

Abstract
Activation of cAMP synthesis by intracellular Ca2+ is thought to be the main mode of cAMP generation in the brain. Accordingly, the Ca2+-activated adenylyl cyclases I and VIII are expressed prominently in forebrain neurons. The present study shows that the novel adenylyl cyclase type IX is inhibited by Ca2+ and that this effect is blocked selectively by inhibitors of calcineurin such as FK506 and cyclosporin A. Moreover, adenylyl cyclase IX is inhibited by the same range of intracellular free Ca2+ concentrations that stimulate adenylyl cyclase I. Adenylyl cyclase IX is expressed prominently in the forebrain. Substantial arrays of neurons positive for AC9 mRNA were found in the olfactory lobe, in limbic and neocortical areas, in the striatum, and in the cerebellar system. These data show that the initiation of the cAMP signal by adenylyl cyclase may be controlled by Ca2+/calcineurin and thus provide evidence for a novel mode of tuning the cAMP signal by protein phosphorylation/dephosphorylation cascades.
Molecular cloning of mammalian adenylyl cyclase (Krupinski et al., 1989) has led to the discovery of nine different isotypes. These can be classified broadly into Ca2+-stimulated cyclases, Ca2+-inhibited cyclases, and protein kinase C-activated cyclases on the basis of their distinct regulation by intracellular Ca2+ and protein phosphorylation (for review, see Sunahara et al., 1996; Antoni, 1997). Current evidence indicates that all three classes of adenylyl cyclase are expressed in the mammalian brain (for review, see Mons and Cooper, 1995; Antoni et al., 1998).
The Ca2+-stimulated enzymes adenylyl cyclase I (AC1) and VIII (AC8) have been assigned important roles in synaptic plasticity (Xia and Storm, 1997). In terms of neurotransmitter function, AC1 (Wayman et al., 1994) and AC8 (Cali et al., 1994) are both activated by Ca2+/calmodulin. Moreover, AC1 may generate cAMP as a coincidence detector for concerted signals from Gs-coupled receptors and ionotropic receptors that trigger changes in the membrane potential and an increase of intracellular free Ca2+.
Adenylyl cyclases II (AC2) and VII (AC7) are activated by protein kinase C (see summary by Ishikawa, 1998). Further, AC2 is also a coincidence detector stimulated by Gβγ subunits in the context of activation by Gsα (Tang and Gilman, 1991; Chen et al., 1995). AC2 mRNA is abundant in the brain, including the neocortex and the limbic lobe (Mons and Cooper, 1995). AC7 has a more restricted distribution in the brain (Mons and Cooper, 1995), and its functional properties have not been analyzed in detail.
With respect to Ca2+-inhibited adenylyl cyclases, only the striatum and the mesolimbic dopaminergic system have been reported to express significant levels of AC5 (Glatt and Snyder, 1993;Mons and Cooper, 1995). Low levels of AC3 (Xia et al., 1992) and AC6 (Mons and Cooper, 1995) mRNA have been reported in whole brain.
By far, the most abundant cerebral adenylyl cyclase at the mRNA level appears to be AC9 (Paterson et al., 1995; Premont et al., 1996). The properties of AC9 are controversial, because studies from this laboratory (for review, see Antoni et al., 1998) suggested that receptor-induced synthesis of cAMP by AC9 is inhibited by calcineurin (protein phosphatase 2B). Others (Premont et al., 1996) have reported no effect of Ca2+ on AC9 overexpressed in Sf9 insect cells. Given the potential functional significance of a calcineurin-regulated adenylyl cyclase in the brain, we have analyzed the properties of AC9 overexpressed in human embryonic kidney (HEK-293) cells and compared the Ca2+ sensitivity of AC9 with that of AC1 in this system. Further, we have investigated the distribution of AC9 in mouse and rat forebrain.
The results showed that AC1 and AC9 are regulated reciprocally by intracellular free Ca2+. Moreover, the inhibition of AC9 by Ca2+ was blocked by the calcineurin inhibitors FK506 and cyclosporin A. Expression of AC9 mRNA was confined to the gray matter and appeared mainly neuronal. The limbic lobe, the neocortex, and the cerebellum all expressed AC9 mRNA. Expression of AC9 mRNA and protein was highest in the hippocampus. In sum, these studies reveal a novel mode of cAMP signaling in the brain in which the initiation of the cAMP signal is controlled by Ca2+/calcineurin.
MATERIALS AND METHODS
Animals. Adult (6–8 weeks old) male BALB/c mice and Wistar rats were housed under constant temperature and lighting (on, 5:00 A.M.; off, 7:00 P.M.) and had free access to pelleted food and tap water.
In situ hybridization histochemistry. Animals were decapitated, and the brains were removed rapidly from the skull and processed as previously described (Rosie et al., 1992; Paterson et al., 1995). 35S-labeled riboprobes for AC9 were prepared by transcribing segments of the mouse (plasmid JP142) (Paterson et al., 1995) or rat AC9 cDNA (nucleotides 1107–1450) (J. M. Paterson, unpublished data) with the requisite RNA polymerases and used as previously described (Rosie et al., 1992; Paterson et al., 1995). Brain sections were exposed to x-ray film for 3–6 d or dipped in Kodak NTB-2 or Ilford K5 emulsion and exposed for 2–6 weeks. Cell nuclei were counterstained with pyronin. Measurements of optical densities and grain counts were performed according to published procedures (Rosie et al., 1992).
Immunodetection of AC9 protein. Antisera against the synthetic pentadecapeptide KQLSSNTHPKHCKYS (Severn Biotech, Kidderminster, UK) at the N-terminal region of AC9 were raised in rabbits against the synthetic peptide coupled to purified protein derivative (Lachmann et al., 1986). A similarly prepared Tyr-peptide-PPD conjugate derived from YEASELSKLNVSKSV at the C terminus of the protein also was used to immunize BCG-primed hens (courtesy of Professor Peter Sharp, The Roslin Institute, Edinburgh, Scotland).
For radioimmunoassay and immunoblot analysis the rats were decapitated, and brain regions were dissected rapidly while the brain was cooled on a glass plate on wet ice. The hippocampus, the cerebellum, the hypothalamus, and the adenohypophysis were homogenized in 50 mm Tris-HCl, 5 μg/ml leupeptin, 7 μg/ml pepstatin, 1 mm EGTA, 20 μl/ml Trasylol (Bayer, Wuppertal, Germany), and 1 mm MgSO4, pH 7.4, at 4°C; crude membranes were prepared as previously described (Antoni et al., 1985). Similar membrane extracts were prepared from HEK-293 cell lines and mouse corticotrope tumor (AtT20) cells.
For immunoblots, membranes prepared from 10 mg of wet tissue or 107 cells were resuspended in SDS-PAGE sample buffer (50 mm Tris, pH 7.2, 2% w/v SDS, and 5% v/v mercaptoethanol). After being heated to 95°C for 5 min and centrifuged at 8000 × g for 2 min, aliquots of the supernatant were separated by SDS-PAGE on 7.5% homogenous microgels. Electroblotting was performed on the same apparatus (Pharmacia PhastSystem, Uppsala, Sweden), and the blots were probed with rabbit anti-AC9 antiserum at 1:4000 dilution in conjunction with an ECL kit (Amersham, Aylesbury, UK).
For radioimmunoassay the membranes prepared from 10 mg of wet tissue or 107 cells were solubilized in 0.5 ml of 154 mm NaCl, 50 mm Tris-HCl, pH 7.4, 1% NP40 (v/v), 0.25% deoxycholate (w/v), 1 mm EGTA, 1 mm PMSF, 1 μg/ml leupeptin, 1 μg/ml pepstatin, and 1.5 μl/ml Trasylol (Bayer) (RIPA buffer). After centrifugation for 30 min at 16,000 × g, the supernatant was collected and the aliquots were assayed for immunoreactive AC9 content. The C-terminally directed chicken serum was used at 1:30,000 final dilution, with radioiodinated peptide antigen prepared by the chloramine T method as the tracer. Bound and free ligand were separated by the double antibody precipitation method, using donkey anti-chicken IgY serum (courtesy of SAPU). The detection limit of the assay was 2 fmol of C-terminal antigen peptide per tube.
Production of stably transfected HEK-293 cells. HEK-293 cells were maintained as described previously (Paterson et al., 1995). The cells were transfected by electroporation with 10 μg of DNA per milliliter of the expression plasmid pcDNA3 (Invitrogen, De Schelp, The Netherlands) or pcDNA3 containing the cDNA of bovine AC1 (courtesy of D. R. Storm) or mouse AC9 (Paterson et al., 1995). Clonal cell lines were selected in 0.6 mg/ml of G418 and subsequently were propagated without antibiotic. Five different AC9-expressing clones that produced similar levels of cAMP were tested for the effects of Ca2+ on cAMP production, and all of these showed the inhibitory effect characterized in the present study. All four AC1-expressing clones that were tested showed stimulation by Ca2+; the cell line showing the highest cAMP response to Ca2+ was used in further experiments.
Assays of cAMP production. Incubations for the measurement of cAMP production were performed in cell suspensions as previously described (Paterson et al., 1995), with modifications as detailed below.
HEK-293 cells display complex intracellular Ca2+handling mechanisms that may be controlled by cAMP and calcineurin (Querfurth and Selkoe, 1994; Lin et al., 1995; Seuwen and Boddeke, 1995). Therefore, intracellular stores of Ca2+ were depleted to reduce the contribution of intracellular Ca2+ pools to the intracellular free Ca2+ signal. All procedures were performed at 37°C; where required, the immunosuppressant drugs FK506 (courtesy of Fujisawa GmbH, Munich, Germany), cyclosporin A (courtesy of Novartis, UK), and L685,818 (courtesy of Merck, Rahway, NJ) or ethanol vehicle (0.1% v/v) were added to the medium at the onset of the Ca2+ depletion period. Two different Ca2+ depletion protocols were used.
In the first protocol the cells were incubated in a balanced salt solution containing (in mm) 133 NaCl, 5.4 KCl, 0.25 Na2HPO4, 0.44 KH2PO4, 1 MgSO4, 2 EGTA, 5.6 d-glucose, 25 HEPES, pH 7.40, and 0.1% (w/v) bovine serum albumin (HBSS/EGTA) with 10 μm ryanodine and 1 μm thapsigargin (both from LC Labs, Boston, MA). These conditions deplete rapidly exchangeable Ca2+ pools and activate capacitative Ca2+ entry mechanisms (Cooper et al., 1994).
In the second protocol 4-Br-A23187 (5–15 μm) was used instead of ryanodine and thapsigargin and Ca/EGTA was added to obtain initial levels of [Ca2+]i similar to those in the studies with ryanodine and thapsigargin. Under these conditions intracellular Ca2+ pools are depleted and, in addition to capacitative Ca2+ entry, Ca2+ influx may take place through the pores formed by the ionophore.
The cells were incubated in Ca2+-depleting medium for 20 min. Subsequently, HBSS/EGTA containing various amounts of CaCl2 (Ca/EGTA) was added to achieve extracellular concentrations of Ca2+ ranging between 0.125 and 2 mm. The pH of this solution was 7.65 to minimize the change of pH caused by the displacement of protons from EGTA—a pH shift from 7.40 to 7.29 occurred on achieving a free Ca2+concentration of 2 mm. This protocol was modified when the effects of divalent cations were compared with those of Ca2+, because Ba2+ and Sr2+ potently displace Ca2+ from EGTA. Hence, cells were incubated for 20 min under Ca2+-depleting conditions and subsequently were pelleted by centrifugation at 200 × g for 5 min. The cells were resuspended in HBSS containing no added Ca2+, 0.2 mm EGTA, 10 μmryanodine, and 1 μm thapsigargin. The additions of Ca2+ and other divalent cations were made assuming that 0.2 mm EGTA was present.
After Ca/EGTA was added, the cells were incubated for 5 min, after which the cyclic nucleotide phosphodiesterase (PDE) inhibitors isobutylmethylxanthine (IBMX) and rolipram, 1 and 0.1 mm, respectively, were introduced. Unless indicated otherwise, the cells were incubated for a further 10 min and then the incubation was terminated by the addition of 0.2 m HCl. The total cAMP content of the cells and the medium was determined after freeze-thawing and acetylation by radioimmunoassay (Antoni et al., 1995).
Membrane adenylyl cyclase assay. Crude membranes from HEK-293 cells were prepared as for immunodetection, except that 50 nm calyculin A also was included in the homogenization buffer to inhibit protein phosphatases. Membranes (6–10 μg of protein per tube) were incubated at 30°C in 50 mmTris-HCl buffer, pH 7.1, containing (in mm) 0.3 ATP, 3 MgCl2, 5 creatine phosphate, 1 EGTA, and 0.5 IBMX plus 0.5 mg/ml creatine phosphokinase, 0.25% (w/v) bovine serum albumin; the reaction was linear up to 20 min. cAMP content was measured by radioimmunoassay as above.
Estimation of intracellular free Ca2+.The measurement of intracellular free Ca2+concentration ([Ca2+]i) was performed in cells loaded with fura-2 AM (4 μm for 30 min at 37°C) in a Shimadzu model RF5000 spectrofluorometer. Approximately 3 × 106 cells/ml were loaded into a cuvette; the solution was stirred and thermostatically heated to 37°C. The values for maximum and minimum fluorescence were obtained by recording the fluorescence intensities of a solution designed to mimic the intracellular environment (in mm): 25 Na-HEPES, pH 7.0, 10 NaCl, 120 KCl, 10 d-glucose, and 1 MgSO4 at 340 and 380 nm excitation; the formula of Grynkiewicz et al. (Grynkiewicz et al., 1985) was applied. The KDof fura-2 for Ca2+ was taken as 224 nm; maximum emission was determined at 4.1 mmCaCl2, and minimum values were determined with the addition of 60 mm EGTA.
Data analysis. The production of cAMP is given as multiples of the cAMP content of those cells not treated with blockers of PDE; the range of these means was 35–80 fmol/well, depending on the cell line used and the passage number of the cells. Data were analyzed by one-way ANOVA, and Newman–Keuls test or Dunnett’s test was used for multiple comparisons as appropriate.
RESULTS
Heterologous expression of AC9 in HEK-293 cells
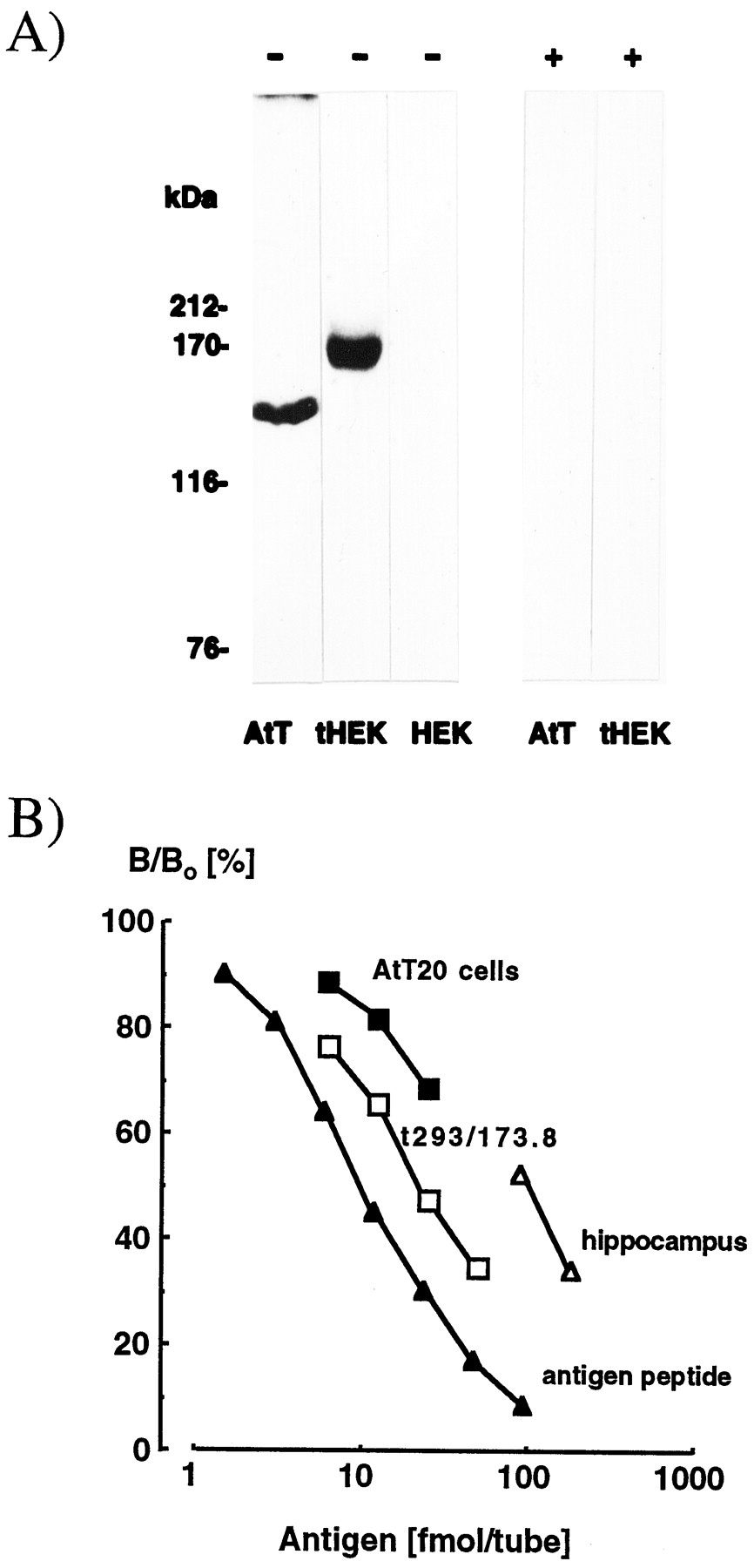
Immunoreactive AC9 was undetectable in membrane extracts prepared from wild-type or pcDNA3-transfected HEK-293 cells (Fig.1A). Cells stably transfected with AC9 cDNA contained a major ∼164 kDa band on immunoblots, whereas in extracts of AtT20 cells, where the cyclase is expressed endogenously (Antoni et al., 1995), an immunoreactive band migrating at ∼150 kDa was observed. The difference is plausibly attributable to cell-specific post-translational modifications such as glycosylation (Cali et al., 1994; Premont et al., 1996), phosphorylation (Kawabe et al., 1994; Antoni et al., 1998), or acylation (Mollner et al., 1995). Radioimmunoassay suggested that the stably transfected AC9 cell line expresses ∼30-fold higher levels of AC9 protein than AtT20 cells (10.1 ± 0.9 vs 0.30 ± 0.02 pmol/mg protein; means ± SEM; n = 3) (Fig.1B).

Detection of AC9 protein by immunoblot and radioimmunoassay. A, Crude membrane fractions of AtT20 D16:16 mouse corticotrope tumor cells (AtT; 0.36 μg of protein/lane), wild-type HEK-293 cells (HEK; 0.36 μg of protein/lane), and HEK-293 cells stably transfected with AC9 protein (tHEK; 0.08 μg of protein/lane) were separated by SDS-PAGE on 7.5% homogenous microgels. Immunoblots were prepared and reacted (lanes marked with −) with rabbit antiserum 149 at 1:4000 directed against the N-terminal region of mouse AC9 protein or the same antiserum preabsorbed with the antigenic pentadecapeptide (1 μg/ml; lanes marked with +). Positions of molecular weight markers are shown on the left. B, Crude membrane fractions prepared from AtT20 D16:16 cells (filled squares), HEK-293 cells stably transfected with AC9 (open squares), and rat hippocampus (open triangles) were solubilized in RIPA buffer and analyzed by radioimmunoassay with a chicken antiserum directed against the C-terminal region of mouse AC9 and the antigen peptide (filled triangles) as a reference standard. The tracer was the radiolabeled antigen peptide; sample extracts were serially diluted twofold.
Selective monitoring of AC1- and AC9-derived cAMP
In wild-type or pcDNA3-transfected HEK-293 cells the PDE inhibitors caused no significant (p > 0.05 by one-way ANOVA) change of cAMP levels in the absence of receptor agonists (Fig. 2A), whereas forskolin and corticotropin-releasing factor markedly stimulated cAMP accumulation in these cells (Antoni et al., 1995,1998).

Effect of Ca2+ on cAMP production by HEK-293 cells stably transfected with adenylyl cyclase cDNAs. A, Time course of cAMP accumulation in cells transfected with AC9 (open triangles) or pcDNA3 vector alone (filled triangles). Cells were incubated under Ca2+-depleting conditions (2 mmEGTA and 5 μm 4Br-A23187). Blockers of phosphodiesterase (1 mm IBMX and 0.1 mm rolipram) were applied at time 0. Data are expressed as multiples of the amounts of cAMP found in cells not receiving PDE blockers at time0. Means ± SEM; n = 4/group.B, Concentration-dependent inhibition of cAMP production by Ca2+ in cells stably transfected with AC9. HEK-293 cells overexpressing AC9 were incubated in medium containing no added calcium, 2 mm EGTA, 10 μm ryanodine, and 1 μm thapsigargin before the addition of various amounts of Ca/EGTA to the extracellular fluid. Theabscissa shows the amount of free Ca2+ in the extracellular medium. Data are expressed as multiples of the amount of cAMP found in cells not receiving PDE blockers. Means ± SEM; n = 4/group.C, Concentration-dependent simulation of cAMP production by Ca2+ in cells stably transfected with AC1. Conditions are as in B except that HEK-293 cells overexpressing AC1 were used.
In contrast, large increases of cAMP were elicited by PDE inhibitors in cells transfected with AC9 (Fig. 2A) or AC1 (data not shown). The addition of forskolin or corticotropin-releasing factor with blockers of PDE produced further increases above the levels observed in the presence of PDE blockers alone. However, these were only partially attributable to the transfected enzymes once stimulated cAMP synthesis by the endogenous adenylyl cyclase system of the host cell line was taken into account (data not shown) (Antoni et al., 1998).
The high cAMP-synthesizing activity of cells expressing AC9 was not attributable to the “stripping” of Gsα from heterotrimeric Gs. In membranes prepared from AC9-transfected cells, basal adenylyl cyclase activity was 532 ± 15 pmol/mg protein per 20 min, whereas in pcDNA3-transfected cells it was 26 ± 1 pmol/mg protein per 20 min (n = 4/group). Importantly, the application of GDP-β-S failed to alter basal adenylyl cyclase activity in AC9 transfected cells, whereas it blocked the stimulatory effect of GTP-γ-S (Table1).
Effect of GDP-β-S (100 μm) on membrane adenylyl cyclase activity of HEK-293 cells stably transfected with mouse AC9 in pcDNA3
In subsequent experiments PDE inhibitors alone were added to intact cells to elicit cAMP production, thus minimizing the contribution of the endogenous adenylyl cyclase system of the host cell line to cAMP biosynthesis. The addition of extracellular Ca2+inhibited or stimulated cAMP production in cells transfected with AC9 (Fig. 2B) or AC1 (Fig. 2C), respectively. The inhibitory effect of Ca2+ on AC9 in this paradigm appeared specific because 2 mmSrCl2, BaCl2, or MgCl2 had no significant influence on cAMP accumulation (Table 2).
Effect of divalent cations on cAMP production in HEK-293 cells stably transfected with AC9
Effect of Ca2+ on AC9 is mediated by calcineurin
The inhibitory effect of Ca2+ on AC9-derived cAMP formation in EGTA/ryanodine/thapsigargin-treated cells (Fig.3A) was blocked by FK506, an inhibitor of calcineurin (Schreiber, 1992). L685,818, an analog of FK506 that does not affect calcineurin activity (Dumont et al., 1992), was without effect at 50 μm (Fig. 3B). Cyclosporin A, another blocker of calcineurin (Schreiber, 1992), also inhibited the effect of Ca2+ on cAMP formation (Fig.3B). The pattern of [Ca2+]iin AC9-transfected cells treated with EGTA/ryanodine/thapsigargin is shown in Figure 3C; identical results were obtained with AC1-transfected cells (data not shown). Overall, in AC9-transfected cells 10 μm FK506 had no significant effect on the initial levels of [Ca2+]i (vehicle, 49.6 ± 8.5 nm vs FK506, 52 ± 6.5 nm; means ± SEM; n = 6) or the plateau levels achieved in the presence of 0.5 mm (94 ± 16 vs 85 ± 6; n = 3) or 2 mmextracellular Ca2+ (124.4 ± 15 vs 119 ± 13 in FK506; means ± SEM; n = 6).

Effect of calcineurin blockers on the inhibitory effect of Ca2+ in HEK-293 cells overexpressing AC9. The cells were incubated in medium containing no added calcium, 2 mm EGTA, 10 μm ryanodine, and 1 μm thapsigargin before the addition of various amounts of Ca/EGTA to the extracellular fluid to produce the free Ca2+ concentrations indicated on theabscissa. A, Concentration-dependent reversal of the Ca2+ inhibitory effect on cAMP production by FK506. cAMP accumulation was evoked by the addition of 1 mm IBMX and 0.1 mm rolipram. Data are expressed as multiples of the amount of cAMP found in cells not receiving PDE. Means ± SEM; n = 4/group. B, Specificity of immunosuppressant action: 2 μm FK506 and 2 μm cyclosporin A (CsA) blocked the effect of 0.5 mm extracellular free Ca2+ on cAMP production, whereas L685,818 (50 μm) had no effect. Conditions are as in A. Open columns, 0 Ca2+ medium; striped columns, 0.5 mm Ca2+. *p < 0.05 when compared with respective 0 Ca2+ medium control; one-way ANOVA, followed by Newman–Keuls test.C, Measurement of [Ca2+]i in a suspension of HEK-293 cells transfected with AC9 and loaded with fura-2 AM. Thelines indicate the start of the application of Ca/EGTA to the medium, and the numbers above thelines indicate the final concentrations (in mm) of free extracellular Ca2+ that are present; PDE-I indicates the application of 1 mm IBMX and 0.1 mm rolipram. The data are representative of six similar experiments.
In ionophore-treated cells, Ca2+ was also inhibitory to cAMP production by AC9 (Fig.4A), whereas AC1 was stimulated robustly (data not shown). FK506 and cyclosporin A blocked the inhibitory effect of 0.25 mm CaCl2 in the presence of 4-BrA23187, whereas L685,818 was without effect (Fig.4B). The pattern of [Ca2+]i under these conditions is shown in Figure 4C; note the lack of an effect of FK506.

Analysis of the effect of Ca2+and calcineurin blockers in ionophore-treated HEK-293 cells stably transfected with AC9. A, Cells were incubated in medium containing no added calcium, 2 mm EGTA, and 5 μm 4Br-A23187 before the addition of various amounts of Ca/EGTA to the extracellular fluid to produce the free Ca2+ concentrations that are indicated on theabscissa. Calcineurin inhibitors were applied 20 min before Ca/EGTA. cAMP accumulation was evoked by the addition of 1 mm IBMX and 0.1 mm rolipram; data are expressed as multiples of the amount of cAMP found in cells not receiving PDE blockers. Means ± SEM; n = 4/group. *p < 0.05 when compared with the respective 0 Ca2+ group. $p < 0.05 when compared with the respective groups receiving vehicle; one-way ANOVA, followed by Newman–Keuls test. B, Specificity of FK506 (10 μm) effect when compared with vehicle and L685,818 (50 μm). Open columns, 0 Ca2+ medium; striped columns, 0.5 mm Ca2+. Conditions are as in A. *p < 0.05 when compared with the respective 0 Ca2+ group; one-way ANOVA, followed by Newman–Keuls test. C, Measurement of [Ca2+]i in HEK-293 cells transfected with AC9, loaded with fura 2-AM, and pretreated with vehicle (filled symbols) or 10 μm FK506 (open symbols). The top line indicates the start of the application of Ca/EGTA yielding 0.25 mmfree extracellular Ca2+ into the medium; theline below PDE-I indicates the application of 1 mm IBMX and 0.1 mm rolipram. The data are representative of six similar experiments.
Higher average levels of [Ca2+]i(200–300 nm) were achieved by using 15 μm4-BrA23187 and 0.25 mm CaCl2. However, under these conditions the addition of PDE blockers to the cells dramatically enhanced [Ca2+]i further to ≥1 μm. An almost complete suppression of cAMP synthesis by AC9 resulted that was not altered by FK506 or cyclosporin A (data not shown). In parallel experiments the stimulatory effect of Ca2+ on AC1 was observed no longer, suggesting the nonspecific toxicity of this ionophore concentration in the presence of Ca2+.
Distribution of AC9 mRNA in the forebrain
A summary of the overall distribution pattern of AC9 mRNA in mouse forebrain is presented in Table3. The distribution of AC9 mRNA was identical in rat brain. The mRNA signal was restricted mainly to the gray matter and appeared to be associated with neuronal perikarya.
Relative abundance of AC9 mRNA in areas of the mouse brain
Olfactory lobe
A large number of neuronal perikarya in the anterior olfactory nucleus, the nucleus of the lateral olfactory tract, and the olfactory bulb were strongly positive for AC9 mRNA.
Limbic lobe and other limbic areas
The most intense mRNA signal was apparent in the limbic areas of the forebrain. In particular, neurons of the hippocampal complex, including the tenia tecta, the anterior hippocampus, indusium griseum, and hippocampus, were labeled very intensely for AC9 mRNA. In the hippocampus (Fig. 5A–E) the CA1–CA3 pyramidal cell layers and the dentate gyrus granule cells showed dense and specific labeling. Examination at higher magnification (Fig. 5E) showed that close to 70% of the perikarya in the pyramidal layer of CA3 and at least 50% of the granule cells in the dentate gyrus expressed AC9 mRNA. Strong specific hybridization signal also was detected in neurons of the subiculum (Fig. 5F,G) and the parasubiculum.

Demonstration of AC9 mRNA in the hippocampus and the subiculum in coronal sections of mouse brain, using a35S-labeled antisense riboprobe (except for B, G). A, Dark-field view of the anterior dorsal hippocampus. d, Dentate gyrus. Scale bar, 400 μm.B, Specificity control with 35S-labeled sense probe in coronal section of dorsal hippocampus; otherwise, the conditions are as in A. C, Dark-field view of the dorsal hippocampus at a more posterior level. Note the intense labeling of pyramidal neurons in the CA1 andCA3 regions as well as granule cells in the dentate gyrus (d). Scale bar, 100 μm. D, Dark-field view of the ventral hippocampus showing intense labeling of neuronal perikarya in the pyramidal layers ofCA1–CA3 as well as the granule cells of the dentate gyrus (d). s, Subiculum. Scale bar, 200 μm. E, Bright-field view of dorsal hippocampus at higher magnification in coronal sections counterstained with pyronin to reveal cell nuclei. Scale bar, 50 μm.F, Bright-field view of the subiculum; intense labeling is evident in neurons. V, Dorsal third ventricle. Scale bar, 100 μm. G, Specificity control forF, using 35S-labeled sense riboprobe.
Neurons in the piriform, the entorhinal, and the cingulate cortices were also highly positive for AC9 mRNA. Labeled cell bodies in the cingulate (Fig. 6A,B) and the entorhinal cortices (data not shown) were most prominent in layers II and III.

Distribution of AC9 mRNA in coronal sections of the cingulate cortex (A, C) and the parietal cortex (B, D) of the mouse brain. A, Cingulate cortex, dark-field view. Scale bar, 100 μm. Arrowheadpoints to layer II–III shown in C as a bright-field image at higher magnification, counterstained with pyronin. Scale bar, 50 μm. B, Parietal cortex, dark-field view. Scale bar, 200 μm. D, Bright-field view of layers II–III, counterstained with pyronin. Scale bar, 50 μm.
Other limbic areas showing distinct low-to-moderate levels of mRNA signal include the dorsal septal nucleus, the lateral, medial, and posterior amygdaloid nuclei, the medial habenular nucleus, and the interpeduncular nucleus.
Neocortex
In the neocortex AC9 mRNA was observed mainly in neurons of layers II, III, and VI and the large pyramidal cells of layer V (Fig.6C,D). There were no apparent topographical variations in this pattern.
Striatonigral system
The caudate putamen contained a large number of cells expressing moderate levels of AC9 (Fig.7A,B) mRNA, whereas the globus pallidus was essentially devoid of specific signal.

Neurons positive for AC9 mRNA in the caudate nucleus (A, B). Scale bars:A, 100 μm; B, 25 μm.
Diencephalon
The anteroventral thalamic nucleus showed moderate mRNA signal; a lower level of signal was found in the paratenial and the centromedian thalamic nuclei. Both the supraoptic and the paraventricular nuclei contained low-to-moderate levels of mRNA. Low-level labeling also was detected in the median preoptic nucleus.
Cerebellar system
In the cerebellar cortex Purkinje cells were weakly positive for AC9 mRNA (Fig. 8). Other areas of the cerebellar system expressing AC9 mRNA included the cerebellar nuclei, the inferior olive, the pontine nuclei, and the red nucleus.

AC9 mRNA labeling in coronal section of the cerebellum. Arrowheads, Purkinje cells;ic, interposed cerebellar nucleus; lc, lateral cerebellar nucleus. Scale bar, 200 μm.
Lower brainstem
Some reticular, serotonergic, and motor cells in the lower brainstem expressed moderate levels of AC9.
Detection of AC9 protein in brain
Immunoblot analysis of detergent extracts of membrane fractions from the hippocampus and hypothalamus showed the presence of a major ∼155 kDa immunoreactive band (Fig. 9). Quantification by radioimmunoassay with a C-terminally directed antiserum suggested that crude membrane fractions prepared from the hippocampus have AC9 levels similar to those seen in AtT20 cells (Table4). The cerebellum, the hypothalamus, and the anterior pituitary gland expressed lower levels of AC9 protein, which is consistent with the lower abundance of the mRNA signal in these tissues (Paterson et al., 1995; present study).

Immunoblots of rat hippocampal (HIP; 2.5 μg protein/lane) and hypothalamic (HYP; 7.5 μg/lane) membranes reacted with anti-AC9 (−lanes) or the same antiserum preincubated with 1 μg/ml antigen peptide (+ lanes). Proteins were separated by SDS-PAGE on 7.5% homogenous microgels. The positions of molecular weight markers are indicated on the left of the graph.
The concentration of immunoreactive adenylyl cyclase 9 in selected regions of the rat brain and the anterior pituitary gland
DISCUSSION
These data show that the Ca2+/calcineurin-inhibited adenylyl cyclase (AC9) is present in areas of the brain previously thought to contain only Ca2+-activated (AC1 and AC8) or protein kinase C-activated (AC2 and AC7) adenylyl cyclases. The distribution of AC9 mRNA in the brain suggests close proximity to or possible colocalization in AC1- and AC2-expressing neurons in limbic and neocortical areas. Functionally, comparative analysis of AC1- and AC9-derived cAMP production in HEK-293 cells showed that AC1 and AC9 are regulated by the same range of intracellular free Ca2+ concentrations. Taken together, these data indicate that AC9 is a major cAMP-synthesizing enzyme in brain and is modulated by Ca2+ by way of the protein phosphatase calcineurin.
Properties of AC9
Previous work from our laboratory demonstrated calcineurin-mediated inhibition of agonist-induced cAMP responses in AtT20 cells that express relatively high levels of endogenous AC9 (Antoni et al., 1995) as well as HEK-293 cells transiently transfected with AC9 cDNA (Paterson et al., 1995). In these studies cAMP production was evoked by receptor agonists, and thus it remained to be demonstrated that the target of calcineurin-dependent control is AC9 itself rather than the efficiency of receptor–Gs–cyclase coupling. Meanwhile, others have reported that mouse AC9 is not influenced by Ca2+/calmodulin (Premont et al., 1996) and that a human AC9 variant recently cloned from the heart is not regulated by Ca2+-dependent processes (Hacker et al., 1998). The plausible explanations for these discrepancies are (1)Premont et al. (1996) used Sf9 insect cell membranes for which the profile of phosphorylation of AC9 may be different from that in mammalian systems, and (2) Hacker and coworkers (1998) reported an enzyme that has no significant homology in the C1b regulatory domain (see below) with mouse AC9 and hence may have different regulatory properties.
All mammalian adenylyl cyclases known so far conform to the “quasi-duplicated transporter” design (Taussig and Gilman, 1995) consisting of two membrane-spanning domains each, followed by substantial cytoplasmic loops that have been designated as C1 and C2, respectively. Current evidence indicates that the physical interaction of C1 and C2 underlies the catalytic activity of the enzyme. The interaction between C1 and C2 is facilitated by forskolin or Gsα–GTP complexes (Tang and Gilman, 1995; Tesmer et al., 1997) that enhance the affinity of the interaction between C1 and C2 by ∼50-fold (Whisnant et al., 1996). Therefore, overexpression of adenylyl cyclase in the region of 30-fold above normal, as in the present study, should produce substantial “basal” adenylyl cyclase activity, because the number of catalytically active C1–C2 complexes is increased by a similar factor as in the presence of forskolin or Gsα–GTP. The adaptation of stably transfected cells to high levels of cAMP via increased gene expression and phosphorylation-dependent activation of PDEs is well documented (Conti et al., 1995; Houslay, 1998). Hence unstimulated levels of cAMP in cells stably transfected with adenylyl cyclase are similar to those in wild-type cells, whereas the pharmacological inhibition of PDE activity unmasks the enhanced production of cAMP by the transfected enzyme (Cali et al., 1994; Wayman et al., 1994; present study). With the use of GDP-β-S we have provided additional evidence that the increased adenylyl cyclase activity in the stably transfected cells is independent of Gsα. The analysis of membrane adenylyl cyclase activity also confirmed previous reports (Premont et al., 1996;Antoni et al., 1998; Yan et al., 1998) that AC9 is stimulated only weakly by forskolin. Taken together, the characteristics of cAMP production in HEK-293 cells stably transfected with AC9 and treated with PDE blockers reflect the regulatory properties of the adenylyl cyclase catalytic moiety.
The present experiments show that intracellular free Ca2+ inhibits cAMP synthesis by AC9. The inhibitory effect of Ca2+ on AC9 was blocked by FK506 and cyclosporin A, but not by L685,818, which conforms with the current knowledge on the pharmacology of calcineurin (Schreiber, 1992; Sigal and Dumont, 1992). The degree of inhibition of AC9 by Ca2+/calcineurin was modest, i.e., 20–40%, which could be attributable to a number of factors, including the high levels of adenylyl cyclase relative to calcineurin expressed in the cells or the inactivation of calcineurin by [Ca2+]i (Wang et al., 1996). Furthermore, depending on the rate of cAMP hydrolysis of the system under study, a relatively small reduction (∼30%) of the rate of cAMP synthesis may lead to a marked fall (∼80%) of intracellular cAMP levels (Chiono et al., 1995).
The concentration of intracellular Ca2+ sufficient for the inhibition of AC9 was in the region of 100 nm, a point at which calcineurin is already active (Perrino et al., 1995;Fruman et al., 1996). Further, this suggests that the fluctuations of [Ca2+]i found under “basal” conditions are sufficient to modulate AC9 activity, i.e., constitutive phosphorylation/dephosphorylation may influence the level to which the enzyme may be activated. Indeed, in AtT20 cells, which display spontaneous [Ca2+]i spikes with amplitudes up to 400 nm (Antoni et al., 1992), the inhibition of calcineurin with FK506 and cyclosporin A leads to enhanced activation of membrane adenylyl cyclase by corticotropin-releasing factor (Antoni et al., 1998).
Whether or not AC9 is a direct target for calcineurin or is regulated by a protein phosphatase downstream of calcineurin (Antaraki et al., 1997), i.e., a protein phosphatase cascade (Cohen, 1989), remains to be established. Moreover, the protein kinases that phosphorylate AC9 have not been identified.
Distribution of AC9 in the forebrain
The AC9 mRNA signal was prominent in neurons found in higher brain regions where substantial levels of mRNA for other Ca2+-inhibited adenylyl cyclases such as AC3, AC5, or AC6 have not been reported. Extraneuronal mRNA for AC9 was not observed. Overall, in the hippocampus, the neocortex, the striatum, and the cerebellum the same arrays of neurons that express significant levels of AC9 mRNA also contain relatively high levels of calcineurin (Kuno et al., 1992; Goto et al., 1993; Dawson et al., 1994) and FKBP12 (Dawson et al., 1994; Sabatini and Snyder, 1995). Within hippocampal pyramidal neurons (Sı́k et al., 1998) calcineurin is found in dendritic spines and the cell body, where adenylyl cyclase immunoreactivity also has been demonstrated with an antibody that recognizes all mammalian adenylyl cyclases (Mons et al., 1995). Thus, on the basis of current knowledge, the respective topographical distributions of AC9, calcineurin, and FKBP12 are consistent with the hypothesis that calcineurin is a regulator of AC9.
A comparison of the patterns of expression of AC9 and other adenylyl cyclases clearly indicates the close proximity of neurons expressing AC9 to those expressing the Ca2+-stimulated cyclases AC1/AC8 as well as AC2, an adenylyl cyclase stimulated by protein kinase C. Indeed, the distribution of AC9 mRNA in the hippocampal formation is identical to that of AC1, except in the pyramidal layer of CA1–CA3 where AC1 mRNA is relatively low (Xia et al., 1991), whereas AC9 mRNA is highly abundant. Thus mRNAs for three classes of adenylyl cyclase are expressed in the pyramidal cells of CA1–CA3 and in the granule cells of the dentate gyrus. Whether or not this is also the case at the level of protein expression and to what extent these cyclases are colocalized in the same neurons remain to be clarified.
The present study provides the first data on AC9 protein concentration in brain regions, which suggest that the level of AC9 in rat hippocampus is comparable to that in the clonal AtT20 cell line. In turn, AtT20 cells were found to express the expected level, i.e., ∼0.05% of total membrane protein of immunoreactive AC9. Given the selective neuronal localization of AC9 mRNA, the concentration of AC9 protein in the hippocampus appears remarkably high. Similar data on other cyclase isotypes are not available at present.
Implications for synaptic function
Currently, any considerations of the physiological function of AC9 are speculative; however, two areas stand out as plausible and biologically significant.
First, in AtT20 cells AC9 is the target of an intracellular Ca2+ feedback loop that also involves BK-type K+ channels and Ca2+/calmodulin-activated PDE. This system regulates the membrane potential and the levels of cAMP to maintain cellular excitability (Antoni, 1996). AtT20 cells show the rhythmic firing of action potentials and intracellular free Ca2+transients (Suprenant, 1982; Adler et al., 1983; Antoni et al., 1992). Prominent expression of AC9, Ca2+-activated K+ channels, and Ca2+/calmodulin PDE, i.e., the components of Ca2+ negative feedback on cAMP levels and the membrane potential, is evident in hippocampal principal neurons, which also exhibit various modes of rhythmic firing. Thus AC9 may be involved in the generation of rhythmic electrical activity in the hippocampus and possibly in other parts of the forebrain.
Second, hippocampal neurons have been analyzed extensively with respect to the role of cAMP in synaptic plasticity. The activation of cAMP synthesis by [Ca2+]i has been attributed as a key role in various types of long-term potentiation (LTP) (for review, see Xia and Storm, 1997). However, the example of the Drosophila dunce mutant, which is deficient in a cyclic nucleotide PDE gene, indicates that the hydrolysis of cAMP is also a key process in memory formation and underscores the issue of a cAMP signal that is optimized in time as well as space (Dudai, 1997).
Calcineurin is a potent inhibitor of postsynaptic activity in hippocampal neurons (Raman et al., 1996; Wang and Kelly, 1997) and recently has been proposed as a “memory suppressor” gene (Abel et al., 1998). The overexpression of calcineurin in hippocampal neurons impaired a cAMP-dependent phase of tetanus-evoked LTP (Winder et al., 1998) as well as hippocampus-dependent memory formation (Mansuy et al., 1998). The prominent expression of AC9 in the hippocampus and the pivotal role of the tuning of the cAMP Ca2+ signal in synaptic plasticity (Blitzer et al., 1995) indicate that AC9 may be an important physiological target for protein kinase and phosphatase cascades that contribute to the long-term modulation of synaptic transmission.
Footnotes
We thank J. Bennie, O. Grace, and S. Carroll for technical support. A generous supply of cAMP antiserum donated by K. J. Catt and A. Baukal (National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD) is gratefully acknowledged. We also thank D. R. Storm and W. L. Zhu (Seattle, WA) for kindly providing the plasmids used in this study and Peter Sharp (Roslin, Scotland, UK) for help in raising antisera in hens.
Correspondence should be addressed to Dr. F. Antoni, Medial Research Council Brain Metabolism Unit, Department of Neuroscience, University of Edinburgh, Edinburgh, EH8 9JZ, Scotland, UK.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}