


Abstract
In situ hybridization with cRNA probes showed A2A receptor and Golf mRNAs to be abundantly expressed in caudate putamen, nucleus accumbens, and olfactory tubercle, whereas Gs mRNA shows a comparatively low expression in regions expressing A2A receptors. In caudate putamen, 49% of the medium-sized neuron-like cells exhibited a strong signal for adenosine A2A receptor mRNA, and 98% showed a strong signal for Golf mRNA. In contrast, Gs mRNA was found in only 12% of the medium-sized neuron-like cells in caudate putamen. The coexpression of adenosine A2A receptor mRNA with that of Golf or Gs mRNAs was studied with double in situ hybridization. A large majority (91–95%) of the neurons in caudate-putamen that contained adenosine A2A receptor mRNA also expressed Golf mRNA, whereas only 3 to 5% of the neurons with adenosine A2A receptor mRNA coexpressed Gs mRNA. The A2A receptor agonist CGS 21680 [2-[p-(2-carbonylethyl)phenylethylamino-5′-N-ethylcarboxamidoadenosine] dose dependently activated Golf subunits in striatal membranes as shown by photolabeling with [α-32P]m-acetylanilido-GTP followed by immunoprecipitation with a specific antibody against Golf. Transfection of Golf cDNA into Chinese hamster ovary cells, which stably express human adenosine A2A receptors, led to an increased efficacy of CGS 21680, as evidenced by a stronger cAMP response, indicating that activation of Golf by A2A receptors leads to a biological signal. In conclusion, these results provide anatomical and biochemical evidence that adenosine A2A receptors stimulate Golf rather than Gs in striatum.
The nucleoside adenosine can influence most mammalian cell types via stimulation of four G protein-coupled receptors: subtypes A1, A2A, A2B, and A3 (Fredholm et al., 1994). The nature of the responses to adenosine and other agonists depends on the selective coupling of the activated receptor to distinct G proteins. The A1 (Freissmuth et al., 1991;Munshi et al., 1991; Jockers et al., 1994) and A3(Zhou et al., 1992; Palmer et al., 1995) receptors are coupled with Gi proteins inhibiting adenylyl cyclase, whereas the A2A and A2B receptors (van Calker et al., 1979; Londos et al., 1980; Pierce et al., 1992) couple to Gs-like proteins and activate adenylyl cyclase. Three stimulatory G proteins have been biochemically characterized: Gs short, Gslong, and Golf (Jones et al., 1990). They show 88% identity in their amino acid sequences (Jones and Reed, 1989). The Gs subunits have a widespread distribution, whereas the Golf subunit is distributed in a more restricted manner (Herve et al., 1993). Golf was first shown to be expressed in olfactory epithelium and was postulated to be exclusively involved in olfactory signaling (Jones and Reed, 1989). However, Golf was later found also in other brain regions and the expression was particularly high in striatum (Drinnan et al., 1991; Herve et al., 1993). The striatal expression may be functionally important: recently developed transgenic mice deficient in Golf are not only anosmic but are also hyperactive (Belluscio et al., 1998).
Thus, Golf present in striatum probably plays a key role in the signal transduction mediated by G protein-coupled receptors in this area. Previous work has shown that Golf is the main stimulatory G protein coupling dopamine D1 receptors to adenylyl cyclase in the striatum (Herve et al., 1993). D1 receptors are abundantly expressed in approximately 50% of the striatal neurons, mostly in those projecting to the substantia nigra and containing γ-aminobutyric acid, substance P, and dynorphin (Gerfen et al., 1990;Le Moine et al., 1991). Adenosine A2A receptors are also highly expressed in the striatum (Jarvis and Williams, 1989;Parkinson and Fredholm, 1990). However, adenosine A2A receptors are segregated from dopamine D1 receptors and are selectively expressed in the striatopallidal neurons that also contain enkephalin and dopamine D2 receptors (Schiffmann et al., 1991; Fink et al., 1992; Svenningsson et al., 1997). This is the other major subpopulation of projection neuron within the striatum. Because Golf is highly expressed in the striatal neurons and A2A receptors stimulate adenylyl cyclase and cAMP-dependent signal transduction in striatum (Fredholm, 1977;Svenningsson et al., 1998), it seemed important to examine whether A2A receptors may couple to Golf in the striatum. The aim of the present study was therefore to investigate the cellular localization of Golf, Gs, and adenosine A2A receptors in rat brain, and to investigate, using a photoaffinity labeling technique, whether adenosine A2A receptors are functionally coupled to Golf.
Materials and Methods
Chemicals.
Aprotinin, m-aminoacetophenone, 1,4-dioxane, Triton X-100, sodium deoxycholate, phenymethylsulfonyl fluoride, protein A-Sepharose, tergitol Nonidet P-40, 4-morpholineethanesulfonic acid,N-ethyl-N′-(3-dimethylamino-propyl)carbodi-imide hydrochloride, and cAMP were purchased from Sigma (St. Louis, MO). CGS 21680 [2-[p-(2-carbonyl-ethyl)-phenylethylamino]-5′-N-ethylcarboxamidoadenosine] was obtained from Research Biochemicals International (Natick, MA). [3H]cAMP was obtained from DuPont-NEN (Boston, MA). SCH 58261 [5-amino-7-(2-phenylethyl)-2-(2-furyl)-pyrazolo[4,3-e]-1,2,4-triazolo[1,5-c]pyrimidine] was a gift from Dr. E. Ongini (Schering-Plough Research Institute, Milan, Italy). Digoxigenin-11-UTP and FuGENE 6 transfection reagent were purchased from Boehringer Mannheim (Mannheim, Germany).35S-UTP, [α-32P]GTP, and 4-morpholinepropanesulfonic acid were purchased from Amersham (Little Chalfont, England).
Tissue Preparation.
Adult male Sprague-Dawley rats (200–250 g) were briefly anesthetized with CO2 and sacrificed immediately by decapitation. For the in situ hybridization experiments, the brains were dissected out and frozen on dry ice. Consecutive coronal sections (14 μm) were made through the rostral part of striatum. For the photolabeling experiments, striatum was dissected out and homogenized on ice in 50 mM Tris·HCl, pH 7.4 with an Ultra Turrax (3 ×10 s). The homogenate was centrifuged for 10 min (1,000g) after which the pellet was discarded. The supernatant was centrifuged for 50 min (30,000g) and washed once with 50 mM Tris·HCl, pH 7.4. After the second centrifugation, the pellet containing the membranes was resuspended in an incubation buffer (20 mM MgCl2, 400 mM NaCl, 120 mM HEPES, and 0.4 mM EDTA) and stored at −80°C in aliquots until used.
Subcloning of cDNA Fragments for In Vitro Transcription Templates.
cDNA fragments corresponding to the rat adenosine A2A receptor, the rat α subunit Gs, and the rat α subunit Golf were amplified with the use of polymerase chain reaction (PCR). The PCR products were subcloned into pBluescript II (Stratagene, La Jolla, CA). To verify the incorporation of the cDNA into pBluescript II, the subcloned cDNA fragments were sequenced using an automated DNA sequencer, ABI 373A (Applied Biosystems Inc., Foster City, CA).
Probe Synthesis and Labeling for In Situ Hybridization.
35S- or digoxigenin-labeled antisense and sense cRNA probes were prepared by in vitro transcription from cDNA clones corresponding to fragments of rat adenosine A2Areceptor (Fink et al., 1992), rat G protein α subunit Gs (Jones and Reed, 1987), and rat G protein α subunit Golf (Jones and Reed, 1989). The transcription was performed using MAXI-script in vitro transcription kit according to the manufacturer's protocol (Ambion Inc., Austin, TX).
Single In Situ Hybridization.
The in situ hybridization experiments were performed as described previously by Le Moine and Bloch (1995). Cryostat sections were postfixed in 4% paraformaldehyde, dehydrated in graded alcohol, and were hybridized overnight at 55°C with 106 cpm of 35S-labeled probe in 50 μl of hybridization solution. After washing, the slides were dipped into Ilford K5 emulsion [diluted 1:3 in standard sodium citrate (SSC) buffer], exposed for 8 to 12 weeks, developed, and stained with cresyl violet.
Double In Situ Hybridization.
In double in situ hybridization experiments, the adenosine A2Areceptor cRNA probe was labeled with 35S-UTP, whereas the probes against Gs and Golf mRNAs were labeled with digoxigenin-11-UTP. Cryostat sections were pretreated as described above. The sections were hybridized overnight at 55°C with a combination of35S- and digoxigenin-labeled probes (106 cpm of 35S-labeled probe and approximately 20 ng of digoxigenin-labeled probe in 50 μl of hybridization solution). The slides were washed in RNase A and various concentrations of SSC, but without dithiothreitol (DTT). At the end of the washes, the slides were put in 0.1× SSC at room temperature. The sections were rinsed twice for 5 min in buffer A (1 M NaCl/0.1 M Tris/2 mM MgCl2, pH 7.5), and then for 30 min in buffer A containing 3% normal goat serum and 0.3% Triton X-100. After 5 h of incubation at room temperature with alkaline phosphatase-conjugated antidigoxigenin antiserum (Boehringer Mannheim; 1:1000 in buffer A/3% normal goat serum/0.3% Triton X-100), the sections were rinsed in buffer A (5 min, twice), then twice for 10 min in buffer B (1 M NaCl/0.1 M Tris/5 mM MgCl2, pH 9.5), and twice for 10 min in 0.1 M buffer B (containing 0.1 M NaCl). The sections were then incubated overnight in the dark at room temperature in 0.1 M buffer B, pH 9.5, containing 0.34 mg/ml nitroblue tetrazolium and 0.18 mg/ml bromochloro-indolylphosphate. The sections were thereafter rinsed in 0.1 M buffer B, pH 9.5, then in 1× SSC, dried and dipped into Ilford K5 emulsion. After being exposed for 7 to 11 weeks, the sections were developed and mounted without counterstaining.
Preparation of Radioactivem-Acetylanilido-GTP.
The preparation of radioactivem-acetylanilido-GTP (m-AcAGTP) was performed as described by Zor et al. (1995). Briefly, [α-32P]GTP (3000 Ci/mmol) was freeze-dried and redissolved in 50 μl of 0.125 M 4-morpholineethanesulfonic acid buffer, pH 6.5.N-Ethyl-N′-(3-dimethylamino-propyl)carbodi-imide hydrochloride (10 μmol) was then dissolved in this solution andm-aminoacetophenone (20 μmol) in 20 μl of 1,4-dioxane was added. After 5 h at room temperature, the mixture was freeze-dried and redissolved in 10 mM 4-morpholinepropanesulfonic acid buffer, pH 7.0. Insoluble m-aminoacetophenone was removed by centrifugation. The product was stored at −18°C until used.
After the synthesis of [α-32P]m-AcAGTP, the photoreactivity of the product was tested by UV irradiation of the compound spotted onto polyethylenimine-cellulose plates followed by chromatography with 1 M LiCl as mobile phase. Approximately 30 to 40% of the irradiated m-AcAGTP did not migrate from the origin (data not shown). Nonirradiated m-AcAGTP had a retardation factor value twice that of [α-32P]GTP, which agrees with that shown previously (Zor et al., 1995).
Photoaffinity Labeling of Striatal Membranes with [α-32P]m-AcAGTP.
First we optimized the photoaffinity reaction with regard to Mg2+, NaCl, and GDP concentrations. The reaction mixture that gave the largest ratio of incorporated [α-32P]m-AcAGTP between stimulated and unstimulated membranes was selected for further use. The optimal reaction mixture contained, in a total volume of 60 μl, 50 μg of membranes, 5 mM MgCl2, 100 mM NaCl, 0.1 mM EDTA, 0.6 mM ATP, 100 μM GDP, and the adenosine A2Aagonist CGS 21680 when indicated. After 3 min incubation at 37°C, 1.5 μCi of [α-32P]m-AcAGTP was added and the samples were incubated for additional 5 min at 37°C. The reaction was terminated by dilution with 400 μl of ice-cold incubation buffer (5 mM MgCl2, 100 mM NaCl, 30 mM HEPES, 0.1 mM EDTA, and 2 mM DTT). Excess photoaffinity label was removed by centrifugation at 4°C (12,000g for 5 min) and the pellet was resuspended in 60 μl of incubation buffer. The membranes were pipetted into dimples made in aluminum foil pressed against an empty Eppendorf tube rack. The aluminum foil was wrapped over a glass dish filled with ice. UV irradiation (λl = 320 nm) was applied for 20 min from a distance of 2 cm from the samples. Photolabeled membranes were pelleted and solubilized in 2% SDS for 10 min at room temperature before SDS/polyacrylamide gel electrophoresis (PAGE; 12%) and autoradiography or immunoprecipitation.
Immunoprecipitation of α Golf Protein.
The solubilized, photolabeled membranes were diluted 1:1 with immunoprecipitation buffer (10 mM Tris·HCl, pH 7.4, 1% Triton X-100, 1% sodium deoxycholate, 0.5% SDS, 150 mM NaCl, 1 mM DTT, 1 mM EDTA, 10 μg/ml aprotinin, and 0.2 mM phenylmethylsulfonyl fluoride) and centrifuged at 12,000g for 10 min at 4°C. The pellet from this centrifugation was discarded and the supernatant was mixed with 3 μl of nondiluted, subtype-specific G protein α subunit Golf antibody (Santa Cruz Biotechnology, Santa Cruz, CA), which does not cross-react with the G protein α subunit Gs, and was incubated for 1 h at 4°C under constant rotation. Then 60 μl of protein A-Sepharose (4 mg) in immunoprecipitation buffer was added to each sample and incubated overnight at 4°C under constant rotation. Thereafter, the Sepharose beads were pelleted (1 min at 12,000g, 4°C) and washed twice with 1 ml of washing buffer A (50 mM Tris·HCl, 600 mM NaCl, 0.5% SDS, and 1% tergitol Nonidet P-40, pH 7.4) and twice with washing buffer B (100 mM Tris·HCl, 300 mM NaCl, and 10 mM EDTA, pH 7.4). The washed Protein A-Sepharose was then resuspended in 100 μl of Laemmli buffer and heated at 80°C for 5 min and then centrifuged as above. Fifty microliters of the supernatant was then subjected to SDS/PAGE (12%), as described by Laemmli (1970). The gel was dried in a gel drier and exposed to X-ray film.
Subcloning of the Rat αGolf cDNA.
The rat G α subunit Golf cDNA was amplified with PCR using a plasmid (Bluescript) containing the rat Golf cDNA sequence (a gift from Dr. A.G. Gilman, Southwestern Medical Center, Dallas, TX). The PCR product was subcloned into the vector pCI-neo (Promega, Scandinavian Diagnostic Services, Falkenberg, Sweden). To verify the incorporation of the cDNA into pCI-neo, the subcloned cDNA fragments were sequenced using an automated DNA sequencer, ABI 373A (Applied Biosystems Inc.).
Transient Transfection of Golf cDNA into Chinese Hamster Ovary (CHO) Cells Expressing Human Adenosine A2AReceptors.
CHO-K1 cells (American Type Culture Collection, Rockville, MD) stably expressing human adenosine A2A receptors (Kull et al., 1999) were grown adherent and maintained in α-minimum essential medium without nucleosides, containing 10% fetal calf serum, penicillin (50 U/ml), streptomycin (50 μg/ml), l-glutamine (2 mM), and geneticin (Life Technologies, Täby, Sweden; 500 μg/ml) at 37°C in 5% CO2/95% air. Transient transfection of Golf cDNA or control plasmid was performed using FuGENE 6 transfection reagent (Boehringer Mannheim) according to the instruction manual.
Measurement of cAMP Accumulation.
Thirty-six hours after transfection, both Golf cDNA-transfected cells and control cells were sown into 12-well plates (200,000 cells per well) and allowed to grow for 36 h. Cells were then washed twice with HEPES-buffered (20 mM) α-minimal essential medium, pH 7.4. The cells were incubated at 37° for 10 min in 0.9 ml of HEPES-buffered medium. The adenosine A2A agonist CGS 21680 was added in 0.1 ml of medium and the cells were incubated for another 10 min. The reactions were terminated by the addition of perchloric acid to a final concentration of 0.4 M. After 1 h at 4°C, the acidified cell suspensions were transferred to tubes and neutralized with 4 M KOH/1 M Tris·HCl. The cAMP content in the samples was determined using a competitive radioligand-binding assay (Nordstedt and Fredholm, 1990). Radioactivity was measured in an LKB/Pharmacia scintillation counter with 3 ml of ReadySafe (Beckman, Bromma, Sweden) scintillation fluid.
Data Analysis.
In the double in situ hybridization experiments with probes against adenosine A2Areceptor mRNA and Golf or Gs mRNAs in the striatum, three categories of neurons were counted: those that only exhibited a radioactive signal (i.e., at least two times the background), those that showed only a nonradioactive signal, and those that showed both signals. Quantification was made in the lateral and medial parts of striatum and in the core and shell regions of nucleus accumbens. All neurons within the examined areas were counted.
Autoradiographic data from photolabeling experiments were quantified using densitometry (MCID system; Imaging Research, St. Catharines, Canada). Analysis of dose response curves from cAMP measurements, nonlinear regression analysis, and statistical analysis (ttest) were performed using GraphPad Prism (ver. 3.00 for Windows; GraphPad, San Diego, CA).
Results
Regional Distribution of Golf, Gs, and A2A Receptors in Rat Forebrain.
To study the abundance and distribution of mRNAs for Golf, Gs, and A2A receptors, we performed in situ hybridization on coronal sections from the rat brain (Fig. 1). Adenosine A2A receptor mRNA was, as shown previously (Svenningsson et al., 1997), abundant in the caudate putamen, the nucleus accumbens, and the olfactory tubercle (Fig. 1a). Golf mRNA was also found in the caudate putamen, the nucleus accumbens, and the olfactory tubercle. In addition, Golf mRNA, but not adenosine A2A receptor mRNA, was detected in e.g., the pyramidal cell layer of piriform cortex and in the Islands of Calleja (Fig. 1b). These neurons in the Islands of Calleja are known to be interconnected with the olfactory tubercle (Fallon, 1983) and to express dopamine D1 and D3receptors (Le Moine and Bloch, 1996; Svenningsson et al., 1997). Gs mRNA was most abundant in areas not expressing adenosine A2A receptor mRNA, such as pyramidal cells of the piriform cortex, cerebral cortex, claustrum, endopiriform nucleus, and the diagonal band of Broca. In addition, Gs mRNA was found throughout septum, whereas mRNA encoding A2A receptors was found only in occasional cells in lateral septum. Moderate levels of Gs mRNA were detected in nucleus accumbens, whereas caudate putamen only exhibited very low expression (Fig. 1c).

Distribution of adenosine A2A, Golf, and Gs in rat brain. Darkfield autoradiograms showing the expression of (a) adenosine A2Areceptor mRNA, (b) Golf mRNA, and (c) Gs mRNA in coronal sections of the rat forebrain. Regions expressing the genes appear white. Scale bars, 1 mm.
Cellular Localization of Golf, Gs, and A2A Receptor mRNA within the Striatum.
There are three major subcategories of striatal neurons: medium-sized (∼20 μm in diameter) neurons (which are further subdivided into spiny and aspiny neurons) and large-sized (∼40 μm in diameter) neurons. Of the striatal neurons, 95% are medium-sized spiny neurons and the remaining 5% are medium-sized aspiny neurons and large-sized neurons. Because no staining for detecting spines was used in the present study, we could only distinguish between neurons based on their size.
In caudate putamen and nucleus accumbens, 49% of the medium-sized-neuron-like cells exhibited a strong signal for adenosine A2A receptor mRNA, which is in agreement with our previous report (Svenningsson et al., 1997). GolfmRNA was found in the majority (98%) of the medium-sized neurons within the striatum. No expression of A2Areceptor or Golf mRNAs was found in large-sized neurons. The labeling of Gs mRNA in striatum was weak and most of the label seemed to be located in glial rather than neuronal cells. Gs mRNA was occasionally found in large-sized neurons. Of the medium-sized neurons, only 12% in caudate putamen and 27% in nucleus accumbens showed detectable Gs mRNA.
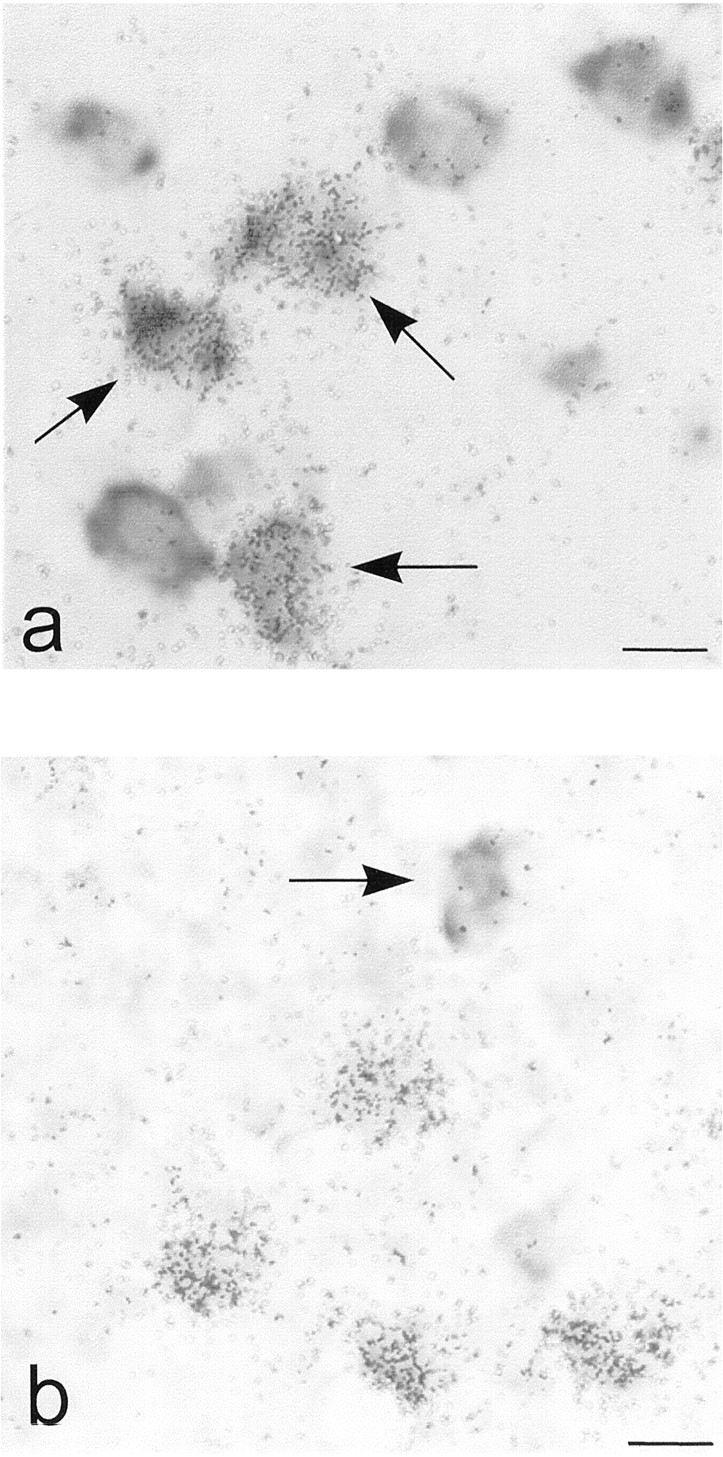
To determine the coexpression of adenosine A2Areceptor mRNA with that of Golf or Gs in caudate putamen and nucleus accumbens, we performed double in situ hybridization. Quantifications were made in the lateral and medial parts of striatum and in the core and shell regions of nucleus accumbens. As expected from the above-mentioned data, the large majority of the adenosine A2Areceptor mRNA-containing neurons also expressed Golf mRNA in the lateral and medial parts of striatum as well as in the core and shell regions of nucleus accumbens (95, 91, 91, and 88%, respectively) (Fig.2a, 3). By contrast, only a minority (3 to 5%) of the adenosine A2A receptor mRNA-containing neurons coexpressed Gs mRNA in caudate-putamen (Figs. 2b and 3). In nucleus accumbens, the extent of coexpression of A2A receptor mRNA and Gs mRNA was somewhat higher (13 to 21%). When Golf mRNA positive neurons were considered, 47 to 56% of them were also positive for A2A receptor mRNA labeling in the examined areas of caudate-putamen and nucleus accumbens (Figs. 2a and 3). Of the few neurons that expressed Gs mRNA, 17 to 36% in caudate putamen and 33 to 42% in nucleus accumbens also expressed A2Areceptor mRNA (Fig. 3). We could also detect a high level of Gs mRNA expression in some large-sized neurons that did not express A2A or Golf mRNAs (data not shown).

Colocalization of adenosine A2Areceptors with Golf and Gs. Emulsion autoradiograms from double in situ hybridization experiments showing (a) the colocalization of adenosine A2A receptor mRNA (silver grains) with Golf mRNA (dark cells) in caudate putamen (arrows indicate colocalization) and (b) the absence of colocalization of adenosine A2A receptor mRNA (silver grains) with Gs mRNA (dark cell) in caudate putamen (arrow indicates a single labeled neuron for Gs mRNA). Scale bars, 10 μm.

Histograms showing the numbers of neurons in the lateral and medial caudate-putamen and in the core and shell of nucleus accumbens that are labeled (+) or not labeled (−) with probes for adenosine A2A receptor mRNA, Golf mRNA and Gs mRNA. All neurons within the examined areas were counted.
Photolabeling of G Proteins in Striatal Membranes.
To investigate whether Golf is activated on stimulation with an A2A receptor agonist, we used a photolabeling technique (Zor et al., 1995). We first established that G proteins present in the membranes can be photolabeled by [α-32P]m-AcAGTP and that activation of G proteins leads to an increase in this photolabeling. We incubated striatal membranes with [α-32P]m-AcAGTP with and without a high concentration (10 μM) of the adenosine A2Aagonist CGS 21680 and with different concentrations (3–100 μM) of GDP. The labeling was then visualized by SDS/PAGE followed by X-ray autoradiography. The crude photolabeled membranes gave a broad band with an apparent molecular mass of 40 to 50 kDa (Fig.4a). In a parallel experiment, the Golf subunits were immunoprecipitated with a specific antibody directed against Golf, and separated with the use of SDS-PAGE. The incorporated [α-32P]m-AcAGTP label was determined by autoradiography (Fig. 4b). The immunoprecipitate gave a single band with an apparent molecular mass of 44 to 45 kDa, a size identical with the molecular mass of the protein visualized in immunoblot experiments (data not shown). By comparing the intensity of [α-32P]m-AcAGTP labeling in the crude membranes, it was not possible to distinguish between control conditions and in the presence of agonist at any of the GDP concentrations. However, by comparing the intensity of labeling after immunoprecipitation, it was clear that the labeling was enhanced in the stimulated samples. At low (3–10 μM) GDP concentrations, the basal labeling was strongest, but the ratio of agonist-stimulated to basal photolabeling of the Golf protein was largest at high (30 to 100 μM) GDP concentrations (Fig. 4, b and c).

Agonist-stimulated photoaffinity labeling of Golf is influenced by GDP. Membranes from striatum were photolabeled with [α-32P]m-AcAGTP at various GDP concentrations in the absence (−) or presence (+) of CGS 21680 (10 μM). Autoradiogram showing solubilized labeled membranes directly subjected to SDS-PAGE (a) or one representative autoradiogram of three independent experiments showing immunoprecipitated membranes with the subtype-specific G protein α subunit Golf antibody subjected to SDS-PAGE (b). c, histograms showing the quantification of 10 μM CGS 21680-induced increased labeling of immunoprecipitated Golf subunits in percent of control at various GDP concentrations (n= 3) (Error bars, S.E.M.).
To determine whether the agonist-induced enhancement of [α-32P]m-AcAGTP labeling of Golf was dose-dependent, photolabeling experiments were performed using different concentrations of the adenosine A2A receptor agonist CGS 21680 in the presence of a high concentration of GDP (100 μM). The immunoprecipitated samples were visualized by SDS/PAGE followed by X-ray autoradiography. As demonstrated by the representative autoradiogram in Fig. 5a, incorporation of [α-32P]m-AcAGTP into Golf protein increased in a concentration-dependent manner. The intensity of the labeling was used to construct a dose-response curve (Fig. 5b). The potency of CGS 21680 in this assay was estimated to 16 nM (4.7–54; 95% confidence interval). The agonist induced increase of labeling was blocked by the specific adenosine A2A receptor antagonist SCH 58261 (30 nM) (data not shown).

Concentration dependent increase of Golfphotolabeling by CGS 21680. Membranes from striatum were stimulated in the presence of 100 μM GDP with increasing concentrations of CGS 21680 (1 nM to 10 μM). Golf was immunoprecipitated with the subtype-specific G protein α subunit Golf antibody and subjected to SDS-PAGE. The autoradiogram (a) showing the 45-kDa region is representative of three independent experiments. b, dose-response curve of CGS 21680 induced incorporation of [α32P]m-AcAGTP as percent of maximal incorporation (n = 3) (Error bars, S.E.M.).
cAMP Accumulation Experiments.
The adenosine A2A agonist CGS 21680 induced a concentration-dependent increase in cAMP accumulation both in the control cells and in the Golf cDNA transfected CHO cells (Fig. 6). There was no significant change in the potency of CGS 21680: 19 nM (13–27) for Golf transfected cells and 29 nM (16–52) for control cells. However, there was a significant increase (ttest P = .0082) in the efficacy of the agonist. The plateau was 2.8 (2.6–3.0) pmol/50 μl for Golftransfected cells and 2.2 (1.9–2.4) pmol/50 μl for control cells (95% confidence intervals within parentheses).

Dose-response curves of CGS 21680 induced cAMP accumulation in CHO cells stably expressing (▴) adenosine A2A receptors and CHO cells transiently transfected (▪) with Golf cDNA and expressing adenosine A2Areceptors. Results are mean of two experiments with duplicates and are presented as picomoles of cAMP per 50 μl (Error bars, S.E.M.). The plateau is significantly increased in Golf transfected cells compared with nontransfected cells (t testP = .0082).
Discussion
It has been taken for granted that adenosine A2A receptors are coupled to the stimulatory G protein subunit Gs. The present data suggest that this may not be the complete truth, at least not in dopamine-rich areas of the brain.
Some previous reports suggest that Gs plays a minor role in striatal neurons, which contain the highest levels of A2A receptors in the brain. Although the highest levels of activated adenylyl cyclase, measured using [3H]forskolin binding, are found in the cautate putamen (Worley et al., 1986), this region possesses the lowest levels of Gs mRNA in brain (Largent et al., 1988) (see also Figs. 1c, 2b, and 3). It can also be mentioned that Albright and McCune-Albright syndromes, which are associated with mutations of the Gs gene, do not exhibit any extrapyramidal neurological symptoms (Weinstein et al., 1990; Schwindinger et al., 1992).
Herve et al. (1993) have shown that selective lesioning of striatonigral neurons, using a retrograde neurotoxin, markedly decreases the levels of dopamine D1 receptors and Golf in striatum. Moreover, 6-hydroxydopamine lesioning of the dopaminergic axons in neonatal rats induces hypersensitivity to dopamine receptor agonists in adulthood without any change in D1 receptor binding—but there is an increase in Golf expression. There is also an increase in Gs protein, but mainly in glial cells (Penit-Soria et al., 1997). Furthermore, mice deficient in Golf are hyperactive (Belluscio et al., 1998). These results could indicate that Golf might, in striatum, play some of the role(s) otherwise attributed to Gs.
Here we confirm that mRNAs for the adenosine A2Areceptor (Svenningsson et al., 1997) and for Golf(Drinnan et al., 1991; Herve et al., 1993) were most abundant in the caudate putamen, the nucleus accumbens, and the olfactory tubercle, whereas these areas express little Gs (Largent et al., 1988). We also show directly that adenosine A2A receptors and Golf are colocalized. In situ hybridization experiments showed that approximately 50% of the medium-sized neuron-like cells exhibited a strong signal for adenosine A2A receptor mRNA and a large majority (95%) of these neurons coexpressed Golf mRNA. Golf mRNA was also abundant in A2A receptor mRNA negative neurons. Thus, the relative enrichment of A2Areceptors and Golf mRNA in the striatum is likely to reflect the preferential localization in the striatopallidal neurons (A2A receptors) and in striatonigral and striatopallidal neurons (Golf). The labeling for Gs mRNA in striatum was weak and seemed to be preferentially located in glial cells, as described previously (Feinstein et al., 1992). In the present study, only 3 to 5% of the neurons in caudate putamen and 13 to 21% of the neurons in nucleus accumbens expressed both A2A receptor mRNA and Gs mRNA.
Using a photoaffinity labeling method, we show that the adenosine A2A receptor is not only colocalized with Golf in striatum, but also functionally coupled to it. Photoaffinity labeling is a method used for identification of distinct molecular components in biochemical processes. The simplest procedure for labeling G proteins is by UV irradiation of [α-32P]GTP (Basu and Modak, 1987). However, this method has low sensitivity. The most widely used photoaffinity label for G proteins is 4-azidoanilido-GTP and was developed byPfeuffer (1977). However, this compound has some disadvantages: it is unstable and its synthesis, purification, and application must be performed in complete darkness. The photoaffinity label we have used,m-acetylanilido-GTP, developed by Zor and coworkers (1995), has several advantages: 1) no need for purification; 2) quantitative conversion of GTP into m-AcAGTP; 3) excellent stability in solution; and 4) resistant to hydrolysis and remains bound to the G protein during centrifugal washing. Using this photolabeling method, we could demonstrate that the adenosine A2A agonist CGS 21680 induced an increased incorporation of [α-32P]m-AcAGTP in immunoprecipitated Golf subunits. The magnitude of the increase was GDP-concentration-dependent, with the largest increase seen at a high GDP concentration (100 μM). CGS 21680 increased labeling in a dose-dependent manner with a calculated EC50 value of 16 nM (4.7–54). This increase was receptor mediated because it was blocked by the A2A receptor antagonist SCH 58261 (data not shown). The CGS 21680 potency is in agreement with the potency in the cAMP assay.
In an attempt to determine whether A2Areceptor-activated Golf subunits could activate adenylyl cyclase, we transiently transfected Golfinto CHO cells stably expressing A2A receptors (Kull et al., 1999). In the Golf transfected cells, there was a significant increase in the maximal agonist stimulated adenylyl cyclase activity without any change in the potency of CGS 21680. A change in the maximal effect was expected from the previous demonstration of increased constitutive accumulation of cAMP and the observation that agonist-stimulated cAMP levels were proportional to the amount of Gs expression (Yang et al., 1997).
Conclusion
The present study shows that adenosine A2Areceptor mRNA was coexpressed with Golf mRNA in striatal medium-sized neurons to a much higher extent than with Gs mRNA. Using a photolabeling technique, we show that activation of adenosine A2A receptors in striatal membranes led to activation of Golf. Moreover, transfection of Golf cDNA into cells that express human adenosine A2A receptors led to an increase of the maximal agonist stimulated cAMP level. When taken together, these results provide strong anatomical and biochemical evidence that adenosine A2A receptors stimulate adenylyl cyclase and cAMP-dependent signal transduction in striatum by activating Golf rather than Gs. These findings have the more general implication that a given receptor may couple to different G proteins in different locations.
Footnotes
- Received December 29, 1999.
- Accepted July 26, 2000.
-
Send reprint requests to: Dr. Björn Kull, Department of Physiology and Pharmacology, Section of Molecular Neuropharmacology, Karolinska Institutet, S-171 77 Stockholm, Sweden. E-mail:bjorn.kull{at}fyfa.ki.se
-
These studies were supported by the Swedish Medical Research Council (proj no. 2553), by Knut and Alice Wallenberg's Foundation, and Karolinska Institutet.
Abbreviations
- PCR
- polymerase chain reaction
- SSC
- saline sodium citrate
- DTT
- dithiothreitol
- m-AcAGTP
- m-acetylanilido-GTP
- PAGE
- polyacrylamide gel electrophoresis
- CHO-K1
- Chinese hamster ovary cells, strain K1
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}