


Abstract
The human herpes virus 6 (HHV-6)-encoded chemokine receptor U51 constitutively activates phospholipase C (PLC) and inhibits cAMP-responsive element (CRE)-mediated gene transcription via the activation of Gq/11 proteins. Yet, chemokines known to bind U51 differentially regulate U51 coupling to G proteins. CCL5/RANTES induced pertussis toxin (PTX)-insensitive increases in PLC activity and changes in intracellular free calcium concentration ([Ca2+]i), whereas both CCL2/MCP-1 and CCL11/eotaxin failed to stimulate PLC activity or increase [Ca2+]i. In contrast, all three chemokines counteracted the effects of U51 on CRE activity via the activation of PTX-sensitive Gi/o proteins. For each of the tested chemokines, coexpression of U51 with a variety of Gα subunits, however, revealed a distinct profile for preferred G-protein coupling, which could be shifted by modulation of the relative expression of G proteins. These findings are consistent with a chemokine-selective trafficking of receptor stimulus to distinct G proteins and suggest that the constitutive activity of U51 and the chemokine-induced signaling involve different active states of the receptor. By virtue of its ability to constitutively activate signaling pathways, its G-protein promiscuity, and the chemokine-directed trafficking of receptor stimulus, U51 can be considered a sensitive and versatile virally encoded signaling device, potentially of importance in HHV-6-related pathologies.
Chemokine receptors belong to the family of G-protein-coupled receptors (GPCRs) and are involved in the regulation of the immune response, inflammation, and leukocyte trafficking (Rossi and Zlotnik, 2000). It is noteworthy that herpesviruses, including the human cytomegalovirus (HCMV), Kaposi's sarcoma-associated virus, and human herpesvirus 6 and 7 (HHV-6 and -7), contain genes that encode for proteins with homology to mammalian chemokine receptors (Murphy, 2001; Sodhi et al., 2004). A prominent feature of these virusencoded receptors (vGPCRs) is their ability to bind a broad spectrum of chemokines and to signal in a constitutively active manner, which is not apparent for their cellular homologs (Arvanitakis et al., 1997; Casarosa et al., 2001, 2003; Waldhoer et al., 2002). The HCMV genome encodes four GPCRs, of which US28 and UL33 are capable of constitutively activating several signaling pathways linked to inflammation in HCMV-infected cells (Casarosa et al., 2003; Minisini et al., 2003). Furthermore, GPCR homologs in rat and murine cytomegalovirus (R78 and M78, respectively) are required for in vivo virulence in the absence of defined ligands, and M78 signaling can affect immediate early mRNA accumulation (Beisser et al., 1999; Oliveira and Shenk, 2001). These findings suggest that constitutive activity of vGPCRs is of physiological relevance and allows or facilitates viruses to take control of infected cells and host immune response (Sodhi et al., 2004). In addition, some of these vGPCRs show promiscuous coupling to different G proteins (Casarosa et al., 2003; Rosenkilde et al., 2004), whereas cellular chemokine receptors mainly couple to Gi/o proteins (Rossi and Zlotnik, 2000).
HHV-6 causes widespread, persistent infection. Under immune suppressive conditions HHV-6 reactivates and is associated with inflammatory conditions, including encephalitis and transplantation diseases and has been linked to multiple sclerosis (Gompels, 2004). The HHV-6 genome encodes two proteins, U12 and U51, with homology to mammalian chemokine receptors considered as major candidates to contribute to inflammatory pathologies (Gompels, 2004). We have shown previously that U51 binds CC-chemokines, such as CCL5/RANTES, CCL11/eotaxin, and CCL2/MCP-1, chemokines associated with HHV-6-linked pathologies (Milne et al., 2000; Dockrell, 2003; Grivel et al., 2003; Gompels, 2004). In addition, U51 signals in a stable cell line of hemapoietic origin, but this could be either ligand-inducible or constitutive, because the cell line also secreted chemokine ligand (Milne et al., 2000). Very recently, U51 has been shown to positively regulate HHV-6 replication and to enhance cell-cell fusion in vitro (Zhen et al., 2005).
So far, there is no detailed information available on the signaling properties of the HHV-6-encoded GPCR U51. In the present work, we present evidence of constitutive and differential chemokine ligand-inducible signaling. U51 constitutively activates phospholipase C (PLC) and inhibits CRE-mediated gene transcription through a PTX-insensitive pathway. This constitutive activity of the receptor can be differentially modulated by human chemokines. Moreover, in systems with reduced constitutive activity induced by expression of various Gα subunits, the tested chemokines display G-protein-selective agonistic behavior. These results support the notion of chemokine-specific trafficking of receptor stimulus to different G-proteins (Kenakin, 1995a), suggesting the existence of chemokine-selective receptor conformations for these versatile vGPCRs. The widespread constitutive activity and receptor-G-protein promiscuity displayed by vGPCRs implies a common behavior within this group of GPCRs, which may contribute to herpesvirus pathogenesis.
Materials and Methods
DNA Constructs. The pcDNA3-derived vector containing HHV-6A-encoded U51 (pcDNA3-U51) was generated as described previously (Milne et al., 2000). The reporter plasmid pTLNC-21CRE was obtained from W. Born (National Jewish Medical and Research Center, Denver, CO). Gifts of pcDNA3-based vectors containing the cDNAs of Gαq (Dr. B. Conklin, University of California, San Francisco, CA), Gα11 (Dr. H. Umemori, Washington University Medical School, St. Louis, MO), Gαt (Dr. B. Defize), GRK2 and GRK2K220R (Dr. S. Cotecchia, Université de Lausanne, Lausanne, Switzerland), and PTX-insensitive mutant Gαo1, Gαi1, Gαi2, and Gαi3 (Dr. G. Milligan, University of Glasgow, Glasgow, Scotland) are gratefully acknowledged.
Cell Culture and Transfection. COS-7 cells were cultured and transiently transfected using the DEAE-dextran method as described previously (Casarosa et al., 2001). In all experiments, the total amount of cDNA transfected was maintained constant by addition of the empty vector (pcDNA3).
Binding Experiments. Labeling of CCL5 and binding in COS-7 Cells were performed as described previously (Gruijthuijsen et al., 2002). In saturation binding studies, cells were incubated in buffer (50 mM HEPES, pH 7.4, 1 mM CaCl2, 5 mM MgCl2, and 0.5% BSA) containing concentrations of 125I-CCL5 ranging between 0.1 and 25 nM. In competition experiments, cells were incubated with 2 nM 125I-labeled CCL5 and various amounts of unlabeled CCL2, CCL5, or CCL11. Nonspecific binding was determined in the presence of 0.1 μM unlabeled CCL5. Cell count was performed in triplicate in reserved wells at the end of each experiment and used to calculate number of binding sites per cell.
[3H]Inositol Phosphates Production. Cells were labeled overnight with myo-[2-3H]inositol (1 μCi/ml) as described previously (Casarosa et al., 2001) in the presence or absence of PTX (100 ng/ml), washed for 10 min with Dulbecco's modified Eagle's medium containing 25 mM HEPES, pH 7.4, 0.5% BSA, and 20 mM LiCl and incubated for 2 h in the same medium with or without the chemokines indicated. [3H]Inositol phosphates (InsP) were isolated by anion exchange chromatography and counted by liquid scintillation.
Reporter Gene Assays. CRE-driven gene transcription was measured as described previously (Casarosa et al., 2003). COS-7 cells were transfected with pTLNC-21CRE and indicated plasmids. Transfected cells were incubated for 18 h in the presence or absence of 100 ng/ml PTX. Then, the indicated chemokines were added together with forskolin (10-5 M). Twenty-four hours after transfection, CRE-driven luciferase expression was measured by luminescence in a Wallac Victor2 microplate reader (PerkinElmer Wallac, Turku, Finland).
Calcium Measurements. Agonist-stimulated increases in [Ca2+]i were quantified by monitoring the fluorescence of Fluo-4 AM-loaded COS-7 cells, using an automated NOVOstar microplate reader (BMG Labtech GmbH, Offenburg, Germany). Twenty-four hours after transfection, cells were treated with 100 ng/ml PTX, when indicated. Forty-eight hours after transfection, cells were loaded in Hanks' balanced salt solution containing 20 mM HEPES, 2.5 mM probenecid, 0.5% BSA, 2 μM Fluo-4 AM, and 0.02% Pluronic F-127, pH 7.4. Cells were washed three times, and fluorescence was measured (one data point; excitation, 485 nm; emission, 520 nm) for 10 s to calculate the mean basal value. After agonist addition, fluorescence was recorded for 50 s. Changes induced by Triton X-100 [0.25% (v/v)] injection were recorded for further 10 s to determine the maximal fluorescence. Basal and maximal values determined for each well were used to normalize the data. Results are expressed as percentage of maximal stimulation induced by Triton X-100.
Western Blot Analysis. Transiently transfected COS-7 cells were lysed 48 h after transfection in phosphate-buffered saline containing 1% Nonidet P-40, 0.1% sodium dodecyl sulfate, 0.5% sodium deoxycholate, 1 mM phenylmethylsulfonyl fluoride, and 2 μg each of aprotinin and leupeptin per milliliter; sonicated; separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis; and blotted onto a polyvinylidene difluoride membrane (NEN Life Science Products, Boston, MA). To estimate Gα subunit expression, increasing amounts of recombinant Gαi1 (sc-4232), Gαi2 (sc-4222), Gαi3 (sc-4223), Gαo1 (sc-4224), and Gαq (sc-4226) subunits (Santa Cruz Biotechnology, Inc., Santa Cruz, CA) were run on the same gel. The antibody recognizing Gαi/o/t/z (D-15) (Santa Cruz Biotechnology, Inc.) was used in combination with a rabbit anti-goat horseradish peroxidase-conjugated secondary antibody (Bio-Rad, Hercules, CA). The antibody recognizing Gαq/11 (C-19) (Santa Cruz Biotechnology, Inc.) was used in combination with a goat anti-rabbit horseradish peroxidase-conjugated secondary antibody (Bio-Rad). Protein bands were detected by an enhanced chemiluminescence assay (PerkinElmer Life and Analytical Sciences, Boston, MA) with an Image station (NEN Life Science Products) and quantified using Kodak Digital Science 1D Image Analysis Software version 3.0.2 (Eastmand Kodak, Rochester, NY). Membranes were stripped, and equal loading of the lanes was verified using an antibody recognizing β-actin (AC-15) (Sigma-Aldrich, St. Louis, MO) in combination with a goat anti-mouse horseradish peroxidase-conjugated secondary antibody (Bio-Rad).
Data Analysis. Curve fitting and data analysis were carried out by nonlinear regression analysis using Prism 4.0 and statistical analyses with InStat 3.0 (GraphPad Software, Inc., San Diego, CA). Specific binding was calculated by subtraction of nonspecific binding from total binding, which was in all of the cases below 5% of the total radioactivity added. Specific binding in cpm was subsequently used to calculate number of binding sites per cell by using the specific activity of the 125I-CCL5, as described previously (Gruijthuijsen et al., 2002). Unpaired t test (two groups) or one-way analysis of variance test (three or more groups) was applied. Data are expressed as mean ± S.E.M. of three independent experiments run in triplicates.

Chemokine binding profile at U51. A, saturation binding using 125I-CCL5. COS-7 cells were transfected with 5 μg of U51 cDNA. Forty-eight hours after transfection, 125I-CCL5 binding was measured. Nonspecific binding was determined in the presence of 0.1 μM unlabeled CCL5. Results are presented as specific binding (number of binding sites/cell). A representative experiment of three independent experiments, performed in triplicate, is shown. Inset, specific binding of 125I-CCL5 (10 nM) to COS-7 cells transfected with increasing amounts of U51 cDNA. B, displacement of 125I-CCL5 binding at U51 by CCL2, CCL5, and CCL11. COS-7 cells were transfected with 5 μg of U51 cDNA. Forty-eight hours after transfection, cells were incubated with 2 nM 125I-CCL5 in the presence of various concentrations of the displacing chemokines. Nonspecific binding was determined in the presence of 0.1 μM unlabeled CCL5. Data are presented as percentage of U51 total specific binding. The average of three experiments, each performed in triplicate, is shown.
Results
Chemokine Binding Properties and Constitutive Activity of HHV-6-Encoded Receptor U51. COS-7 cells were transfected with cDNA coding for the HHV-6A U51 open reading frame (Milne et al., 2000). Using 125I-CCL5 as radio-ligand, specific and saturable binding to one single binding site was obtained, similar to that found previously using the stable cell line U51-K562 (Milne et al., 2000) (Kd = 2.4 ± 0.8 nM, Bmax = 53,732 ± 7327 sites/cell) (Fig. 1A). Increasing concentrations of transfected U51 cDNA resulted in concomitant increases in the number of binding sites (Fig. 1A, inset). 125I-CCL5 was homologously displaced by unlabeled CCL5 and heterologously displaced by two other human chemokines, CCL2 and CCL11, showing similar pKi values for each chemokine ligand [pKi = 8.78 ± 0.19 (CCL5), 8.50 ± 0.15 (CCL11), and 8.18 ± 0.24 (CCL2); n = 3] (Fig. 1B).
Transfection of increasing amounts of U51 cDNA in COS-7 cells resulted in a PTX-insensitive, agonist-independent enhancement of InsP levels, indicating that the U51 protein exists in a spontaneously active conformation presumably coupling to Gq/11 proteins (Fig. 2A). In addition, U51 expression resulted in a PTX-insensitive, agonist-independent decrease in CRE-mediated transcription (Fig. 2B), whereas the constitutive activity of HHV-8 ORF74 was sensitive to PTX, as described previously (Smit et al., 2002). In the absence of forskolin, U51 expression had no detectable effect on CRE activity (data not shown), indicating the inability of U51 to activate Gs proteins.
Involvement of Gαq/11 Proteins in U51-Mediated Signaling. The receptor kinase GRK2 is known to scavenge both Gαq/11 subunits as well as Gβγ subunits, thus regulating signaling through a phosphorylation-independent mechanism as well (Premont et al., 1995; Sallese et al., 2000). Therefore, coexpression of U51 with GRK2 or the kinase-deficient mutant GRK2K220R (Diviani et al., 1996) led to an effective inhibition of U51-mediated InsP production (Fig. 3, A and B). In contrast, coexpression with Gαt, known to scavenge Gβγ subunits (Clapham and Neer, 1997), did not affect U51 constitutive signaling significantly (Fig. 3C). These results suggest that in COS-7 cells, U51 constitutively activates phospholipase C interacting with endogenous Gαq/11 subunits as described previously for the Gq-coupled HCMV US28 GPCR (Casarosa et al., 2001).

Constitutive activation of signaling pathways by U51. A, PLC activity. COS-7 cells (1 × 106 cells) were transiently transfected with increasing amounts of U51 cDNA. Forty-eight hours after transfection, InsP accumulation was measured in cells treated with or without 100 ng/ml PTX. B, CRE-mediated gene transcription. COS-7 cells (1 × 106 cells) were transiently transfected with increasing amounts of U51 cDNA, HHV-8 ORF74 (2 μg), or empty vector (mock; 5 μg) and pTLNC-21CRE (5 μg) and incubated in the presence or absence of 100 ng/ml PTX. Eighteen hours after transfection, 10 μM forskolin was added. Twenty-four hours after transfection, CRE-driven luciferase expression was determined. The asterisk denotes a statistically significant difference versus basal levels in cells expressing only U51 without PTX treatment. The average of four experiments, performed in triplicate, is shown.
Treatment of COS-7 cells with 0.3 μM calcium ionophore A23187 (calcimycin) but not with 0.2 μM protein kinase C activator phorbol ester phorbol 12-myristate 13-acetate led to a marked reduction CRE transcriptional activity (82 ± 4% reduction of the maximal response induced by 10 μM FSK; n = 4), suggesting that the observed effects probably involve the modulation of [Ca2+]i. Moreover, treatment of COS-7 cells, which endogenously express the Gq/11-coupled histamine H1 receptor, with 10 μM histamine resulted in a PTX-insensitive partial inhibition of CRE transcriptional activity (53 ± 4% reduction of maximal response induced by 10 μM FSK; n = 4) and in a PTX-insensitive increase in [Ca2+]i (data not shown). The H1 receptor antagonist mepyramine (1 μM) completely inhibited the effects of histamine on CRE transcriptional activity (93 ± 3% of maximal response induced by 10 μM FSK; n = 4). These data demonstrated that activation of Gq/11-mediated pathways leads to inhibition of CRE-mediated transcription in COS-7 cells, indicating that the constitutive PTX-insensitive effects of U51 on CRE-mediated transcription probably involve Gq/11 proteins.
Ligand-Induced Modulation of Signaling Pathways Activated by U51. To determine the effects of human chemokines on the constitutive signaling of U51, we stimulated U51-expressing cells with increasing concentrations of CCL2, CCL5, or CCL11. As shown in Fig. 4, A and B, CCL5 induced a small but significant dose-dependent increase in basal InsP levels in the absence of PTX (185 ± 4 versus 210 ± 5%, n = 4; p < 0.01, basal and stimulated, respectively; pEC50 = 8.8 ± 0.3). Moreover, in cells pretreated with PTX (100 ng/ml; 24 h), the effect of CCL5 on InsP production was further increased (180 ± 5 versus 229 ± 4%; n = 4; p < 0.01, basal and stimulated, respectively), whereas for CCL2 and CCL11, a small statistically nonsignificant effect was observed only at 100 nM (Fig. 4B). The pEC50 value of CCL5 was not affected by PTX treatment (data not shown). These data demonstrate that if coupling of U51 to PTX-sensitive G-proteins is prevented, CCL5-induced signaling through PTX-insensitive G-proteins is facilitated.
It is noteworthy that treatment of U51-expressing cells with CCL2, CCL5, and CCL11 counteracted the reduction of CRE-mediated gene transcription generated by the constitutive activity of U51 (Fig. 4C). Chemokine-induced effects were completely abolished by pretreatment with PTX, indicating coupling of U51 to Gi/o proteins upon chemokine binding.
Cellular chemokine receptors transduce extracellular signals, in particular, chemokine-induced increases in [Ca2+]i mainly through Gi/o proteins (four), although coupling to a broader range of G-proteins has been suggested (Arai and Charo, 1996). In COS-7 cells expressing U51, only CCL5 (30 nM) induced a marked increase in [Ca2+]i (20.2 ± 3.5% of the maximal response), whereas CCL2 and CCL11 had no significant effect within the dose range tested (up to 30 nM) (Fig. 4D). Yet, preincubation of cells with CCL2 and CCL11 completely antagonized the effect of CCL5 on [Ca2+]i, thus suggesting interaction with a common binding site on U51 (Fig. 4D, inset). In contrast to cellular chemokine receptors, in U51-expressing cells the transient increases in [Ca2+]i induced by CCL5 were insensitive to PTX treatment (Fig. 4E). None of the tested chemokines had detectable effects on mock-transfected COS-7 cells (CCL5; Fig. 6, A and B; CCL2 and CCL11, data not shown). These results indicate that both the constitutive activity and the CCL5-induced stimulation of U51 promote receptor coupling to PTX-insensitive G-proteins, presumably of the Gq/11 family.
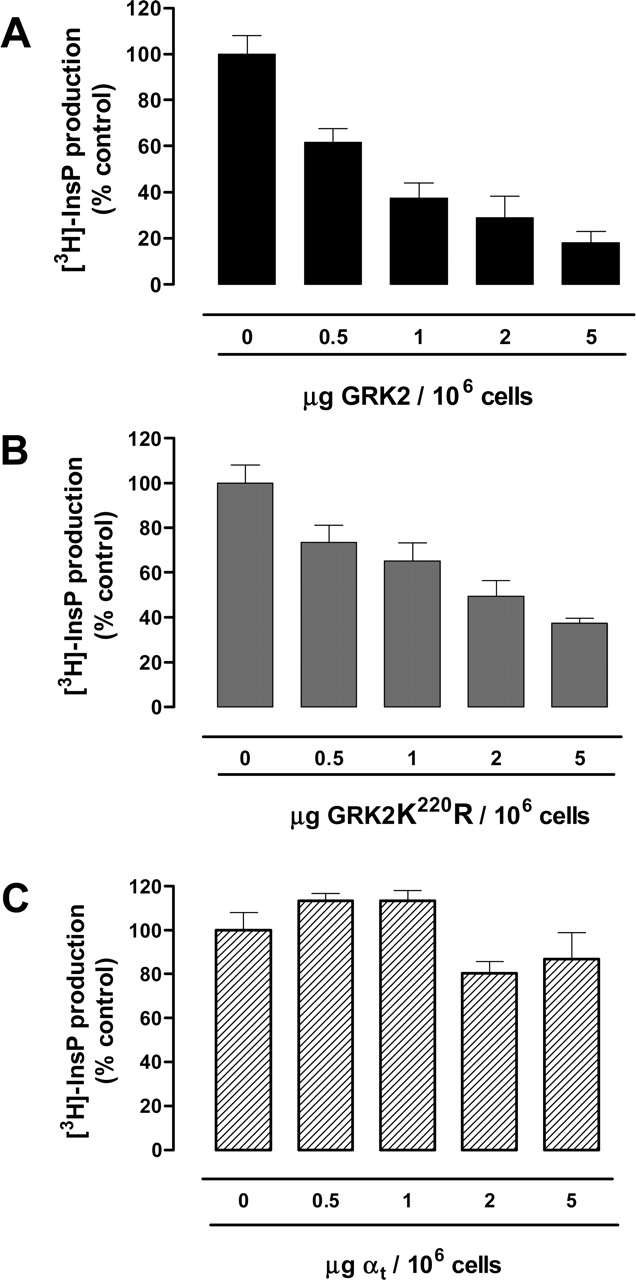
Regulation of U51-mediated constitutive activity by GRK2. COS-7 cells (1 × 106 cells) were transiently cotransfected with U51 (2 μg) plus 5 μg of vector (control; 0) or increasing amounts of GRK2 (A), GRK2-K220R (B), or Gαt (C) cDNAs as indicated. Forty-eight hours after transfection, U51-mediated InsP accumulation was measured. Data are expressed as percentage of maximal stimulation in control cells. The average of three experiments, performed in triplicate, is shown.
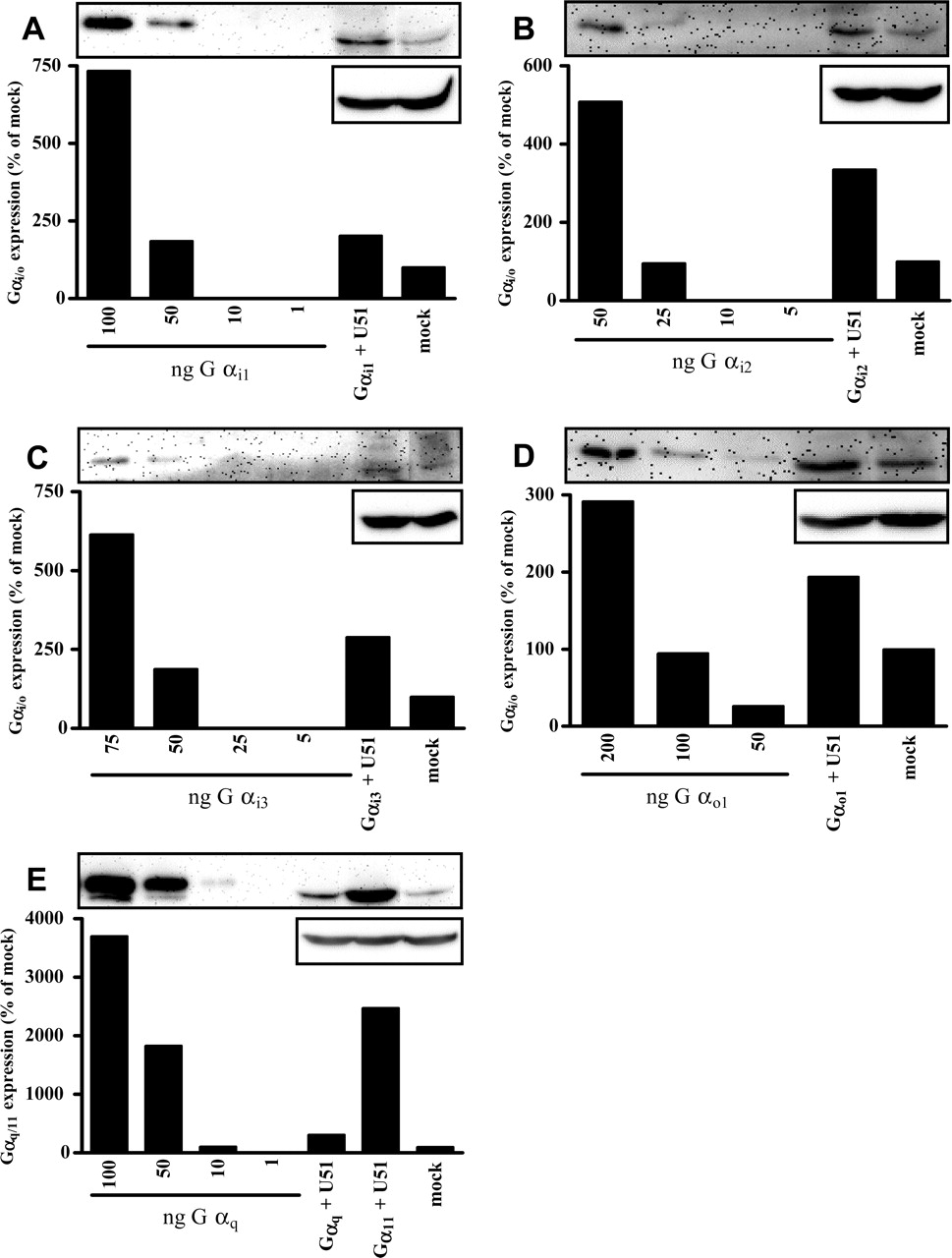
Promiscuous Coupling of U51 to Different G Proteins. To study the ability of U51 to activate different G-proteins in more detail, we examined the effects of different G-proteins, including Gα subunits of both Gq/11 family and PTX-insensitive Gα subunits of the Gi/o family (Wise et al., 1997) on constitutive U51-mediated signaling. To estimate the levels of overexpression of the various Gα subunits achieved by transient transfection of COS-7 cells, we used defined amounts of recombinant G-proteins as standards in Western blots, along with lysates from cells transfected with U51 and each Gα subunit (Fig. 5). All Gα subunits seem to be properly expressed as determined by Western blot analysis (Fig. 5), as reported previously (Wise et al., 1997). All transfected Gα subunits were found to be expressed within a (patho)physiological range (Davis et al., 2000) and at comparable levels (Fig. 5). All Gα subunits of the Gi family (Fig. 5, A-C) and Gα11 (Fig. 5E) were expressed at approximately 50 ng of protein/50 μg of total protein, whereas Gα01 and Gαq were expressed at approximately 150 ng of protein/50 μg of total protein and 25 ng of protein/50 μg of total protein, respectively (Fig. 5, D and E). Binding of 125I-CCL5 to U51 was not altered upon coexpression of Gαi subunits U51, only increased upon expression of Gαq (2.6-fold), Gαo1 (5.5-fold), or Gα11 (12.4-fold).
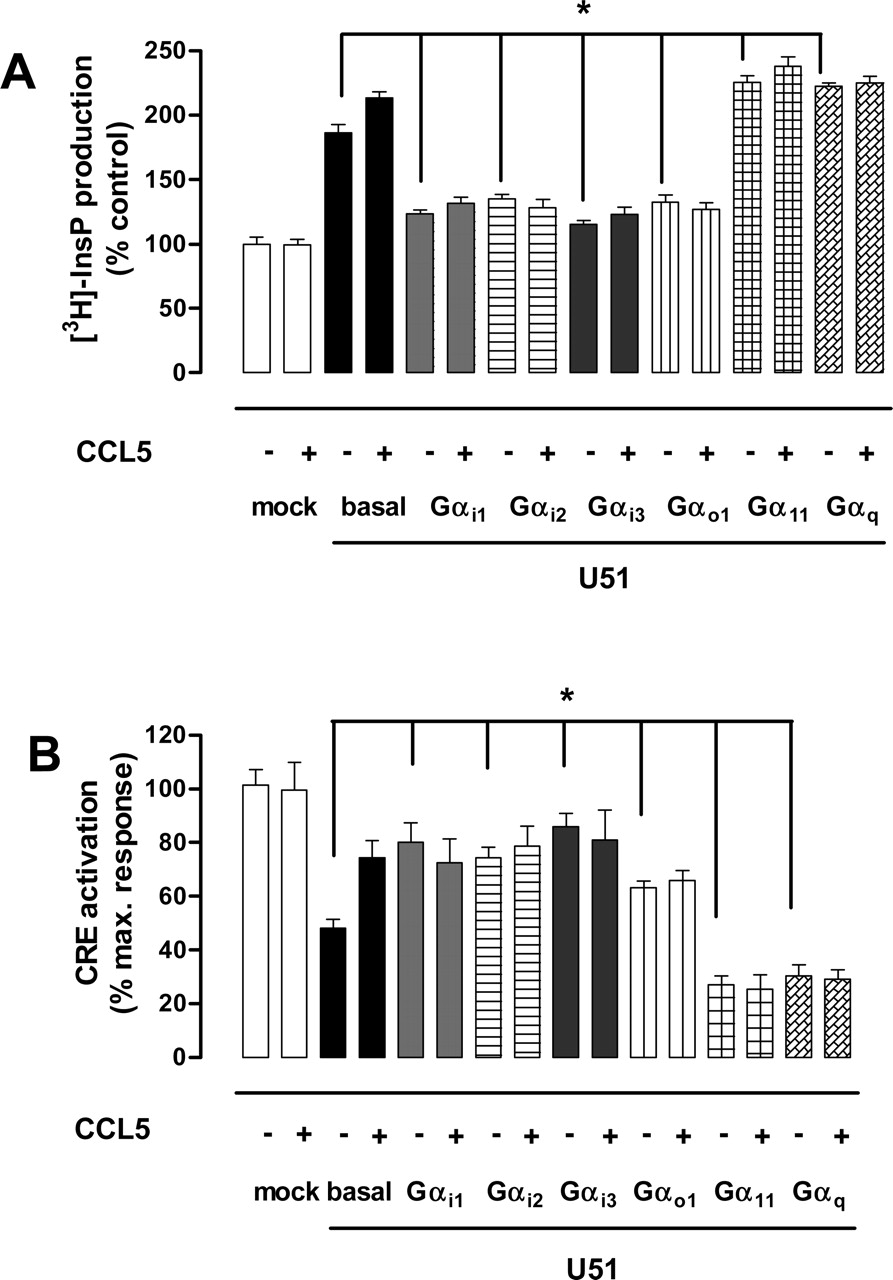
The modulating effects on basal signaling, observed upon introduction of all the Gα subunits indicated functional expression of these proteins in COS-7 cells, as reported previously (Wise et al., 1997; Casarosa et al., 2003). Coexpression of U51 with Gαi1, Gαi2, Gαi3, or Gαo1 led to a significant decrease in basal levels of InsP (Fig. 6A). Conversely, coexpression of U51 with Gαq or Gα11 led to a small but significant increase in the basal levels of InsP, compared with cells expressing U51 only.
Likewise, coexpression of U51 with Gαi1, Gαi2, Gαi3, and Gαo1 reversed the constitutive, U51-mediated inhibition of CRE activity to levels similar to those induced by CCL5 in the absence of exogenous G-proteins (Fig. 6B). In contrast, coexpression of U51 with Gα11 or Gαq further enhanced the U51-mediated constitutive inhibition of CRE activity (Fig. 6B).
In the presence of exogenous G-proteins, CCL5 (Fig. 6), CCL2 and CCL11 (not shown) failed to modulate both InsP production (Fig. 6A) and CRE activity (Fig. 6B) over the basal levels induced by the constitutive activity of U51.
Chemokine-Specific Stimulus Trafficking to Different G Proteins Revealed by Stimulus-Biased Systems. To further evaluate the influence of G-protein expression on inducible U51 signaling, we used stimulus-biased assay systems allowing detection of ligand-specific effects upon differential G-protein expression (Watson et al., 2000). Cells were transfected with U51 and various PTX-insensitive mutant Gαi/o subunits and pretreated with PTX to inactivate endogenous Gi/o proteins (Wise et al., 1997). Thereafter, the ability of chemokines to induce increases in intracellular calcium in the presence of different G-proteins was determined. Expression of the different Gα subunits seemed comparable, as shown by Western blot analysis in Fig. 5. These studies revealed an interesting chemokine-specific G-protein activation (Fig. 7). In cells coexpressing U51 and Gαi1 or Gαi2, CCL2 and CCL11 induced increases in [Ca2+]i, with CCL2 being the most efficacious agonist in those conditions, whereas CCL5 had no effect (Fig. 7, A and B). Coexpression of Gαi3, however, revealed a different rank order of efficacies of the tested chemokines. In this condition, CCL5 induced a partial response, whereas CCL11 had no effect, and CCL2 remained the most efficacious agonist under these conditions (Fig. 7C). In contrast to results using Gαi proteins, coexpression of U51 with Gαo1 (Fig. 7D), Gα11 (Fig. 7E), or Gαq (data not shown) revealed a receptor phenotype that was only sensitive to CCL5. Coexpression of Gαo1 resulted in a slightly reduced pEC50 for CCL5 (Gαo1 = 8.60 ± 0.15 versus no additional Gα subunits = 9.02 ± 0.09; n = 4; p = 0.05; unpaired t test). The pEC50 values for CCL5 in the presence of Gα11 or Gαq were comparable with the pEC50 for CCL5 without expression of additional Gα subunits (Fig. 7).
Discussion
Most virally encoded GPCRs of a variety of β- and γ-herpesviruses signal constitutively and show promiscuous G-protein coupling, unlike their cellular homologs (for review, see Sodhi et al., 2004). In this study, we show constitutive and inducible signaling combined with novel vGPCR chemokine-induced trafficking of receptor stimulus.

Ligand-induced modulation of signaling pathways by U51. A, chemokine effect on U51-mediated PLC activity. COS-7 cells were transiently transfected with cDNA encoding U51 (2 μg/106 cells). Cells were incubated with the indicated chemokines (100 nM), and InsP production was measured. Results are expressed as percentage of vector-transfected (mock) control cells. The average of four experiments, each performed in triplicate, is shown. The asterisk denotes a statistically significant difference versus nontreated U51-expressing cells. B, concentration-response curves for CCL2, CCL5, and CCL11. Cells were incubated with increasing concentrations of the indicated chemokine, and InsP production was measured. Results are expressed as percentage of maximal response induced by CCL5. The average of three experiments, each performed in triplicate, is shown. The effects shown for CCL2 and CCL11 were measured in PTX-treated cells. C, effects of various chemokines on U51-modulatory effects on CRE-mediated gene transcription. COS-7 cells (1 × 106 cells) were transiently transfected with U51 cDNA (2 μg) or empty vector (mock; 2 μg) and pTLNC-21CRE (5 μg) and incubated in the presence or absence of PTX. The asterisks denote a statistically significant difference versus basal levels in nontreated U51-expressing cells. Results are expressed as percentage of the maximal response induced by forskolin in vector-transfected (mock) cells. The average of four experiments, performed in triplicate, is shown. D, effects of chemokines on [Ca2+]i. COS-7 cells were transiently transfected with U51 cDNA, (2 μg/106 cells). Forty-eight hours after transfection, cells were loaded with Fluo-4 AM and exposed to chemokines indicated (30 nM). Inset, preincubation of U51-expressing cells (10 min) with 30 nM CCL2, or CCL11 on CCL5-induced (30 nM) intracellular Ca2+ mobilization. E, insensitivity to pertussis toxin of CCL5-mediated intracellular Ca2+ mobilization. COS-7 cells were transiently transfected with U51 cDNA (2 μg/106 cells) and treated with or without PTX (100 ng/ml) for 24 h. Forty-eight hours after transfection, cells were loaded with Fluo-4 AM and exposed to CCL5 (30 nM). Results are shown as percentage of maximal response induced by Triton X-100. A representative experiment of three independent experiments, performed in triplicate, is shown.
We report for the first time that the HHV-6-encoded GPCR U51 constitutively activates PLC, via a PTX-insensitive Gq/11-linked pathway. The involvement of Gq/11 proteins in U51-mediated signaling was corroborated by coexpression of U51 with Gq and G11 proteins, GRK2 or its kinase-deficient mutant GRK2K220R, which scavenges Gαq/11 and Gβγ subunits, or the Gβγ subunits scavenger Gαt. Coexpression with Gq and G11 led to a further increase in U51-mediated InsP production, whereas coexpression with GRK2 or GRK2K220R resulted in efficient inhibition. Gαt had no effect on the constitutive activity of U51, indicating involvement of Gαq/11 but not Gβγ subunits, as described for other constitutively active vGPCRs (Casarosa et al., 2001, 2003).

Expression of Gα subunits. A to D, expression of Gαi/o. COS-7 cells (1 × 106 cells) were transiently transfected with cDNA encoding U51 (2 μg) and different Gα subunits (2 μg each) or empty vector (4 μg). Expression of Gαi/o subunits was determined by Western blot analysis using an antibody against Gαi/o/t/z (top). To ensure equal loading of the lanes, the blots were stripped and checked for expression of β-actin (bottom). Increasing amounts of recombinant Gαi1, Gαi2, Gαi3, or Gαo1 subunits were run on the same gel to estimate Gα subunit expression. E, expression of Gαq/11. COS-7 cells (1 × 106 cells) were transiently transfected with cDNA encoding U51 (2 μg) and Gαq or Gα11 subunits (2 μg each) or empty vector (4 μg). Expression of Gαq/11 subunits was determined by Western blot analysis using an antibody against Gαq/11 (top). To ensure equal lane loading, the blots were stripped and checked for expression of β-actin (bottom). Increasing amounts of recombinant Gαq subunits were run on the same gel to estimate Gα subunit expression. Quantification of the bands (bar diagrams) was done with Kodak Digital Science 1D Image Analysis Software version 3.0.2. A representative experiment of three independent experiments is shown.
In contrast to other vGPCRs, U51 significantly and constitutively decreases CRE-mediated gene transcription in a PTX-insensitive manner. Because cellular chemokine receptors are known to couple to Gi/o proteins (Rossi and Zlotnik, 2000), this PTX insensitivity was surprising. CRE activity, however, is regulated by various signals, including cAMP, [Ca2+]i, and/or other signaling pathways (Shaywitz and Greenberg, 1999). Control experiments indicate that in COS-7 cells, activation of Gq/11 proteins increases [Ca2+]i and inhibits CRE-mediated gene transcription as well. This could be explained by the endogenous expression of adenylyl cyclase isoform IX in COS-7 cells, which is negatively regulated by increases in [Ca2+]i (Paterson et al., 2000). Thus, in nonstimulated conditions U51 can adopt an active conformation that preferentially couples to Gq/11 proteins, leading to PLC activation, subsequent increases in [Ca2+]i, and inhibition of CRE-mediated transcription.

Effect of various Gα subunits on the chemokine-induced and constitutive signaling of U51. A, effect of coexpression of U51 with various Gα subunits on U51-induced PLC activity. COS-7 cells (1 × 106 cells) were transfected with either U51-encoding cDNA (basal; 2 μg) or empty vector (mock; 2 μg) and cDNAs encoding different Gα subunits (2 μg/106 cells) or not. Cells were incubated with CCL5 (100 nM) when indicated, and InsP accumulation was measured. Results are shown as percentage of vector-transfected (mock) control cells. B, effect of coexpression of U51 with various Gα subunits on U51-modulated CRE-mediated gene transcription. COS-7 cells (1 × 106 cells) were transfected with either U51-encoding cDNA (basal; 2 μg) or empty vector (mock; 2 μg), and cDNAs encoding different Gα subunits (2 μg) or not and pTLNC-21CRE (5 μg). Cells were incubated with CCL5 (100 nM) when indicated, and CRE-driven luciferase expression was determined. Results are shown as percentage of maximal response induced by forskolin in vector-transfected cells. The asterisks denote a statistically significant difference versus basal levels present in U51-expressing cells (p < 0.05; n = 3). The average of three experiments, performed in triplicate, is shown.
U51 binds a variety of human CC-chemokines, among which are CCL2, CCL5, and CCL11 (Milne et al., 2000). Only CCL5 significantly induced increments in [Ca2+]i and modestly activated PLC over basal signaling in a Gi-independent manner. CCL2 and CCL11 rather acted as competitive ligands, antagonizing the effects of CCL5 on calcium mobilization. All three chemokines induced reversal of the U51-mediated inhibition of CRE activity. These counteracting effects of the three chemokines could be considered as negative efficacy and hence the chemokines might be classified as inverse agonists (Neubig et al., 2003). Yet, their effects were completely inhibited by PTX treatment, indicating involvement of an active state of the receptor.
Our findings indicate that CCL2, CCL5, and CCL11 act as agonists at U51, trafficking the receptor signal between Gq/11 and Gi/o proteins (CCL5) or only to Gi/o proteins (CCL2 and CCL11), whereas under nonstimulated conditions U51 constitutively signals mainly to Gq/11 proteins (Fig. 8). Based on this, we speculate that chemokine binding to U51 (partly) redirects U51 signaling to Gi/o proteins. This is consistent with our observation that coexpression of U51 with different Gαi/o subunits results in a reduced activation of PLC. When relative expression levels of Gi/o proteins are high, a redistribution of constitutively active U51 receptor states can occur and chemokines differentially traffic U51 signaling to Gi/o proteins. Likewise, their PTX-sensitive effects on CRE-mediated transcription were abolished by overexpression of Gαq or Gα11 subunits, indicating that shifting the relative expression in favor of Gq/11 proteins, eliminates any further coupling of U51 to Gi/o proteins.
To study the promiscuous coupling of U51, we used PTX-resistant Gαi/o subunits (Wise et al., 1997) within stimulus-biased assay systems to detect chemokine-induced trafficking of receptor stimulus to individual Gi/o proteins (Watson et al., 2000). These PTX-insensitive Gi/o isoforms are similarly expressed and seem functional.

Responses to chemokines in G-protein-biased assay systems. COS-7 cells were transfected with 2 μg of pcDNA3-U51/106 cells and 2 μg/106 of different Gα subunits or pcDNA3 vector alone. Twenty-four hours after transfection, cells were exposed to 100 ng/ml PTX. Forty-eight hours after transfection, cells were loaded with Fluo 4-AM and exposed to increasing concentrations of chemokines. Calculated pEC50 values in the presence of the different Gα subunits are as follows: for CCL2, 9.92 ± 0.17 (A; Gαi1), 9.53 ± 0.16 (B; Gαi2), and 9.91 ± 0.19 (C; Gαi3) (p = 0.25; not significant); for CCL5, 8.88 ± 0.18 (C; Gαi3), 8.60 ± 0.15 (D; Gαo1), 9.05 ± 0.11 (E; Gα11), and 9.02 ± 0.09 (F; no additional Gα subunits) (p = 0.15; not significant; analysis of variance). Data are shown as percentage of maximal response induced by Triton X-100. The average of four experiments, performed in triplicate, is shown.
In the presence of additional Gαi1 or Gαi1subunits, CCL2 and CCL11 selected receptor states that could couple to Gαi1 and Gαi2, although with different efficacies and potencies. Under these conditions, CCL2 was the most efficacious and potent agonist. It is noteworthy that CCL5 did not induce any detectable response but was still able to specifically bind 125I-CCL5 (data not shown). In contrast, when the system was biased to Gαi3, CCL2 evoked a response but CCL11 had no detectable effect, suggesting different ways for CCL2 and CCL11 to induce U51 signaling. It is noteworthy that the introduction of Gα11 and Gαq induced significant increases in 125I-CCL5 binding. This was also the case for Gαo1, which also recognizes receptor states selected by CCL5 (Fig. 7D). This is to be expected according to the extended ternary complex model (ETCM) of receptor occupancy, because the degree to which agonist binding would be enhanced depends upon the relative stoichiometries of the G-proteins involved and their affinities for the receptor states (Watson et al., 2000). In the absence of additional Gα subunits, where U51 most probably couple to Gαq/11 proteins, CCL2 and CCL11 could not induce calcium mobilization and antagonized the Gαq/11-mediated effects of CCL5. Therefore, CCL2 and CCL11 were unable to stimulate U51 when the system was biased to Gαq or Gα11. Under these conditions or without additional Gα subunits, only CCL5 induced a robust calcium mobilization via PTX-insensitive Gαq/11 proteins.
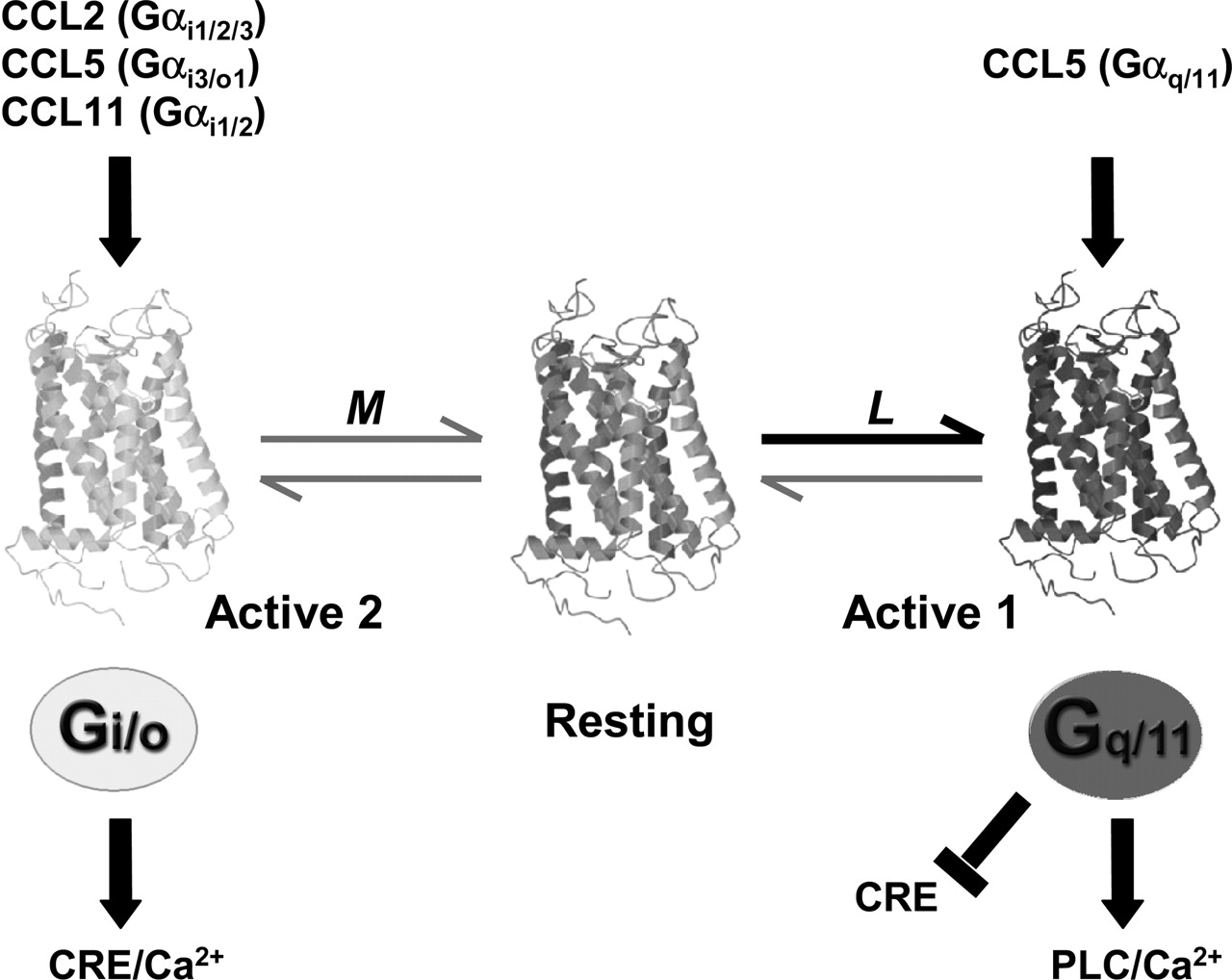
Schematic representation of constitutive and chemokine-induced signaling of U51. The scheme is based on the three state model of agonist action (Leff et al., 1997). U51 exists in a resting conformation (middle) and at least two active conformations, which interact with different G-proteins. The active conformation 1 (right) interacts with Gq/11 proteins, and the active conformation 2 (left) interacts with Gi/o proteins. In the absence of an agonist, the distribution of the receptor states is governed by the equilibrium constants L and M. In view of the observed constitutive activity of U51, the equilibriums are spontaneously displaced to the active conformation 1, resulting in constitutive activation of Gq/11 proteins. The presence of an agonist redistributes the receptor states, trafficking the signaling of U51 to different G-proteins. CCL5, a more pleiotropic agonist, selects Gq/11- and Gi/o-coupled active states, resulting in CCL5-induced increases in PLC activity and intracellular Ca2+ mobilization. Conversely, CCL2 and CCL11 select only Gi/o-coupled active states. The redistribution of receptor states induced by CCL2 and CCL11 results in an apparent decrease in the inhibition of CRE activity induced by the spontaneously favored active state 1. In the presence of exogenous Gαi/o subunits, CCL2 and CCL11 differentially induced intracellular Ca2+ mobilization.
In cells coexpressing U51 and Gαo1, only CCL5 was able to induce signaling. Cotransfected Gαo1 is expressed and induced significant changes in the basal activity of U51 (Figs. 5 and 6), and slightly reduces the pEC50 to CCL5 in calcium mobilization experiments (Fig. 7D). It cannot be ruled out that the calcium response induced by CCL5 in Gαo1-expressing cells reflects a response by endogenous Gq/11 proteins. These data reinforce the idea that each chemokine transfers U51 signaling to a distinct set of G-proteins, most probably by selecting various active receptor conformations (Fig. 8).
Various examples of GPCRs exhibiting ligand-specific trafficking of receptor signals have been reported (Kenakin, 2003, and references therein). Simulations with the ETCM showed that reversals of relative agonist efficacies could be affected by different ratios of two G-proteins (Kenakin, 2003). For constitutively active receptors, the ETCM predicts increases in basal activity result in diminished maximal response to agonists (Chen et al., 2000), as observed here for CCL5 in cells expressing only U51 and in biased systems with added Gq/11 proteins. Moreover, modeling differential binding of agonists to multiple receptor conformations redistributes their relative abundances and the ratios of the corresponding signaling species (Chen et al., 2000). Therefore, our data show that different chemokines (i.e., CCL2, CCL5, and CCL11) possess distinct efficacies toward U51, depending on the cellular context and the G-protein expression profile (Fig. 8).
Agonist-directed trafficking of receptor stimulus to different G-proteins (Kenakin, 1995a) has been used to explain protean agonism (Kenakin, 1995b) invoking a redistribution of receptor species (Leff et al., 1997; Brink, 2002). For U51, the putative protean behavior of chemokines could entirely be explained within the concept of ligand-directed trafficking of receptor stimulus. To our knowledge, results in this article represent the first example of a chemokine/(viral)chemokine receptor system for which observations of this type can be explained directly by invoking such a mechanism. This may help to understand how the ligand-induced signaling is routed at chemokine receptors. Inducible and constitutive signaling has recently been shown for the herpesvirus saimiri ECRF3 and equine herpesvirus 2 chemokine receptor (Rosenkilde et al., 2004, 2005). Ligand-induced and constitutive signaling occurred through different G-proteins, suggesting that the G-protein selectivity is unique for each vGPCR. Our current study on U51 takes these observations further in demonstrating not only selectivity in constitutive signaling but also selectivity in chemokine-induced signaling depending on the expression of specific G-proteins.
In conclusion, we present evidence for the existence of differential chemokine-induced activation of a constitutively active vGPCR, HHV-6 U51. The constitutive activity of U51 and the chemokine-induced signaling involve activation of different molecular pathways. U51 exists in a constitutively active state preferentially coupled to Gq/11 proteins, which can be differentially redistributed to different Gi/o proteins upon binding of different chemokines. Considering the widespread observation of constitutive activity and receptor-G-protein promiscuity for (v)GPCRs, these observations on ligand-dependent redirected signaling could be more common than previously recognized and may help to interpret the currently widespread claim of protean behavior of receptor ligands, particularly in different cellular contexts (Kenakin, 2003). Ligand-selected U51 conformations may be favored in membrane microdomains, such as lipid rafts, linked with Gi/o proteins or caveolins linked with Gq/11 proteins (Oh and Schnitzer, 2001). This may be important for infections in vivo where resting lymphocytes lack caveolin, thereby affecting chemokine receptor activity (Venkatesan et al., 2003). Moreover, marked increases in G-proteins of the Gi/o family have been reported upon differentiation of hematopoietic cells, which are well known targets of HHV-6 (Davis et al., 2000). These increases are much larger that the levels of overexpression induced by our transient transfections, suggesting that our observations are of physiological relevance. Thus, in a conceivable scenario in which U51 is expressed after HHV-6 infection in different cell types with distinct G-protein composition and chemokine expression profile, U51 signaling properties will lead to altered signaling, which could affect HHV-6 infection and its associated pathologies.
Acknowledgments
C.P.F. thanks Dr. S. A. Fratantoni for critical revisions.
Footnotes
-
This work was funded in part by The Netherlands Organization for Scientific Research (Jonge Chemische Wetenschappen) (to C.P.F. and D.V.), Dutch Technology Foundation (Stichting Technische Wetenschappen) (to D.V. and H.F.V.), and the Royal Netherlands Academy of Arts and Sciences (to M.J.S.). U.A.G. acknowledges support, in part, by the Royal Society and the Biotechnology and Biological Sciences Research Council (UK) at London School of Hygiene and Tropical Medicine.
-
ABBREVIATIONS: GPCR, G-protein-coupled receptor; HCMV, human cytomegalovirus; HHV, human herpesvirus; vGPCR virus-encoded G-protein-coupled receptor; RANTES, regulated on activation, normal T cell expressed and secreted; PLC, phospholipase C; CRE, cAMP response element; PTX, pertussis toxin; BSA, bovine serum albumin; InsP, inositol phosphates; AM, acetoxymethyl ester; FSK, forskolin; ETCM, extended ternary complex model.
- Received May 27, 2005.
- Accepted December 5, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}