


Abstract
Classical chemoattractant receptors are of fundamental importance to immune responses. The two major roles of such receptors are the modulation of chemotaxis and the generation of reactive oxygen species. The formyl peptide receptor-like 1 (FPRL1) can be stimulated by two different ligands, Trp-Lys-Tyr-Met-Val-Met-CONH2 (WKYMVM) and lipoxin A4 (LXA4). Although leukocyte chemotaxis mediated by activated FPRL1 has been reported, the role of FPRL1 in superoxide generation remains to be studied. In this study, we examined the effect of WKYMVM or LXA4 on chemotactic migration and superoxide generation in human neutrophils. WKYMVM and LXA4 stimulated neutrophil chemotaxis via tyrosine phosphorylation events. In terms of reactive oxygen species generation, WKYMVM but not LXA4 stimulated superoxide generation in neutrophils. To understand this difference on superoxide generation via the same receptor, FPRL1, we compared the signaling pathways downstream of FPRL1 by the two different ligands. At first, we confirmed that both WKYMVM and LXA4 caused intracellular calcium ([Ca2+]i) increase in a pertussis toxin-sensitive manner and that these ligands competitively inhibited each other with respect to [Ca2+]i increase in neutrophils. This result suggests that WKYMVM and LXA4 share the same receptor, FPRL1. By investigating cellular signaling by WKYMVM and LXA4, we found that WKYMVM but not LXA4 induced extracellular signal-regulated protein kinases (ERKs), c-Jun NH2-terminal kinase, and phospholipase A2 (PLA2) activation. We also found that ERK-mediated cytosolic PLA2 activity is essential for superoxide generation. These results indicate that the activation of FPRL1 by the two different ligands can induce differential cellular signaling and unique functional consequences in human neutrophils.
Neutrophils perform a critical role in innate immune responses, including extravasation from the peripheral blood stream, migration into an infected area, and the generation of reactive oxygen species such as superoxide (Baggiolini et al., 1993; Bokoch, 1995). Moreover, the activities of neutrophils involved in these roles are modulated by many diverse molecules. Among these, chemoattractants including classic chemoattractants [such as formyl-methionyl-leucyl-phenylalanine (fMLF), complement component C5a, and leukotriene B4] and several chemokines stimulate leukocytes via the activation of pertussis toxin (PTX)-sensitive G protein-coupled receptor (GPCR) (Baggiolini et al., 1993; Bokoch, 1995). The ligation of the receptors by appropriate chemoattractants initiates the dissociation of the Gαi subunit from the Gβγ subunit complex, which in turn induces downstream signaling by activating effector molecules. This signaling in turn results in cytoskeletal rearrangements, exocytosis, histamine release, receptor induction, adhesion, the production of bioactive lipids, and the activation of the respiratory burst system via NADPH oxidase activation (Bokoch, 1995; Wu et al., 2000).
Because these chemoattractants have important roles in the modulation of host immunity, their molecular interactions with receptors and the subsequently induced cellular signaling pathways have received much attention (Le et al., 2001b). The formyl peptide receptor (FPR) system is one of the most extensively studied chemoattractant receptor systems. When fMLF binds to FPR, FPR transmits a signal to the heterotrimeric G proteins, which induces the activation of phospholipase C (PLC) and phophoinositide-3-kinase (PI3K) (Jiang et al., 1996; Pan et al., 2000). PLC then hydrolyzes phosphatidylinositol 4,5-bisphosphate into phosphatidylinositol 1,4,5-triphosphate and diacylglycerol, which induce intracellular calcium ([Ca2+]i) increase and protein kinase C activation, respectively (Rhee and Bae, 1997). Moreover, PI3K converts phosphatidylinositol 4,5-bisphosphate into phosphatidylinositol 1,4,5-triphosphate, which is essential for modulating cellular enzymes, including 3-phosphoinositide–dependent protein kinase 1 (Shepherd et al., 1998). The downstream signaling initiated by fMLF binding also induces other signaling cascades, which include mitogen-activated protein kinases (MAPKs), phospholipase A2 (PLA2), and phospholipase D (PLD) activation (Marshall et al., 2000; Bechoua and Daniel, 2001).
FPR and two homologs of FPR have been identified in humans: formyl peptide receptor-like 1 (FPRL1) and formyl peptide receptor-like 2 (FPRL2) (Le et al., 2001b). Recently, several different agonists have been identified for FPR and FPRL1 (Le et al., 2001b). One of the natural ligands for FPRL1 is lipoxin A4 (LXA4), an eicosanoid (Maddox et al., 1997). Although FPRL1 is important in the modulation of inflammatory responses, by mediating the recruitment of leukocytic cells into an infected area, its downstream signaling pathway has not been examined. Previously, we reported on a synthetic peptide, Trp-Lys-Tyr-Met-Val-Met-CONH2 (WKYMVM), that can stimulate leukocyte activity (Baek et al., 1996). WKYMVM was found to stimulate phosphoinositide hydrolysis and extracellular signal-regulated protein kinase (ERK) activation in U937 cells, and these events proved to be PTX-sensitive, suggesting the involvement of PTX-sensitive G protein (Baek et al., 1996, 1999). Recently Christophe et al. (2001) demonstrated that WKYMVM is also a ligand for FPRL1. In this study, we compared the signaling pathways and functional consequences downstream of FPRL1 ligation by WKYMVM and LXA4 in human neutrophils.
Materials and Methods
Materials. WKYMVM was synthesized and purified in the Peptide Library Support Facility at Pohang University of Science and Technology, as described previously (Baek et al., 1996). RPMI 1640 was purchased from Invitrogen (Carlsbad, CA). Amyloid β peptide 1–42 (Aβ42) was purchased from Bachem Bioscience (King of Prussia, PA). Human interleukin-8 (IL-8) was obtained from Genzyme (Cambridge, MA). Dialyzed fetal bovine serum and supplemented bovine calf serum were from Hyclone Laboratories (Logan, UT). Pertussis toxin and [9,10(n)-3H]myristic acid (53 Ci/mmol) were from Amersham Biosciences UK, Ltd. (Little Chalfont, Buckinghamshire, UK). [5,6,8,9,11,12,14,15-3H]arachidonic acid (AA) (100 Ci/mmol) was from PerkinElmer Life Sciences (Boston, MA). Precoated silica gel TLC plates (F-254) were from Merck (Darmstadt, Germany); GF109203X, Ro-31–8220, LY294002, and genistein were from Calbiochem (San Diego, CA); 5(S),6(R),15(S)-trihydroxyeicosa-7E,9E,11Z,13E-tetraenoic acid (lipoxin A4), methyl arachidonylfluorophosphonate (MAFP), arachidonyltrifluoromethyl ketone (AACOCF3), and bromoenol lactone (BEL) were from BIOMOL Research Laboratories (Plymouth Meeting, PA); rabbit anti-human antibodies to ERK2, phospho-ERKs, JNK, phospho-JNK, and phospho-paxillin (Y118) were from Cell Signaling Technology Inc. (Beverly, MA), and horseradish peroxidase-conjugated antibodies to mouse and rabbit IgG were purchased from Kirkegaard and Perry Laboratories (Gaithersburg, MD).
Isolation of Human Neutrophils. Peripheral blood was collected from healthy donors, and human neutrophils were isolated by dextran sedimentation, hypotonic lysis of erythrocytes, and by using a lymphocyte separation medium gradient as described previously (Bae et al., 2001). Isolated human neutrophils were used promptly.
Cell Culture. RBL-2H3 cells and FPRL1-expressing RBL-2H3 cells were cultured in Dulbecco's modified Eagle's medium supplemented with 20% FBS and 200 μg/ml of G418, as described previously (He et al., 2000).
Measurement of [Ca2+]i. The level of [Ca2+]i was determined by Grynkiewicz's method using fura-2/AM (Bae et al., 2001). Briefly, prepared cells were incubated with 3 μM of fura-2/AM at 37°C for 50 min in fresh serum-free RPMI 1640 medium with continuous stirring. Cells (2 × 106) were aliquoted for each assay into Ca2+-free Locke solution (154 mM NaCl, 5.6 mM KCl, 1.2 mM MgCl2, 5 mM HEPES, pH 7.3, 10 mM glucose, and 0.2 mM EGTA). Fluorescence changes at two excitation wavelengths (340 and 380 nm) and an emission wavelength of 500 nm were measured, and the fluorescence ratio obtained was translated into [Ca2+]i.
Chemotaxis Assay. Chemotaxis assays were performed using multiwell chambers (Neuroprobe Inc., Gaithersburg, MD) (Bae et al., 2001). Briefly, prepared human neutrophils were suspended in RPMI 1640 medium at a concentration of 1 × 106 cells/ml, and 25 μl of this suspension was placed on the upper well of a chamber onto a 3-μm polyhydrocarbon filter, which separated the suspension from the WKYMVM- or LXA4-containing lower well. After incubation for 2 h at 37°C, nonmigrated cells were removed by scraping. Cells that migrated across the filter were dehydrated, fixed, and stained with hematoxylin (Sigma, St. Louis, MO) and were counted in five randomly chosen high-power fields (400×).
Stimulation of Cells with WKYMVM or LXA4. Prepared cells were aliquoted into 2 × 106 cells and stimulated with the indicated concentrations of WKYMVM or LXA4 for predetermined periods. After stimulation, the cells were washed with serum-free RPMI 1640 medium and lysed in lysis buffer (20 mM HEPES, pH 7.2, 10% glycerol, 150 mM NaCl, 1% Triton X-100, 50 mM NaF, 1 mM Na3VO4, 10 μg/ml of leupeptin, 10 μg/ml of aprotinin, and 1 mM phenylmethylsulfonyl fluoride). The detergent-insoluble material was pelleted by centrifugation (12,000g for 15 min at 4°C), and the soluble supernatant fraction was removed and stored at –80°C or used immediately. Protein concentrations in the lysates were determined using Bradford protein assay reagent.
Electrophoresis and Immunoblot Analysis. Protein samples were prepared for electrophoresis by adding concentrated Laemmli sample buffer and separated using an 8% SDS-polyacrylamide gel and a buffer system described previously (Bae et al., 2000). After electrophoresis, the proteins were blotted onto a nitrocellulose membrane, which was then blocked by incubation with Tris-buffered saline/0.05% Tween 20 containing 5% nonfat dry milk. The membranes were then incubated with anti–phospho-ERK antibody, anti-ERK antibody, anti–phospho-JNK antibody, or anti-JNK antibody and washed with Tris-buffered saline. Antigen-antibody complexes were visualized after incubating the membrane with 1:5,000 diluted goat anti-rabbit IgG or goat anti-mouse IgG antibody coupled to horseradish peroxidase by using the enhanced chemiluminescence detection system.
Measurement of Superoxide Anion Generation. Superoxide anion generation was determined by measuring cytochrome c reduction using a microtiter 96-well plate enzyme-linked immunosorbent assay reader (EL312e; Bio-Tek Instruments, Winooski, VT) as described previously (Bae et al., 2001). Human neutrophils (2 × 106 cells in RPMI 1640 medium) were preincubated with 50 μM of cytochrome c at 37°C for 1 min and then incubated with 100 nM of WKYMVM. To study the roles of PLA2 and PLD in peptide-induced superoxide generation, aliquots of cells were pretreated with 20 μM of the PLA2 inhibitor butan-1-ol or butan-3-ol before peptide stimulation. Superoxide generation was determined by measuring changes of light absorption at 550 nm over 5 min at 1-min intervals.
Measurement of Phosphatidylbutanol Formation in Human Neutrophils. Phosphatidylbutanol (PBt) production was determined as described in an earlier report (Bae et al., 2000) with a slight modification. Briefly, human neutrophils were resuspended to 1 × 106 cells/ml in RPMI 1640 medium containing 2.5% FBS and loaded with [3H]myristic acid (5 μCi/ml) for 90 min at 37°C. The loaded neutrophils were then washed twice with serum-free RPMI 1640 medium and stimulated with WKYMVM in the presence of 0.5% butan-1-ol. After 30 min, the reaction was quenched by adding 0.5 ml of ice-cold methanol, and the medium was aspirated. Chloroform (1 ml) and 0.5 ml of 1 M NaCl were then added to the medium, and total lipids were extracted by vigorous vortexing. The lower chloroform phase was obtained by centrifuging at 550g for 10 min and dried under nitrogen. The lipids were then dissolved in chloroform-methanol (95:5), spotted onto silica gel 60 TLC plates, and separated using a solvent containing chloroform-methanol-acetic acid (90:10:10), as described previously (Bae et al., 2000). The amounts of PBt and total lipid were determined using a Fuji BAS-2000 image analyzer (Fuji Photo Film Co., Ltd., Tokyo, Japan).
Measurement of PLA2 Activity in Cells. Isolated human neutrophils (107 cells/ml) were prelabeled with 0.5 μCi/ml of [3H]AA in RPMI 1640 medium containing 10% FBS at 37°C for 24 h in a humidified incubator supplied with 95% air and 5% CO2, as described previously (Bae et al., 2000). The labeled cells were then washed twice with serum-free RPMI 1640 and incubated in RPMI 1640 medium containing 0.1% fatty acid-free BSA for 15 min at 37°C. After discarding the medium, the cells were stimulated with various concentrations of WKYMVM for the indicated times. The radioactivities of the medium and of the collected cells were measured using a liquid scintillation counter. When investigating the effects of inhibitors on the WKYMVM-stimulated PLA2 activity, the cells were preincubated with the indicated concentrations of each inhibitor, or vehicle, for 15 min before stimulation.
Results
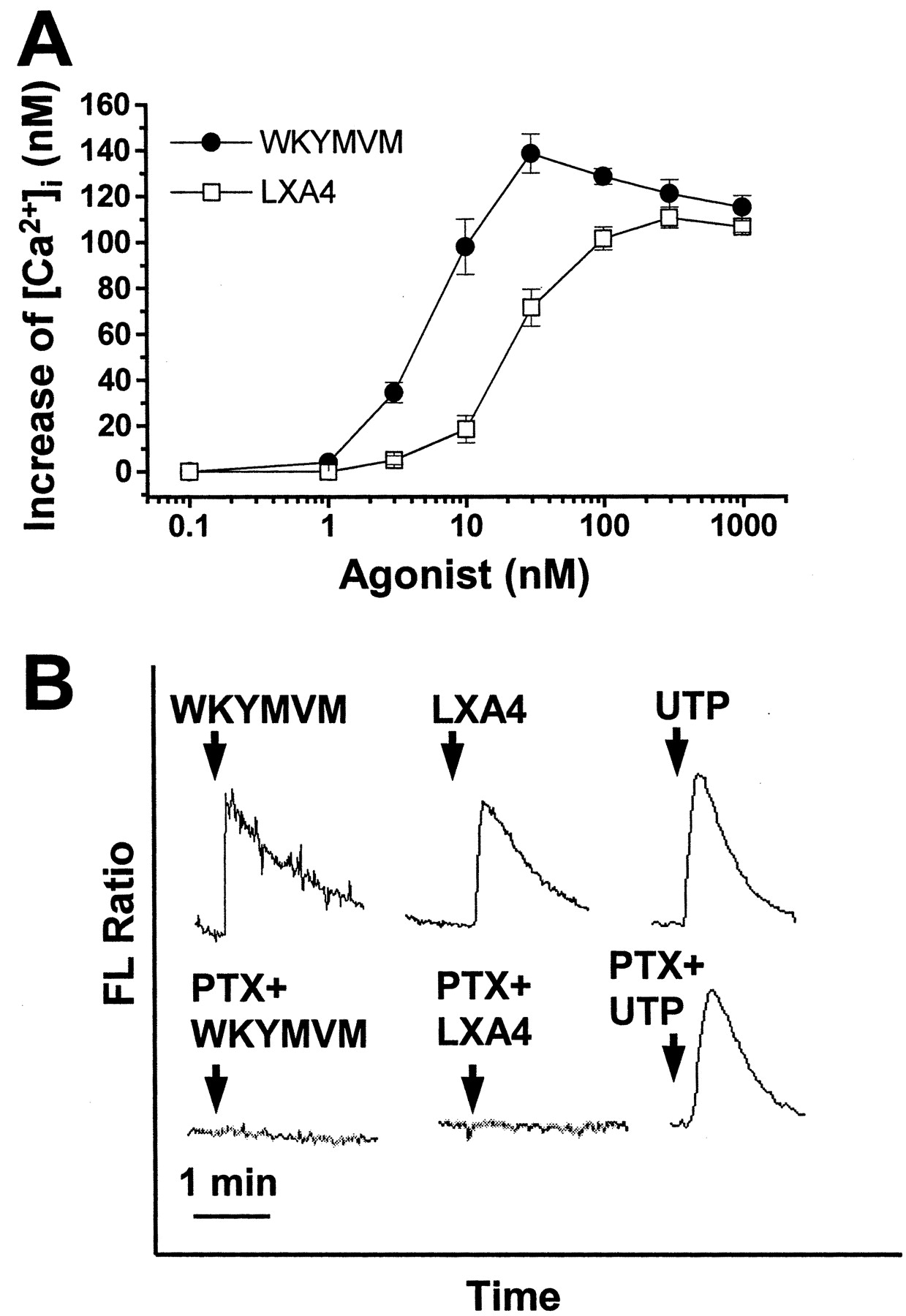
WKYMVM and LXA4 Stimulate [Ca2+]i Increase in a PTX-Sensitive Manner in Human Neutrophils. Many chemoattractants that bind to specific GPCRs induce phosphoinositide hydrolysis and [Ca2+]i increase (Bokoch, 1995; Jiang et al., 1996; Le et al., 2001b). In this study, we examined the effects of WKYMVM and LXA4 on [Ca2+]i changes in human neutrophils. Fura-2–loaded human neutrophils were stimulated with various concentrations of WKYMVM or LXA4. As shown in Fig. 1A, WKYMVM or LXA4 caused [Ca2+]i increase in a concentration-dependent manner, showing maximal activity at 30 and 100 nM, respectively. The effect of WKYMVM or LXA4 on calcium activity was consistently observed with different donors. In a parallel experiment, we also verified the effect of WKYMVM or LXA4 on [Ca2+]i increase in HL60 (dHL60) cells differentiated by DMSO. Both WKYMVM and LXA4 were found to stimulate [Ca2+]i increase in the dHL60 cells with similar potencies as in neutrophils (data not shown). This result further supports our observation that LXA4 stimulates human neutrophils to induce [Ca2+]i increase. The majority of chemoattractants have been found to bind to a family of receptors that are coupled to PTX-sensitive G proteins (Bokoch, 1995; Le et al., 2001b). Thus, we examined the effect of PTX on WKYMVM or LXA4-induced [Ca2+]i increase. When neutrophils were pre-incubated with 100 ng/ml of PTX for 24 h, WKYMVM- or LXA4-induced cytosolic calcium changes were completely inhibited (Fig. 1B). As a control experiment, we checked the effect of UTP, which stimulates myeloid cells in a PTX-insensitive manner, on [Ca2+]i increase in PTX-treated neutrophils (Alemany et al., 2000). We confirmed that UTP-induced [Ca2+]i increase was not affected by PTX treatment (Fig. 1B). These results indicate that both WKYMVM and LXA4 stimulate [Ca2+]i increase in human neutrophils and that these events occur in a PTX-sensitive G protein-dependent manner.

Both WKYMVM and LXA4 stimulate [Ca2+]i increase in human neutrophils. A, human neutrophils were loaded with 3 μM fura-2/AM for 50 min at 37°C in serum-free RPMI 1640 medium and washed three times with serum-free RPMI 1640. Before measurement, the cells (2 × 106) were suspended in Ca2+-free Locke solution containing 0.2 mM EGTA. Fura-2–loaded cells were stimulated with various concentrations of WKYMVM or LXA4. Maximum [Ca2+]i increases were noted. Data are presented as means ± S.E.M. of four independent experiments. B, isolated human neutrophils were preincubated in the absence or presence of 100 ng/ml of PTX for 24 h at 37°C in a 5% CO2 incubator. Cells were then loaded with fura-2, and the [Ca2+]i increases because of additions of 100 nM of WKYMVM, 1 μM of LXA4, or 100 μM UTP were measured. The charts shown are from a single experiment and are representative of three independent experiments. FL, fluorescence.
WKYMVM and LXA4 Stimulate Neutrophils via FPRL1. LXA4 has been reported to be a lipid agonist for FPRL1 (Maddox et al., 1997). Recently, it was also demonstrated that WKYMVM could activate FPRL1 but not FPR (Christophe et al., 2001). Because WKYMVM and LXA4 are specific agonists for FPRL1, we checked whether the two agonists competitively increase [Ca2+]i. As shown in Fig. 2A, the stimulation of neutrophils with WKYMVM caused the complete inhibition of LXA4-induced [Ca2+]i increase. Similarly, LXA4 pretreatment also completely inhibited WKYMVM-induced [Ca2+]i increase (Fig. 2A). These results suggest that WKYMVM and LXA4 share the same receptor in neutrophils and further suggest that WKYMVM and LXA4 stimulate human neutrophils via FPRL1. To confirm that WKYMVM and LXA4 act on the same receptor, FPRL1, on the surface of human neutrophils, we investigated the effect of Aβ42 on WKYMVM or LXA4 signaling. Aβ42 is a well-known FPRL1-specific agonist (Le et al., 2001a). When human neutrophils were pretreated with Aβ42, [Ca2+]i increase by WKYMVM- or LXA4 was completely inhibited (Fig. 2B). The result strongly supports our notion that WKYMVM and LXA4 act on FPRL1 in the cells. We also investigated the effect of WKYMVM or LXA4 on the [Ca2+]i increase by IL-8, a specific ligand of IL-8 receptor. Pretreatment of WKYMVM or LXA4 did not affect on the IL-8–induced [Ca2+]i increase in human neutrophils (Fig. 2C). This result suggests that inhibition of LXA4-induced signaling caused by WKYMVM pretreatment is mediated through specific desensitization of FPRL1 but not through other receptors.
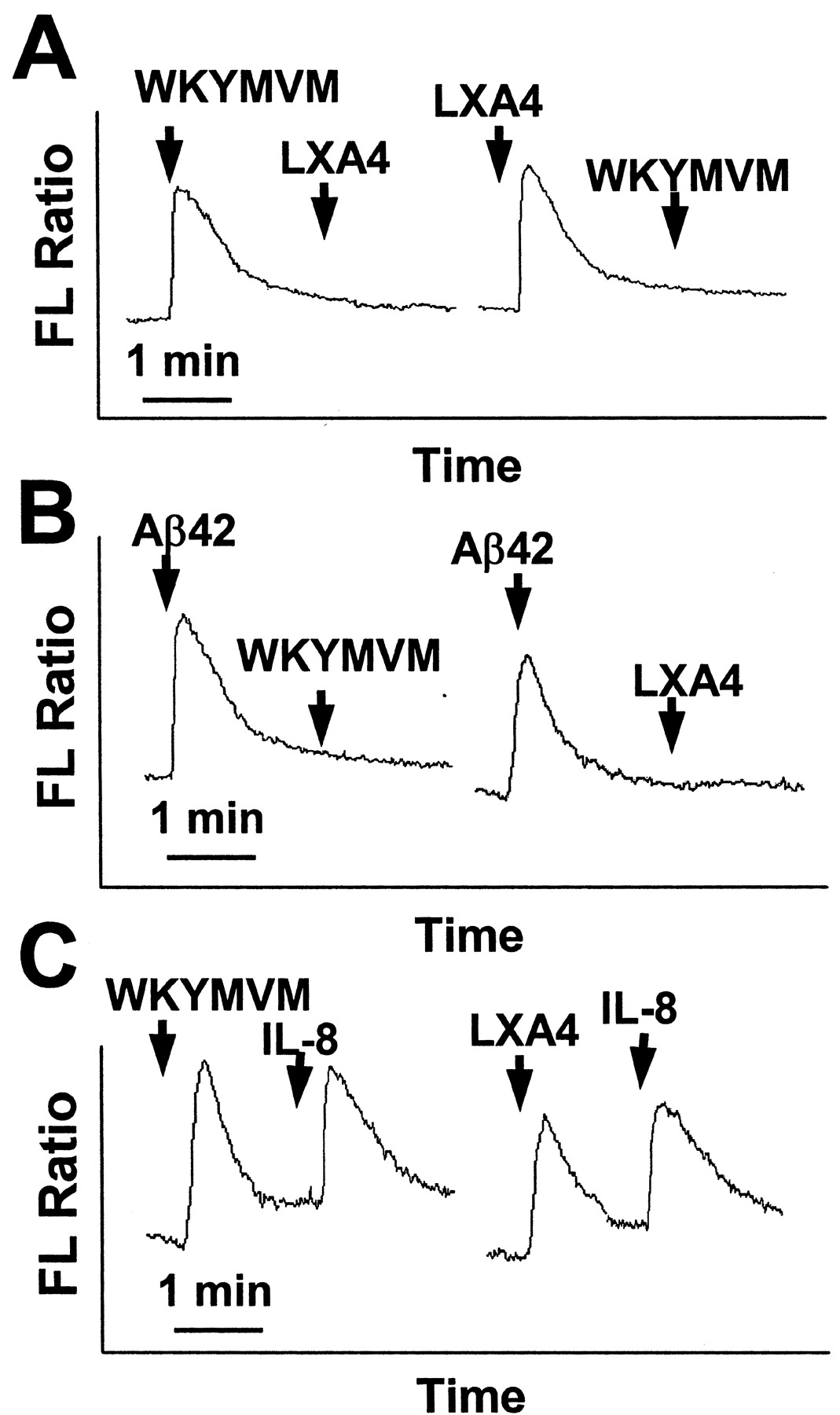
WKYMVM and LXA4 act on the same receptor, FPRL1, in human neutrophils. A, fluorescence (FL) ratios of fura-2–loaded human neutrophils were monitored before and during the sequential addition of agonists (100 nM WKYMVM or 1 μM LXA4) at the times indicated by the arrows. B, 20 μM Aβ42, 100 nM WKYMVM, or 1 μM LXA4 were used. C, 100 nM WKYMVM, 1 μM LXA4, or 1 ng/ml IL-8 were used. The charts shown are from a single experiment, which was representative of at least three independent experiments (A, B, and C).
WKYMVM and LXA4 Induce Neutrophil Chemotaxis via Tyrosine Phosphorylation. FPRL1 is one of the classic chemoattractant receptors (Le et al., 2001b). Because WKYMVM and LXA4 have been reported to be ligands of FPRL1, we examined the effect of WKYMVM or LXA4 on neutrophil chemotaxis. As shown in Fig. 3A, WKYMVM induced neutrophil chemotaxis within the concentration range of 1 to 500 nM; similarly, LXA4 also caused neutrophil chemotaxis in the range of 10 to 500 nM (Fig. 3A). WKYMVM induced neutrophil chemotaxis more potently than did LXA4. A number of reports have shown that several chemoattractants stimulate the tyrosine phosphorylation of cellular proteins and that tyrosine kinases are involved in the chemotaxis of leukocytes stimulated by these chemoattractants (Rodriguez-Frade et al., 1999; Wang et al., 2000). We examined the effects of WKYMVM or LXA4 at the tyrosine phosphorylation level in human neutrophils by Western blotting and by probing with anti–phospho-tyrosine antibodies. When cells were stimulated with either of these agonists, a rapid increase in the phospho-tyrosine of several proteins, including a 70-kDa phosphorylated protein, was observed. These proteins were rapidly tyrosyl-phosphorylated (in less than 5 min) and dephosphorylated 30 min after stimulation with WKYMVM or LXA4 (data not shown). We examined the identity of p70 by Western blotting with anti-phospho (Y118) paxillin antibody. As shown in Fig. 3B, stimulation of the cells with 100 nM of WKYMVM or with 1 μM of LXA4 caused rapid tyrosine phosphorylation of paxillin. This suggests that the occupation of FPRL1 elicits the activation of tyrosine kinases, such as p125 FAK, thus resulting in the phosphorylation of paxillin. Next, we decided to investigate whether tyrosine kinase activity is required for WKYMVM- or LXA4-induced neutrophil chemotaxis. Preincubation of human neutrophils with various concentrations of genistein for 15 min at 37°C, before stimulation with WKYMVM or LXA4, did indeed affect neutrophil chemotaxis in a concentration-dependent manner (Fig. 3C). These results indicate that WKYMVM and LXA4 elicit the tyrosyl-phosphorylation of several proteins and that this signaling is required for the chemotaxis of human neutrophils.

WKYMVM- and LXA4-induced neutrophil chemotaxis. Assays were performed using a modified Boyden chamber assay, as described under Materials and Methods. Various concentrations of WKYMVM or LXA4 were loaded into the lower chambers. A, isolated human neutrophils (1 × 106 cells/ml of serum-free RPMI 1640 medium) were added to the upper wells of a 96-well chemotaxis chamber and allowed to migrate across a polycarbonate membrane with a pore size of 3 μm; migration was assessed after 2 h of incubation at 37°C. B, human neutrophils were stimulated with 100 nM of WKYMVM or with 1 μM of LXA4 for the indicated lengths of time. Each sample (30 μg of protein) was subjected to 8% SDS-PAGE, and phosphorylated paxillin was determined by immunoblot analysis with anti-phospho (Y118) paxillin antibody. The shown digits are the quantitative result of paxillin phosphorylation. Immunoblot analysis with anti-actin antibody was conducted to confirm that the same amounts of protein were loaded. The results shown are representative of three independent experiments. C, to test the effect of tyrosine kinase on WKYMVM- or LXA4-induced neutrophil chemotaxis, prepared human neutrophils were exposed to 100 nM of WKYMVM or 300 nM of LXA4 in the absence or in the presence of various concentrations of genistein. The numbers of cells that had migrated were determined by counting in a high-power field (400×). Data are presented as means ± S.E.M. of three independent experiments, each performed in duplicate (A and C).
WKYMVM but Not LXA4 Stimulates Superoxide Generation. One of the major physiological functions of classic chemoattractants, including fMLF, is the production of reactive oxygen species such as superoxide (Baggiolini et al., 1993; Bokoch, 1995). In this study, we compared the effects of WKYMVM and LXA4 on superoxide generation in neutrophils. WKYMVM (100 nM) caused superoxide generation in human neutrophils and was similar to 1 μM of fMLF in this respect (Fig. 4). LXA4, however, did not affect superoxide generation at the 1-μM level (Fig. 4). We also confirmed that superoxide was not generated at concentrations of LXA4 in the range of 1 to 500 nM (data not shown). These results indicate that WKYMVM induces chemotaxis and superoxide generation and that LXA4 only stimulates chemotaxis but not superoxide generation in human neutrophils.

Effects of WKYMVM and LXA4 on superoxide generation in neutrophils. Isolated human neutrophils (2 × 106 cells/100 μl/assay) were preincubated for 1 min at 37°C with 50 μM of cytochrome c before being stimulated with 100 nM WKYMVM, 1 μM LXA4, or 1 μM fMLF. Cytochrome c reduction was monitored as a change in absorption at 550 nm at 1-min intervals over 5 min, as described under Materials and Methods.
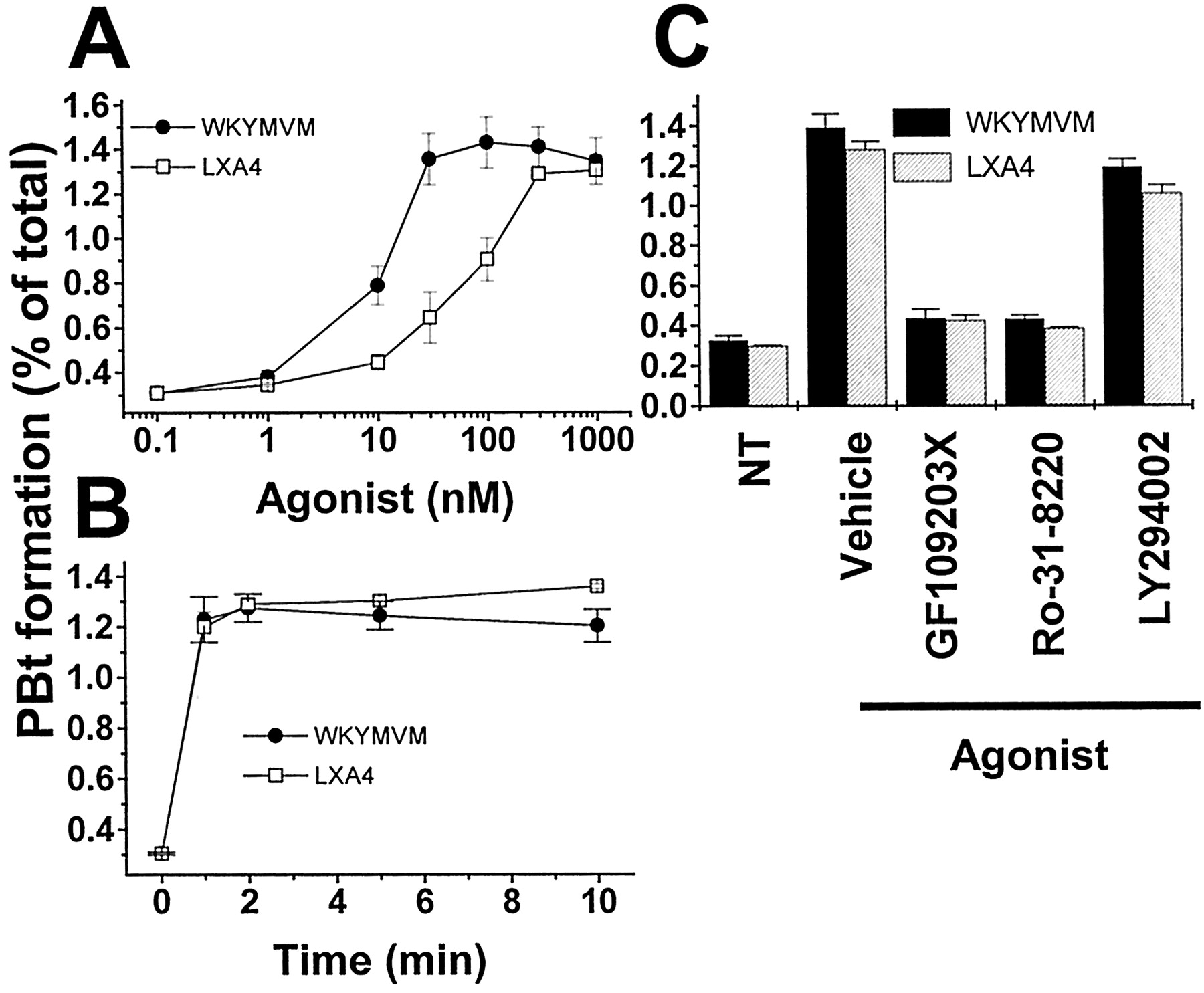
WKYMVM and LXA4 Stimulate PLD Activity. Phosphatidic acid, a product of PLD, has been suggested to play a critical role in the activation of the NADPH oxidase complex, resulting in superoxide generation (Perry et al., 1992; Bae et al., 2000). Figure 4 shows that superoxide generation was induced by WKYMVM but not by LXA4. One of the possible reasons for this differential effect concerns the activation status of PLD. We examined the effect of WKYMVM or LXA4 on PLD activity in neutrophils by measuring PBt formation in the presence of butan-1-ol. WKYMVM treatment caused PBt formation in a concentration-dependent manner, showing maximal activity at 100 nM (Fig. 5A). Stimulation of neutrophils with 100 nM of WKYMVM elicited transient PBt formation (Fig. 5B). Neutrophils treated with 30 to 1,000 nM of LXA4 showed similar transient PBt formation (Fig. 5, A and B). In terms of the signaling pathways of WKYMVM- and LXA4-induced PLD activation, it was observed that PLD activation by WKYMVM or LXA4 was sensitive to GF109203X and Ro-31–8220, which suggests the involvement of PKC in this process (Fig. 5C).

Effects of WKYMVM and LXA4 on PBt formation in neutrophils. Isolated human neutrophils were labeled with [3H]myristic acid for 90 min at 37°C in RPMI 1640 medium with 10% FBS. After three washes with phosphate-buffered saline, the labeled cells were stimulated with various concentrations of WKYMVM or LXA4 in the presence of 0.5% butan-1-ol for 10 min (A) or with 100 nM WKYMVM or 1 μM LXA4 in the presence of 0.5% butan-1-ol for various times (B). C, the neutrophils were suspended in RPMI 1640 medium and incubated for 15 min in the presence or absence of 5 μM GF109203X, 5 μM Ro-31–8220, and 50 μM LY294002 at 37°C and then stimulated for 10 min with 100 nM WKYMVM or 1 μM LXA4. PBt was resolved from total lipids by thin-layer chromatography. A Fuji BAS-2000 instrument was used to quantify each lipid. Data are presented as means ± S.E.M. of three separate experiments.
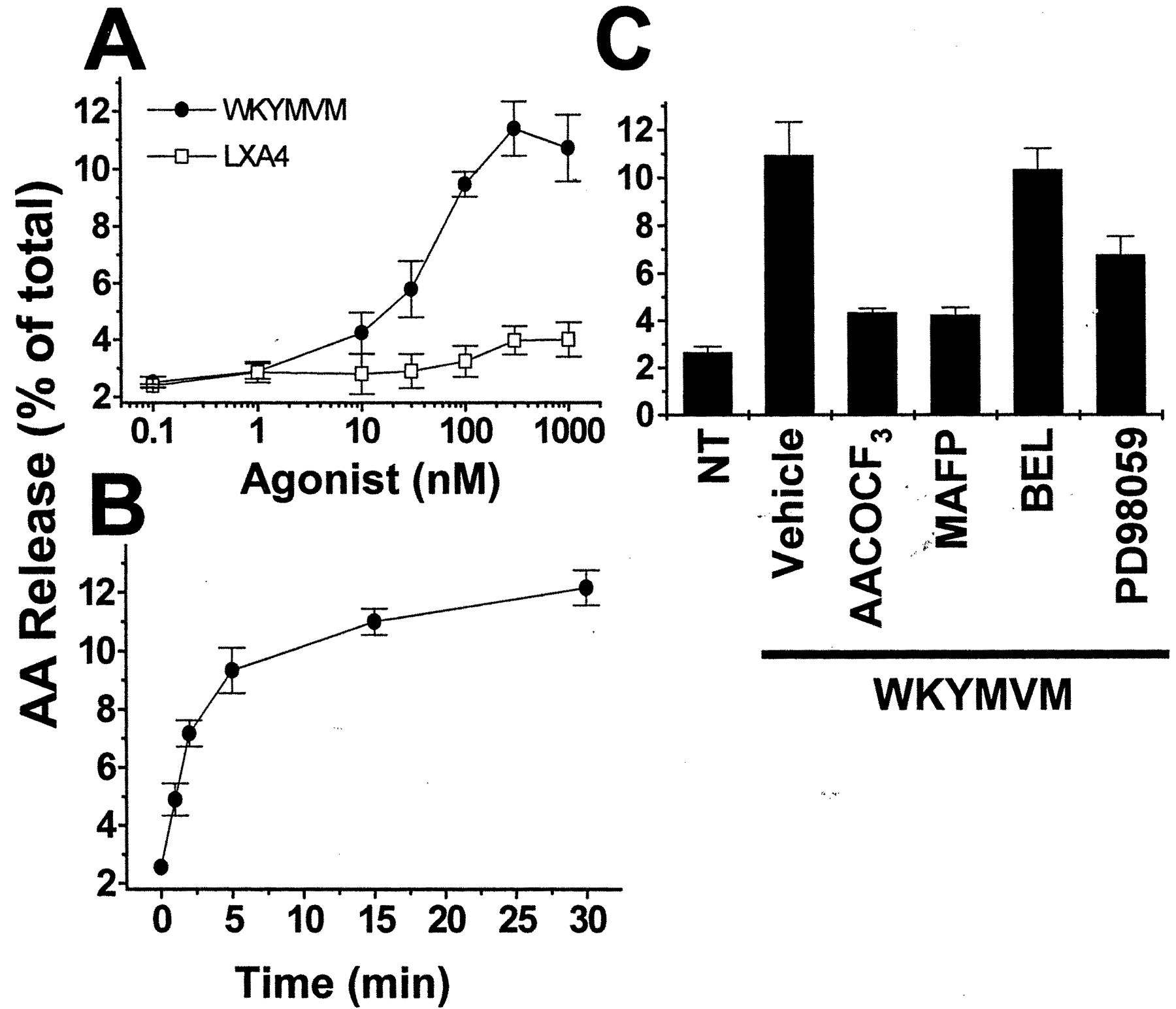
WKYMVM but Not LXA4 Stimulates AA Release. PLA2 activation is regarded as an important requirement for the activation of the superoxide-generating machinery (Dana et al., 1998; Bae et al., 2000). We investigated the effect of WKYMVM and LXA4 on PLA2 activity by measuring AA release in neutrophils. As shown in Fig. 6A, WKYMVM at 10 to 1,000 nM stimulated AA release, which leveled off at approximately 5 min (Fig. 6B); however, LXA did not affect AA release significantly at concentrations lower than 1 μM. This is a very interesting result in view of the fact that LXA4 stimulated [Ca2+]i increase and PLD activation in neutrophils. To identify the isozyme of PLA2 activated by WKYMVM, we examined the effects of different isozyme-specific PLA2 inhibitors on WKYMVM-induced AA release. Pretreatment with AACOCF3 (an inhibitor for cPLA2) or with MAFP (an inhibitor for cPLA2 and iPLA2) before the WKYMVM stimulation of neutrophils significantly blocked the WKYMVM-induced AA release (Fig. 6C), whereas preincubation of the cells with BEL (an inhibitor for iPLA2) did not affect WKYMVM-stimulated AA release (Fig. 6C). This result implies that WKYMVM stimulates cPLA2 but not iPLA2 activity in neutrophils. A previous report has demonstrated that ERK activity is important for cPLA2 phosphorylation and activation (Gijon and Leslie, 1999). We examined the role of ERK on WKYMVM-induced cPLA2 activation using PD98059, an inhibitor of MAPK kinase. Pretreatment of neutrophils with PD98059 before WKYMVM stimulation elicited the partial inhibition of WKYMVM-induced AA release, indicating the participation of mitogen-activated protein kinase kinase-dependent ERK activity (Fig. 6C).

Effect of WKYMVM and LXA4 on AA release in neutrophils. Human neutrophils preloaded with [3H]AA were stimulated with various concentrations of WKYMVM or 1 μM LXA4 for 30 min (A) and for various periods of time with 100 nM of WKYMVM or 1 μM of LXA4 (B) in the presence of 0.1% fatty acid-free BSA at 37°C. C, the neutrophils were suspended in Hanks' balanced salt solution containing 0.1% fatty acid-free BSA; incubated with MAFP (20 μM for 15 min), AACOCF3 (20 μM for 15 min), BEL (20 μM for 15 min), or PD98059 (50 μM for 60 min) at 37°C; and stimulated for 30 min with 100 nM WKYMVM or vehicle. The neutrophils were then suspended in Hanks' balanced salt solution containing 0.1% fatty acid-free BSA, and stimulated for the predetermined times with the indicated concentrations of WKYMVM, LXA4, or vehicle. Release of [3H]AA into the extracellular medium was determined using a liquid scintillation counter. Results are expressed as percentages of total cellular radioactivity and are presented as mean values ± S.E.M. (n = 6).
WKYMVM but Not LXA4 Stimulates Mitogen-Activated Protein Kinases Activities. The results that WKYMVM, but not LXA4, stimulated cPLA2 activation in a PD98059-sensitive manner (Fig. 6C) led us to investigate the effect of WKYMVM or LXA4 on MAPK activation in human neutrophils. Accordingly, the effects of various concentrations of WKYMVM or LXA4 were studied on isolated human neutrophils. It was observed that the phosphorylation of ERKs was mediated by WKYMVM but not by LXA4 (Fig. 7, A and B). Moreover, WKYMVM-induced ERK phosphorylation was concentration-dependent and showed maximal activity at 300 nM (Fig. 7A). When we treated the cells with 1 μM of WKYMVM, ERK phosphorylation increased in a time-dependent manner, and maximal activity was reached after 5 min (Fig. 7B). However, in the case of the LXA4-stimulated cells, no significant change in the phosphorylation level of ERKs was detected (Fig. 7, A and B). JNK phosphorylation was also monitored by Western blot analysis using anti–phospho-JNK antibody and was found to be induced by WKYMVM treatment but not by LXA4 treatment (Fig. 8, A and B). These results indicate that the stimulation of FPRL1 by WKYMVM, but not by LXA4, induces the activation of signaling pathways that lead to the activation of ERKs and JNK.

Activation of ERKs by WKYMVM but not by LXA4. Human neutrophils were stimulated with various concentrations of WKYMVM or LXA4 for 5 min (A) or with 100 nM of WKYMVM or 1 μM of LXA4 for various times (B). Each sample (30 μg of protein) was subjected to 8% SDS-PAGE, and phosphorylated ERK levels were determined by immunoblot analysis with anti–phospho-ERK antibody. Immunoblot analysis with anti-ERK antibody was performed to confirm that the same amounts of proteins were loaded. The results shown are representative of at least two independent experiments.

Activation of JNK by WKYMVM but not by LXA4. Human neutrophils were stimulated with various concentrations of WKYMVM or LXA4 for 5 min (A) or with 100 nM of WKYMVM or 1 μM of LXA4 for various times (B). Each sample (30 μg of protein) was subjected to 8% SDS-PAGE, and phosphorylated JNK was determined by immunoblot analysis with anti–phospho-JNK antibody. Immunoblot analysis with anti-JNK antibody was performed to confirm that the same amounts of proteins were loaded. The results shown are representative of at least two independent experiments.
Both cPLA2 and PLD Activities Are Essential for Superoxide Generation in Neutrophils. In Fig. 4, we showed that WKYMVM but not LXA4 stimulated superoxide generation in human neutrophils. Previously, we reported on the independent functioning of cPLA2 and PLD on chemoattractant-induced superoxide generation in human monocytes (Bae et al., 2000). In terms of the signaling mechanisms involved in the differential activation of the superoxide-generating machinery by WKYMVM and LXA4, we suspected a differential role of MAPK in mediating cPLA2 activation and PLD activation. Furthermore, in the present study, WKYMVM-induced superoxide generation was almost completely ablated by cPLA2 inhibitors (AACOCF3 and MAFP) or by a phosphatidic acid acceptor (butan-1-ol) (Fig. 9). The iPLA inhibitor (BEL) or control alcohol not acting as a phosphatidic acid acceptor (butan-3-ol) did not affect superoxide generation induced by WKYMVM (Fig. 9). The result indicates that both of cPLA2 and PLD activation are essential for the activation of the NADPH oxidase complex. Because WKYMVM stimulates both PLA2 and PLD activation, it can induce superoxide generation in neutrophils. However, in the case of LXA4, PLD is activated but PLA2 is not, which leads to the lack of cellular superoxide generation in neutrophils.

Effect of WKYMVM-evoked stimulation of cPLA2 and PLD on superoxide generation in neutrophils. Neutrophils were incubated for 15 min in the presence or absence of 20 μM of MAFP, 20 μM of AACOCF3, or 20 μM of BEL at 37°C and stimulated for 5 min with 100 nM WKYMVM. Butan-1-ol (0.4%) or butan-3-ol (0.4%) was added just before stimulation with 100 nM WKYMVM for 5 min. Cytochrome c reduction was monitored as a change in absorption at 550 nm over 5 min, as described under Materials and Methods. Data are presented as means ± S.E.M. of three separate experiments.
WKYMVM and LXA4 Induce Different Signaling in FPRL1-Expressing RBL-2H3 Cells. To further support our notion that WKYMVM and LXA4 differentially stimulate human neutrophils via FPRL1 activation, we investigated the effects of WKYMVM and LXA4 on [Ca2+]i increase and ERK activation in FPRL1-expressing RBL-2H3 cells. As shown in Fig. 10A, stimulation of FPRL1-expressing RBL-2H3 cells with 1 μM WKYMVM or with 1.4 μM LXA4 elicited [Ca2+]i increase. Because no significant [Ca2+]i increase was seen in response to WKYMVM or LXA4 in vector-expressing RBL-2H3 cells (Fig. 10B), the result indicates that both agonists stimulate FPRL1, resulting in [Ca2+]i increase. We also tested the effects of the two agonists on ERK activation in FPRL1-expressing RBL-2H3 cells. Stimulation of FPRL1-expressing RBL-2H3 cells with 1 μM WKYMVM caused transient phosphorylation of ERKs in a time-dependent manner (Fig. 10C). However, when we stimulated FPRL1-expressing RBL-2H3 cells with 1.4 μM LXA4, we could not observe any significant enhancement of phosphorylation of ERKs (Fig. 10C). To confirm whether WKYMVM-induced ERK phosphorylation was mediated by the activation of FPRL1, we investigated the effect of WKYMVM or LXA4 on ERK phosphorylation in vector-expressing RBL-2H3 cells. As shown in Fig. 10D, neither WKYMVM nor LXA4 stimulated ERK phosphorylation in vector-expressing RBL-2H3 cells. The results indicate that although WKYMVM stimulates FPRL1, resulting in both [Ca2+]i increase and ERK activation, LXA4 stimulates [Ca2+]i increase but not ERK activation.

Differential signaling of WKYMVM or LXA4 in FPRL1-expressing RBL-2H3 cells. FPRL1-expressing RBL-2H3 cells (A) or vector-transfected RBL-2H3 cells (B) were loaded with 3 μM fura-2/AM for 50 min at 37°C in serum-free RPMI 1640 medium and washed three times with serum-free RPMI 1640. Before measurement, the cells (2 × 106) were suspended in Ca2+-free Locke solution containing 0.2 mM EGTA. Fura-2–loaded cells were stimulated with 100 nM of WKYMVM or 1.4 μM of LXA4. The charts shown are from a single experiment and are representative of three independent experiments. FPRL1-expressing RBL-2H3 cells (C) or vector-transfected RBL-2H3 cells (D) were stimulated with 100 nM WKYMVM or 1.4 μM of LXA4 for various times. Each sample (30 μg of protein) was subjected to 8% SDS-PAGE and phosphorylated ERK levels were determined by immunoblot analysis with anti–phospho-ERK antibody. Immunoblot analysis with anti-ERK antibody was performed to confirm that the same amounts of proteins were loaded. The results shown are representative of four independent experiments.
Discussion
Phagocytic leukocytes play a key role in innate immune responses, and classic chemotactic peptide receptors such as FPR or FPRL1 are involved in the modulation of this phagocyte activity (Le et al., 2001b). Although the signaling mechanisms of FPR have been well studied, the signaling pathways of FPRL1 have not been investigated systematically. In the present study, we examined the events downstream of FPRL1 in human neutrophils and found that the activation of FPRL1 caused Ca2+ increase via PLC-β activation in a PTX-sensitive manner, resulting in PKC/PLD-mediated NADPH oxidase stimulation. FPRL1 stimulation also caused PI3K activity-dependent Akt (data not shown) or ERK activation that elicited cPLA2 activation. Nonreceptor tyrosine kinases, such as FAK, were also activated (data not shown), and this FAK activity induced paxillin phosphorylation.
Although the role of chemotactic peptide receptors is generally regarded to be important, and many different ligands for receptors have been identified, the differential modulation of receptors has not been investigated. In the past, the activation of one chemoattractant receptor by different ligands has been regarded to induce the same cellular signaling and physiological responses. In this study, we suggest for the first time that FPRL1, a chemoattractant receptor, can be activated in a ligand-specific manner by WKYMVM or LXA4. WKYMVM, a peptide ligand of FPRL1, was found to stimulate superoxide generation and chemotactic migration in neutrophils by activating Ca2+ increases, MAPKs, cPLA2, PLD, and paxillin. However, LXA4 could stimulate chemotactic migration but not superoxide generation in human neutrophils. In terms of the associated cellular signaling, LXA4 stimulated Ca2+ increase and PLD and paxillin phosphorylation but not the activation of MAPKs and PLA2, which are essential for superoxide generation. With the neutrophil data, it was still unclear whether another receptor could be involved in the WKYMVM-stimulated neutrophil signaling. However, we subsequently demonstrated that differential signaling of FPRL1 in terms of Ca2+ increase and ERK activation was induced by WKYMVM or LXA4 in FPRL1-expressing RBL-2H3 cells but not in vector-transfected RBL-2H3 cells (Fig. 10). Taking our results together, it is reasonable to assume that FPRL1 can mediate distinct cellular signals and functional modulations by ligating different agonists in human neutrophils.
Recent reports have suggested that GPCRs could be modulated in a ligand-specific manner (Robb et al., 1994; Palanche et al., 2001), and ligand-specific GPCR modulation has been found to cause the activation of signaling molecules such as adenylate cyclase (Palanche et al., 2001). In the present study, we show that although WKYMVM and LXA4 stimulate neutrophils and that this results in [Ca2+]i increase (Fig. 1A), the activation of MAPKs was induced by WKYMVM but not by LXA4 (Figs. 7 and 8). To the best of our knowledge, this is the first time that MAPKs such as ERKs and JNK could be differentially modulated via the activation of the same receptor by different ligands. Concerning the mechanism involved in the differential modulation of one GPCR by different ligands, ligand-specific receptor active states have been suggested (Kenakin, 2001). Distinct receptor conformational change induced by different ligands may cause differential coupling patterns of receptor and the heterotrimeric G proteins, which results in the differential activation of effector molecules (Kenakin, 2001). Many chemoattractant receptors, including FPRL1, which is coupled to PTX-sensitive G proteins, induce diverse intracellular signaling through the transient coupling of the βγ subunits of G protein to effector molecules (Bokoch, 1995; Le et al., 2001b). Two of the major effector molecules of the Gβγ subunits are PLCβ and PI3Kγ (Metjian et al., 1999; Scott et al., 2001). Activation of PLCβ induces intracellular Ca2+ increases and PKC activation, which are essential for the activation of PLD (Bacon et al., 1995). PI3Kγ, however, induces Ras/Raf-mediated ERK activation (Avdi et al., 1996) and Akt activation by modulating PDK1 (Sasaki et al., 2000). Previous report demonstrated that different combinations of Gβγ subunits selectively modulate distinct phospholipid-dependent enzymes such as PLCβ or PI3Kγ (Maier et al., 2000). Taking together the previous report (Maier et al., 2000) and our data, it is possible that a distinct combination of Gβγ subunits will be involved in the activation of PLCβ or PI3Kγ in the downstream of FPRL1, resulting in [Ca2+]i increase or ERK activation. Because the binding of LXA4 to FPRL1 could stimulate [Ca2+]i increase/PLD but not ERK/cPLA2, it seems that the associated ligand binding-induced GPCR conformational change disables Gβγ subunit coupling with PI3Kγ. LXA4 also failed to stimulate Akt activity, the downstream event of PI3Kγ of FPRL1 by WKYMVM (data not shown).
Several different FPRL1 ligands have been identified. They include host-derived agonists (LL-37 and SAA), HIV Env domains (F peptide and V3 peptide), and the synthetic peptides (MMK-1 and WKYMVM) (Le et al., 1999, 2001b; Chiang et al., 2000; Christophe et al., 2001). MMK-1 was originally derived from a random peptide library and was identified by a novel autocrine selection method in yeasts engineered to express human FPRL1 (Klein et al., 1998). MMK-1 is a specific chemotactic factor for FPRL1-transfected human embryonic kidney 293 cells and an inducer of Ca2+ increase through FPRL1 (Klein et al., 1998). Recently, Serhan et al. demonstrated that two different ligands of FPRL1 (LXA4 and MMK-1) could compete with each other (Chiang et al., 2000). FPRL1 has been reported to be N-glycosylated at the NH2 terminus (Asn-4) and at the second extracellular loop (Asn-179) (Chiang et al., 2000). Moreover, these N-glycosylated moieties are regarded to be an important feature of intracellular trafficking (Ludwig et al., 2000). Deglycosylation of FPRL1 was reported to dramatically decrease the binding of the peptide ligand MMK-1 but not that of LXA4 (Chiang et al., 2000), which demonstrated the divergent domain requirements of LXA4 and MMK-1 for FPRL1. The FPRL1 binding site for WKYMVM has not been characterized. However, preliminary data suggest the possibility that LXA4 and WKYMVM interact with nonoverlapping sites on FPRL1. For example, pretreatment of neutrophils as well as FPRL1-expressing RBL-2H3 cells with LXA4 does not block subsequent activation of ERK, JNK, and PLA2 by WKYMVM (data not shown), despite the observed desensitization between the two ligands (Fig. 2). This result is consistent with those published by Serhan and colleagues (Chiang et al., 2000) and together indicates a different structural requirement for LXA4 interaction with FPRL1. We believe that WKYMVM and LXA4 can induce differential intracellular signaling by binding to the differential sites of FPRL1.
The activation of MAPKs such as ERKs, JNK, or p38 kinase is essential to transcriptional activation of many inflammatory cytokines (Craig et al., 2000). Generally, the stimulation of chemoattractant receptors that are coupled to PTX-sensitive GPCRs is known to induce MAPK activation (Bokoch, 1995). Recently, Hu et al. (2001) reported that FPRL1 activation by MMK-1 enhanced proinflammatory cytokine production in human monocytes. We also found that WKYMVM stimulation caused IL-6 production and showed PD98059 sensitivity in monocytes (data not shown), which indicates that WKYMVM-induced IL-6 production is ERK activity-dependent. Knowing that chemoattractant receptor activation-induced cytokine production is MAPK activity-dependent, it is apparent that the failure to activate MAPKs when LXA4 binds to its receptor would not cause proinflammatory cytokine production. Previous reports suggested that LXA4 plays an anti-inflammatory role in immune responses (Takano et al., 1997). Recalling that the production of several proinflammatory cytokines and superoxide generation are mediators of inflammatory responses, the failure of MAPK activation or superoxide generation will illustrate a possible differential role of LXA4 against peptide ligands for FPRL1.
In conclusion, although both WKYMVM and LXA4 bind to FPRL1, a classic chemoattractant receptor in human neutrophils, they transmit differential downstream signals. Functionally, the chemotactic migratory effects overlap; however, superoxide generation is only seen with WKYMVM. We suggest for the first time that FPRL1 is modulated differentially in a ligand-specific manner.
Footnotes
-
This study was supported by grant FPR02A5-43-110 of 21C Frontier Functional Proteomics Project from Korean Ministry of Science and Technology, the POSCO innovative research project, and the Medical Science and Engineering Research Center for Cancer Molecular Therapy from the Korea Science and Engineering Foundation. R.H. and R.D.Y. are supported by United States Public Health Service grant AI33503.
-
ABBREVIATIONS: fMLF, formyl-methionyl-leucyl-phenylalanine; PTX, pertussis toxin; GPCR, G protein-coupled receptor; FPR, formyl peptide receptor; PLC, phospholipase C; PI3K, phosphoinositide-3-kinase; MAPK, mitogen-activated protein kinase; PLA2, phospholipase A2; cPLA2, cytosolic phospholipase A2; iPLA2, calcium-independent phospholipase A2; PLD, phospholipase D; FPRL1, formyl peptide receptor-like 1; WKYMVM, Trp-Lys-Tyr-Met-Val-Met-NH2; ERK, extracellular signal-regulated protein kinase; AA, arachidonic acid; PBt, phosphatidylbutanol; GF109203X, 2-[1-(3-dimethylaminopropyl)-1H-indol-3-yl]-3-(1H-indol-3-yl)-maleimide; Ro-31–8220, 3-[1-[3-(amidinothio)propyl-1H-indol-3-yl]-3-(1-methyl-1H-indol-3-yl) maleimide methane sulfonate]; LY294002, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one; LXA4, lipoxin A4, 5(S),6(R),15(S)-trihydroxyeicosa-7E,9E,11Z,13E-tetraenoic acid; MAFP, methyl arachidonylfluorophosphonate; AACOCF3, arachidonyltrifluoromethyl ketone; BEL, bromoenol lactone; Y118, phospho-paxillin; Aβ42, amyloid β peptide 1–42; IL, interleukin; JNK, c-Jun NH2-terminal kinase; FBS, fetal bovine serum; BSA, bovine serum albumin; PKC, protein kinase C; FAK, focal adhesion kinase; PAGE, polyacrylamide gel electrophoresis; PD98059, 2′-amino-3′-methoxyflavone.
- Received July 24, 2002.
- Accepted May 30, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}