


Abstract
Efflux transport of para-aminohippuric acid (PAH) across the blood-brain barrier (BBB) has been demonstrated by use of the brain efflux index (BEI) method. PAH was eliminated from the ipsilateral cerebrum extensively with an apparent efflux rate constant of 0.0587 (min−1) after microinjection into a cerebral cortex region termed Par2. This efflux transport showed a saturation with the Michaelis constant of approximately 400 μM. No more than 3% dose of PAH and carboxyl-inulin, used as a reference compound showing limited permeability at the BBB, were found in the contralateral cerebrum, cerebellum or cerebrospinal fluid up to 20 min after administration. Under saturated conditions for carrier-mediated efflux of PAHvia the blood-cerebrospinal fluid barrier, the BEI value of PAH did not change significantly, which suggested that blood-cerebrospinal fluid barrier was not responsible for the elimination of PAH from the brain after microinjection. No significant metabolism of PAH was demonstrated in the brain for at least 20 min after microinjection, and most of the radioactivity in the ipsilateral and contralateral carotid veins was as the intact form. With the distribution volume of PAH, 0.800 ml/g brain, obtained from the brain slice uptake experiment, the apparent efflux clearance was calculated as 46.9 μl/min/g brain. In addition, the influx clearance of PAH across the BBB determined by the in vivo brain uptake index method was much smaller than the efflux clearance, which demonstrates that BBB transports PAH selectively from the brain to the circulating blood.
Numerous drugs, including nonsteroidal anti-inflammatory drugs, β-lactam antibiotics, quinolone antibiotics and the anti-AIDS agent azidothymidine, have exhibited limited distribution in the brain (Terasaki and Pardridge, 1988; Suzuki et al., 1989a). Based on pharmacokinetic analysis, it has been suggested that this low distribution is caused by a significant efflux compared with the influx at the BBB (Adkison et al., 1994; Wang and Sawchuk, 1995). However, with the exception of P-glycoprotein, only limited information is available regarding selective efflux transport at the BBB (Suzukiet al., 1996).
Recently we developed a novel method, termed the “brain efflux index method,” to estimate directly the efflux rate of various substances at the BBB (Kakee et al., 1996). The validity of the BEI method has been demonstrated with water and 3-O-methyl-d-glucose, the former used as a substrate representing blood flow limited elimination and the latter a substrate representing symmetrical elimination at the BBB. The BEI method may enable us to detect the selective efflux transport systems.
PAH, an organic anion, is known to be excreted in the urinevia the active organic anion transport systems located in the kidney (Pritchard, 1987, 1988; Hori et al., 1993; Takanoet al., 1994). It has also been reported as a substrate for the organic anion efflux transport systems at the BCSFB (Holloway and Cassin, 1972; Bass and Lundborg, 1973; Domer, 1973). Azidothymidine or valproic acid is also expected to be transported by putative probenecid-sensitive organic anion efflux systems at the BBB (Wonget al., 1993; Adkison et al., 1994), leading to extremely limited cerebral distribution. It would be very useful to know if the BBB has a role in pumping out organic anions from brain to blood.
The present study investigated the hypothesis that the BBB transports PAH, an organic anion, selectively from the brain to the circulating blood by the BEI method.
Materials and Method
Reagents.
[3H]PAH (181.3 Gbq/mmol), [3H]3-O-methyl-d-glucose (2782.4 GBq/mmol), [3H]inulin (11.7 GBq/g), [14C]carboxyl-inulin ([14C]inulin; 0.093 GBq/g) and [14C]butanol (0.059 GBq/mmol) were purchased from New England Nuclear (Boston, MA). HEPES, from Dojin Chemicals (Kumamoto, Japan), and xylazine, from Sigma Chemical Co. (St. Louis, MO), were both analytical grade. Ketaral 50 [ketamine hydrochloride (Sankyo Co., Tokyo, Japan)] was used as an anesthetic. All other reagents were of reagent grade and used without further purification.
BBB efflux study of [3H]PAH.
[3H]PAH and [14C]inulin, used as a reference compound, were dissolved in physiological buffer containing 122 mM NaCl, 25 mM NaHCO3, 10 mM d-glucose, 3 mM KCl, 1.4 mM CaCl2, 1.2 mM MgSO4, 0.4 mM K2HPO4 and 10 mM HEPES. This buffer was adjusted to pH 7.4 by 2 N NaOH (ECF buffer). Sprague-Dawley male rats (supplied by Charles River, Yokohama, Japan) weighing 200 to 250 g were anesthetized with an intramuscular injection of ketamine-xylazine (1.22 mg xylazine and 125 mg ketamine per kg b.wt.) and placed in a stereotaxic frame (Narishige Co., Tokyo, Japan). The BEI experiments were carried out as reported previously (Kakee et al., 1996). After removing part of the scalp, a midline incision was performed to expose the reference point on the skull called the bregma. A small hole was drilled at the Par2 region (0.2 mm anterior and 5.5 mm lateral to the bregma and 4.5 mm deep) of the left cerebrum to allow entry of an injection needle, and then 0.5 μl of a mixture of [3H]PAH (2.5 nCi or 0.2 μCi/rat) and [14C]inulin (0.25 nCi or 1.5 nCi/rat) was injected by use of a 5-μl microsyringe (Hamilton, Reno, NE) fitted with a needle (i.d. 100 μm, o.d. 350 μm, Seiseido Medical Industry, Tokyo, Japan) in the absence and presence of unlabeled PAH. The osmolarity of the injectate was adjusted by removal of NaCl from the ECF buffer to give an isotonic solution, if required. The craniometric data and the precise localization of the region to be injected were determined with a stereotaxic atlas (Paxinos and Watson, 1986). After microinjection of drug into the cerebrum, CSF was sampled from the cisterna magna at appropriate times. A small hole was opened at the sagittal midline through the suture between the interparietal and supraoccipital bones with an electrical drill (Natsume Seisakusho Co., Ltd., Tokyo, Japan). A syringe was introduced into the hole to a depth of 6 mm. Gentle suction was applied by means of this syringe to withdraw 50 to 150 μl CSF. Immediately after CSF sampling, rats were decapitated and left and right cerebrum and cerebellum were removed. After measuring the wet weight of each excised cerebrum or cerebellum, they were dissolved in 2.5 ml 2 N NaOH by incubating at 50°C for 3 hr. Then, 14 ml liquid scintillation cocktail (Hionic-fluor; Packard Instruments Corp., Meriden, CT) was added to the sample at room temperature. Radioactive counting was performed by a double-channel system for the 3H, 14C mixed samples using an LC-6000 liquid scintillation counter (Beckmann Instruments Corp., Fullerton, CA). Each sample was measured twice for 5 min and the average was used for calculations. The BEI value was obtained by the following equation (Kakee et al., 1996):
The Vbrain was determined by thein vitro uptake into brain slices as described below.
Kinetic parameters for the efflux transport of PAH across the BBB were estimated according to the following equation:
where Km,Vmax and C are the Michaelis constant (μM), the maximum transport velocity across the BBB (nmol/min/g brain) and the concentration of PAH in the brain (μM), respectively. The concentration of PAH in the brain was estimated from the concentration in the injectate, assuming that the 0.5-μl injectate was diluted 30.3-fold in the brain as reported previously (Kakee et al., 1996). Accordingly, PAH concentration in the brain was calculated by dividing the drug concentration in the injectate by 30.3. To obtain the kinetic parameters,Km and Vmax, Eadie-Hofstee plot analysis was performed by the following equation derived from equation 3.
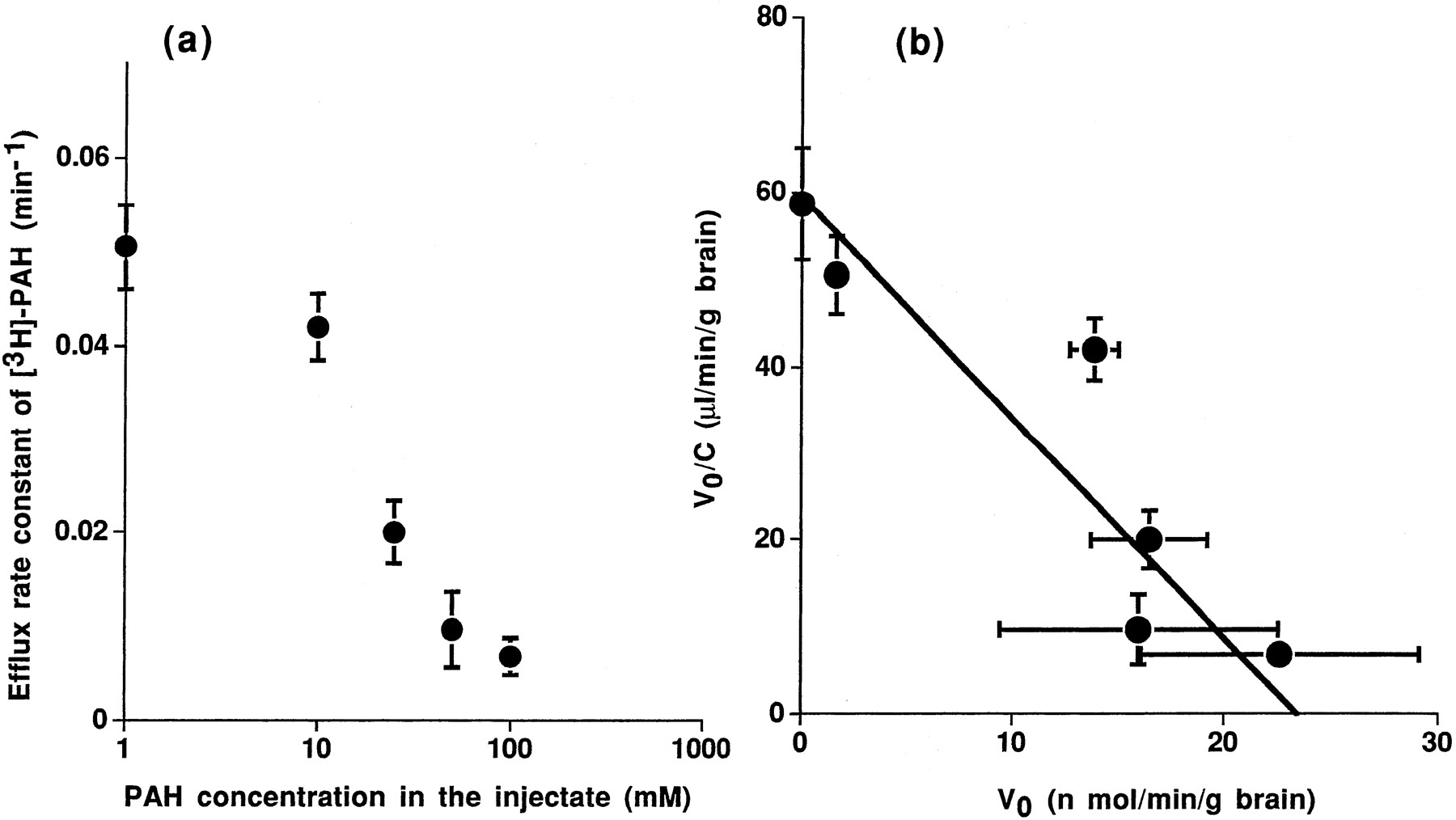
Fitting equation 4 to the data sets was carried out by an iterative nonlinear least-squares method using a program “MULTI” to obtain the kinetic parameters (Yamaoka et al., 1981). The input data were weighed as the reciprocal of the square of the observed values, and the algorithm used for the fitting was the damping Gauss Newton Method. The Km andVmax for the efflux of PAH across the BBB obtained were 396 ± 73 (μM) and 23.4 ± 2.7 (nmol/min/g brain), respectively.
Intracerebroventricular injection of [3H]PAH and [14C]inulin.
[3H]PAH (2.5 μl; 1 μCi/rat) and [14C]inulin (0.05 μCi/rat) were administered to the lateral cerebral ventricle of the left cerebrum in the absence or presence of 100 mM unlabeled PAH as reported previously (Suzukiet al., 1985, 1988, 1989b). After injection of drug, CSF was sampled from the cisterna magna at appropriate times as described above. The elimination clearances of [3H] PAH and [14C]inulin in the CSF were calculated from the following equation:
Because the CSF volume has been reported to be 250 μl/rat (Cserr and Berman, 1978), the PAH concentration in the CSF was estimated assuming that the injectate was immediately and homogeneously distributed in the CSF compartment.
To investigate the effect of a saturable efflux system at the BCSFB on the apparent elimination of PAH from the brain after microinjection into Par2, 100 mM unlabeled PAH was administered to the rat cerebral ventricle 30 sec before the BEI study with [3H]PAH. The osmolarity of the injectate was, if required, adjusted by removal of NaCl from the ECF buffer to give an isotonic solution.
Metabolism of [3H]PAH after cerebral microinjection.
An aliquot of 1 μl [3H]PAH (2 μCi/rat) was administered to the Par2 region of the left cerebrum. One milliliter of blood from the ipsilateral and contralateral carotid veins was withdrawn 5 min after administration; then rats were decapitated and the left cerebrum was removed. Plasma samples were obtained by centrifugation at 3,000 rpm for 10 min. Analytical samples were prepared following deproteinization with methanol. In the case of brain samples, they were suspended in 1.8 ml ECF buffer using a Teflon homogenizer (Iuchi Seieido Co., Ltd., Tokyo, Japan), and then centrifuged at 3,000 rpm for 10 min. A 3 ml supernatant was added to 6 ml methanol to denature proteins. After centrifugation at 3,000 rpm for 10 min, 8 ml supernatant was evaporated under N2 and then the samples were analyzed by the imaging analyzer system (BAS-3000; Fuji Photo Co., Ltd., Tokyo, Japan) following 10 × 10 cm thin-layer chromatography (Kieselgel 60F254, Merck Co., Ltd., Darmstadt, Germany). The solvent system for the thin-layer chromatography wasn-butanol/acetic acid/water (25:4:10).
Measurement of distribution volume of [3H]PAH in the brain.
The distribution volume of PAH in the brain was determined by the in vitrobrain slice uptake technique. Brain slices were prepared as reported previously with minor modification (Newman et al., 1991). After decapitating rats, brains were immediately removed and dissected in ice-cold oxygenated ECF buffer. A hypothalamic slice, 300 μm thick, was cut using a brain microslicer (DTK-2000, Dosaka EM Co., Ltd., Kyoto, Japan), and kept in oxygenated ECF buffer equilibrated with 95% O2-5% CO2. After preincubation for 5 min at 37°C, the brain slice (40–50 mg) was transferred to 50 ml oxygenated incubation medium containing 0.05 μCi/ml [3H]PAH and 0.01 μCi/ml [14C]inulin at 37°C. At appropriate times, brain slices and part of the incubation medium were stored at −20°C for the determination of drug concentrations. The apparent zero-time intercept of the [14C]inulin uptake time profile, i.e., 0.136 ± 0.006 ml/g slice (mean ± S.E., n = 6), obtained from the same study with the brain slices, was used to correct for the adsorbed water volume.
BUI of [3H]PAH, [3H]3-O-methyl-d-glucose or [3H]inulin.
The influx clearance of [3H]PAH across the BBB was determined by the BUI method reported previously (Oldendorf, 1970; Terasaki et al., 1986). An aliquot of 250 μl Ringer’s/HEPES buffer (pH 7.4, 5 mM HEPES) was injected rapidly into the left common carotid artery. The injection solution contained [3H]PAH (100 μCi/ml) and [14C]butanol (0.05 μCi/ml). In the BUI studies with [3H]3-O-methyl-d-glucose or [3H]inulin, [3H]3-O-methyl-d-glucose (10 μCi/ml) and [14C]butanol (0.5 μCi/ml) or [3H]inulin (100 μCi/ml) and [14C]butanol (0.05 μCi/ml) were dissolved in the injectate, respectively. Fifteen seconds after the carotid artery injection, the rats were decapitated. The radioactivity in the injection solution and the hemisphere ipsilateral to the injection were determined. The BUI value, extraction ratio, apparent influx clearance (CLinf) and intrinsic BBB permeability surface area product (PSinf) were calculated as follows (Oldendorf, 1970):
Results
Efflux of [3H]PAH from the brain across the BBB.
Figure 1 shows the time course of the remaining percentage of [3H]PAH in the ipsilateral cerebrum after microinjection into Par2 of rat brain. Approximately 60% of the administered dose of [3H]PAH was eliminated from the ipsilateral cerebrum within 20 min, which indicates a significant elimination of [3H]PAH from the cerebrum (fig. 1). The apparent elimination rate constant, kel, was found to be 5.87 × 10−2 ± 0.65 × 10−2min−1 by nonlinear least squares regression analysis of the observed values up to 20 min after microinjection. As shown in table 1, no significant amount of administered [3H]PAH was found in the contralateral cerebrum, cerebellum or CSF compartment at 20 min, even after an intracerebral microinjection of a high dose of radioactivity. In addition, no significant difference in the remaining percentage of [14C]inulin, an internal reference compound, was observed at 20 min after microinjection (table 1). The compartmental model analysis based on the drug elimination obtained by the BEI study and i.c.v. injection study as described below indicated that only 0.52% of the apparent elimination rate constant in the BEI study contributed to efflux from the CSF.
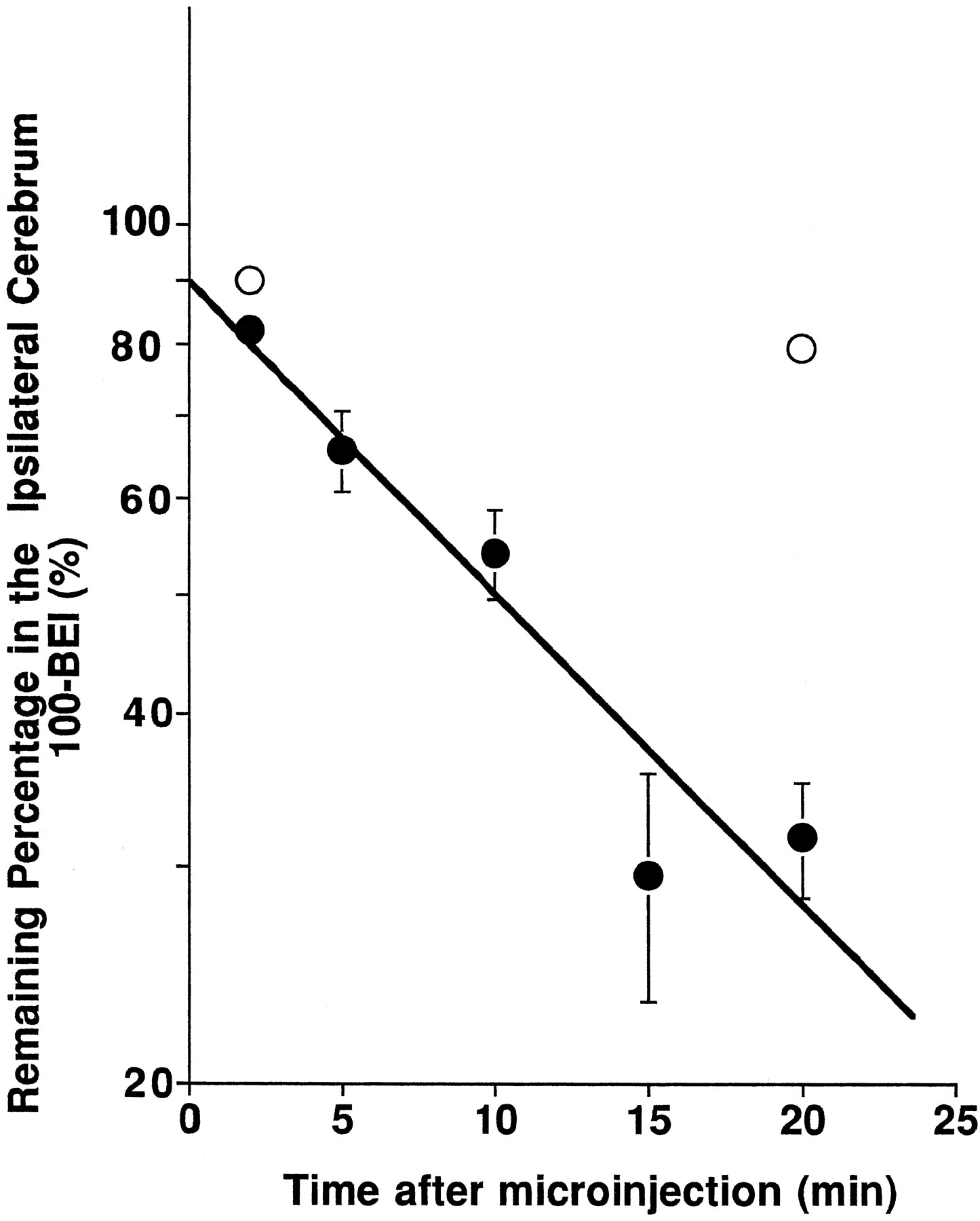
Time courses of [3H]PAH in the ipsilateral cerebrum after intracerebral microinjection in the presence of [14C]inulin as an internal reference. A mixture of [3H]PAH (2.5 nCi) and [14C]inulin (0.25 nCi) dissolved in 0.5 μl ECF buffer was injected into Par2 of rat cerebrum in the absence or presence of 100 mM unlabeled PAH. Rats were decapitated at 2, 5, 10, 15 and 20 min after microinjection. Closed and open symbols represent the time courses of the percentage of [3H]PAH remaining in the brain after administration in the absence and presence of 100 mM PAH (the mean ± S.E.;n = 3–7). The slope of the solid line represents the elimination rate constant of tracer amount of [3H]PAH, i.e. 5.87 × 10−2 ± 0.65 × 10−2 min−1, obtained by the nonlinear least squares regression analysis.
Recovered percentage of [3H]PAH and [14C]inulin in the ipsilateral, contralateral cerebrum, cerebellum or CSF after intracerebral microinjection
The BEI study of [3H]PAH was also performed at 2 and 20 min with 100 mM PAH in the injectate. Twenty minutes after injections, the remaining percentage of [3H]PAH after coadministration of unlabeled PAH was found to be 79.6 ± 1.9%, which was significantly greater that that in the absence of unlabeled PAH, i.e., 31.8 ± 3.4% (fig. 1).
The apparent elimination rate constant was decreased in a dose-dependent manner (fig. 2a). The Michaelis constant, Km, and the maximum velocity, Vmax, for the efflux of PAH across the BBB obtained were 396 ± 73 μM and 23.4 ± 2.7 nmol/min/g brain by Eadie-Hofstee plot analysis, respectively (fig.2b).

(a) Concentration dependence of the efflux of [3H]PAH at the BBB. A mixture of [3H]PAH (2.5 nCi) and [14C]inulin (0.25 nCi) dissolved in 0.5 μl ECF buffer was injected into Par2 of rat cerebrum in the presence of 1, 10, 25, 50 or 100 mM unlabeled PAH. Rats were decapitated at 2 and 20 min after microinjection. Closed symbols represent the efflux rate constant of [3H]PAH at the BBB (the mean ± S.E.; n = 6). (b) Eadie-Hofstee plot of the efflux transport of [3H]PAH at the BBB. The reciprocal of the slope of solid line, obtained by the nonlinear least squares regression analysis, represents the Km value for the transport of PAH at the BBB, as described under “Materials and Method.”
Efflux of [3H]PAH from the CSF after i.c.v. injection.
Figure 3 shows the time course of the percentage dose of [3H]PAH and [14C]inulin in the CSF after cerebroventricular administration. Based on equation 5, the apparent elimination clearance of [3H]PAH from the cerebral ventricle, CLefflux,csf, was estimated to be 26.7 and 4.0 μl/min/rat for PAH and inulin, respectively. In the presence of 1 mM PAH in the CSF, the elimination of [3H]PAH from the CSF compartment was delayed significantly (fig. 3).

Time courses of the percentage of the dose of [3H]PAH and [14C]inulin remaining in the CSF after i.c.v. injection. A mixture of [3H]PAH (1 μCi) and [14C]inulin (0.05 μCi) dissolved in 2.5 μl ECF buffer was injected into the cerebroventricle in the absence and presence of 100 mM unlabeled PAH in the injectate. Rats were decapitated at 5, 15 and 30 min after administration. Closed and open circles represent the time courses of the percentage dose of [3H]PAH remaining in the CSF in the absence and presence of 100 mM unlabeled PAH in the injectate, respectively (n = 4). Closed squares represent the time courses of the percentage dose of [14C]inulin remaining in the CSF (n = 6). The elimination clearance of tracer amounts of [3H]PAH and [14C]inulin was calculated to be 26.7 and 4.0 μl/min/rat, respectively.
After administering unlabeled PAH in the cerebral ventricle, the BEI study of [3H]PAH was also performed to examine the contribution of the saturable elimination pathway of PAHvia the BCSFB to the apparent elimination from the ipsilateral cerebrum. As shown in table2, no significant difference was observed in the BEI values of [3H]PAH at 20 min compared in the absence and presence of 1 mM unlabeled PAH in the CSF, which suggests the minimal contribution of the elimination pathwayvia the BCSFB.
BEI of [3H]PAH after intracerebroventricular injection of excess unlabeled PAH
Metabolism of [3H]PAH after cerebral microinjection.
Metabolism of [3H]PAH was investigated by the autoradiogram of the thin-layer chromatography of [3H]PAH in the ipsilateral cerebrum and carotid vein after microinjection into Par2. All the major spots of the ipsilateral cerebrum, and ipsilateral and contralateral carotid veins had the same Rf value (Rf = 0.58) as that of authentic [3H]PAH used as a standard (data not shown). Moreover, no significant metabolites were found for the brain and blood samples examined.
Comparison of efflux and influx clearance of [3H]PAH at the BBB.
The distribution volume of PAH in the brain, Vbrain, was determined in the in vitro brain slice uptake study. Figure4 shows the time course of the brain slice-to-medium concentration ratio of [3H]PAH. No significant difference in the slice-to-medium concentration ratio between 60 and 120 min was observed after incubation, giving a steady-state slice-to-medium ratio of 0.800 ± 0.051 ml/g brain (n = 6, mean ± S.E.). Incorporating the apparent elimination rate constant (5.87 × 10−2 ± 0.65 × 10−2 min−1, fig. 1), and the distribution volume in the brain (0.800 ± 0.051 ml/g brain) into equation 2, the apparent BBB efflux clearance of PAH was calculated to be 46.9 ± 3.8 μl/min/g brain.

Time courses of [3H]PAH uptake by rat brain slices. Rat brain slices were incubated with 0.05 μCi/ml [3H]PAH and 0.01 μCi/ml [14C]inulin at 37°C. At appropriate times, the radioactivity in the brain slices and incubation medium were measured, and the slice-to-medium concentration ratio was estimated.
By means of the in vivo carotid artery injection technique, the BBB influx clearances for [3H]PAH, [3H]inulin and [3H]3-O-methyl-d-glucose were determined. As shown in table 3, [3H]3-O-methyl-d-glucose was significantly taken up by the brain and the brain extraction percentage, i.e., 10.1 ± 0.8%, was very similar to that reported by us and others (Oldendorf et al., 1982;Kakee et al., 1996). In contrast to the significant efflux of [3H]PAH from the brain after cerebral microinjection, very limited BBB influx clearance of [3H]PAH was obtained, i.e.,12.1 ± 0.9 μl/min/g brain, which was not significantly different from that of [3H]inulin, used as a vascular space marker (table 3).
BUI, brain extraction and BBB influx clearance of [3H]PAH, [3H]inulin or [3H]3-O-methyl-d-glucose3-a
Discussion
Several acidic drugs have been reported to show significantly limited cerebral distribution compared with neutral or basic drugs, by BUI, brain perfusion, brain microdialysis or in vivointravenous injection techniques (Nau and Loscher, 1982; Cornfordet al., 1985; Suzuki et al., 1989a). Also, theKp value, the concentration ratio of drug in brain to that in plasma, has been found to be well below unity at steady state. The following four possibilities have been proposed to explain the limited cerebral distribution of acidic drugs: 1) protein binding in plasma was very high compared with that in brain, 2) the efflux clearance was greater than the influx clearance across the BBB, 3) the efflux via BCSFB was significant and 4), all these possibilities combined. Recently, pharmacokinetic model analysis after i.c.v. injection, perfusion or brain microdialysis experiments have led us to consider the second possibility to explain the limited distribution of the acidic drugs such as valproic acid (Adkisonet al., 1994). As reported previously, the BEI method has been shown to characterize the BBB efflux transport process (Kakeeet al., 1996). By this newly developed BEI method, we have determined the efflux rate of PAH, selected as a model substrate of an acidic drug.
PAH is well known to be actively excreted at the kidney or choroid plexus via organic anion transporters (Holloway and Cassin, 1972; Bass and Lundborg, 1973; Domer, 1973). This compound also has a low distribution in the brain similar to several other substances. Because the protein binding of PAH in the plasma has been reported to be only 2.4% (Elbourne et al., 1990), the first possibility can be rejected as an explanation of the limited distribution of PAH in the brain. PAH molecules microinjected into Par2 of the rat hemisphere disappeared rapidly from the ipsilateral cerebrum with an elimination rate constant of 5.87 × 10−2min−1 (fig. 1). Also, we could not detect the eliminated PAH in the contralateral cerebrum, cerebellum or CSF (table 1), which indicated that only a limited amount of [3H]PAH is transferred from the injection site to the other cerebral regions, and the elimination of [3H]PAH is attributed to efflux from the brain to blood across the BBB.
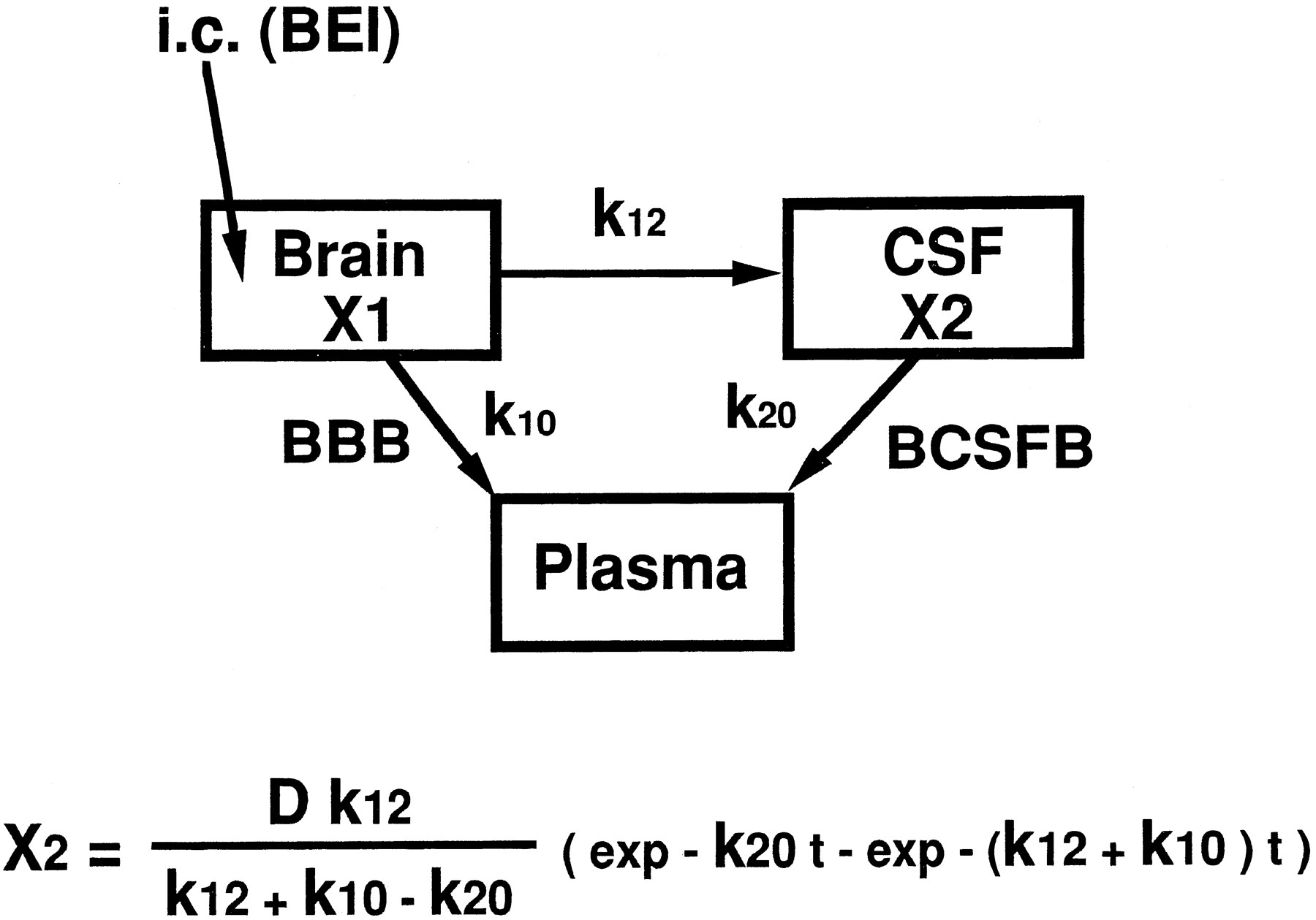
Several acidic drugs such as β-lactam antibiotics, including benzylpenicillin, or valproic acid have been recognized as substrates for probenecid-sensitive anion transport systems at the BCSFB, by use of in vivo i.c.v. injection, perfusion or in vitro uptake techniques involving the isolated choroid plexus (Suzuki et al., 1987a,b, 1989b, 1996; Adkison et al., 1994; Ogawa et al., 1994). The elimination clearance of these compounds in the CSF after i.c.v. injection was approximately 22.5 μl/min/rat (Suzuki et al., 1989b), which was 5.6-fold greater than that of inulin used as a CSF bulk-flow marker. Similarly, the CSF elimination clearance of PAH was 26.7 μl/min/rat after i.c.v. injection, which was 6.7-fold greater than the bulk-flow rate (fig. 3), and its elimination profile was similar to that reported previously (Jakobson, 1987). The elimination clearance of PAH in the CSF can be considered to include the bulk-flow rate, active efflux at the BCSFB and efflux at the BBB after diffusion in the brain interstitial space. The active efflux clearance of PAH at the BCSFB could be estimated as 26.7 μl/min/rat at the most. On the other hand, in [3H]inulin, the apparent elimination clearance from the CSF, 4.0 μl/min/rat, which was similar to the reported value, 2.9 μl/min/rat estimated by use of blue dextran (Suzuki et al., 1985), indicated only the CSF bulk-flow rate, because its diffusion was extremely limited by its high molecular weight, 5000. The elimination of [3H]PAH was significantly reduced by unlabeled 1 mM PAH in the CSF, which shows that the active efflux system at the BCSFB was significantly inhibited under these conditions (fig. 3). In this situation, it would be possible to predict the contribution of BCSFB to the elimination from the brain by the BEI method. The proposed BEI pharmacokinetic model contained three compartments, brain, CSF and plasma, and the rate constants among each compartment were defined as follows:k12, brain to CSF which means the diffusion process in the brain interstitial space,k10; brain to plasma which means the efflux process at the BBB; and k20, CSF to plasma which means the efflux process at the BCSFB (fig.5). The diffusion process in the brain interstitial space from CSF to brain was neglected in this model to give the maximum value to the rate constant ofk12 in the BEI study. The drug amount in the CSF compartment (X2) after intracerebral administration can be obtained by the equation as described in figure5. In this condition, the summation of k12and k10 was already obtained as 0.058 min−1 by the BEI study. Similarly, the maximum k20 value was also obtained as 0.520 min−1 by the initial elimination curve of [3H]PAH up to 5 min in the i.c.v. injection study. According to the equation as described in figure 5, the drug amount in the CSF (X2) showed the highest level at 4.7 min after intracerebral microinjection to the Par2 region. Considering the injected dose (0.2 μCi/rat) and the detection limit of radioactivity in the CSF, the ratio ofk12 to the summation ofk12 and k10 was calculated as 5.2 × 10−3. Accordingly, only 0.52% of the apparent elimination constant, estimated by the BEI technique, was estimated to be caused by elimination from the CSF. Moreover, to assess the contribution of CSF elimination, we have performed a BEI study of [3H]PAH under saturated conditions of active efflux at the BCSFB. No significant difference between the two conditions was observed for the BEI values at 20 min after microinjection into Par2 (table 2), which suggests that elimination from the BCSFB is not responsible for the apparent elimination of [3H]PAH from the ipsilateral cerebrum.

A schematic diagram of the three-compartmental pharmacokinetic model in the BEI study.
PAH has been known to exist in its intact form in plasma after intravenous administration, but excreted into urine, 80% as the intact form and 20% as an acetylated metabolite, which indicates that PAH is partly transformed to acetylated PAH in the kidney (Elbourne et al., 1990). However, there is no information on the metabolism of PAH in the brain. No major metabolite was found in the ipsilateral cerebrum, ipsilateral and contralateral carotid veins up to at least 20 min after administration, which shows that PAH was effluxed from the brain at the BBB in its intact form.
Recently, several reports have shown asymmetrical transport at the BBB and these have used pharmacokinetic model analysis after i.c.v. injection, cerebroventricular perfusion or brain microdialysis with model compounds such as valproic acid, an anti-AIDS agent azidothymidine or quinolone antibiotics, all well known to exhibit limited distribution in the brain (Wong et al., 1993;Adkison et al., 1994; Wang and Sawchuk, 1995). Moreover, it has been also found that P-glycoprotein, i.e. the MDR-1 gene product, functions at the BBB, pumping out several antitumor agents including vincristine and immunosuppressants such as cyclosporin A; this has been achieved by in vitro uptake experiments with primary brain capillary endothelial cells (Tatsuta et al., 1992; Tsuji et al., 1992, 1993), an in vivo brain ischemia model (Sakata et al, 1994; Ohnishi et al, 1995) and the in vivo MDR 1a gene knockout mouse (Schinkel et al., 1994). The influx rate of PAH has been determined by the intracarotid artery injection technique and has an extremely low BUI value similar to that of inulin used as a vascular space marker (table 3). The apparent PSinf value was calculated to be 12 μl/min/g brain (table 3). Suzuki et al. have already reported that the relevance of carrier-mediated transport can be detected using the brain perfusion technique for the influx of benzylpenicillin, an organic anion with an influx clearance of 5.48 μl/min/g brain (Suzuki et al., 1989a). This value is below the detection limit of the BUI experiment. Considering that the apparent BBB efflux clearance of PAH obtained using BEI and thein vitro uptake technique with brain slices (figs. 1 and 4) was 46.9 μl/min/g brain, the efflux clearance is at least 3-fold greater than the influx clearance. These results led us to conclude that there is asymmetrical transport of PAH across the BBB, with selective transportation from the brain to blood.
We have measured the partition coefficient (P) of PAH in octanol/ECF buffer and obtained 8.69 × 10−4 as the P value. Levin (1980) has already reported a good relationship between the parameter given by the octanol/water partition coefficient divided by the square root of the molecular weight and rat brain capillary permeability. Based on this report, the permeability of PAH was calculated to be 2.6 μl/min/g brain, which was 1/18 the observed efflux clearance. Moreover, the apparent elimination constant obtained by the BEI method decreased 88% in the presence of 100 mM PAH in the injectate, which suggests that passive diffusion is not a significant factor in the elimination from the brain. In fact, this efflux transport showed the saturation with the Kmvalue of 396 μM (fig. 2). This value was very similar to theKm value for the transport of PAH in the kidney (Hori et al., 1993). As yet, only MDR1 has been reported to pump out several agents from the brain to blood at the BBB (Tsuji et al., 1992; Schinkel et al., 1994). However, because it is generally believed that the substrates for MDR 1 seem to be cationic and hydrophobic in nature, one could assume that non-P-glycoprotein efflux transport systems are responsible for the efflux of PAH from the brain to blood across the BBB.
In a previous report, we demonstrated the validity of the BEI method, with [3H]water and [3H]3-O-methyl-d-glucose, the former used as a substrate representing blood flow limited elimination and the latter a substrate representing symmetrical elimination at the BBB (Kakee et al., 1996). This sensitive method allowed us to obtain an accurate efflux rate for the test substances. With the BEI method, we have demonstrated the asymmetrical transport of PAH across the BBB. It is important to identify the efflux transport mechanisms of PAH across the BBB and clarify the difference between the putative efflux system at the BBB and the organic anion transport systems in the kidney.
In conclusion, the present study provides direct in vivoevidence that the BBB selectively transports PAH, an organic anion, from brain to circulating blood.
Footnotes
-
Send reprint requests to: Yuichi Sugiyama, Ph.D., Professor, Department of Pharmaceutics, Faculty of Pharmaceutical Sciences, University of Tokyo, Hongo, Bunkyo-ku, Tokyo 113, Japan.
-
↵1 This work was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science and Culture, Japan and by Japan Health Foundation Drug Innovation Science Project.
- Abbreviations:
- BBB
- blood-brain barrier
- CSF
- cerebrospinal fluid
- BCSFB
- blood-cerebrospinal fluid barrier
- BEI
- brain efflux index
- BUI
- brain uptake index
- Par2
- Parietal Cortex Area2
- i.c.v.
- intracerebroventricular
- HEPES
- 4-(2-hydroxyethyl)-piperazineethanesulfonic acid
- PAH
- para-aminohippuric acid
- AIDS
- acquired immune deficiency syndrome
- MDR
- multi-drug resistance
- Received December 13, 1996.
- Accepted July 29, 1997.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}