


Abstract
Human estrogen receptors α (ERα) and β (ERβ) are ligand-inducible transcription factors that are highly homologous in their central DNA-binding and carboxyl-terminal ligand-binding domains. In contrast, there is very little conservation between ERα and ERβ in the amino-terminal domain. Using different human cell lines, we show that wild-type ERβ transcriptional activity is lower or similar to that of ERα, depending on the cell type. Deletion of the amino-terminal domain in both ER subtypes resulted in no or a lower decrease of transcriptional activity of ERβ compared with ERα, suggesting that the ERβ amino-terminal domain contains a weaker transcriptional activation function-1. Using ERα and ERβ deletion mutants, we showed that the amino-terminal transcriptional activity of ERβ maps to amino acids 1-31. Interestingly, this domain contains a six amino-acid motif (amino acids 5–10 in human ERβ) that is part of the ERα-activation function-1 region (amino acids 49–54 in human ERα) and highly conserved among all mammalian ERα amino-terminal domains. Despite this similarity between the two ER subtypes, no autonomous and ligand-independent activity of the ERβ-amino-terminal domain was observed in yeast and mammalian cells in contrast to ERα. This study provides a molecular basis for the difference in transcriptional activity between ERα and ERβ and establishes that ERβ contains a structurally and functionally restricted amino-terminal transcriptional activity.
Estrogen receptors (ERs) are ligand-inducible transcription factors that mediate the biological effects of estrogens and antiestrogens. Two ER subtypes, encoded by different genes have been isolated in mammals, ERα and ERβ (Greene et al., 1986; Kuiper et al., 1996; Tremblay et al., 1997). ERα and ERβ belong to the large superfamily of nuclear receptors that includes receptors for steroid and thyroid hormones, retinoic acid, vitamin D3, fatty acids, eicosanoids, oxysterols, and ecdysone, as well as orphan receptors that have no known ligand (Gronemeyer and Laudet, 1995; Mangelsdorf et al., 1995). Members of this superfamily share a similar modular structure, including a central DNA-binding domain (DBD), a carboxyl-terminal ligand-binding domain (LBD), and a variable amino-terminal domain. Like the other steroid hormone nuclear receptors, ERα and ERβ homodimerize on hormone binding, and interact with specific palindromic DNA sequences termed estrogen response elements (EREs) located in target genes (Beato et al., 1995). In addition ERα and ERβ can form heterodimers at ERE-binding sites (Cowley et al., 1997; Pace et al., 1997; Pettersson et al., 1997). Transcription is then stimulated by interaction of the DNA-bound receptor with coactivators and recruitment of a preinitiation complex (Xu et al., 1999).
Transcriptional activity of ERα is controlled by two different transcription activation functions (AFs) located in the amino-terminal region (AF-1) and in the carboxyl-terminal region of the LBD (AF-2), respectively (Tora et al., 1989b). The AF-1 transcription activation function is hormone-independent, whereas activity of AF-2 requires the presence of hormone (Tora et al., 1989b). Furthermore, both AF activities are dependent on the promoter context and cell type (Tora et al., 1989b; Berry et al., 1990; Pham et al., 1992; Tzukerman et al., 1994). The AF-1 and AF-2 activities have been shown to synergize in certain cell types and this is thought to occur via the interaction of the corresponding regions with coactivators such as SRC-1 (Tora et al., 1989b; McInerney et al., 1996). In ERα, the AF-1 region also has been shown to mediate the ligand-independent activation of the receptor on phosphorylation (Kato et al., 1995; Bunone et al., 1996). Mutational analysis of human ERα has shown that within the amino-terminal region itself, AF-1 activity maps to several different domains, depending on the cell type and the biocharacter of the ligand (Metzger et al., 1995; McInerney and Katzenellenbogen, 1996). For instance, deletion of the first 101 amino acids resulted in a more pronounced reduction of the AF-1 activity in chicken embryo fibroblasts than in cervix carcinoma HeLa cells and had a moderate effect in yeast cells (Metzger et al., 1995). It also was shown that deletion of the first 87 amino acids had no effect on 17β-estradiol-stimulated transcription in the breast cancer MDA-MB 231 cells, whereas the agonistic response to the antiestrogentrans-hydroxytamoxifen was completely abolished (McInerney and Katzenellenbogen, 1996).
ERβ exhibits a high degree of homology with ERα in the DBD region (96% amino acid identity) and in the LBD region (58% amino acid identity), including an identical AF-2 core domain (Kuiper et al., 1996). This structural homology correlates with the findings that both receptors have similar DNA-binding properties (Cowley et al., 1997;Pace et al., 1997; Pettersson et al., 1997), bind 17β-estradiol with similar affinity (Kuiper et al., 1997), and physically interact with the coactivator SRC-1 (Tremblay et al., 1997). In contrast, the amino-terminal domain of ERβ is shorter by about 80 amino acids and shows no sequence homology compared with that of ERα. However, this region is well conserved between the rat, mouse, and human ERβ, suggesting an evolutionary constraint and a functional importance. Variability of the amino-terminal region between subtypes or isoforms of receptors for the same hormone is common within the nuclear receptor superfamily and generally results in functional differences. For instance, progesterone receptor isoforms A and B diverge exclusively in this domain and activate transcription from target genes differentially and in a cell-specific manner (Tora et al., 1988; Kastner et al., 1990;Vegeto et al., 1993). Interestingly, it has been reported that the 17β-estradiol stimulated transcriptional response of ERα is higher than that of ERβ in some cell types (Cowley et al., 1997; Pettersson et al., 1997; McInerney et al., 1998). Furthermore, no partial agonistic activity of the antiestrogen 4-hydroxytamoxifen could be observed with ERβ in contrast to ERα (Cowley et al., 1997; Tremblay et al., 1997). Finally, ERα but not ERβ is able to activate transcription from a reporter gene controlled by an AP-1 site in response to 17β-estradiol or diethylstilbestrol (Peach et al., 1997). Collectively, these observations suggest that despite well conserved DBD and LBD, ERα and ERβ have different transcriptional activation properties that could result at least in part from structurally divergent amino-terminal domains.
In this study, we have compared the transcriptional activities of human ERα and ERβ by using different human cell lines and promoter configurations. With amino-terminal ERβ and ERα mutants we show that amino-terminal domains of both ER subtypes exhibit clear functional differences.
Materials and Methods
Reagents.
Estradiol-17β was purchased from Sigma (St. Louis, MO). Moxestrol was kindly provided by Stefan Nilsson (KaroBio AB, Huddinge, Sweden). Media, antibiotics, fetal bovine serum, Lipofectamin-Plus, and Lipofectin were purchased from Life Technologies, Inc. (Gaithersburg, MD). Vent DNA polymerase was purchased from New England Biolabs (Beverly, MA). The luciferase detection reagents were purchased from Bio-Orbit Oy (Turku, Finland).
Plasmids Constructs.
The luciferase reporter plasmids 2xERE-Tk-Luc and 3xERE-TATA-Luc have been described (McInerney et al., 1996; Pettersson et al., 1997; Kuiper et al., 1998). The expression plasmid pSG5-hERα also has been described (Tora et al., 1989a). The human ERα amino-terminal deletion mutants ERαΔ63 and ERαΔ142 were constructed by inserting linkers containing a start codon between the EcoRI andNotI (ERαΔ63) or FseI (ERαΔ142) sites of pSG5-hERα. The ERαΔ182 mutant was constructed by replacing theEcoRI-HindIII fragment of pSG5-hERα (coding for residues 1–340) with a polymerase chain reaction (PCR)-generated fragment coding for residues 183 to 340 and including a start codon immediately before amino acid 183. The expression plasmid pSG5-hERβ was constructed by inserting a SacII (blunted)-BamHI fragment containing the human ERβ cDNA (Enmark et al., 1997) into the pSG5 plasmid after digestion withBamHI (blunted) and BglII. The human ERβ amino-terminal deletion mutants ERβΔ31, ERβΔ46, ERβΔ67, and ERβΔ93 were constructed by replacement of the EcoRI fragment (coding for residues 1–332) from pSG5-hERβ with PCR-generated fragments containing a start codon and coding for residues 32 to 332, 47 to 332, 68 to 332, and 94 to 332, respectively. The pSG5-mERαΔLBD was constructed by ligating a 275-base pair-long PCR-generated KpnI/EagI fragment (Pettersson et al., 1997) and an EagI/SacI fragment of the plasmid MOR (Hillier et al., 1989) into the KpnI andSacI sites of pSP72. The resulting construct was digested with BamHI and FspI, giving a fragment coding for residues 1 to 246 subsequently introduced into pSG5. The pSG5-mERβΔLBD was made by subcloning a BamHI fragment from pSP72-mERβ (Pettersson et al., 1997) (coding for residues 1–171) into the BamHI site of pSG5. The yeast expression plasmids were derived from the 2μ plasmid pGBT9 expressing the Gal4 DNA-binding domain (residues 1–147) (Clontech, Palo Alto, CA). Both plasmids pGBT-ERα184 and pGBT-ERβ103 were constructed by insertion of PCR fragments coding, respectively, for the human ERα (residues 1–184) and the human ERβ (residues 1–103) into theEcoRI-BamHI linearized pGBT9. All PCR generated constructs were verified by sequencing (Cybergene AB).
Electromobility Shift Assays.
Wild-type ERα and ERβ and cognate deletion mutants were in vitro transcribed/translated in the rabbit reticulocyte lysate system (Promega, Madison, WI) with T7 RNA polymerase, according to the instructions of the manufacturer. Six microliters of each lysate was incubated at room temperature for 20 min in binding buffer (15% glycerol, 50 mM KCl, 20 mM HEPES, pH 7.9, 5 mM MgCl, 10 mM dithiothreitol, 5 mM phenylmethylsulfonyl fluoride, 3 mg of pdIdC) containing ∼5 fmol of 32P-labeled consensus ERE. Protein-DNA complexes were separated on a 5% polyacrylamide/0.25× Tris-borate-EDTA gel at ∼10 V/cm, followed by drying and autoradiography at −70°C.
Cell Culture and Transfections.
The human embryonic kidney HEK293 cells and human hepatoma HepG2 cells were grown in 1:1 mixture of Dulbecco's modified Eagle's (DME) medium and Ham's F-12 medium supplemented with 10% fetal bovine serum, 2 mMl-glutamine, 100 I.U./ml penicillin, and 100 μg/ml streptomycin at 37°C in 5% CO2. The human cervix carcinoma HeLa and 3Y1 rat embryonic fibroblasts cells were grown under the same conditions except that DME medium supplemented with 10 or 5% fetal bovine serum, respectively, was used. Cells were plated at approximately 20% confluence in 24-well multidishes 24 h before transfection. HEK293 and HepG2 cells were transiently transfected overnight with Lipofectin according to the recommendations of the manufacturer and Hela cells were transfected for 3 h with the Lipofectamin-Plus reagent. Transfected DNA included 100 ng of reporter plasmid and 10 ng (HEK293 and HepG2 cells) or 1 ng (HeLa cells) of expression plasmid. After exposure to the DNA/lipid mixture, cells were cultured for 24 h in phenol red-free DME/F-12 medium (HEK293 and HepG2 cells) or DME medium (HeLa and 3Y1 cells) supplemented with 5% charcoal/dextran-treated fetal bovine serum, in the absence or presence of hormone. Transfected cells were washed with phosphate-buffered saline and lysed in 0.1 M Tris-acetate, 2 mM EDTA, 1 mM dithiothreitol lysis buffer. Luciferase activities were measured with the GenGlow detection kit (Bio-Orbit Oy) and a Lucy I luminometer (Anthos GmbH, Salsburg, Austria). Luciferase values were corrected for variations in protein concentrations and data are expressed as mean ± S.D. of at least three independent transfection experiments.
Yeast Transactivation Assay.
All details for yeast manipulation can be found in the Clontech Yeast Protocols Handbook. The plasmid pGBT9 or its derivatives pGBT-ERaAF-1 and pGBT-ERbAF-1 were introduced in the yeast reporter strain Y187 (MATa, ura3-52, his3-200, ade2-101, trp1-901, leu2-3, 112, gal4Δ, met − , gal80Δ, URA3::GAL1 UAS -GAL1 TATA -lacZ) with a conventional polyethylene glycol/lithium acetate transformation procedure. Recombinant yeasts were selected for growth at 30°C on selective medium lacking tryptophan. Single colonies were grown overnight in 5 ml of liquid medium lacking tryptophan and then in 25 ml of YPD medium. Mid-log phase cultures were harvested and protein extracts were prepared in 150 μl of Z buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, pH 7.0) by cell disruption with 425- to 600-mm acid-washed glass beads (Sigma). β-Galactosidase activity was determined in triplicate in 20 μl of protein extract with the Galacto-Light Plus detection system (Tropix, Bedford, MA) and a Lucy I luminometer (Anthos GmbH). β-Galactosidase values were corrected for variations in protein concentrations and data are expressed as a mean ± S.D. of three independent experiments.
Results
Cell Type-Dependent Differences between Transcriptional Activities of ERα and ERβ.
The transcriptional activities of the wild-type human ERα and ERβ were first compared with the 2xERE-Tk-Luc reporter plasmid. In HEK293 cells, ERβ transcriptional activity was approximately 50% of that of ERα when cells were treated with 10 nM 17β-estradiol (Fig.1A). The same experiment done in HepG2 cells showed that ERβ had a basal and induced transcriptional activity that was less than 15% of that of ERα (Fig. 1A). In contrast, ERβ transcriptional activity was only slightly reduced compared with that of ERα in HeLa cells (Fig. 1A). The ERβ isoform with an extended amino-terminal region, termed ERβ530 (Ogawa et al., 1998), had the same activity as ERβ in HEK293 cells (Fig. 1A) and other cell lines (data not shown and see Tremblay et al., 1999), suggesting that this additional region is not critical for transactivation. Furthermore, a recent study showed that rat ovary, uterus, and prostate express the 55-kDa ERβ protein (Hiroi et al., 1999). The different transcriptional activity between ERα and ERβ in HEK293 and HepG2 cells is unlikely to be due to autosquelching because we used a dose of ERβ expression vector (10 ng) resulting in the highest inducibility and which was at least 10 times lower than that required for transcriptional autoinhibition (data not shown). Alternatively, this difference between ERα and ERβ could be due to metabolism of 17β-estradiol in these particular cell types, resulting in actual intracellular concentrations too low for fully activating ERβ. To test this possibility we performed the same transfection experiments in HEK293 and HepG2 cells by using the nonmetabolizable estrogen agonist Moxestrol. Similar results were obtained in HEK293 cells and a slightly higher ERβ activity was observed in HepG2 cells, suggesting that no significant metabolism of 17β-estradiol occurred in these two cell lines (Fig. 1B). These results indicate that ERβ is either as transcripitionally active as ERα (HeLa cells) or less active, depending on the cell type (HEK 293 and HepG2 cells). Because ERα contains a cell-specific amino-terminal transcriptional AF-1, the observed cell type-dependent differential transcriptional activity between ERα and ERβ may reflect different functional properties of the highly diverging amino-terminal region of ERβ.

Cell-type-dependent differential transcriptional activity of ERα and ERβ. A, human HEK293, HepG2, and HeLa cells were transfected with a luciferase reporter gene containing a tandem consensus ERE in front of the thymidine kinase promoter (2xERE-Tk-Luc) and ERα or ERβ expression plasmids as indicated. Cells were treated for 24 h with vehicle (−, ■) or 10 nM 17β-estradiol (E2, ▪). B, HEK293 and HepG2 cells were transfected as in A and were treated for 24 h with vehicle (−, ■) or 10 nM Moxestrol (Mox, ▪). Data are expressed as percentage of ERα-activity in the presence of hormone and represent the mean ± S.D. of at least three independent experiments.
Amino-Terminal Deletion Differentially Affects Transcriptional Activity of ERα and ERβ.
To determine whether ERα and ERβ amino-terminal regions contain a similar transcriptional activity, the entire amino-terminal region was deleted in both receptors and the activity of the resulting truncated receptors (termed ERαΔ182 and ERβΔ93, respectively) was compared with that of the corresponding wild-type receptors in different cell type and promoter contexts. Results show that in HEK293 cells, the ERαΔ182 mutant retained slightly more activity compared with the ERβΔ93 mutant when using the 2xERE-Tk promoter (Fig.2A). In contrast, the ERαΔ182 was unable to transactivate the 3xERE-TATA-Luc reporter gene in these cells. The ERβΔ93 mutant showed a 50% decrease of the wild-type activity. In HepG2 cells, ERαΔ182 was inactive with both promoters, whereas the ERβΔ93 retained more than 50% of the wild-type activity (Fig. 2B). In HeLa cells, deletion of the amino-terminal region of ERα decreased its transcriptional activity significantly more when tested with the 3xERE-TATA-Luc reporter gene compared with the 2xERE-Tk-Luc construct (Fig. 2C). Interestingly, the ERβΔ93 mutant showed a substantially higher activity than the wild-type receptor with the 3xERE-TATA-Luc reporter gene, whereas ERα showed a decreased transcriptional activity. These data demonstrate that deletion of the ERβ-amino-terminal domain has a moderate yet a different cell type- and promoter context-dependent effect on the transcriptional activity compared with ERα, suggesting functional differences between the two ERs.

Differential cell type and promoter context effect of the amino-terminal deletion of ERα and ERβ. HEK293 (A), HepG2 (B), and HeLa (C) cells were transfected with luciferase reporter gene under the control of either a 2xERE-Tk or 3xERE-TATA promoter and wild-type (WT) or mutant ERα or ERβ expression plasmids as indicated. Cells were treated for 24 h with vehicle (−, ■) or 10 nM 17β-estradiol (E2, ▪). Data are expressed as percentage of wild-type ER activity in the presence of hormone and represent the mean ± S.D. of at least three independent experiments.
Identification of a Short Amino-Terminal Transcriptional Activation Domain in ERβ.
As shown in Fig. 2, both ERα and ERβ transcriptional activities were decreased in HEK293 cells transfected with the 3xERE-TATA promoter, on deletion of the entire amino-terminal domain of the receptors. These conditions were therefore used to localize the region of ERβ responsible for the amino-terminal transcriptional activity of the receptor and compare it with that of ERα. To this end, ERα and ERβ amino-terminal deletion mutants depicted in Fig. 3A were created. All these mutants showed similar DNA-binding activities when tested in an electromobility shift assay with a consensus ERE as a probe (Fig. 3 B). Deletions of the first 63 amino acids in ERα (mutant ERαΔ63) resulted only in a moderate decrease of approximately 30% of the transcriptional activity (Fig. 3C). Further deletion of ERα up to amino acid 142 resulted in a dramatic decrease of the activity. The ERαΔ182 mutant behaved similarly to the ERαΔ142 mutant. These results with the HEK293 cell line are in agreement with previous findings that used other cell lines and indicate that the domain encompassing amino acids 50 to 143 is critical for ERα AF-1 activity in most cell types. In contrast to ERα, deletion of the extreme amino-terminal region of ERβ (ERβΔ31 mutant) produced a 60% decrease of the transcriptional activity compared with wild-type receptor (Fig. 3D). When ERβ was deleted beyond amino acid 31 up to amino acid 93, no additional loss of transcriptional activity was observed. These data show that most if not all the amino-terminal transcriptional activity of ERβ is attributable to a short domain mapping to the extreme amino terminus of the protein. This situation contrasts with that observed with ERα, in which the amino-terminal transcriptional AF-1 extends over 80 to 100 amino acids. Collectively, these comparative data clearly show a different functional organization of the amino-terminal domains of ERα and ERβ.
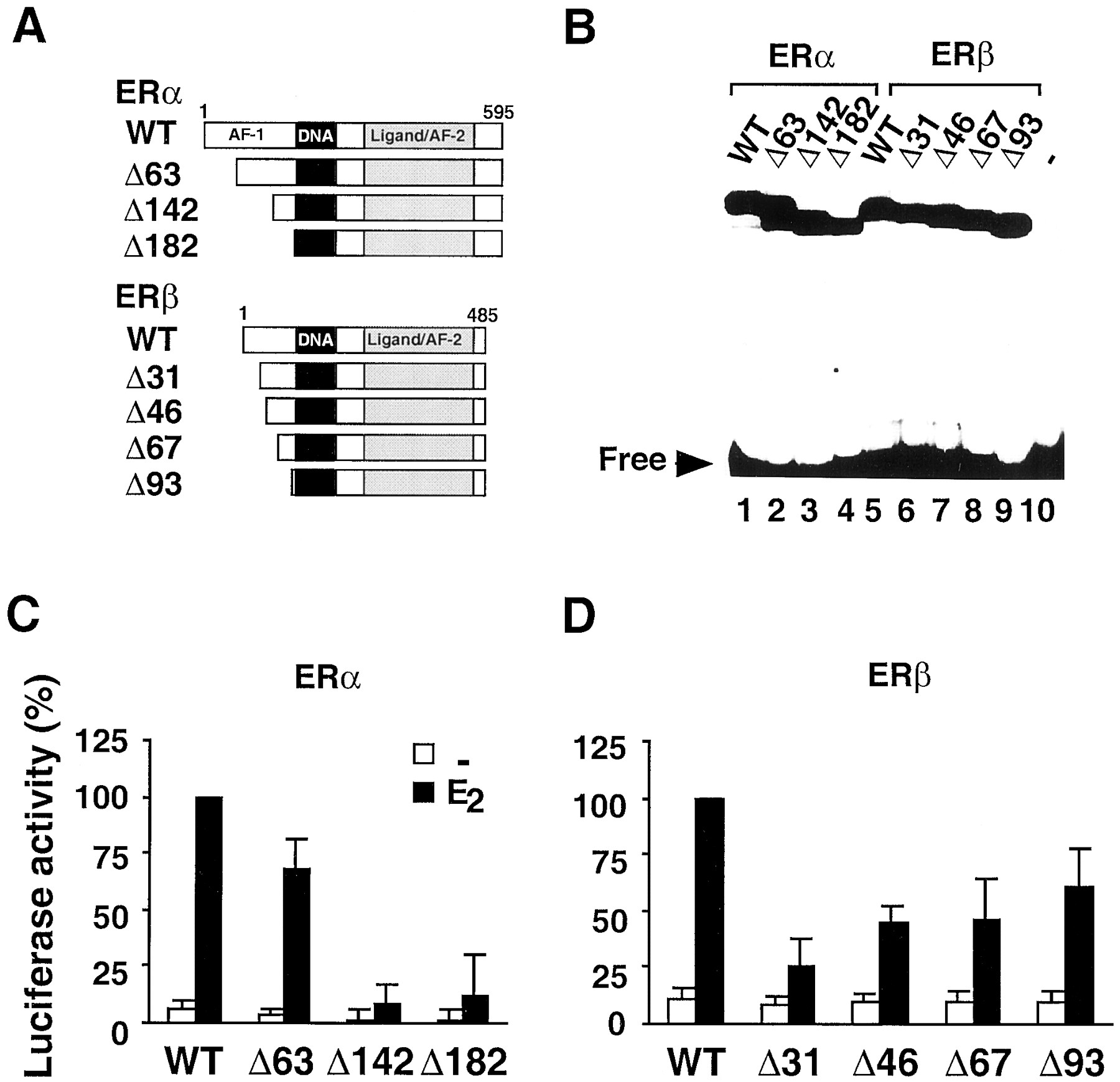
Transcriptional activity of ERα and ERβ amino-terminal deletion mutants in HEK293 cells. The structure (A) of the deletion mutants derived from wild-type ERα and ERβ is shown. The domains corresponding to the AF-1 and AF-2 in ERα, DBD, and LBD are indicated. The ERα- and ERβ-deletion mutant nomenclature refers to the number of amino acids deleted from the amino-terminus. For analysis of the DNA-binding activity (B) of the ER mutants, wild-type ERα (lane 1) and ERβ (lane 5) and the various corresponding deletion mutants (lanes 2–4 and lanes 6–9, respectively) or unprogrammed lysate (lane 10) were in vitro translated and incubated with 32P-labeled consensus ERE, and DNA-protein complexes were separated on a nondenaturing polyacrylamide gel. Cells were transfected with expression plasmid for wild-type or mutant ERα (C) and the 3xERE-TATA-Luc reporter plasmid. Cells were treated with vehicle (−, ■) or 10 nM 17β-estradiol (E2, ▪) for 24 h and assayed for luciferase activity. D, transfection conditions were as in C, except that expression plasmids for wild-type or mutant ERβ were used. Data are expressed as percentage of ERα or ERβ wild-type activity in the presence of hormone and represent the mean ± S.D. of at least three independent experiments.
ERβ Amino-Terminal Transcripitonal Activity Is Not Autonomous and Ligand-Independent.
Fusion of the ERα amino-terminal domain to the heterologous Gal4 DNA-binding domain demonstrated that this domain exhibits a constitutive and ligand-independent transcriptional activity in yeast as previously reported (Metzger et al., 1995). In contrast, the amino-terminal domain of ERβ did not show any activity in this transactivation assay, suggesting that the ERβ amino-terminal domain cannot function in an autonomous manner (Fig.4). To further investigate this lack of constitutive activity in ERβ and exclude a possible yeast inhibitory effect, we used 3Y1 rat embryonic fibroblasts in which ERα exhibits a significant ligand-independent AF-1 activity. These cells were cotransfected with expression plasmids coding for ERα and ERβ mutants with no ligand-binding domain and the 3xERE-TATA-Lucreporter gene. In these cells, the ERα mutant showed a ligand-independent constitutive activity, whereas the ERβ mutant had no activity. Collectively, these data show that in contrast to ERα, ERβ has no autonomous and ligand-independent amino-terminal transcriptional activity.

ERβ has no autonomous and ligand-independent amino-terminal transcriptional activity. A, Gal4 DBD was fused to the amino-terminal domains of ERα and ERβ, respectively, and the resulting chimeric proteins were introduced into the yeast reporter strain Y187. β-Galactosidase activities were expressed as percentage of that obtained with the Gal4-ERα construct. Data represent mean ± S.D. of three independent experiments. B, LBD was deleted from ERα and ERβ to create the ERαDLBD and ERβDLBD deletion mutant, respectively. The mutant receptors were transfected in 3Y1 rat embryonic fibroblasts together with the 3xERE-Luc reporter plasmid and luciferase activity was measured 24 h later. Data are expressed as percentage of ERα-activity in the presence of hormone and represent the mean ± S.D. of at least three independent experiments.
Discussion
Estrogen receptors α and β are highly homologous in their DNA- and ligand-binding domains. Likewise, functions associated with these structural domains such as ERE binding, dimerization, and affinity to the natural estrogen 17β-estradiol are very similar for ERα and ERβ (Cowley et al., 1997; Kuiper et al., 1997; Pace et al., 1997; Pettersson et al., 1997). Yet, there is accumulating evidence that the two receptors function differently. These differences include, for instance, lower transcriptional activity of ERβ in several cell lines (Fig. 1) (Vegeto et al., 1993; Cowley et al., 1997), higher binding affinity of ERβ for the phytoestrogens coumestrol and genistein (Kuiper et al., 1997), absence of agonistic activity of the antiestrogen 4-hydroxytamoxifen on ERβ through ERE sites (Cowley et al., 1997; Tremblay et al., 1997), opposite signaling between ERα and ERβ at AP-1 sites with 17β-estradiol and raloxifene (Peach et al., 1997), and binding and activation of SF1 response elements by ERα but not ERβ (Vanacker et al., 1999). The structural determinants and molecular mechanisms for such differences are not understood, but the poor conservation of the amino-terminal region between ERα and ERβ may play a significant role by, for instance, providing the receptors with different AF-1 domains.
Using amino-terminal deletions of ERα and ERβ in various cell types and promoter configurations, we show specific transcriptional activity properties for the two ER subtypes. Recently, McInerney et al. (1998)showed that substitution of the amino-terminal region of ERβ by that of ERα could confer to the resulting chimeric receptor an improved transcriptional activity in response to estradiol in some cell types and an agonistic response to antiestrogens that contrasts to wild-type ERβ (McInerney et al., 1998). These data together suggest that the amino-terminal region plays an important role in the control of the transcriptional activity specificity exhibited by both ER subtypes.
In this study, we show that in cell types and promoter contexts under which ERβ exhibits an amino-terminal transcriptional activity, this activity is significantly weaker than that of ERα. This observation correlates with the finding that the region responsible for the amino-terminal activity in ERβ is restricted to a short domain of 30 amino acids, whereas it extends over most of the amino-terminal region in ERα. Interestingly, this domain contains a six amino-acid motif that is highly conserved between all mammalian ERαs and ERβs and in ERα, this motif is part of the conserved box 1 that was defined as one of the subdomains controlling AF-1 activity (Fig.5) (White et al., 1987; Metzger et al., 1995). Thus, despite a poor sequence homology and important functional differences between ERα and ERβ amino-terminal regions, the presence of this conserved motif suggests that it plays a role in the control of the amino-terminal transcriptional activity of both ER subtypes. However, deletion of residues 2 to 63 in ERα (Δ63ERα mutant), thus excluding this motif, had a moderate effect, suggesting that this motif is not a major activating domain. This is consistent with the weak ERβ amino-terminal transcriptional activity (see below). In an evolutionary perspective, this observation also implies that this motif was present in the nonduplicated ancestral ER gene in early vertebrates. The differential loss of transcriptional activity on deletion of the amino-terminal domain of ERα and ERβ suggests that their interactions with transcriptional intermediary factors are qualitatively and quantitatively different. Moreover, the opposite effect of the amino-terminal deletion in ERα (loss of activity) and ERβ (increased activity) observed in HeLa cells transfected with the 3xERE-TATA-luc reporter gene (Fig. 2C) indicates that the ERα and ERβ amino-terminal regions are targets for different cofactors, and suggests that ERβ is a putative target for corepressors. This possibility also is supported by recent data showing that GRIP1, a coactivator member of the p160 transcriptional intermediary factor group directly interacts with the ERα amino-terminal domain and potentiates 4-hydroxytamoxifen transcriptional activity, whereas no significant interaction was detected with ERβ (Webb et al., 1998).

Mammalian ERα and ERβ amino-terminal domains contain a highly conserved motif. Localization and alignment of the conserved motif found in the AF-1 domain of ERα and ERβ from various mammalian species. Position of the proline residue is indicated in parentheses. Accession number for the ER sequences used are human ERα, X03635; mouse ERα, M38651; rat ERα, Y00102; camel ERα,X98107; sheep ERα, Z49257; human ERβ, X99101; mouse ERβ,AJ000220; and rat ERβ, U57439.
Although our deletion analysis identified the extreme amino-terminus of ERβ as a transcriptional activation domain in the context of the wild-type receptor, we found surprisingly that the ERβ amino-terminal domain was not transcriptionally autonomous when fused to the heterologous yeast Gal4 DBD, in contrast to that of ERα (Metzger et al., 1995) (Fig. 4). This may explain why the amino-terminal transcriptional activity of ERβ is generally weaker than that of ERα. This finding also suggests that synergism between the amino- and carboxyl-terminal regions is required irrespective of the cell type for ERβ-transcriptional activation, whereas it is cell type-dependent for ERα. Nevertheless, the lack of ligand-independent autonomous activity of the ERβ AF-1 domain does not rule out the possibility for such an activity under certain physiological conditions. For instance, it was recently shown that phosphorylation of the ERβ-amino-terminal domain resulted in the recruitment of the coactivator SRC-1 and subsequent ligand-independent transcriptional activation (Tremblay et al., 1999).
Interestingly, in HeLa cells transfected with the 2xERE-Tk-Luc construct, a condition showing very low ligand-independent ERα AF-1 activity (Berry et al., 1990), ERβ transcriptional activation potency was only slightly reduced compared with that of ERα (Fig. 1). This supports the idea that in cell types and promoter contexts that require an AF-1 for full ER trancriptional activity, the lower transactivation capacity of ERβ results from a weaker amino-terminal transcriptional activity compared with ERα. This is also in agreement with the finding that 4-hydroxytamoxifen acts only as a pure antagonist through ERβ, whereas it is an agonist for ERα on a promoter from which transcription can be regulated by the ERα AF-1 on its own (Berry et al., 1990; McInerney and Katzenellenbogen, 1996).
Collectively, our comparative data show that ERα- and ERβ-amino-terminal domains are functionally different and establish that ERβ does not contain a typical AF-1 domain in contrast to ERα. This study provides a molecular basis for at least some of the differential transcriptional activities between both ER subtypes.
Acknowledgments
We thank Dr. Bart van der Burg for the gift of the 3xERE-TATA-Luc reporter plasmid.
Footnotes
- Received March 3, 2000.
- Accepted June 6, 2000.
-
Send reprint requests to: Jan-Åke Gustafsson, Department of Medical Nutrition, NOVUM, Karolinska Institute, S-141 86 Huddinge, Sweden. E-mail: jan-ake.gustafsson{at}csb.ki.se
-
↵1 Present address: Ecole Normale Supérieure, CNRS UMR 5665, 46 allée d'Italie, Lyon 69364 Cedex, France.
-
This work was supported by grants from the Swedish Cancer Society and from KaroBio AB (to J.-Å.G.).
Abbreviations
- ER
- estrogen receptor
- DBD
- DNA-binding domain
- LBD
- ligand-binding domain
- ERE
- estrogen response element
- AF
- activation function
- PCR
- polymerase chain reaction
- DME
- Dulbecco's modified Eagle's
- SRC-1
- steroid receptor coactivator-1
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}