together imply that drugs acting at the DAT have potential as pharmacotherapies and that those with decreased cocaine-like effects are prime candidates for the development of medications for cocaine dependence.
Acknowledgments
We thank Patty Ballerstadt for administrative assistance, Gianluigi Tanda and S. Barak Caine for generous donations of time and advice, and Santosh Kulkarni and Jianjing Cao for generously synthesizing sufficient quantities of the BZT analogs for the studies.
Footnotes
-
This work was supported by the Intramural Research program of the National Institutes of Health National Institute on Drug Abuse.
-
Parts of this work were previously presented at the following conference: Hiranita T, Newman AH, and Katz JL (2008) Assessment of reinforcing effects of benztropine analogs and their effects on cocaine self-administration in rats: comparisons with monoamine uptake inhibitors; 2008 Annual Meeting of the American Society for Pharmacology and Experimental Therapeutics; 2008 Apr 5–9; San Diego, CA. American Society for Pharmacology and Experimental Therapeutics, Bethesda, MD.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.108.145813.
-
ABBREVIATIONS: DAT, dopamine transporter; SERT, serotonin transporter; NET, norepinephrine transporter; BZT, benztropine; AHN 1-055, 3α-[bis(4′-fluorophenyl)methoxy]-tropane; AHN 2-005, N-allyl-3α-[bis(4′-fluorophenyl)methoxy]-tropane; JHW 007, N-(n-butyl)-3α-[bis(4′-fluorophenyl)methoxy]-tropane; LED, light-emitting diode; FR, fixed ratio; inj, injection; ANOVA, analysis of variance; GBR 12909, 1-{2-[bis-(4-fluorophenyl)methoxy]ethyl}-4-(3-phenylpropyl)piperazine; WIN 35,428, (-)-carbomethoxy-3β-(4-fluorophenyl)tropane.
- Received September 5, 2008.
- Accepted February 18, 2009.
- U.S. Government work not protected by U.S. copyright





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}