


Abstract
The effects of several N-sulfonyl-polyamines, including N1-dansyl-spermine (N1-DnsSpm) andN 1-(n-octanesulfonyl)-spermine (N1-OsSpm), were studied at recombinantN-methyl-d-aspartate (NMDA) receptors expressed in Xenopus laevis oocytes. N1-DnsSpm and N1-OsSpm inhibited NMDA receptors and were ∼1000-fold more potent than spermine in oocytes voltage-clamped at −70 mV. Block by N1-DnsSpm and N1-OsSpm was strongly voltage dependent, being more pronounced at hyperpolarized membrane potentials. With the Woodhull model of voltage-dependent channel block, the values of K d(0) were 779 μm, 882 μm, and 7.4 mm and those of zδ were 2.58, 2.57, and 1.07 for N1-DnsSpm, N1-OsSpm, and spermine, respectively. This suggests that an increase in the voltage dependence of block together with an increase in affinity contributes to the increased potencies of N1-DnsSpm and N1-OsSpm compared with spermine. Sensitivity to N1-DnsSpm was reduced by mutation NR1(N616Q) and was increased by mutations NR1(N616G) and NR2A(N615G). The NR1(N616G) and NR2A(N615G) mutations decreased the K d(0) value of N1-DnsSpm without affecting zδ, whereas the NR1(N616Q) mutation reduced zδ. These mutations may alter the accessibility of part of the polyamine binding site within the channel pore or directly alter the properties of that site. Block by N1-DnsSpm (0.3 μm) was almost complete at −100 mV, and there was no relief of block at extreme negative membrane potentials (−100 to −200 mV) at wild-type NR1/NR2A channels. In contrast, block by N1-DnsSpm was partially relieved at extreme negative potentials at receptors containing NR1(N616G) or NR2A(N615G), suggesting that N1-DnsSpm can permeate these mutant channels but not wild-type NR1/NR2A channels. This is hypothesized to be due to an increase in the pore size of channels containing NR1(N616G) or NR2A(N615G), which allows passage of the bulky head group of N1-DnsSpm. In contrast to N1-DnsSpm, N1-OsSpm could easily permeate wild-type NR1/NR2A channels, presumably because the head group of N1-OsSpm can pass through the narrowest part of the channel pore.N-Sulfonyl-polyamines such as N1-DnsSpm and N1-OsSpm represent a new class of polyamine antagonists with which to study glutamate receptor ion channels.
The endogenous polyamine spermine has a variety of effects on NMDA and non-NMDA glutamate receptors (1,2). At NMDA receptors, spermine has both stimulatory and inhibitory effects when applied extracellularly (3-7). Inhibition of NMDA receptors by spermine is strongly voltage dependent and may be caused by an open-channel block and/or screening of surface charges around the mouth or vestibule of the ion channel (3, 5, 8). Intracellular spermine can block the ion channel of some subtypes of AMPA and kainate receptors, an effect that is responsible for inward rectification of these receptors (9-12) and may be mechanistically similar to the block of inward-rectifier K+ channels by polyamines (13,14).
When applied extracellularly, spermine is a relatively weak antagonist at NMDA receptors and at polyamine-sensitive AMPA and kainate receptors, blocking these receptors at high micromolar to millimolar concentrations (3, 4, 10, 15). A number of polyamine-conjugated spider and wasp toxins are more potent antagonists than spermine at glutamate receptors (16). These toxins, which include the philanthotoxins, argiotoxins, and α-agatoxins, are characterized structurally by the presence of an aromatic amino acid head group linked through a carbonamide bond to a polyamine tail such as spermine or a pentamine or hexamine (17-21). Because of their potencies and specificities, polyamine-conjugated toxins are potentially valuable tools for studying the pharmacological and structural properties of glutamate receptor ion channels and as tools to discriminate subtypes of native glutamate receptors. However, it is often difficult to obtain these toxins because they have to be purified from spider venom, requiring access to the appropriate spiders, or to be synthesized in the laboratory. The syntheses of argiotoxins and α-agatoxins, which are potent NMDA channel blockers, are far from straightforward (22), and commercially available toxins are often prohibitively expensive.
To look for polyamine derivatives that have activities similar to those of the polyamine-conjugated spider toxins, we have studied the properties of several N1-substituted polyamines. Derivatives of spermine and spermidine, such as N1-DnsSpm and N1-OsSpm (Fig. 1), that have an alkyl- or aryl-sulfonyl group attached to one of the terminal amino groups, were recently found to be potent inhibitors of calmodulin-activated phosphodiesterase activity and of polyamine uptake.1 These compounds are stable and are relatively easy to synthesize. In the current study, N1-DnsSpm and N1-OsSpm were found to be potent voltage-dependent blockers that could differentially block and/or permeate recombinant NMDA receptors. N1-Sulfonyl-polyamines are useful new tools to study polyamine block of glutamate receptors and channel structure of those receptors.

Structures of some of the compounds used in this study.
Experimental Procedures
Expression in oocytes and voltage-clamp recording.
The preparation of cRNAs and the preparation, injection, and maintenance of oocytes were carried out as described previously (7, 23, 24). Oocytes were injected with NR1 plus NR2 cRNAs in a ratio of 1:5 (0.5–4 ng of NR1 plus 2.5–20 ng of NR2). For experiments with NR2C and NR2D, the mouse cDNA clones ε3 and ε4 were used (25, 26).
Macroscopic currents were recorded with a two-electrode voltage-clamp using a GeneClamp 500 amplifier (Axon Instruments, Foster City, CA) or an OC-725 amplifier (Warner Instruments, Hamden, CT) as previously described (7, 23). Electrodes were filled with 3 m KCl and had resistances of 0.4–3 MΩ. Oocytes were continuously superfused (∼5 ml/min) with a Mg2+-free saline solution (96 mm NaCl, 2 mm KCl, 1.8 mmBaCl2, 10 mm HEPES, pH 7.5), which contained BaCl2 rather than CaCl2 to minimize Ca2+-activated Cl− currents (27).In most experiments, oocytes were injected with K+-BAPTA (50–100 nl of 40 mm, pH 7.4) on the day of recording to eliminate a slowly activating Cl− current that is seen even in the presence of extracellular Ba2+ (23).
I-V curves were measured by using linear voltage ramps over 6–24 sec as described in Results. In some experiments, control ramps with glutamate were measured before and after ramps with polyamines and the control ramps were averaged. In other experiments, control ramps were measured before but not after ramps with polyamines. In all experiments, leak currents were measured with ramps before and after the test ramps, and leak currents were digitally subtracted.
Site-directed mutagenesis.
The NR1 mutants were prepared by using a 2.6-kb SphI/SalI fragment of plasmid pN60 (28) inserted into the same sites of M13mp18 (29). Similarly, the NR2 mutants were prepared using a 2.2-kb BamHI/XmaI fragment of pBSNR2A and a 2.1-kb BamHI/SphI fragment of PBSNR2B inserted into the same sites of M13mp18 and M13mp19, respectively. Mutagenesis was carried out according to the method of Kunkel et al. (30) or Sayers et al.(31) with the Sculptor in vitro mutagenesis system (Amersham International, Buckinghamshire, UK). The oligonucleotides for preparation of mutants were CAA TGC CGG AGC CGA GCA GGA CGC (antisense) for NR1(N616G), GCC TGG TCT TCCAGA ATT CTG TGC C (sense) for NR2A(N614Q), TGG TCT TCA ACC AGT CTG TGC CTG T (sense) for NR2A(N615Q), TGG TCT TCA ACG GTT CTG TGC CTG (sense) for NR2A(N615G), GTC TGG TGT TTC AGA ACT CCG TAC C (sense) for NR2B(N615Q), and TGG TGT TTA ACCAGT CCG TAC CTG T (sense) for NR2B(N616Q) (mutated nucleotides are underlined). Mutated DNA fragments were isolated from the replicative form of M13 and religated into the corresponding sites of pN60, pBSNR2A, and pBSNR2B. Mutations were confirmed by DNA sequencing (32). The NR1(N616Q) mutant (33) was provided by Dr. S. Nakanishi (Institute for Immunology, Kyoto University Faculty of Medicine, Kyoto, Japan). The NR1(N616R) mutant (34) was provided by Dr. R. J. Dingledine (Department of Pharmacology, Emory University, Atlanta, GA). Amino acids are numbered from the initiator methionine in NR1 and NR2 clones (28, 35). This numbering system differs from the system used in some laboratories in which amino acids are numbered from the first residue in the mature peptide (36). Thus, residues NR1(N616) and NR2A(N615) correspond to NR1(N598) and NR2A(N596), respectively, in the article by Wollmuth et al. (36).
Data analysis.
Data analysis and curve fitting were carried out using Axograph (Axon Instruments) or SigmaPlot (Jandel Scientific, San Rafael, CA) on Macintosh computers. To obtain values for the IC50 and Hill slope (nH
) of antagonists, concentration-inhibition curves were fit to eq.1:
For analysis of the voltage dependence of block by polyamines, data were analyzed using a model based on that of Woodhull (37) by fitting the data to eq. 2:
Materials.
The syntheses of N1-DnsSpm, N1-OsSpm, N1-DnsSpd, and N8-DnsSpd (hydrochloride salts) are reported elsewhere.1 l-Glutamate and glycine were purchased from Sigma Chemical (St. Louis, MO). Spermine tetrahydrochloride was purchased from Aldrich Chemical (Milwaukee, WI) or Calbiochem (San Diego, CA). The NR1 clone (28) was a gift from Dr. S. Nakanishi (Institute for Immunology, Kyoto University, Kyoto, Japan). The splice variant of NR1 used in these studies was NR1A (28, 38). The NR2A and NR2B clones (39) were gifts from Dr. P.H. Seeburg (Center for Molecular Biology, University of Heidelberg, Germany). The ε3 and ε4 (mouse NR2C and NR2D) clones (25, 26) were gifts from Dr. M. Mishina (Department of Pharmacology, University of Tokyo, Japan).
Results
Potencies of polyamine derivatives.
Polyamines and theirN-dansylated derivatives inhibited responses to glutamate and glycine at NR1/NR2A receptors. The potencies of spermine, spermidine, and their derivatives were measured in oocytes voltage-clamped at −70 mV (Fig. 2 and Table1). N1-DnsSpm was 1700-fold more potent than the parent compound spermine (Fig. 2 and Table 1). Similarly,N-dansylation at either terminal amino group of spermidine produced compounds, N1-DnsSpd and N8-DnsSpd, that were 200–400-fold more potent than spermidine (Table 1). To determine whether the dansyl moiety itself was a potent NMDA receptor antagonist, we measured the effects of dansylamide and of dansylethylamide at NR1/NR2A receptors in oocytes voltage-clamped at −70 mV. These two compounds, at a concentration of 10 μm, inhibited responses to glutamate by only 5–9% (data not shown). Thus, the potent inhibitory effects of dansylated polyamines do not lie within the dansyl group itself and require the polyamine tail in addition to the dansyl head group. Another derivative of spermine, N1-OsSpm (Fig. 1), which has an N1-alkyl rather than an N1-aryl substitution, had a potency similar to that of N1-DnsSpm (Table 1). The subunit-specificity of N1-DnsSpm was determined by measuring its potency at NR1/NR2 receptors containing different NR2 subunits. N1-DnsSpm was ∼50-fold more potent at NR1/NR2A and NR1/NR2B receptors than at NR1/NR2C and NR1/NR2D receptors (Table 1). The block of NR1/NR2A receptors by N1-DnsSpm (0.3 μm) was noncompetitive with respect to glutamate and glycine, with the degree of block being unaffected by concentrations of glutamate and glycine over a range of 0.3–10 μm (data not shown).

Effects of N1-DnsSpm and spermine at NR1/NR2A receptors. The inhibitory effects of various concentrations of N1-DnsSpm and spermine on stimulation by glutamate (10 μm; with 10 μm glycine) were measured in oocytes expressing NR1/NR2A receptors and voltage-clamped at −70 mV. Values are mean ± standard error from six oocytes for each compound and are expressed as a percentage of the control response to glutamate. Inset, representative trace showing inhibition by 0.3 μm N1-DnsSpm.
Effects of polyamine analogs at NMDA receptors
Block of NMDA receptors by N1-DnsSpm is strongly voltage dependent (see below), similar to block by spermine. In addition to voltage-dependent block, spermine has three other macroscopic effects at native and recombinant NMDA receptors, all of which are subunit dependent (3, 4, 6, 7, 40). These effects are “glycine-dependent” stimulation, which involves an increase in the affinity for glycine and is seen with sub-saturating concentrations of glycine; “glycine-independent” stimulation, which is seen in the presence of saturating concentrations of glycine; and a decrease in agonist affinity, which is mechanistically related to the glycine-independent form of stimulation. All of the effects of spermine are seen at NR1/NR2B receptors (containing the NR1A splice variant of NR1), but only voltage-dependent block and glycine-dependent stimulation are seen at NR1/NR2A receptors (6, 7). To determine whether N1-DnsSpm shows either form of polyamine stimulation, we measured the effects of 0.3 μm N1-DnsSpm at NR1/NR2B receptors in oocytes voltage-clamped at −20 mV to minimize voltage-dependent block. No stimulation by N1-DnsSpm was seen when experiments were carried out with high (10 μm) or low (0.1 μm) concentrations of glycine (data not shown). This suggests that polyamine stimulation does not occur with concentrations of N1-DnsSpm that produce a profound voltage-dependent block and that, if N1-DnsSpm does have stimulatory effects at NMDA receptors, the potency of those effects is not increased to the same extent as the potency of the voltage-dependent block. Thus, although it remains possible that N1-DnsSpm can have stimulatory effects on NMDA receptors, such effects are not seen when studying macroscopic currents using low micromolar concentrations of N1-DnsSpm. These observations are consistent with the hypothesis that the stimulatory effects of spermine and related polyamines involve binding sites that are outside the ion channel pore, distinct from the site that mediates voltage-dependent block (1, 3, 6). Experiments were also carried out to look for stimulation by high (10–100 μm) concentrations of N1-DnsSpm at NR1/NR2B receptors (data not shown), but those concentrations produced a large block of glutamate responses even at depolarized membrane potentials, which may mask stimulatory effects of N1-DnsSpm.
Voltage dependence of block and effects of mutations in the M2 regions of NR1 and NR2.
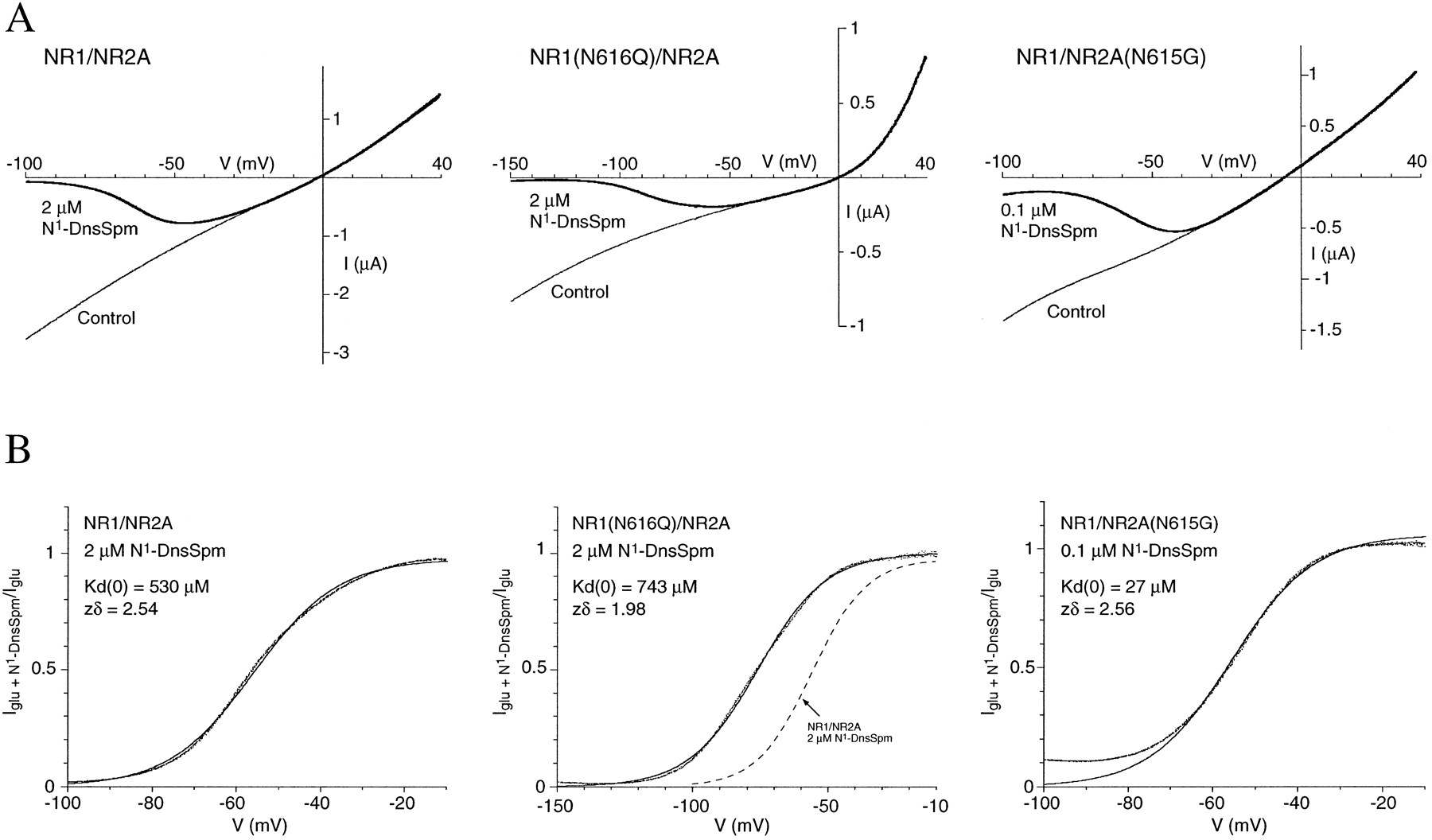
The inhibitory effects of spermine and of polyamine-derived spider toxins are voltage dependent. Therefore, experiments were carried out to study the voltage dependence of block by N1-sulfonyl-polyamines. In some experiments, we measured steady state currents induced by glutamate or glutamate plus N1-DnsSpm in oocytes voltage-clamped at different holding potentials. Inhibition by N1-DnsSpm was strongly voltage dependent, being more pronounced at hyperpolarized than at depolarized membrane potentials (data not shown). The voltage dependence of block by N1-DnsSpm and N1-OsSpm was studied quantitatively by using voltage ramps analyzed according to the model of Woodhull (37) (Fig. 3 and Tables 2 and3). At wild-type NR1/NR2A receptors, the values ofKd (0) were similar for N1-DnsSpm (779 μm) and N1-OsSpm (882 μm), as were the values for zδ (2.58 for N1-DnsSpm and 2.57 for N1-OsSpm; Tables 2 and 3). In these two polyamine derivatives, the sulfonamide nitrogen is a weak acid and is not charged at pH 7.5, and the dimethylamino group on the dansyl moiety of N1-DnsSpm is also uncharged at this pH. Thus, N1-DnsSpm and N1-OsSpm each have a total charge of +3, due to protonation of the amino groups in the polyamine tail. Assuming that all three amino groups enter the transmembrane electric field (i.e., z = 3), the value of δ (the average depth of the transmembrane field sensed by the polyamines) is 0.86. We also studied block by spermine (300 μm) at NR1/NR2A receptors by using voltage ramps. The value of Kd (0) was 7.4 ± 3.5 mm and that of zδ was 1.07 ± 0.17 (mean ± standard error, 5 oocytes) for block by spermine. These values are similar to those reported for block by spermine at native NMDA receptors on cultured hippocampal neurons, at which theKd (0) value was 27 mm and the zδ value was 1.17 (3).

Voltage-dependent effects of N1-DnsSpm at wild-type and mutant NR1/NR2A receptors. A, I-V curves were measured by voltage ramps (−100 to +40 mV or −150 to +40 mV over 6 sec) in oocytes expressing wild-type or mutant NMDA receptors activated by glutamate (10 μm; with 10 μm glycine) in the absence (control) or presence of N1-DnsSpm (2 μm or 0.1 μm). Leak currents have been subtracted. B, Currents from A, measured in the presence of N1-DnsSpm, are expressed as a fraction of the control current in each oocyte. Solid lines, fits to the Woodhull model (text eq. 2), which, in these cells, was fit from −10 to −80 mV [NR1/NR2A], −10 to −100 mV [NR1(N616Q)/NR2A], and −10 to −70 mV [NR1/NR2A(N615G)] and extrapolated to −100 or −150 mV. The values of K d(0) and zδ for N1-DnsSpm that were derived for each oocyte are shown in the corresponding panels. Broken line in the middle (B) [NR1(N616Q)/NR2A], fitted curve for block by N1-DnsSpm at wild-type NR1/NR2A channels is replotted from the left(B) [NR1/NR2A].
Voltage-dependent block of NR1/NR2A receptors by N1-DnsSpm
Voltage-dependent block of NR1/NR2A receptors by N1-OsSpm
Block and permeation of NMDA receptor channels by inorganic divalent cations and by synthetic channel blockers such as MK-801 and TCP are controlled by an asparagine residue (N616) in the NR1 subunit (33, 34,41, 42). This residue is in a position analogous to the Q/R site that controls Ca2+ permeability of AMPA and kainate receptors (43). In AMPA and kainate receptors, the residue at the Q/R site also controls sensitivity to block by intracellular and extracellular polyamines (9, 15). The NR2 subunits contain two asparagine residues in the M2 loop that correspond to N616 and S617 of NR1. In NR2A, these are residues N614 and N615, and in NR2B, they are N615 and N616 (Fig. 4A). In NR1/NR2A receptors, NR1(N616) and NR2A(N615) (i.e., the second asparagine residue in NR2A) have a major influence on ion permeability, and these residues seem to be directly in the ion permeation pathway, where they form the narrowest part of the ion channel pore (36). However, the critical asparagine residues in M2 of NR1 and NR2 subunits seem to provide nonsymmetrical contributions to channel structure and cation permeability (36, 41). To determine whether the asparagine residues in the M2 regions of NR1 and NR2 influence sensitivity to N1-DnsSpm, we studied the effects of mutations at NR1(N616), NR2A(N614), NR2A(N615), NR2B(N615), and NR2B(N616).

Block of mutant NR1/NR2 receptors by N1-DnsSpm. A, Schematic showing the positions of the M1-M4 segments of NMDA receptor subunits. NR1 and NR2 subunits have amino- and carboxy-terminal domains of different sizes flanking the M1-M4 region. The amino acid sequences in the M2 regions (solid line above sequences) of NR1, NR2A, and NR2B are shown below the schematic. Amino acids are numbered from the initiator methionine in each subunit according to Moriyoshi et al. (28) and Ishii et al. (35). Bold and numbered, positions of the asparagine residues where mutations were studied. B, Effects of N1-DnsSpm on stimulation by glutamate (10 μm; with 10 μm glycine) were measured in oocytes expressing wild-type and mutant NR1/NR2A receptors and voltage-clamped at −70 mV. Values are mean ± standard error from four to nine oocytes for each subunit combination and are expressed as a percentage of the control response to glutamate.
Mutation of NR1(N616) to glutamine (N616Q) or arginine (N616R) reduced the potency of N1-DnsSpm by 15- 200-fold when the mutants were coexpressed with wild-type NR2A or NR2B (Fig. 4B and Table4), whereas mutation of this residue to glycine, NR1(N616G), increased the potency of N1-DnsSpm by ∼40-fold (Table 4). Mutations NR2A(N614Q) and NR2B(N615Q) produced a small increase in sensitivity to N1-DnsSpm, whereas N-to-Q mutations at the second asparagine in NR2A or NR2B, NR2A(N615Q) and NR2B(N616Q), produced small decreases in sensitivity to N1-DnsSpm when these mutants were expressed with wild-type NR1 (Table 4). This suggests that the second asparagine residue in the M2 region of NR2A and NR2B has a role similar to NR1(N616). Furthermore, mutation of NR2A(N615) to glycine, NR2A(N615G), increased sensitivity to N1-DnsSpm by 20–40-fold when NR2A(N615G) was coexpressed with wild-type NR1 or NR1(N616Q) (Fig. 4B and Table 4).
Properties of mutant NMDA receptors
Results of experiments using voltage ramps analyzed according to the Woodhull model suggest that the increased potency of N1-DnsSpm seen with the NR1(N616G) and NR2A(N615G) mutations is due to an increase in the affinity of the binding site for N1-DnsSpm because the values ofKd (0) are significantly lower in these mutants than in the wild-type NR1/NR2A receptor, whereas the values of zδ are unchanged (Fig. 3 and Table 2). In contrast, the NR1(N616Q) mutation had only a small effect on theKd (0) of N1-DnsSpm, but this mutation significantly reduced the value of zδ for N1-DnsSpm (Fig. 3 and Table 2).
We also studied the effects of some of the mutations in M2 on block of NR1/NR2A receptors by N1-OsSpm. IC50 values for N1-OsSpm were derived from concentration-inhibition curves using oocytes voltage-clamped at −70 mV. The potency of N1-OsSpm at NR1/NR2A receptors at −70 mV was reduced by mutation NR1(N616Q) and increased by mutation NR2A(N615G) (Table 3), similar to the effects of these mutations on the potency of N1-DnsSpm. However, mutation NR1(N616G), which increases the potency of N1-DnsSpm, had no effect on the potency of N1-OsSpm (Table 3). The voltage dependence of block by N1-OsSpm was studied at wild-type and mutant receptors by using voltage ramps and Woodhull modeling. Because N1-OsSpm showed marked permeation of wild-type and mutant channels (see below), we used concentrations of N1-OsSpm that were 5–10-fold higher than their IC50 values measured at −70 mV. Mutation NR1(N616Q) did not alter the Kd (0) value of block by N1-OsSpm but significantly reduced the value of zδ (Table 3). In contrast, mutation NR2A(N615G) reduced the Kd (0) of N1-OsSpm without affecting the zδ (Table 3). These results are similar to the effects of NR1(N616Q) and NR2A(N615G) on the zδ and Kd (0) values of block by N1-DnsSpm (Table 2).
An aspartate residue (D669) in the extracellular loop of NR1, just distal to the M3 segment, has been found to influence stimulation by spermine and inhibition by protons (44). This residue also has a small influence on voltage-dependent block by spermine. Mutations that neutralize the negative charge at D669 (D669N and D669A) reduced voltage-dependent block by spermine, whereas a mutation that retains the negative charge (D669E) did not alter voltage-dependent block by spermine, although all three mutations decrease sensitivity to pH and to spermine stimulation (44). It was proposed that screening of the negative charge at D669 of NMDA receptors may contribute to voltage-dependent block by spermine (44). It is notable that in the linear amino acid sequence D669 is in a position analogous to D652 of the GluR1 subunit of AMPA receptors, a position also occupied by an aspartate residue in all other GluR subunits. In a study that generated three-dimensional models of this region of GluR1 and GluR6 subunits, GluR1(D652) (equivalent to D669 in NR1) was one of two acidic residues that, after agonist binding, was proposed to move to a position near the mouth of the ion channel (45). This residue could be important for attracting cations to the channel (45). If D669 in NR1 occupies a position in the tertiary structure of NR1 analogous to that of D652 in GluR1 (45), then NR1(D669) could be positioned near the entrance of the ion channel of NMDA receptors. In this case, the inhibitory effect of spermine could be due in part to an interaction of an amino group of spermine with NR1(D669), masking the negative charge at D669, and thus reducing the attraction of Na+ and Ba2+ and, consequently, the unitary conductance of the channel. To determine whether D669 influences block by N1-DnsSpm, we measured the effects of 1 μm N1-DnsSpm on responses to glutamate (10 μm; with 10 μm glycine) at NR1/NR2A receptors containing NR1(D669E) and NR1(D669N) in oocytes voltage-clamped at −70 mV. N1-DnsSpm inhibited responses to glutamate by 77 ± 5% (wild-type), 80 ± 2% (D669E), and 58 ± 2% (D669N; p < 0.01, one-way analysis of variance with Dunnett’s test). Thus, mutation D669N but not D669E reduces block by N1-DnsSpm, similar to effects seen with spermine (44), although the effect of the NR1(D669N) mutation is much smaller than the effects of mutations at NR1(N616). This suggests that charge screening, at least of residue D669, contributes only in small part to block by N1-DnsSpm. It may be that N1-DnsSpm has more points of interaction with the channel pore than does spermine, and thus mutations at one contact point (D669) have a smaller effect on block by N1-DnsSpm than on block by spermine.
Permeation through wild-type and mutant channels.
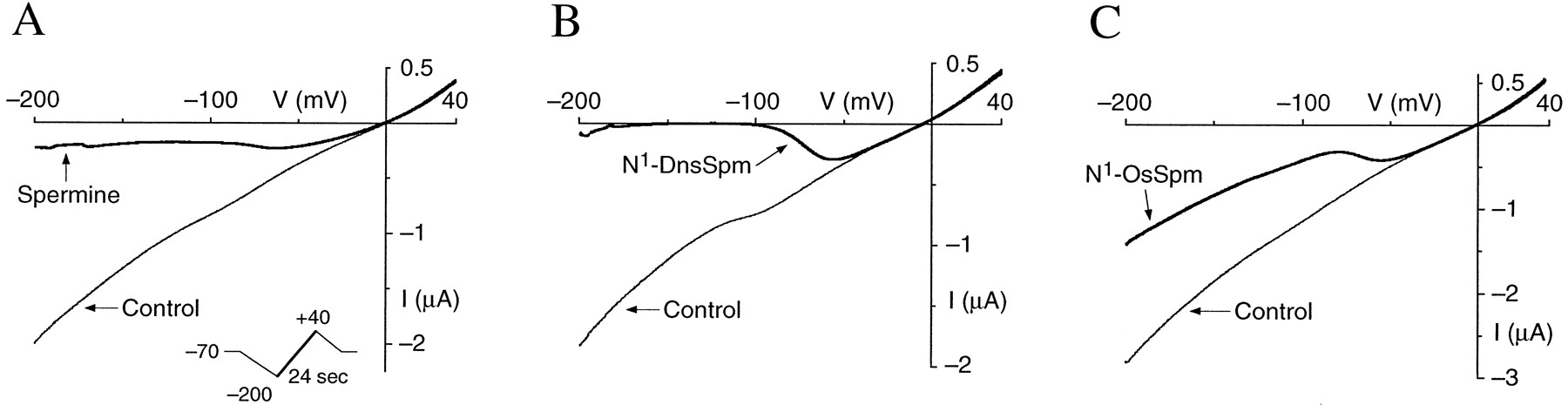
A number of different characteristics have been reported for the voltage-dependent block of macroscopic NMDA currents by spermine and polyamine analogs. Spermine produces an incomplete block of NMDA receptors even at extremely hyperpolarized potentials of −100 to −200 mV (3, 46). The shallow slope conductance and incomplete block seen with spermine (Fig.5A) (3, 46) may reflect permeation of spermine through the ion channel of NMDA receptors (3) or screening of surface charges rather than fast channel block (8). Some polyamine analogs, such as 1,10-diaminodecane, seem to act as classic channel blockers, producing a complete block of macroscopic currents (47, 48). Other analogs, in particular, long-chain penta-amines such as BE4444, cause a complete block of NMDA responses, but the block is relieved by ≥50% at extreme negative membrane potentials, presumably reflecting permeation of BE4444 through the ion channel of NMDA receptors (46). To determine whether the N1-sulfonyl-polyamines can permeate NMDA channels, we compared the effects of spermine with those of N1-DnsSpm and N1-OsSpm at extreme negative potentials (Fig. 5) by using concentrations of spermine (300 μm), N1-DnsSpm (0.3 μm), and N1-OsSpm (0.3 μm) that are close to the IC50 values for these antagonists at −70 mV (see Table 1). N1-DnsSpm produced a complete block of NR1/NR2A receptors at membrane potentials of ∼−100 mV, and little or no recovery of the response was seen at extreme negative potentials (Fig. 5B). In contrast, block by N1-OsSpm was incomplete and showed a partial recovery at extreme negative membrane potentials (Fig. 5C). These results suggest that N1-OsSpm, but not N1-DnsSpm, can easily permeate the ion channel of NR1/NR2A receptors.

I-V relationships at NR1/NR2A receptors. I-V curves were measured by voltage ramps (−200 to +40 mV, 24 sec; see A,inset) during steady state responses induced by glutamate (10 μm; with 10 μmglycine) in the absence and presence of (A) 300 μmspermine, (B) 0.3 μm N1-DnsSpm, and (C) 0.3 μm N1-OsSpm. Leak currents have been subtracted. Similar results were obtained with 3–12 oocytes for each compound.
Mutation of the asparagine residues at NR1(N616) and NR2A(N615) to glycine residues has been shown to increase the permeability of organic cations such as tetramethylammoniun, leading to estimates of the effective pore size in wild-type and mutant NMDA receptors (36). The size of the narrowest constriction in wild-type NR1/NR2A receptors was estimated to be ∼0.55 nm, and this was increased to 0.67 nm in NR1/NR2A(N615G) receptors, 0.75 nm in NR1(N616G)/NR2A receptors, and 0.87 nm in NR1(N616G)/NR2A(N615G) receptors (36, 49, 50). Using a space-filling model, we estimated the diameter of the naphthalene ring on N1-DnsSpm to be 0.8–0.85 nm and hypothesized that this may prevent permeation of N1-DnsSpm through wild-type NR1/NR2A receptors but that N1-DnsSpm may permeate NR1/NR2A receptors containing NR1(N616G) and NR2A(N615G) mutants. To test this hypothesis, we measured the I-V profile for block by N1-DnsSpm at receptors containing these mutants. We also studied N-to-Q (rather than N-to-G) mutants at these positions (Fig.6). Even though the N-to-Q mutation at NR1(N616Q) increases the size of the side chain at this position, the mutation has been reported to increase rather than decrease the size of the narrowest part of the ion channel pore, possibly because the bulkier side chain of the glutamine (Q) mutant does not pack well in the channel pore and disrupts channel structure (36).

Permeation of N1-DnsSpm through mutant NR1/NR2 receptors. A, I-V curves were constructed by voltage ramps (−185 mV to +40 mV; 24 sec) at wild-type and mutant NR1/NR2A receptors activated by glutamate (10 μm; with 10 μm glycine) in the absence and presence of N1-DnsSpm. Leak currents have been subtracted. B, The fractional block of the glutamate response by N1-DnsSpm was measured at −80 and at −170 mV from data obtained with voltage ramps similar to those shown in A. Values are mean ± standard error. The concentrations of N1-DnsSpm that were used in A and B and the number of oocytes for each subunit combination in B were 0.3 μm N1-DnsSpm for NR1/NR2A (20 oocytes), 0.01 μm for NR1(N616G)/NR2A (7 oocytes), 6 μm N1-DnsSpm for NR1(N616Q)/NR2A (7 oocytes), 0.03 μm N1-DnsSpm for NR1/NR2A(N615G) (7 oocytes), 0.01 μmN1-DnsSpm for NR1(N616Q)/NR2A(N615G) (12 oocytes), and 0.3 μm N1-DnsSpm for NR1(N616Q)/NR2A(N615Q) (11 oocytes).
To assess permeation of N1-DnsSpm at wild-type and mutant NR1/NR2A receptors, we measured I-V relationships in the presence and absence of N1-DnsSpm using concentrations of N1-DnsSpm that were close to their IC50 values measured at −70 mV (Table 4). These concentrations of N1-DnsSpm produced a 60–80% block of glutamate responses at −80 mV in the wild-type and mutant receptors (Fig. 6B). I-V curves were constructed by using linear voltage ramps from −200 or −185 mV to +40 mV at a rate of 9–10 mV/sec (Fig. 6A). To measure recovery from block (which is assumed to reflect permeation of N1-DnsSpm), the ratio of the glutamate-induced current in the absence and presence of N1-DnsSpm was measured at −80 and −170 mV (Fig. 6B). This ratio will be larger at −170 mV than at −80 mV if there is recovery from block at extreme negative membrane potentials. At wild-type receptors, no recovery was seen at −170 mV. However, a pronounced recovery was seen at NR1(N616G)/NR2A, NR1/NR2A(N615G), and NR1(N616Q)/NR2A(N615G) receptors (Fig. 6). These data suggest that N1-DnsSpm, which does not easily permeate wild-type NR1/NR2A receptors, can readily permeate receptors containing NR1(N616G) or NR2A(N615G). We also studied permeation at channels containing NR1(N616Q) together with wild-type NR2A and at channels containing NR2A(N615Q) together with NR1(N616Q) (Fig. 6); permeation by N1-DnsSpm was not seen at these mutants. Thus, the increase in permeation of N1-DnsSpm is selective for the NR1(N616G) and NR2A(N615G) mutations.
Discussion
One of the goals of this study was to identify polyamine derivatives that share the potencies of the polyamine-conjugated toxins such as the argiotoxins and philanthotoxins but are easier to synthesize than those toxins. The sulfonamide polyamine derivatives described here represent a new class of polyamine antagonists of glutamate receptors. Because these compounds are stable and their syntheses are straightforward, N1-DnsSpm, N1-OsSpm, and related analogs should be useful tools for studies of glutamate receptor ion channels. The N1-substituted polyamines that we have studied were 200-1700-fold more potent than the parent compounds, with N1-DnsSpm and N1-OsSpm being the most potent. Indeed, N1-DnsSpm (IC50 = 0.3 μmat −70 mV) was only ∼30-fold less potent than argiotoxin636 and Agel-505 (IC50 = 0.01 μm at −70 mV) (18, 19). The potency of N1-DnsSpm is similar to that reported for philanthotoxin-343 (IC50 = 10–56 μm at −60 mV) (20, 21). Although N1-DnsSpm and N1-OsSpm were ∼1000-fold more potent than spermine at −70 mV, the values ofKd (0) determined for these compounds (800–900 μm) were only ∼10-fold lower than the Kd (0) value for spermine (7.4 mm). However, N1-DnsSpm and N1-OsSpm showed a much steeper voltage dependence (zδ = 2.58) than did spermine (zδ = 1.07), suggesting that an increase in voltage dependence, together with a modest increase in the affinity of binding, is responsible for the much greater potency, at −70 mV, of the N1-sulfonyl polyamines compared with spermine.
The subunit selectivity of N1-DnsSpm was similar to that reported for a number of structurally diverse channel blockers acting at NMDA receptors. Thus, N1-DnsSpm was less potent at NR1/NR2C and NR1/NR2D receptors than at NR1/NR2A and NR1/NR2B receptors, similar to the profile seen with Mg2+, MK-801 (25, 35, 39), spermine (6, 51), and argiotoxin636 (19). The structural features of NR2 subunits that control sensitivity to many of these antagonists are unknown, although regions of NR2B and NR2C that influence Mg2+ block have been described (52). Spermine itself blocks NR1/NR2A and NR1/NR2B receptors at micromolar concentrations but has no effect on NR1/NR2C and NR1/NR2D receptors at concentrations of ≤300 μm (6, 51). Because of its increased potency compared with spermine, N1-DnsSpm may be a useful probe for studying the subunit-specific properties that influence sensitivity to polyamines.
At wild-type NR1/NR2A channels, Woodhull (37) analysis of the block yielded very similar values of zδ (∼2.58) for N1-DnsSpm and N1-OsSpm. In this analysis, z represents the valence of the blocker, and δ represents the depth of the binding site for the blocker within the membrane field. Thus, if the charge on the polyamine analogs is +3 at pH 7.5, the value of δ is 0.86, suggesting that the “binding site” for polyamines lies deep within the channel. Indeed, the zδ values for the N1-sulfonyl polyamines (zδ = 2.57–2.58) were larger than the value for spermine seen at NR1/NR2 receptors (zδ = 1.07) or at native NMDA receptors (zδ = 1.17) (3), suggesting that N1-DnsSpm and N1-OsSpm may bind much deeper in the channel pore than spermine itself, even though spermine has four positively charged amino groups at physiologic pH. However, there are a number of limitations and caveats to using the Woodhull model to analyze block by polyamines. First, it is not known whether only one molecule of N1-DnsSpm (or any of the other polyamines) enters and binds in the channel at a given time or whether two or more molecules can bind simultaneously. In the case of inward rectifier K+ channels, for example, it has been proposed that two molecules of spermine can simultaneously enter and block the channel in an end-to-end manner (53). Second, it is not known whether the entire polyamine tail of N1-DnsSpm and N1-OsSpm enters the pore of the NMDA channel, or if only part of the polyamine tail, containing, for example, one or two charged amino groups, enters the membrane electric field. However, if only one molecule of N1-DnsSpm enters the channel, most or all or the polyamine tail would have to be within the membrane electric field to yield a zδ value of 2.58, assuming that z = 3. A third limitation to analyzing the voltage-dependence of block is that N1-OsSpm shows marked permeation of wild-type channels and N1-DnsSpm permeates some of the mutant NMDA channels. Although we used concentrations of the polyamines that produced a large block and analyzed data over a voltage range where the block develops, it is not known to what extent permeation of a polyamine molecule influences the block caused by another molecule subsequently entering the channel. It is conceivable that in channels where permeation occurs, two polyamines could be present simultaneously: one blocking at the polyamine binding site or approaching that site and another unblocking and passing through the channel into the oocyte.
Polyamines have several stable conformations. In the fully extended all-trans conformation of N1-DnsSpm and N1-OsSpm, the distances between the amino groups are ∼0.52 nm (diaminopropane moieties) and ∼0.65 nm (diaminobutane moiety), with a total distance of ∼1.2 nm between the first and third charged amino groups. These positively charged groups may each interact with one or more amino acid side chains within the channel pore, constituting part of the polyamine binding site. Again, this places limitations on the interpretation of the depth-of-field (δ) value derived from the apparent valence, zδ. If the polyamine derivatives enter the membrane field in an extended conformation, the separation between the first and third charged amino groups could span 20% of the membrane electrical field, assuming a lipid bilayer thickness of 6 nm.
Mutations at the critical asparagine residues in the M2 regions of NR1 and NR2A had complex effects on the potency (at −70 mV), theKd (0) value, and the zδ value of block by N1-DnsSpm and N1-OsSpm. Mutations NR1(N616G) and NR2A(N615G) increased the potency of N1-DnsSpm, an effect that was probably due to an increase in the affinity of the binding site for N1-DnsSpm because these mutations reduced the Kd (0) value of N1-DnsSpm without affecting zδ. A possible explanation for these results is that the asparagine residues present at position NR1(N616) and NR2A(N615) normally hinder the binding of N1-DnsSpm and that when these residues are replaced by glycine residues, in NR1(N616G) and NR2A(N615G), the N1-DnsSpm has increased access to the interaction points of its binding site. Mutation of NR1(N616) to glutamine (Q), which has a bulkier side chain than asparagine, had a different effect on block by N1-DnsSpm, reducing the potency and the zδ but having only a small effect on the Kd (0) value of N1-DnsSpm. The NR1(N616Q) mutation may reduce the interaction of one of the charged groups on N1-DnsSpm with the channel protein or may reduce the depth to which N1-DnsSpm can enter into the channel. Indeed, the NR1(N616Q) mutation had a similar effect on block by N1-OsSpm, reducing the potency and the zδ of that compound, suggesting that the mutation affects the interaction of the polyamine tail rather than the head group in these polyamine derivatives. As was seen with N1-DnsSpm and N1-OsSpm, block by MK-801 and TCP is influenced by mutations at NR1(N616) (33, 34, 42). Thus, NR1(N616) may contribute directly to the binding sites for N1-DnsSpm and MK-801. Alternatively, mutations at this site could alter block by perturbing other features of the channel structure or by alterations in pore size.
The NR2A(N615G) mutation decreased theKd (0) of N1-OsSpm, similar to its effect on N1-DnsSpm, whereas NR1(N616G), which increases the affinity for N1-DnsSpm, had no effect on block by N1-OsSpm. The NR1(N616G) mutation may have a predominantly volume-specific effect, allowing easier access of the bulky head group of N1-DnsSpm to the channel but having little or no effect on the accessibility and binding of N1-OsSpm. On the other hand, the NR2A(N616G) mutation affects the affinities of both N1-DnsSpm and N1-OsSpm, suggesting that this residue may alter the properties of another part of the polyamine binding site or may have effects on the accessibility of this site that are similar for N1-DnsSpm and N1-OsSpm. These results also indicate a nonequivalent or nonsymmetrical role for the asparagines at the narrowest part of the channel in NR1 and NR2A, as was seen in studies of the permeability of small organic cations (36).
Block by N1-OsSpm was incomplete and was partially relieved at extreme negative membrane potentials, whereas block by N1-DnsSpm was complete and showed no relief at extreme negative potentials. The relief of block by N1-OsSpm at −100 to −200 mV is similar to that seen with linear penta-amines such as BE4444 (46) and presumably reflects permeation of N1-OsSpm through the ion channel of NMDA receptors. Thus, if N1-OsSpm can permeate the ion channel, presumably in a linear conformation, but N1-DnsSpm cannot, it is likely that the bulky aromatic head group of N1-DnsSpm (see Fig.1) is responsible for the impermeant properties of this molecule. In receptors containing the NR1(N616G) or NR2A(N615G) mutants, permeation of N1-DnsSpm was seen at extreme negative membrane potentials. Based on the permeability ratios of K+ and organic cations such as tetraethylammonium, the NR1(N616G) and NR2A(N615G) mutants have been estimated to increase the size of the narrowest region of the NMDA channel from 0.55 nm in NR1/NR2A receptors to 0.75 nm in NR1(N616G)/NR2A receptors and to 0.67 nm in NR1/NR2A(N615G) receptors (36). The results of the present work are consistent with the hypothesis that the bulky head-group of N1-DnsSpm normally prevents it from easily permeating the channel, but that increasing the pore size with the N-to-G mutations allows N1-DnsSpm to easily permeate NR1/NR2A channels. We estimated the diameter of the naphthalene ring on N1-DnsSpm to be 0.8–0.85 nm, which is too large to easily permeate wild-type channels with a pore size of 0.55 nm (36, 49, 50). Because N1-DnsSpm can easily permeate receptors with NR2A(N615G), the narrowest constriction in NR1/NR2A(N615G) channels may be > 0.67 nm (36). However, it is not known whether the size of the head group on N1-DnsSpm, rather than its chemical structure, is the key determinant of permeation through NMDA channels. Nevertheless, N1-DnsSpm and other sulfonamide polyamine derivatives represent new tools that can be used to study block and permeation of glutamate receptor channels. Studying reversal of block at extreme negative membrane potentials using modified polyamines such as N1-DnsSpm provides a means of probing channel pore structure using extracellular application of N1-DnsSpm and whole-cell recording, without the need to use the polyamine itself as the major charge carrier through the channels.
Acknowledgments
We are grateful to Drs. S. Nakanishi, P. H. Seeburg, and M. Mishina for providing the wild-type NR1 and NR2 clones; to Drs. S. Nakanishi and R. J. Dingledine (Emory University, Atlanta, GA), for providing some of the NR1 mutants; and to Albert Pahk for technical assistance with some experiments.
Footnotes
- Received August 27, 1996.
- Accepted January 17, 1997.
-
Send reprint requests to: Keith Williams, Ph.D., Department of Pharmacology, University of Pennsylvania School of Medicine, Philadelphia, PA 19104-6084.
-
↵1 N. Seiler, F. Douaud, J. Renault, J.-G. Delcros, R. Havouis, P. Uriac, and J.P. Moulinoux. Targeting of tumors via the polyamine uptake system: a model study with N1-dansyl-spermine. I. Effects on enzymes and on cells in culture. Submitted for publication.
-
This work was supported by United States Public Health Service Grant NS35047 from the National Institute of Neurological Disorders and Stroke, by a Grant-in-Aid from the American Heart Association, and by a grant from the Japan Health Sciences Foundation.
Abbreviations
- NMDA
- N-methyl-d-aspartate
- AMPA
- α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
- N1-DnsSpm
- N1-dansyl-spermine
- N1-OsSpm
- N1-(n-octanesulfonyl)-spermine
- N1-DnsSpd
- N1-dansyl-spermidine
- N8-DnsSpd
- N8-dansyl-spermidine
- HEPES
- 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
- TCP
- N-[1-(2-thienyl)cyclohexyl]piperidine
- I-V
- current-voltage
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}