


Abstract
Genomic studies have identified a D398N variation in the α5 subunit of nicotinic acetylcholine receptors (AChRs) that increases risk of nicotine dependence and lung cancer. (α4β2)2α5 AChRs are a significant brain presynaptic subtype in brain. Their high sensitivity to activation by nicotine and high Ca2+ permeability give them substantial functional impact. α3β4* and α3β2* AChRs are predominant postsynaptic AChRs in the autonomic nervous system, but rare in brain. The amino acid 398 of α5 is located in the large cytoplasmic domain near the amphipathic α helix preceding the M4 transmembrane domain. These helices have been shown to influence AChR conductance by forming portals to the central channel. We report that α5 Asn 398 lowers Ca2+ permeability and increases short-term desensitization in (α4β2)2α5 but not in (α3β4)2α5 or (α3β2)2α5 AChRs. This suggests that a positive allosteric modulator would augment nicotine replacement therapy for those with this risk variant. α5 D398N variation does not alter sensitivity to activation. The high sensitivity to activation and desensitization of (α4β2)2α5 AChRs by nicotine results in a narrow concentration range in which activation and desensitization curves overlap. This region centers on 0.2 μM nicotine, a concentration typically sustained in smokers. This concentration would desensitize 60% of these AChRs and permit smoldering activation of the remainder. The low sensitivity to activation and desensitization of (α3β4)2α5 AChRs by nicotine results in a broad region of overlap centered near 10 μM. Thus, at the nicotine concentrations in smokers, negligible activation or desensitization of this subtype would occur.
Introduction
Genome-wide association studies have identified sequence variants in the region encoding α3, β4, and α5 subunits of nicotinic acetylcholine receptors (AChRs) that increase risks of nicotine dependence and, probably thereby, lung cancer (Amos et al., 2008; Berrettini et al., 2008; Bierut et al., 2008; Hung et al., 2008; Thorgeirsson et al., 2008; Weiss et al., 2008; Saccone et al., 2009; Wang et al., 2009). Only one of these causes a change in amino acid sequence, the α5 variation D398N. Asn is the risk variant. It reduced the response to epibatidine of (α4β2)2α5 AChRs assayed using Ca2+ flux (Bierut et al., 2008).
α5* AChRs are important in brain. (α4β2)2α5 AChRs are a significant presynaptic subtype accounting for 10 to 37% of total α4β2* AChRs, depending on the brain region (Brown et al., 2007; Mao et al., 2008). Their high sensitivity to activation by nicotine and high Ca2+ permeability give them substantial functional impact (Brown et al., 2007; Tapia et al., 2007; Kuryatov et al., 2008; Grady et al., 2010). They are sensitive to nicotine-induced up-regulation in cell culture (Kuryatov et al., 2008). In brain, up-regulation was not observed (Mao et al., 2008). This may be because the amount of α5 was limiting. α5 assembles very avidly with α4β2 (Kuryatov et al., 2008). Partially assembled pools of subunits are susceptible to the pharmacological chaperone effects of nicotine that account for rapidly increased AChR assembly in the presence of nicotine (Kuryatov et al., 2005). Excess α5 results in formation of dead-end intermediates that cannot assemble into mature AChRs (e.g., α5α4β2α5) (Kuryatov et al., 2008). Transfected cells that express sufficient excess α5 or β3 to express only (α4β2)2α5 or (α4β2)2β3 AChRs accumulate dead end intermediates, have unusual morphologies, and grow slowly. The amount of α5 in brain may be limited to avoid forming dead-end intermediates.
α3β4* and α3β2* AChRs are rare in the brain, except in the medial habenula (Gotti et al., 2007; Salas et al., 2009). They are predominant postsynaptic AChRs in the autonomic nervous system (Xu et al., 1999). Human α3β2*, but not the more predominant α3β4* AChRs, are sensitive to nicotine-induced up-regulation (Wang et al., 1998). α5 subunits increase the Ca2+ permeability and desensitization rates of these subtypes (Gerzanich et al., 1998). α5 increases sensitivity to nicotine activation of α3β2 but not α3β4 AChRs (Gerzanich et al., 1998).
The α5 amino acid 398 is in the large cytoplasmic domain adjacent to the conserved amphipathic α-helix that immediately precedes the M4 transmembrane domain. This is far from the extracellular location of ACh binding sites, so it is unlikely to influence the sensitivity of agonist binding. A ring of charged amino acids in the M2 transmembrane domain that lines the cation channel is a prominent location that influences Ca2+ permeability of α4β2* AChRs (Tapia et al., 2007). In that location, a negatively charged α4 Glu 261 promotes Ca2+ permeability, as does Glu 261 of α5, whereas the positively charged β2 Lys 258 inhibits it. The cytoplasmic amphipathic helix has been shown to influence α4β2 AChR conductance by forming a vestibule to the channel with lateral windows through which cations must pass (Hales et al., 2006). In such a region, the negatively charged Asp 398 might promote Ca2+ permeability, whereas Asn 398, in which an amide group has replaced the negatively charged carboxyl group of aspartate, might inhibit it.
We report that the α5 Asn 398 high-risk variant has lower Ca2+ permeability and desensitizes more quickly than the Asp 398 variant in (α4β2)2α5 but not in (α3β4)2α5 or (α3β2)2α5 AChRs. This suggests that a positive allosteric modulator (PAM) would enhance nicotine replacement therapy for those with the Asn 398 risk variant. A PAM might achieve the benefits of increasing the amount of (α4β2)2α5 AChRs without the potential liabilities of increasing the amount of α5.
We report that α5 D398N variation does not alter sensitivity to activation by nicotine. The high sensitivity to activation and desensitization of (α4β2)2α5 AChRs by nicotine results in a narrow concentration range in which activation and desensitization concentration/response curves overlap. This region centers on 0.2 μM, a concentration of nicotine typically sustained by smokers (Benowitz, 1996). This concentration would desensitize 60% of (α4β2)2α5 AChRs and permit smoldering activation of the remainder. The low sensitivity to activation and desensitization of (α3β4)2α5 AChRs by nicotine results in a broad region of overlap of concentration/response curves centered near 10 μM. At nicotine concentrations sustained in smokers, negligible activation or desensitization of these AChRs would occur.
Materials and Methods
cDNAs and cRNAs.
The cDNAs for human α3, β2, and α4 subunits were cloned in this laboratory (Gerzanich et al., 1998; Wang et al., 1998). The cDNA for human α5 (Asp 398) was kindly provided by Dr. Francesco Clementi (University Degli Studi di Milano, Milano, Italy). Synthesis of tandem constructs of β2(AGS)6α4 and α4(AGS)12β2 (Zhou et al., 2003) were described previously. Mutation Asn 398 was introduced into the human α5 subunit using the QuikChange site-directed mutagenesis kit (Stratagene, La Jolla, CA) and sequenced to verify that only the desired mutation was present. To prepare the β2(AGS)6α4(AGS)12β2(AGS)6α4 tetrameric concatamer, β2(AGS)6α4 was cut with SalI and NsiI, α4(AGS)12β2 with NsiI and BamHI, and β2(AGS)6α4 with BamHI and SalI. Ligation of the SalI-NsiI fragment from β2(AGS)6α4 with the NsiI-BamHI fragment from α4(AGS)12β2 into BamHI-SalI vector from β2(AGS)6α4 produced the β2(AGS)6α4(AGS)12β2(AGS)6α4 tetramer. This concatamer is referred to hereafter as β2-α4-β2-α4. cRNAs from linearized cDNA templates were synthesized in vitro using the SP6 mMessage mMachine kit (Ambion, Austin, TX). All chemicals were purchased from Sigma-Aldrich (St. Louis, MO), unless otherwise noted.
Oocyte Removal and Injection.
Oocytes were removed surgically from Xenopus laevis and placed in a solution containing 82.5 mM NaCl, 2 mM KCl, 1 mM MgCl2, and 5 mM HEPES, at pH 7.5. They were defolliculated in this buffer containing 2 mg/ml collagenase type IA (Sigma, St. Louis, MO) for 1.5 h. After defolliculation, oocytes were incubated at 18°C in semisterile L-15 medium (Invitrogen, Carlsbad, CA) diluted by half in 10 mM HEPES buffer with 10 U/ml penicillin and 10 μg/ml streptomycin, pH 7.5.
Oocytes were injected with 5 ng of the β2-α4-β2-α4 concatamer cRNA and 2.5 ng of α5 cRNA. (α3β2)2α5 or (α3β4)2α5 AChRs were expressed using free subunits: 3 ng of α3, 3 ng of β2 or β4, and 6 ng of α5. Function was assayed 4 to 6 days after injection of cRNAs. Oocytes were voltage-clamped at −50 mV. Current amplitudes were normalized to maximum ACh responses and averaged for four to five oocytes.
Electrophysiological Recordings and Data Analysis.
Four to six days after injection, whole-cell membrane currents evoked by ACh were recorded in oocytes at room temperature (∼22°C) with a standard two-electrode voltage-clamp amplifier (Oocyte Clamp OC-725; Warner Instrument, Hamden, CT). All recordings were performed at a holding potential of −50 mV. All perfusion solutions contained 0.5 μM atropine to block responses of endogenous muscarinic AChRs that might be present in oocytes. Agonists were applied by means of a set of 2-mm glass tubes directed to the animal pole of the oocytes.
The “Ca2+-only” currents measured in response to 10 μM ACh for (α4β2)2α5 and (α3β2)2α5 or 1000 μM for (α3β4)2α5 reflect only Ca2+ flow through AChR channels. The maximum ACh responses were normalized to the peak currents induced in a Cl−-free version of normal ND-96 buffer composed of 90 mM NaOH, 2.5 mM KOH, 1.8 mM Ca(OH)2, and 10 mM HEPES buffered to pH 7.3 with methanesulfonic acid. Intracellular electrodes in this case were filled with 2.1 M potassium methanesulfonate. The “Ca2+-only” solution was composed of 1.8 mM Ca(OH)2 adjusted to pH 7.5 with HEPES as well as 178 mM dextrose to preserve osmolarity. Data are means ± S.E. for five oocytes for each AChR subtype.
The Hill equation was fitted to the concentration-response relationship using a nonlinear least-squares error curve-fit method (Kaleidagraph; Abelbeck/Synergy, Reading, PA): I(x) = Imax [xnH/(xnH + EC50nH)], where I(x) is the peak current measured at the agonist concentration x, Imax is the maximal current peak at the saturating agonist concentration, EC50 is the agonist concentration required for the half-maximal response, and nH is the Hill coefficient.
Cell Culture.
The human embryonic kidney tsA201 cell lines expressing human (α4β2)2α5, (α3β2)2α5, or (α3β4)2α5 AChRs were described previously (Wang et al., 1998; Kuryatov et al., 2008). All cell lines were maintained in Dulbecco's modified Eagle's medium (high glucose; Invitrogen) supplemented with 10% fetal bovine serum (HyClone, Logan, UT), 100 units/ml penicillin, 100 μg/ml streptomycin, and 2 mM l-glutamine (Invitrogen) at 37°C, 5% CO2 at saturating humidity.
FLEXstation Experiments.
AChR function was determined in the cell lines using a FLEXstation II (Molecular Devices, Sunnyvale, CA) bench-top scanning fluorometer as described by Kuryatov et al. (2005). The day before the experiment, cells were plated at 105/well on black-walled, clear-bottomed 96-well plates (Costar; Corning Life Sciences, Acton, MA). The membrane potential kit (Molecular Devices, Sunnyvale, CA) was used according to the manufacturer's protocol. Serial dilutions of drugs were prepared in V-shaped 96-well plates (Thermo Fisher Scientific, Waltham, MA) and were added in separate wells. Each point on the curves represents the average of four peak responses from different wells.
Statistics.
Data sets are expressed as means ± S.E. Statistical significance between data were determined by Student's t test. Differences were considered significant at the level of p ≤ 0.05.
Results
Conductance and Agonist Sensitivities of α5 Variants in α3* and α4* AChRs Expressed in X. laevis Oocytes.
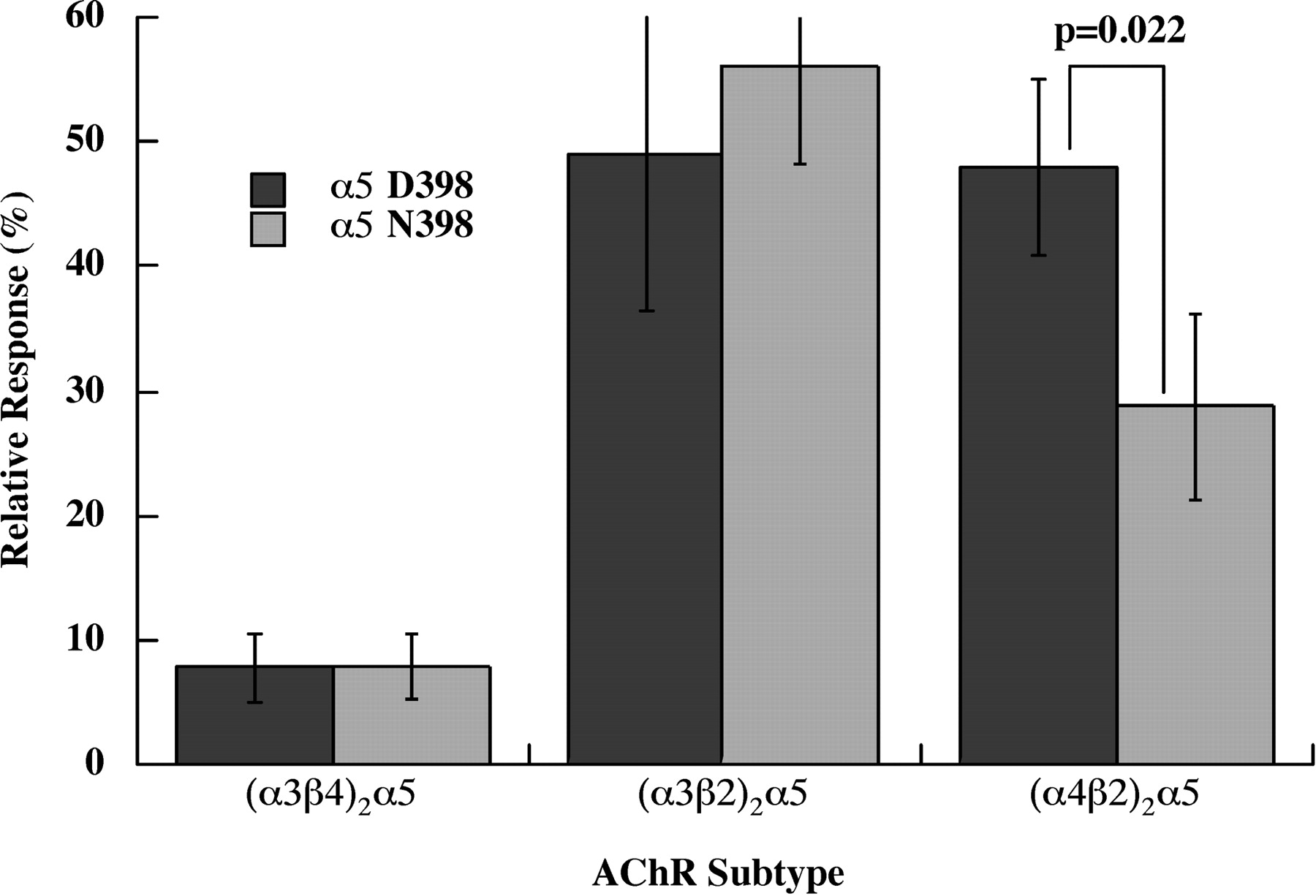
The α5 subunit variant Asn 398 causes reduced Ca2+ permeability in (α4β2)2α5 AChRs, but not in (α3β2)2α5 or (α3β4)2α5 AChRs (Fig. 1). Human AChR subunit mRNAs were expressed in X. laevis oocytes, and responses were assayed electrophysiologically under voltage-clamp conditions. Concatameric β2-α4-β2-α4 subunits plus free α5 were used to insure homogeneous populations of (α4β2)2α5 AChRs, free of any contamination with high agonist sensitivity/low Ca2+ permeability (α4β2)2β2 AChR subtypes or low agonist sensitivity/high Ca2+ permeability (α4β2)2α4 AChR subtypes (Tapia et al., 2007). The α5 accessory subunits, either the Asp 398 allele or the Asn 398 risk allele, were added to the concatamer in equal molar ratio. The concatamer alone at these concentrations gives virtually no response. For α3* AChRs, free subunits were used in a 1:1:2 ratio of α3/β/α5, with the excess α5 intended to insure that all of the AChRs had the subunit composition (α3β)2α5. The currents provoked by a saturating concentration of ACh were measured in solutions lacking Cl− to eliminate currents through Cl− channels. Both a conventional cation mix and a solution containing only Ca2+ were used. The relative response was calculated as the ACh-induced current observed with Ca2+ as the only cation as a percentage of the current in the normal cation mix. Ca2+ permeabilities of both (α3β2)2α5 and (α4β2)2α5 subtypes were quite high. The α5 D398N variation affected only (α4β2)2α5 and not (α3β2)2β5. (α4β2)2α5 Asn 398 had 40% lower Ca2+ permeability than (α4β2)2α5 Asp 398.
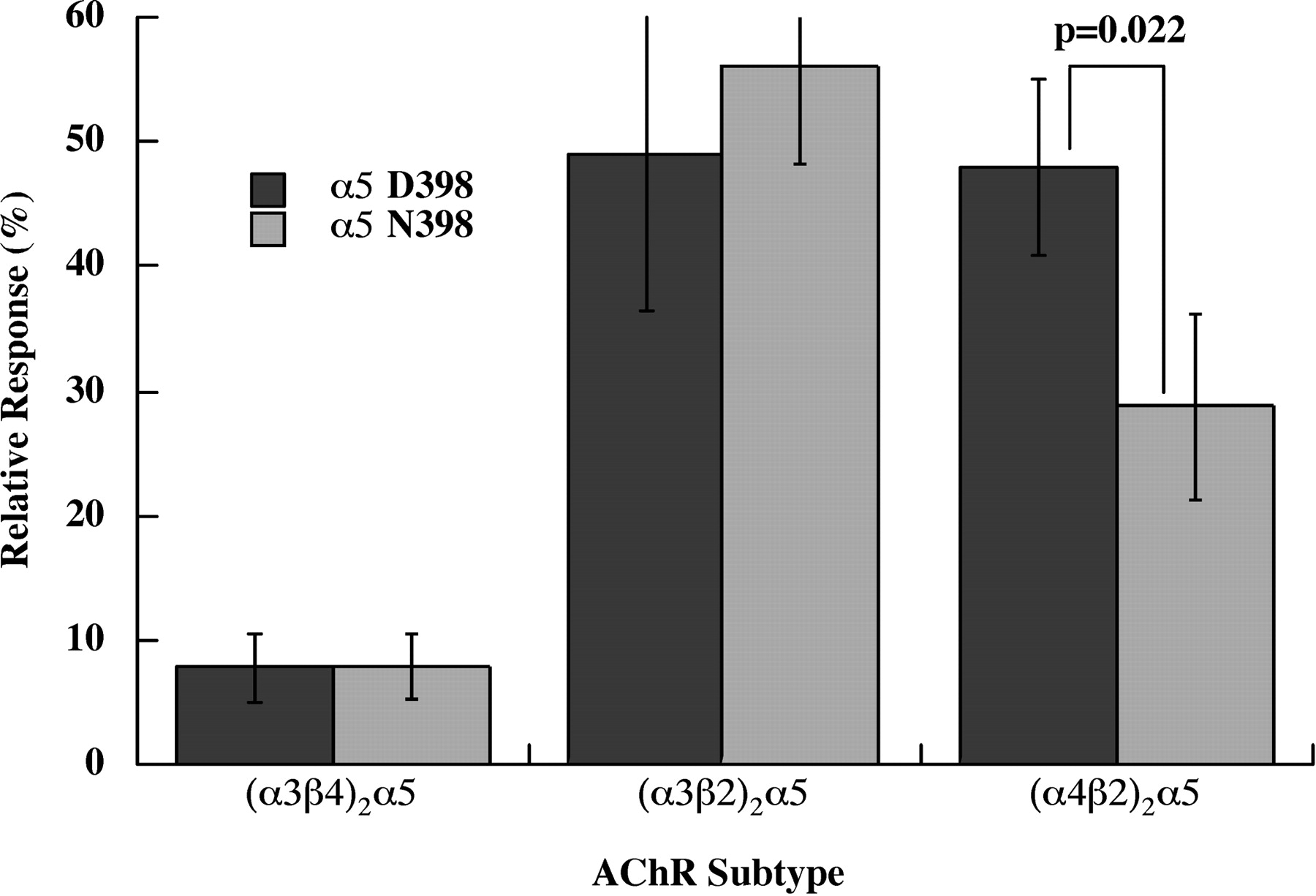
α5 Asn 398 exhibited lower Ca2+ permeability than α5 Asp 398 in (α4β2)2α5 AChRs, but this variation had no effect on Ca2+ permeabilities of α3* AChRs. Human AChRs were expressed in X. laevis oocytes. (α4β2)2α5 AChRs were expressed using a tetrameric concatamer β2-α4-β2-α4 plus free α5 subunits. α3* AChR subtypes used all free subunits. The relative response is the current obtained using 1.8 mM Ca2+ as the sole cation compared with the current obtained using a buffer containing 90 mM Na+, 2.5 mM K+, and 1.8 mM Ca2+. Thus, currents reflect only Ca2+ flow through AChRs. Maximum responses were obtained using 10 μM ACh for the sensitive (α3β2)2α5 and (α4β2)2α5 subtypes and 1000 μM ACh for the less sensitive (α3β4)2α5 subtype.
The α5 D398N variation did not alter sensitivity to activation by agonists (Fig. 2). (α4β2)2α5 AChRs in the α5 Asp 398 and α5 Asn 398 forms were equally sensitive, respectively, to ACh (EC50 = 1.64 ± 0.06 or 1.44 ± 0.09 μM), nicotine (EC50 = 0.499 ± 0.063 or 0.640 ± 0.115 μM), or varenicline (EC50 = 0.0235 ± 0.0059 or 0.0440 ± 0.0098 μM) (p = 0.32). (α3β2)2α5 AChRs in the α5 Asp 398 and Asn 398 forms were equally sensitive, respectively, to ACh (EC50 = 1.83 ± 0.07 or 1.70 ± 0.09 μM). Likewise, the α5 Asp 398 and Asn 398 forms of (α3β4)2α5 AChRs were equally sensitive to ACh (EC50 = 122 ± 17 or 115 ± 20 μM).

Assay of effects of the α5 D398N variation on sensitivity to activation by ACh and short-term desensitization of the (α4β2)2α5, (α3β2)2α5, and (α3β4)2α5 AChR subtypes. Human AChRs were expressed in X. laevis oocytes as in Fig. 1. The (α4β2)2α5 subtype is shown in A to C. A, there is no significant difference in sensitivity to activation by ACh in the two α5 variants. Each response is the average of four to five oocytes. B and C, kinetics of short-term desensitization in a single oocyte expressing each variant exposed to some of the ACh concentrations used in the dose/response curve. The averaged desensitization of four to five oocytes at 3 s after application of ACh are shown in Table 1, revealing that the N variant desensitizes significantly more. The responses in the dose-response curve were the peak responses. The (α3β2)2α5 subtype is shown in D to F. D, there is no significant difference in sensitivity to activation in the two α5 variants. E and F, kinetics of desensitization. It is much faster and more extensive than in (α4β2)2α5 AChRs. The extent of desensitization by the two α5 variants in the (α3β2)2α5 subtype is not significant, as shown in Table 1. The (α3β4)2α5 subtype is shown in G to I. G, there is no significant difference in sensitivity to activation in the two α5 variants. H and I, kinetics of desensitization. It is slower and less extensive than in (α3β2)2α5 AChRs. The extent of desensitization by the two α5 variants is not significant, as shown in Table 1.
Short-term desensitization varied greatly between (α4β2)2α5, (α3β2)2α5, and (α3β4)2α5 AChR subtypes. The D398N variation in α5 had an effect only in the (α4β2)2α5 subtype (Fig. 2, Table 1). With the (α4β2)2α5 subtype, over 3 seconds, 3 μM ACh caused 22% desensitization with respect to the peak response with the Asp 398 variant, but 46% desensitization with the Asn 398 variant. Thus, there is a significant (p = 0.015) difference in short-term desensitization of this subtype in the two α5 variants. The apparent extent of desensitization of the (α3β2)2α5 subtype under the same conditions was much greater (86.9% for Asp 398), with little difference between α5 variants (89.7% for Asn 398). The faster and more extensive desensitization of the (α3β2)2α5 subtype would cause the integrated currents mediated by this subtype to be much less than those of the (α4β2)2α5 subtype. Thus, with presynaptic AChRs tonically activated to promote transmitter release, (α4β2)2α5 AChRs would have a much larger effect than (α3β2)2α5 AChRs. Another striking subtype-specific difference of α3β2 from α4β2 or α3β4 AChRs is that nicotine causes channel block of α3β2 AChRs, resulting in it behaving as a partial agonist (Rush et al., 2002). The apparent extent of desensitization of the (α3β4)2α5 subtype at 3 s after 100 μM ACh (which similarly approximates the EC50 for this lower sensitivity subtype) is only 8 to 15%.
Effect of α5 D398N Variants on extent of acute desensitization
Using ACh concentrations that approximate the EC50 values for each subtype [3 μM for (α4β2)2α5 and (α3β2)2α5, 100 μM for (α3β4)2α5], the percentage desensitization at 3 s after the application of ACh was calculated by comparing the peak current to the current after 3 s. Each value is the average of four to five oocytes. The extent of desensitization was significantly different between the D398N variants only in the case of the (α4β2)2α5 subtype.
Sensitivities to Activation and Desensitization by Sustained Exposure to Nicotine of (α4β2)2α5, (α3β2)2α5, and (α3β4)2α5 AChRs Expressed in Permanently Transfected Cell Lines.
Smokers sustain concentrations of nicotine ≤0.2 μM in blood for many hours (Benowitz, 1996). Under these conditions, sensitive AChR subtypes may be substantially desensitized, whereas less sensitive subtypes may be affected little by such low concentrations of nicotine. Permanently transfected cell lines are better than oocytes for studies of prolonged desensitization because many parallel cultures can be treated with nicotine for hours to achieve the equilibrium desensitization expected in vivo from exposure to sustained concentrations of nicotine. We prepared permanently transfected human embryonic kidney cell lines that express (α4β2)2α5, (α3β2)2α5, and (α3β4)2α5 AChRs using the Asp 398 α5 variant and used these to compare the effects of activation and desensitization by nicotine on these subtypes (Fig. 3). The similar short-term activation and desensitization properties of AChR subtypes with the Asn 398 α5 variant for the (α3β2)2α5 and (α3β4)2α5 subtypes suggest that this variation would not alter the fundamental subtype desensitization properties associated with prolonged exposure to nicotine. Future studies using the (α4β2)2α5 Asn 398 variant might reveal more extensive desensitization in this variant.
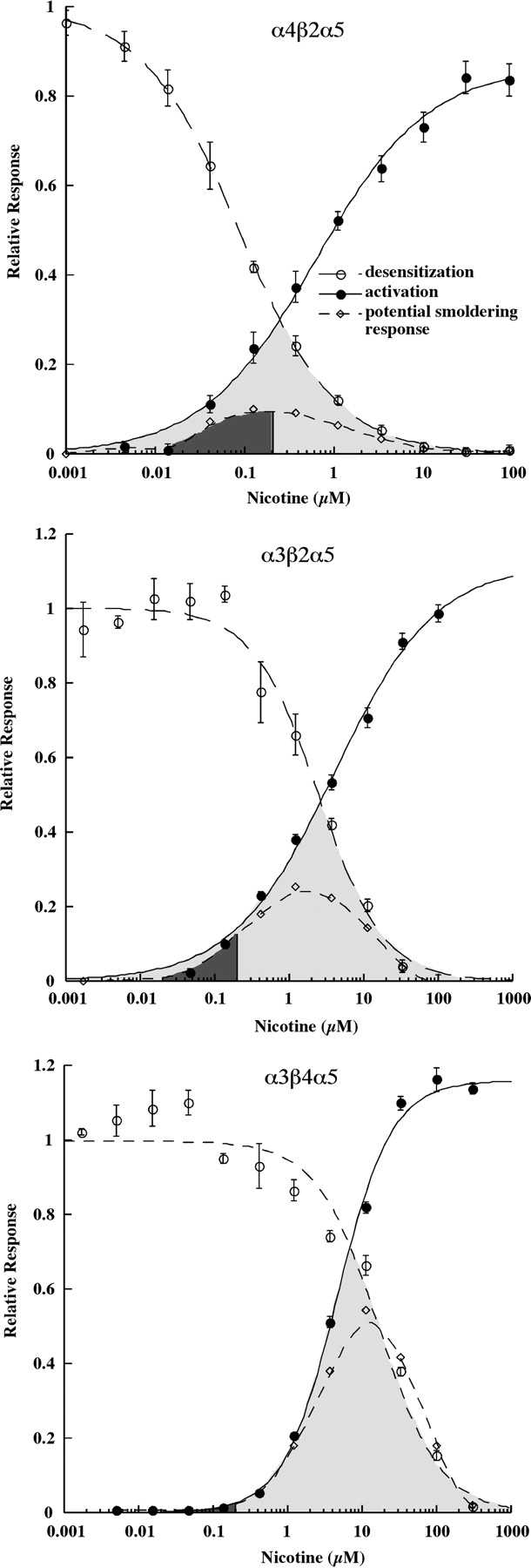
Nicotine sensitivities differ dramatically among the (α4β2)2α5, (α3β2)2α5, and (α3β4)2α5 subtypes. Human AChRs were expressed in permanently transfected human embryonic kidney cell lines (Wang et al., 1998; Kuryatov et al., 2008). Responses were assayed in microwell cultures using a Flexstation microplate fluorometer and a fluorescent probe sensitive to membrane potential. Desensitization was measured after 1.5 h. This allows enough time for equilibrium desensitization but not enough time for up-regulated AChRs to be expressed on the cell surface. The fraction of AChRs remaining activatable after desensitization was assayed using 1 mM ACh. The overlapping region between concentration/response curves for desensitization and activation is highlighted in gray. AChRs remaining undesensitized could undergo smoldering activation in this concentration range. The potential smoldering response at each concentration of nicotine was calculated by multiplying the short-term response to nicotine at that concentration by the fractional response remaining after desensitization. The area under the smoldering response curve below 0.2 μM was blackened to indicate the range of responses that might be sustained in smokers.
The (α4β2)2α5 AChR subtype is sensitive to activation by nicotine (EC50 = 0.620 ± 0.107 μM) (Fig. 3). It is also quite sensitive to desensitization (IC50 = 0.0872 ± 0.0027 μM). The overlap between the activation and desensitization curves is shaded. This indicates the range of nicotine concentrations at which, at equilibrium, “smoldering activation” could be sustained as a result of significant activation despite a substantial degree of desensitization of the AChR population. This zone of smoldering activation is approximately centered on 0.2 μM nicotine, a concentration typically sustained in smokers' blood (Benowitz, 1996). The potential amplitude of the smoldering response at each nicotine concentration was calculated by multiplying the relative short-term response by the relative response remaining after desensitization. The area beneath this curve at concentrations ≤0.2 μM nicotine was blackened to indicate the sustained amplitude of response that would be expected from the subtype at nicotine concentrations likely to be sustained in smokers. In the case of (α4β2)2α5 AChRs, in the presence of 0.2 μM nicotine, sustained smoldering activation equivalent to 9% of the maximum short-term response would be sustained. This tonic effect on membrane potential or calcium concentration in synaptic endings, where these AChRs modulate transmitter release, might substantially promote transmitter release. Most α4β2* AChRs are thought to be located presynaptically (Gotti et al., 2007). The sustained 60% desensitization caused by 0.2 μM nicotine would be likely to substantially impair short-term synaptic transmission at synapses where postsynaptic (α4β2)2α5 AChRs were required for mediating transmission of patterns of short-term signals. If the Asn 398 α5 form produced more extensive desensitization, as suggested by the results shown in Fig. 2 B and C, and Table 1, this would move the equilibrium desensitization curve shown in Fig. 3 to the left. The net effects of this would be to narrow the range of potential smoldering activation, to reduce the amplitude of smoldering activation, to reduce transmitter release potentiated by presynaptic (α4β2)2α5 AChRs, and, through greater desensitization, to more completely block transmission at synapses at which this subtype served a postsynaptic role.
The (α3β2)2α5 AChR subtype is 4.7-fold less sensitive to activation (EC50 = 2.91 ± 0.35 μM) and 34-fold less sensitive to desensitization (IC50 = 2.92 ± 0.19 μM) by nicotine than is the (α4β2)2α5 AChR subtype (Fig. 3). The overlap between activation and desensitization curves centers around 2 μM nicotine, a concentration unlikely to be reached even transiently in smokers. At 0.2 μM nicotine, the sustained smoldering response of (α3β2)2α5 AChRs would be 13% of the maximum short-term response.
The (α3β4)2α5 AChR subtype is 7.5-fold less sensitive to activation (EC50 = 4.64 ± 0.18 μM) and 192-fold less sensitive to desensitization (IC50 = 16.7 ± 0.8 μM) by nicotine than is the (α4β2)2α5 AChR subtype (Fig. 3). The overlap between activation and desensitization curves is very large, resulting in an extensive range of smoldering activation and a high maximum potential amplitude of smoldering activation. However, the overlap centers around 8 μM, more than 40 times the concentration of nicotine sustained in smokers. Consequently, at 0.2 μM nicotine, a concentration sustained in smokers, there is negligible desensitization or smoldering activation. The lack of desensitization means that transmission is unimpaired at autonomic ganglia synapses, where (α3β4)2α5 AChRs are a major postsynaptic subtype. The functional roles of α3* AChRs in the medial habenula, where they are most abundant in brain, are less clear (Gotti et al., 2007). However, the highest transient nicotine concentrations likely to be found in brain would stimulate very little of the maximum potential short-term response. α3* AChRs in lung might be exposed to larger transient concentrations of nicotine.
Discussion
We observed that the α5 Asn 398 variant had lower Ca2+ permeability and greater desensitization than the α5 Asp 398 variant in (α4β2)2α5 AChRs but not in (α3β2)2α5 or (α3β4)2α5 AChRs. α3, β4, and α5 subunits are organized sequentially in the genome and expressed together in the major autonomic ganglia postsynaptic AChR subtype (α3β4)2α5 (Gotti et al., 2007). All α3* AChR subtypes make up 3.5% of brain AChRs, whereas α4β2* AChR subtypes make up >45% (Marks et al., 2010). α4β2*AChRs are important for nicotine addiction. Knockout of α4, β2, or α6 AChRs prevents nicotine self-administration, and re-expression of the knocked out subunit in the ventral tegmental area restores self-administration (Pons et al., 2008). (α4β2)2α5 AChRs make up 10 to 37% of brain α4β2* AChRs in various regions (Brown et al., 2007; Mao et al., 2008). These primarily presynaptic AChRs promote the release of various transmitters. Decreased Ca2+ permeability and increased desensitization in the α5 Asn 398 variant associated with increased risk for tobacco addiction and lung cancer might result in decreased release of a transmitter in which release is modulated by (α4β2)2α5 AChRs (e.g., GABA) (McClure-Begley et al., 2009). The α5 D398N variation did not alter sensitivity to agonist activation of α3* or α4* AChRs. This is not surprising, because ACh binding sites are in the extracellular surface, whereas α5 398 is in the cytoplasmic surface. α5 398 is not part of the cation channel, which has a major role in determining Ca2+ permeability (Tapia et al., 2007). However, it is in a cytoplasmic region thought to form access portals to the channel, which have been shown to have large influence on conductance (Hales et al., 2006). Ion selectivity filters have been found throughout the extracellular vestibule of the AChR, and similar considerations may apply to the cytoplasmic vestibule (Sine et al., 2010).
We showed that (α4β2)2α5 AChRs are sensitive to activation and desensitization by nicotine concentrations sustained in smokers. Thus, this subtype is likely to be important in mediating effects of nicotine. Smoldering activation of (α4β2)2α5 AChRs by nicotine concentrations that desensitize a large fraction of these AChRs may contribute significantly to increasing presynaptic Ca2+ concentrations and depolarization, thereby promoting transmitter release in synapses at which tonic activation of this subtype modulates transmitter release. Extensive desensitization of this subtype by nicotine concentrations in smokers would impair transmission at synapses, where it served a postsynaptic role. By contrast, (α3β4)2α5 AChRs were shown to be very insensitive to activation or desensitization by concentrations of nicotine sustained in smokers. Thus, this subtype is unlikely to be important in mediating effects of nicotine. (α3β2)2α5 AChRs exhibited intermediate sensitivities to activation and desensitization. AChR subtypes containing α5 in the medial habenula may play roles in the effects of nicotine (Salas et al., 2009).
Genome-wide association studies have shown that risk for tobacco addiction and lung cancer are associated not only with the α5 Asn 398 variant but also with variants that reduce the amount of α5 expressed (Wang et al., 2009). Reducing the amount of the (α4β2)2α5 subtype would compound the loss of function produced by decreased Ca2+ permeability and increased desensitization.
An (α4β2)2α5 AChR-selective PAM could compensate for the deficit in function associated with risk for nicotine addiction and lung cancer. Knockout of α5 subunits increases nicotine reward and decreases aversion to high doses of nicotine (Jackson et al., 2010). A PAM could provide genetically based personalized medical therapy for those with the α5 Asn 398 variant when combined with nicotine replacement therapy using nicotine or varenicline. Such a PAM might also have cognitive enhancing effects on those not at risk. The α5 subunit plays a key role in attention circuitry (Bailey et al., 2010). An α4β2α5 AChR PAM might enhance nicotinic antinociception (Jackson et al., 2010). A PAM would specifically enhance the impact of ACh, especially where volume transmission is mediated by low concentrations of ACh. The effects of an (α4β2)2α5-selective PAM would differ from those of agonists such as nicotine or varenicline, which are much less subtype-selective. More importantly, a primary effect of such high-affinity agonist drugs is to desensitize AChRs, thereby blocking the normal ACh effect. Even if smoldering activation of nondesensitized AChRs were produced by agonist drugs to emulate the tonic effects of ACh, desensitization would also block transmission of information inherent in the changes in ACh concentration produced by changes in the firing rates of cholinergic neurons (Sarter et al., 2009). By contrast, a PAM would leave this information intact and amplify it.
PAMs have been discovered that are selective for AChR subtypes (Bertrand and Gopalakrishnan, 2007). For example, N-(5-chloro-2,4-dimethoxyphenyl)-N′-(5-methyl-3-isoxazolyl)-urea (PNU 120596) is highly selective and potent on α7 AChRs (Hurst et al., 2005; Young et al., 2008). Remarkably, it can activate completely desensitized AChRs. Galanthamine is a weak PAM on many AChR subtypes (Samochocki et al., 2003). We found, using cell lines that express this subtype and other α4β2* human AChR subtypes, that it was relatively selective for (α4β2)2α5 AChRs (Kuryatov et al., 2008). It is very likely that potent PAMs selective for (α4β2)2α5 AChRs can be discovered. The unique interface between α5 and α4 subunits, or the interface between β2 and α5 subunits, are sites at which such PAMs might be directed. Benzodiazepines, which act as PAMs on GABAA receptors, bind at an αγ interface in these receptors (Hanson and Czajkowski et al., 2008). Galanthamine binds at such an interface in ACh binding protein (Hansen and Taylor, 2007). An intrasubunit PAM site on α5, homologous to the α7-selective transmembrane site for PNU 120596 (Young et al., 2008), could potentiate all α5* AChR subtypes, yet perhaps be useful. In any case, the α4β2α5 cell line used here could be employed with assays using fluorescent indicators of membrane potential in high throughput screens to select for PAMs specific for (α4β2)2α5 AChRs.
A PAM might act by increasing the probability of activation of a liganded AChR, or by reducing desensitization, or both. The effects of a PAM could be analyzed using overlapping dose/response curves for activation and desensitization by nicotine as in Fig. 3. The ideal PAM would effect only the (α4β2)2α5 subtype. It would probably shift the activation curve to the left, shift the desensitization curve to the right, or both. The net effect could be to greatly increase the range of overlap of these curves at which smoldering activation could take place and increase the amplitude of the maximum response sustained by nicotine. This would result in greater release of transmitters modulated presynaptically by nicotine. Reduced desensitization would reduce impairment of synaptic transmission mediated by postsynaptic (α4β2)2α5 AChRs. Both increased activation and decreased desensitization would increase the activation of (α4β2)2α5 AChRs by ACh in the absence of nicotine.
Authorship Contributions
Participated in research design: Lindstrom and Kuryatov.
Conducted experiments: Kuryatov.
Performed data analysis: Kuryatov.
Wrote or contributed to the writing of the manuscript: Lindstrom, Kuryatov, and Berrettini.
Other: Lindstrom and Berrettini acquired funding for the research.
Acknowledgments
We thank Barbara Campling for her comments on the manuscript.
Footnotes
This work was supported by the National Institutes of Health National Institute of Neurological Disorders and Stroke [Grant NS11323] and the National Institutes of Health National Institute on Drug Abuse [Grant DA025201].
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
doi:10.1124/mol.110.066357.
-
ABBREVIATIONS:
- AChR
- acetylcholine receptor
- PAM
- positive allosteric modulator
- ACh
- acetylcholine
- PNU 120596
- N-(5-chloro-2,4-dimethoxyphenyl)-N′-(5-methyl-3-isoxazolyl)-urea.
- Received May 14, 2010.
- Accepted September 29, 2010.
- Copyright © 2011 The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}