


Abstract
Positive and negative allosteric modulators (PAMs and NAMs, respectively) of the type 5 metabotropic glutamate (mGlu5) receptor have demonstrable therapeutic potential in an array of neurological and psychiatric disorders. Here, we have used rat cortical astrocytes to investigate how PAMs and NAMs mediate their activity and reveal marked differences between PAMs with respect to their modulation of orthosteric agonist affinity and efficacy. Affinity cooperativity factors (α) were assessed using [3H]2-methyl-6-(phenylethynyl)-pyridine (MPEP)-PAM competition binding in the absence and presence of orthosteric agonist, whereas efficacy cooperativity factors (β) were calculated from net affinity/efficacy cooperativity parameters (αβ) obtained from analyses of the abilities of PAMs to potentiate [3H]inositol phosphate accumulation in astrocytes stimulated with a submaximal (EC20) concentration of orthosteric agonist. We report that whereas 3,3′-difluorobenzaldazine (DFB) and 3-cyano-N-(1,3-diphenyl-1H-prazol-5-yl)benzamide (CDPPB) primarily exert their allosteric modulatory effects through modifying the apparent orthosteric agonist affinity at the astrocyte mGlu5 receptor, the effects of S-(4-fluoro-phenyl)-{3-[3-(4-fluoro-phenyl)-[1,2,4]oxadiazol-5-yl]-piperidinl-1-yl}-methanone (ADX47273) are mediated primarily via efficacy-driven modulation. In [3H]MPEP-NAM competition binding assays, both MPEP and 2-(2-(3-methoxyphenyl)ethynyl)-5-methylpyridine (M-5MPEP) defined similar specific binding components, with affinities that were unaltered in the presence of orthosteric agonist, indicating wholly negative efficacy-driven modulations. It is noteworthy that whereas M-5MPEP only partially inhibited orthosteric agonist-stimulated [3H]inositol phosphate accumulation in astrocytes, it could completely suppress Ca2+ oscillations stimulated by quisqualate or (S)-3,5-dihydroxyphenylglycine. In contrast, MPEP was fully inhibitory with respect to both functional responses. The finding that M-5MPEP has different functional effects depending on the endpoint measured is discussed as a possible example of permissive allosteric antagonism.
Introduction
The major excitatory neurotransmitter glutamate exerts its actions in the central nervous system through binding to two distinct classes of cell-surface receptor: the ionotropic and metabotropic glutamate (mGlu) receptors. The eight mammalian mGlu receptors are family C G protein-coupled receptors, with mGlu1 and mGlu5 receptors constituting the group I subfamily (Conn and Pin, 1997; Niswender and Conn, 2010). Group I mGlu receptors are widely expressed in the mammalian central nervous system, and in neurons, they have a predominantly postsynaptic localization. mGlu5 receptors are also expressed in glia, particularly in astrocytes, and the expression of the mGlu5 receptor subtype in astrocytes has been highlighted with respect to a number of potential physiological and pathophysiological roles (Verkhratsky and Kirchhoff, 2007; Wierońska and Pilc, 2009).
Group I mGlu receptors couple via Gαq/11 proteins to regulate phospholipase C activity, as well as mediating G protein-dependent and -independent effects on ion channels and other cellular effector proteins (Hermans and Challiss, 2001; Gerber et al., 2007). Despite sequence, structure, and coupling similarities, it is generally believed that mGlu1 and mGlu5 receptors subserve distinct functional roles within the brain (Hermans and Challiss, 2001; Bonsi et al., 2008). One consistently reported signal transduction difference between mGlu1 and mGlu5 receptors is the robust Ca2+ oscillation initiated by mGlu5, not generally observed after mGlu1 receptor activation (Kawabata et al., 1996; Nakahara et al., 1997; Nash et al., 2001, 2002; Bradley et al., 2009). Work by us and others has provided evidence for the mGlu5 receptor causing Ca2+ oscillations via a “dynamic uncoupling” mechanism (Nash et al., 2002), involving a reversible, protein kinase C-dependent phosphorylation of the receptor at Ser839 (Kim et al., 2005), essentially turning Gq/11/phospholipase C/Ca2+ signaling on and off to create the oscillatory pattern. This unusual mechanism of generating Ca2+ oscillations endows this system with specific properties. For example, whereas changes in mGlu5 receptor expression can alter Ca2+ oscillation frequency, this readout is essentially insensitive to changes across the stimulatory concentration range of an orthosteric agonist (e.g., glutamate) (Nash et al., 2002; Bradley et al., 2009). In contrast, allosteric modulation can concentration-dependently increase [positive allosteric modulator (PAM)] or decrease [negative allosteric modulator (NAM)] orthosteric agonist-initiated Ca2+ oscillation frequency in both recombinant (CHO-lac-mGlu5) and native (rat cortical astrocyte) expression systems (Nash et al., 2002; Bradley et al., 2009). Therefore, allosteric modulators can achieve effects beyond the pharmacological repertoire of orthosteric ligands, allowing them to “retune” the Ca2+ oscillation frequency generated by mGlu5 receptor (orthosteric) occupancy.
An array of molecules have been reported over the past 10 years to interact allosterically with the mGlu5 receptor (Marino and Conn, 2006; Rodriguez and Williams, 2007; Conn et al., 2009); these include NAMs, such as 2-methyl-6-(phenylethynyl)-pyridine (MPEP) and 2-(2-(3-methoxyphenyl)ethynyl)-5-methylpyridine (M-5MPEP); PAMs, such as 3,3′-difluorobenzaldazine (DFB), N-{4-chloro-2-[(1,3-dioxo-1,3-dihydro-2H-isoindol-2-yl)methyl]phenyl}-2-hydroxybenzamide (CPPHA), 3-cyano-N-(1,3-diphenyl-1H-pyrazol-5-yl)benzamide (CDPPB), and ADX47273; and the neutral allosteric modulator 2-(2-(3-methoxyphenyl)ethynyl)-5-methylpyridine (5MPEP) (Gasparini et al., 1999; O'Brien et al., 2003, 2004; Kinney et al., 2005; Rodriguez et al., 2005; Chen et al., 2007; Liu et al., 2008). Positive or negative allosteric modulation of mGlu5 receptor function has been postulated as an approach for the treatment of a number of pathological conditions, including anxiety (Swanson et al., 2005), schizophrenia (Marino and Conn, 2006), and fragile X syndrome (Dölen and Bear, 2008).
Here, we have investigated orthosteric/allosteric interactions at the mGlu5 receptor, primarily in rat cortical astrocytes, with a particular objective of applying quantitative pharmacological principles to determine the nature of the agonist/modulator interaction and to extend our previous work examining how NAMs and PAMs modulate mGlu5 receptor function at the single cell level.
Materials and Methods
Compounds.
Tissue culture reagents, G5 supplement, and Fura-2 acetoxymethyl ester were from Invitrogen (Paisley, UK). [myo-3H]Inositol was from GE Healthcare (Chalfont St. Giles, Buckinghamshire, UK). l-Quisqualic acid, l-glutamic acid, MPEP, DFB, and (S)-3,5-dihydroxyphenylglycine (DHPG) were obtained from Tocris Cookson (Bristol, UK). CPPHA, CDPPB, S-(4-fluoro-phenyl)-{3-[3-(4-fluoro-phenyl)-[1,2,4]oxadiazol-5-yl]-piperidinl-1-yl}-methanone (ADX47273), and 5-methyl-2-(phenylethynyl)pyridine (5MPEP) were synthesized in-house by GlaxoSmithKline (Harlow, UK). M-5MPEP was a kind gift from Dr. P. J. Conn (University of Nashville, Nashville, TN). [3H]MPEP was from American Radiolabeled Chemicals (Stevenage, UK). All other chemicals and reagents were purchased from Sigma-Aldrich (Poole, Dorset, UK).
Astrocyte Preparation and Culture.
Rat cortical astrocytes were prepared as described previously (Bradley et al., 2009). In brief, neocortices from 2- to 3-day-old Wistar rat pups were subject to proteolysis and dissociation and centrifuged at 1000 rpm for 8 min. Cells were resuspended in poly(d-lysine)-coated T175 tissue culture flasks in Dulbecco's modified Eagle's medium containing GlutaMAX-1 with sodium pyruvate, 4500 mg/l glucose, 15% dialyzed fetal bovine serum, 2.5 μg/ml amphotericin B, 105 U/ml penicillin, and 100 μg/ml streptomycin. Cell cultures were maintained at 37°C in a humidified 5% CO2/air atmosphere. Medium was replaced after 3 to 4 days in vitro. At 6 days in vitro, medium was replaced, and flasks were shaken overnight (320 rpm at 37°C). After this, confluent cell monolayers were washed twice with PBS (without Ca2+/Mg2+) and harvested with 0.25% (w/v) trypsin and 0.02% (w/v) EDTA. Cells were centrifuged, resuspended in medium (outlined above), and seeded onto poly(d-lysine)-coated borosilicate coverslips for imaging experiments or poly(d-lysine)-coated 24-well plates for [3H]inositol phosphate experiments. After 24 h, medium was replaced with medium devoid of fetal bovine serum containing the G5 supplement. Experiments were performed 2 to 4 days later.
Single-Cell Intracellular Ca2+ Concentration Assay.
Rat cortical astrocytes were seeded onto 22-mm borosilicate coverslips at 300,000 cells/well and grown to approximately 80% confluence in medium containing G5 supplement. Cells were loaded with Fura-2 acetoxymethyl ester (2 μM) in Krebs-Henseleit bicarbonate containing Pluronic acid F127 (0.36 mg/ml) for 45 to 60 min at room temperature. Coverslips were then transferred to the stage of a Nikon Diaphot inverted epifluorescence microscope, with an oil immersion objective (40×) and a SpectraMASTER II module (PerkinElmer Life and Analytical Sciences, Waltham, MA). Cells were excited at wavelengths of 340 and 380 nm using a SpectraMASTER II monochromator, and emission was recorded at wavelengths greater than 520 nm. The ratio of fluorescence intensities at these wavelengths is given as an index of [Ca2+]i. All experiments were performed at 37°C; drug additions were made via a perfusion line.
Total [3H]Inositol Phosphate Accumulation Assay.
Rat cortical astrocytes were seeded at 150,000 cells/well in poly(d-lysine)-coated 24-well plates. The following day, cells were incubated in fresh medium containing G5 supplement and 2.5 μCi/ml [3H]inositol for 48 h. Cell monolayers were washed twice and incubated in Krebs-Henseleit buffer containing GPT (3 U/ml) and pyruvate (5 mM) for 25 min at 37°C. LiCl (10 mM) was added for a further 20 min before agonist incubations for 20 min. Incubations were terminated by aspiration of buffer and rapid addition of 500 μl of ice-cold trichloroacetic acid (0.5 M). After extraction on ice for 20 to 30 min, samples were transferred to tubes containing 100 μl of EDTA (10 mM, pH 7.0) and 500 μl of a 1:1 mixture of tri-n-octylamine and 1,1,2-trichlorofluoroethane added. Samples were centrifuged at 14,000 rpm for 2 min, and 400 μl of the upper aqueous phase was transferred into fresh tubes containing 100 μl of NaHCO3 (62.5 mM). [3H]inositol mono-, bis-, and trisphosphates ([3H]IPx) were recovered by anion exchange chromatography on Dowex AG1-X8 formate columns as described previously (Mistry et al., 2005).
Radioligand Binding.
[3H]MPEP competition binding versus PAMs was conducted as described previously (Liu et al., 2008). In brief, aliquots of membranes (100 μg), prepared from astrocytes or adult rat cortex, were added to tubes containing vehicle or test compounds (final dimethyl sulfoxide concentration, 0.33% in all assay tubes) and [3H]MPEP (2 nM final concentration in 50 mM Tris/HCl, 0.9% NaCl, pH 7.4). Tubes were incubated at room temperature for 60 min, with gentle shaking. Membrane-bound ligand was separated from free ligand by rapid filtration onto GF/B glass microfiber filters presoaked in wash buffer (20 mM HEPES, 2 mM CaCl2, and 2 mM MgCl2, pH 7.2). [3H]MPEP competition binding versus NAMs was conducted as described previously by Anderson et al. (2002). In brief, membranes (100 μg) from astrocytes or adult rat cortex were added to assay buffer (50 mM HEPES and 2 mM MgCl2, pH 7.4) containing approximately 10 nM [3H]MPEP and increasing concentrations of each NAM to be studied. Tubes were incubated for 2 h at 4°C. Membrane-bound ligand was separated from free ligand by rapid filtration onto GF/B glass microfiber filters followed by washing twice with ice-cold wash buffer (50 mM Tris/HCl, pH 7.4). In all cases, membrane-bound radioactivity was extracted overnight and determined by liquid scintillation counting.
Data Analysis.
Concentration-response relationships were analyzed by nonlinear regression using GraphPad Prism 5.0 software (GraphPad Software Inc., San Diego, CA). Where used, eqs. 3 to 5 were fitted to mean datasets and estimates of standard error and confidence intervals are from the best-fit parameters.
For [3H]MPEP saturation binding data, the following equation was globally fitted to nonspecific and total binding data:
 where Y is radioligand binding, Bmax is the total receptor density, [A] is the radioligand concentration, KA is the equilibrium dissociation constant of the radioligand, and NS is the fraction of nonspecific radioligand binding. For radioligand inhibition binding experiments, a one-site binding equation was fitted to the specific binding of each competitive ligand.
where Y is radioligand binding, Bmax is the total receptor density, [A] is the radioligand concentration, KA is the equilibrium dissociation constant of the radioligand, and NS is the fraction of nonspecific radioligand binding. For radioligand inhibition binding experiments, a one-site binding equation was fitted to the specific binding of each competitive ligand.
 where Top and Bottom are the maximal and minimal asymptotes of the curve, respectively, log[B] is the concentration of inhibitor, logIC50 is the logarithm of the concentration of inhibitor that reduces half the maximal radioligand binding for each binding site and nH is the Hill slope (constrained to unity). IC50 values were converted to KA values (equilibrium dissociation constant) using the Cheng and Prusoff (1973) equation. For the inhibition of [3H]MPEP binding by CPPHA in the presence and absence of quisqualate, the following version of a simple allosteric ternary complex model (Lazareno and Birdsall, 1995) was also fitted to inhibition binding data:
where Top and Bottom are the maximal and minimal asymptotes of the curve, respectively, log[B] is the concentration of inhibitor, logIC50 is the logarithm of the concentration of inhibitor that reduces half the maximal radioligand binding for each binding site and nH is the Hill slope (constrained to unity). IC50 values were converted to KA values (equilibrium dissociation constant) using the Cheng and Prusoff (1973) equation. For the inhibition of [3H]MPEP binding by CPPHA in the presence and absence of quisqualate, the following version of a simple allosteric ternary complex model (Lazareno and Birdsall, 1995) was also fitted to inhibition binding data:
 where Y denotes the percentage of specific binding, [A] is the radioligand concentration, KA is the equilibrium dissociation constant of the radioligand, KB denotes the allosteric modulator dissociation constant and α denotes the affinity cooperativity factor. Values of α > 1 denote positive cooperativity, values α < 1 (but greater than 0) denote negative cooperativity, and values of 1 denote neutral cooperativity.
where Y denotes the percentage of specific binding, [A] is the radioligand concentration, KA is the equilibrium dissociation constant of the radioligand, KB denotes the allosteric modulator dissociation constant and α denotes the affinity cooperativity factor. Values of α > 1 denote positive cooperativity, values α < 1 (but greater than 0) denote negative cooperativity, and values of 1 denote neutral cooperativity.
Datasets for the positive modulator concentration-response curves in [3H]IPX accumulation assays and the quisqualate and modulator titration curves were analyzed globally according to a modified form of the operational model of allosterism, where it is assumed that the orthosteric agonist is full (Leach et al., 2007):
 where Basal is the response in the absence of ligand, EC50 is the midpoint of the full agonist concentration-response curve, KB is the equilibrium dissociation constant of the allosteric ligand, τB denotes the capacity of the allosteric ligand to exhibit agonism (constrained to 0 because none of the allosteric modulators exhibit agonist activity), and αβ is the net affinity/efficacy cooperativity parameter describing the combined effect of the allosteric modulator on quisqualate function (both affinity and efficacy). The terms EM and n denote the maximal possible system response and the slope factor of the transducer function that links occupancy to response, respectively.
where Basal is the response in the absence of ligand, EC50 is the midpoint of the full agonist concentration-response curve, KB is the equilibrium dissociation constant of the allosteric ligand, τB denotes the capacity of the allosteric ligand to exhibit agonism (constrained to 0 because none of the allosteric modulators exhibit agonist activity), and αβ is the net affinity/efficacy cooperativity parameter describing the combined effect of the allosteric modulator on quisqualate function (both affinity and efficacy). The terms EM and n denote the maximal possible system response and the slope factor of the transducer function that links occupancy to response, respectively.
To estimate the degree of affinity cooperativity between quisqualate and the PAMs, inhibition binding curves for PAMs versus [3H]MPEP were determined in the presence and absence of a saturating concentration of quisqualate (30 μM). Because total [3H]MPEP binding was unaffected by quisqualate (in the absence of PAMs), the cooperativity between the two ligands must be neutral (α = 1). Therefore, to estimate the degree of affinity cooperativity between quisqualate and the PAMs, a ratio of IC50 values for each PAM (except for CPPHA) in the presence and absence of quisqualate was determined.
NAM inhibition curves for quisqualate and DHPG-stimulated [3H]IPX accumulation were analyzed according to a four-parameter logistic equation as described above. For datasets studying the effect of multiple concentrations of NAMs on quisqualate concentration-response curves, data were analyzed according to the full operational model of allosterism (Leach et al., 2007):

This model estimates separately the affinities and efficacies of both orthosteric (KA, τA) and allosteric (KB, τB) ligands as well as the affinity (α) and efficacy (β) cooperativity between the ligand pair. The terms Basal, EM, and n are as described for eq. 4. Estimates of NAM affinities (KB) were constrained to those generated by [3H]MPEP binding in astrocytes. Because quisqualate had no effect on the equilibrium binding of MPEP or M-5MPEP, the affinity cooperativity (α) was constrained to unity. Finally, because the NAMs exhibit no positive agonist activity, τB was constrained to 0.
Results
[3H]MPEP Binding in Astrocyte and Cerebrocortical Membranes.
Saturation analysis of [3H]MPEP binding to membranes prepared from cortical astrocytes treated for 3 to 4 days with G5 supplement yielded a Bmax of 667 ± 149 fmol/mg of protein and a KD value of 2.9 ± 0.5 nM (Fig. 1A). For membranes prepared from adult rat cortex, [3H]MPEP saturation binding analysis yielded a Bmax of 621 ± 27 fmol/mg of protein and a KD value of 2.1 ± 0.1 nM (Fig. 1B).
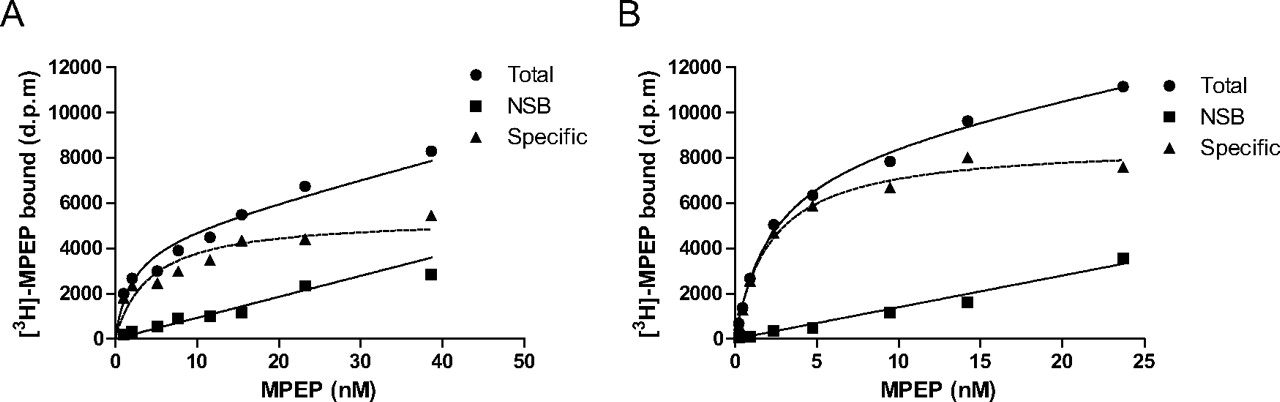
Saturation binding of [3H]MPEP to membranes prepared from rat cortical astrocytes (A) or adult rat cerebral cortex (B). To characterize the MPEP binding site, membranes were incubated with different concentrations of [3H]MPEP (0.1–40 nM) in the absence (Total) or presence (NSB) of 1 μM MPEP (see Materials and Methods). Single representative experiments are shown, with similar data being obtained on at least two additional occasions.
Modulation of [3H]MPEP Binding by Positive Allosteric Modulators (PAMs).
In the presence of a fixed concentration of [3H]MPEP (2 nM), each of the PAMs caused concentration-dependent decreases in [3H]MPEP binding to astrocyte (Fig. 2, left) and cerebrocortical membranes (Fig. 2, right). In astrocyte membranes, ADX47273 caused a similar maximal decrease in [3H]MPEP binding to that seen in the presence of a maximally effective concentration of MPEP (Fig. 2D, left). CDPPB and DFB caused apparently lesser maximal decreases in specific [3H]MPEP binding (78 and 68% decreases, respectively), probably because of a combination of low affinity and poor solubility (O'Brien et al., 2003, 2004; Kinney et al., 2005; Mühlemann et al., 2006), given that previous studies strongly suggest that these PAMs interact with the same binding site as MPEP. All of the modulators display approximate micromolar affinity for the native mGlu5 receptor (see Table 2; MPEP pKi values of 8.03 ± 0.06 and 8.12 ± 0.12 in astrocytes and cortex, respectively).
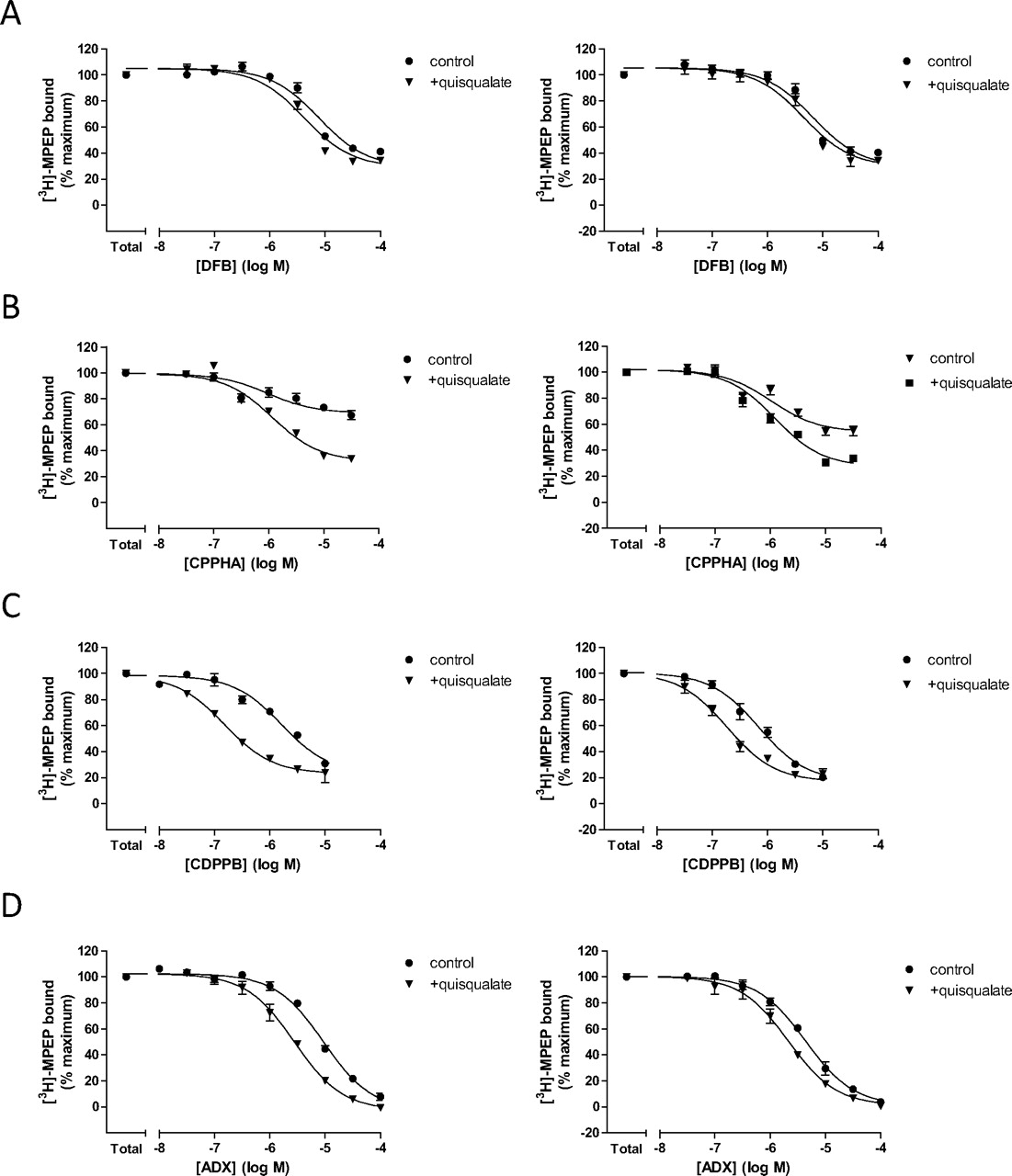
Effects of orthosteric binding site occupancy on the specific binding of [3H]MPEP in the presence of DFB, CPPHA, CDPPB, or ADX47273 in membranes prepared from rat cortical astrocytes (left) or rat cortex (right). The abilities of DFB (A), CPPHA (B), CDPPB (C), or ADX47273 (D) to affect specific [3H]MPEP binding was assessed in the absence and presence of quisqualate (30 μM). Data are presented as means ± S.E.M. of three to seven separate experiments performed in duplicate. Quantitative and statistical analyses of these data are presented in Table 1.
To assess the effects of agonist occupation of the orthosteric mGlu5 receptor binding site on binding at the [3H]MPEP allosteric site, competition binding studies were repeated in the absence and presence of quisqualate (30 μM) at an orthosteric site-saturating concentration (Mutel et al., 2000). The presence of quisqualate had no effect on total [3H]MPEP binding in membranes prepared from cortical astrocytes or adult rat cortex (1488 ± 27 versus 1507 ± 30 dpm and 1168 ± 112 versus 1211 ± 113 dpm in the absence or presence of quisqualate, respectively). However, quisqualate increased the apparent binding affinity of DFB, CDPPB, and ADX7273 in both astrocyte and rat cortex membranes (Table 1), causing parallel leftward shifts of the inhibition binding curves to each of these PAMs (Fig. 2, A, C, and D). The parallel leftward shifts of the [3H]MPEP-DFB, -CDPPB, or -ADX47273 competition curves demonstrate that quisqualate increases the affinity of the PAMs for the mGlu5 receptor. Because the concentration of quisqualate used fully occupies mGlu5 receptors (see above), the fold shift in affinity in the presence and absence of quisqualate represents the degree of affinity cooperativity (termed α) between quisqualate and the respective PAMs. Quisqualate caused a small increase in DFB affinity, which only achieved statistical significance in astrocyte membranes; in contrast, the affinities of both CDPPB and ADX47273 were significantly enhanced in the presence of the agonist, suggesting a high degree of orthosteric-allosteric positive cooperativity (Table 1). It is noteworthy that these cooperativity effects, although evident in rat cortex membranes, were more pronounced in astrocyte preparations. We have also shown that if the orthosteric site is occupied by DHPG (30 μM) or glutamate (300 μM) similar effects are seen on [3H]MPEP versus DFB, CPPHA, CDPPB, or ADX47273 interactions to those illustrated here for quisqualate (data not shown) indicating that these modulatory effects are not demonstrably orthosteric probe-dependent.
Equilibrium dissociation constants (expressed as pKi values) for mGlu5 receptor PAMs in the absence and presence of quisqualate (30 μM) in membranes prepared from cortical astrocytes or rat cortex
Data are shown as means ± S.E.M. for five to seven experiments in the absence of quisqualate and three experiments in the presence of quisqualate all performed in duplicate. Also shown is the shift in pKi value in the presence and absence of quisqualate and its antilogarithm, which represents the affinity cooperativity (α) between the PAM and agonist.
The profile of CPPHA inhibition of [3H]MPEP binding was markedly different; this PAM caused only an approximately 30% decrease in specific binding in astrocyte membranes (Fig. 2B), whereas in adult cortex membranes, an approximately 50% decrease in specific [3H]MPEP binding was observed. Furthermore, the effect of quisqualate on CPPHA was different to that observed for the other PAMs, causing an increase in the inhibition of specific [3H]MPEP binding (Fig. 2B) without affecting apparent pKi values (Table 1). These data are consistent with previous observations that CPPHA acts via an allosteric site on the mGlu5 receptor distinct from the MPEP binding site (Chen et al., 2008). Global analysis of the CPPHA inhibition binding isotherms according to a simple ternary complex model (Ehlert, 1988) suggests that CPPHA displays moderate affinity, with pKB values of 6.16 and 6.15 in astrocytes and cortex, respectively. CPPHA displays weak negative cooperativity with respect to [3H]MPEP binding in astrocytes (α = 0.57) and cortex (α = 0.37), with the degree of negative cooperativity being increased in the presence of quisqualate (α = 0.22 and 0.16 in astrocytes and cortex, respectively).
Effects of Positive Allosteric Modulators on [3H]IPx Responses in Astrocytes.
To compare the effects of mGlu5 receptor PAMs on orthosteric agonist-stimulated phospholipase C responses in astrocyte populations, we assessed [3H]IPx accumulation in the presence of Li+ as an index of phospholipase C activity. None of the mGlu5 receptor PAMs studied here (DFB, CPPHA, CDPPB, and ADX47273) increased [3H]IPx accumulation over basal levels when added alone to astrocytes (data not shown). Therefore, the ability of PAMs to increase [3H]IPx accumulation was assessed in the presence of 50 nM quisqualate, a concentration that alone causes a [3H]IPx response that is ∼20% (EC20) of the maximal quisqualate response (Fig. 3). CDPPB and ADX47273 were each able to potentiate [3H]IPx accumulation stimulated by 50 nM quisqualate to a level comparable with that seen in the presence of a maximally effective concentration of quisqualate alone. In contrast, in the presence of DFB or CPPHA less potentiation was observed (approximately 10 and 65% of maximum quisqualate response, respectively). Therefore, the efficacy rank order for these PAMs was ADX47273 ≈ CDPPB > CPPHA > DFB, whereas the potency rank order was CDPPB > CPPHA ≥ ADX47273 ≥ DFB (see Table 2).

Concentration-dependent effects of DFB (A), CPPHA (B), CDPPB (C), and ADX47273 (D) on quisqualate-stimulated [3H]IPx accumulation in rat cortical astrocytes. Astrocytes were preincubated (10 min) with increasing concentrations of mGlu5 receptor PAMs followed by stimulation with an approximate EC20 quisqualate concentration (50 nM). Concentration-dependent increases in [3H]IPx accumulation caused by mGlu5 receptor PAMs are shown in each panel compared with concentration-dependent [3H]IPx accumulations stimulated by quisqualate alone. Data are shown as means ± S.E.M. for four to five separate experiments, each performed in duplicate.
Potency and maximal response indices for potentiation by mGlu5 receptor PAMs of [3H]IPx accumulations stimulated by an EC20 concentration (50 nM) of the orthosteric agonist quisqualate in astrocytes
pEC50 values are given as −log (molar) values, and maximum potentiation values are given as a fold increase in [3H]IPx accumulation compared with the response stimulated in the presence of 50 nM quisqualate alone. Data are shown as means ± S.E.M. for four to five separate experiments each performed in duplicate. Also shown are equilibrium dissociation constants (pKB) and net cooperativity (αβ) with respect to quisqualate; the efficacy cooperativity (β) is estimated by dividing the overall cooperativity (αβ) by the affinity cooperativity (α; Table 1).
To further understand PAM effects, the modulator concentration-response curves were analyzed according to a modified form of the operational model of allosterism (Leach et al., 2007; eq. 4 under Materials and Methods) to yield an estimate of modulator affinity (pKB) and overall cooperativity with respect to quisqualate (αβ; comprising any effects on affinity and/or efficacy). Parameter estimates are shown in Table 2. The affinity rank order was identical with that seen for the EC50 values of the PAMs, whereas the estimate of cooperativity (αβ) correlated to the potentiation (fold increase) of the EC20 quisqualate response (Table 2).
Quantification of the effects of the PAMs on the quisqualate [3H]IPx accumulation response yields estimates of the overall degree of cooperativity between the respective PAMs and quisqualate (a product of the effect on affinity and efficacy, αβ). Using the value for affinity cooperativity (α) derived from the [3H]MPEP binding studies, it is possible to estimate the efficacy cooperativity between each of the PAMs (except CPPHA) and quisqualate (Table 2). These data suggest that DFB exerts its modest positive modulatory effect through marginal effects on affinity and efficacy (α = 1.5, β = 1.4). It is noteworthy that despite qualitatively similar effects on quisqualate-stimulated [3H]IPx accumulation, quantitative analysis reveals that CDPPB and ADX47273 achieve their positive modulation in different ways. The modulatory effect of CDPPB is largely achieved by an increase in quisqualate affinity, with only a modest effect on efficacy (α = 11, β = 2.3), whereas ADX47273 exerts its effects mainly through increases in quisqualate efficacy, with a more modest effect on affinity (α = 3.2, β = 9.4).
NAM Affinity Estimates at mGlu5 Receptors.
mGlu5 receptor binding affinities for MPEP and M-5MPEP were determined by competition analysis of [3H]MPEP binding to astrocyte (Fig. 4A; Table 3) and rat cortex membranes (Fig. 4B; Table 3). Both NAMs defined similar nonspecific binding components for [3H]MPEP binding to astrocyte and rat cortex membranes: pKi values obtained in the two membrane preparations revealed that M-5MPEP and the neutral allosteric site compound 5MPEP (data not shown) are equieffective in displacing [3H]MPEP binding and both compounds are 30- to 100-fold less potent compared with MPEP (Table 3). To assess the effects of orthosteric site occupation, adult rat cortex membranes were incubated with a high (30 μM) concentration of quisqualate and [3H]MPEP versus MPEP or M-5MPEP competition analyses performed (Fig. 4, C and D; Table 3). Agonist occupation of the orthosteric site had no effect on the interaction between negative/neutral allosteric modulators and [3H]MPEP. These data suggest that unlike the PAMs, the NAMs exhibit neutral affinity cooperativity with respect to quisqualate (α = 1). The above findings were wholly recapitulated if studies were repeated under the assay conditions of Liu et al. (2008) used for the [3H]MPEP-PAM radioligand studies (data not shown).

Displacement of specific [3H]MPEP binding by mGlu5 receptor NAMs, MPEP, and M-5MPEP in membranes prepared from rat cortical astrocytes (A) or rat cortex (B). The indicated concentrations of NAMs were added to membranes immediately before the addition of [3H]MPEP (10 nM final concentration; see Materials and Methods). Effects of orthosteric binding site occupancy on the displacement of specific [3H]MPEP binding by MPEP (C) and M-5MPEP (D) was also investigated in rat cortex membranes. [3H]MPEP binding was performed in the absence and presence of quisqualate (30 μM). Data are shown as means ± S.E.M. of three to six separate experiments performed in duplicate. Quantitative and statistical analyses of these data are presented in Table 3.
Comparison of binding affinity constants for mGlu5 receptor NAMs in the absence and presence of quisqualate (30 μM) in astrocyte and rat cortex membranes
There were no statistical differences between control and quisqualate-treated rat cortex membranes as determined by unpaired Student's t test.
Effects of NAMs on Astrocyte Single-Cell Ca2+ Responses.
The NAM MPEP has been shown previously to decrease glutamate-stimulated Ca2+ oscillations in a concentration-dependent manner in CHO-lac-mGlu5a cells and astrocytes (Nash et al., 2002; Bradley et al., 2009). Here, we have extended these previous studies to include a comparison between MPEP and the “partial” NAM, M-5MPEP (Rodriguez et al., 2005), and to investigate whether any probe-dependence is observed by assessing the effects of these NAMs on Ca2+ oscillations initiated by quisqualate, DHPG, or glutamate. Although the frequency of Ca2+ oscillations stimulated by each of these orthosteric agonists varied slightly, the concentration-dependence for MPEP-induced suppressions of agonist-stimulated responses did not seem to be probe-dependent, giving a pIC50 ≈ 8 and full inhibition versus each of the orthosteric agonists tested (Fig. 5; Table 4). M-5MPEP also caused concentration-dependent, probe-independent, full inhibition of orthosteric agonist-stimulated Ca2+ oscillations in astrocytes (Fig. 6; Table 4), and observations are in agreement with results obtained previously in CHO-lac-mGlu5a cells (Bradley et al., 2009). Given that the affinity cooperativity between the NAMs and quisqualate is neutral, the inhibitory effects of the NAMs must be mediated by a change in agonist efficacy. Although it was not possible to quantify the degree of efficacy cooperativity between NAMs and quisqualate (because the data are only for a single curve), the full inhibition of agonist-stimulated Ca2+ oscillations suggests that the value of β must tend toward 0.
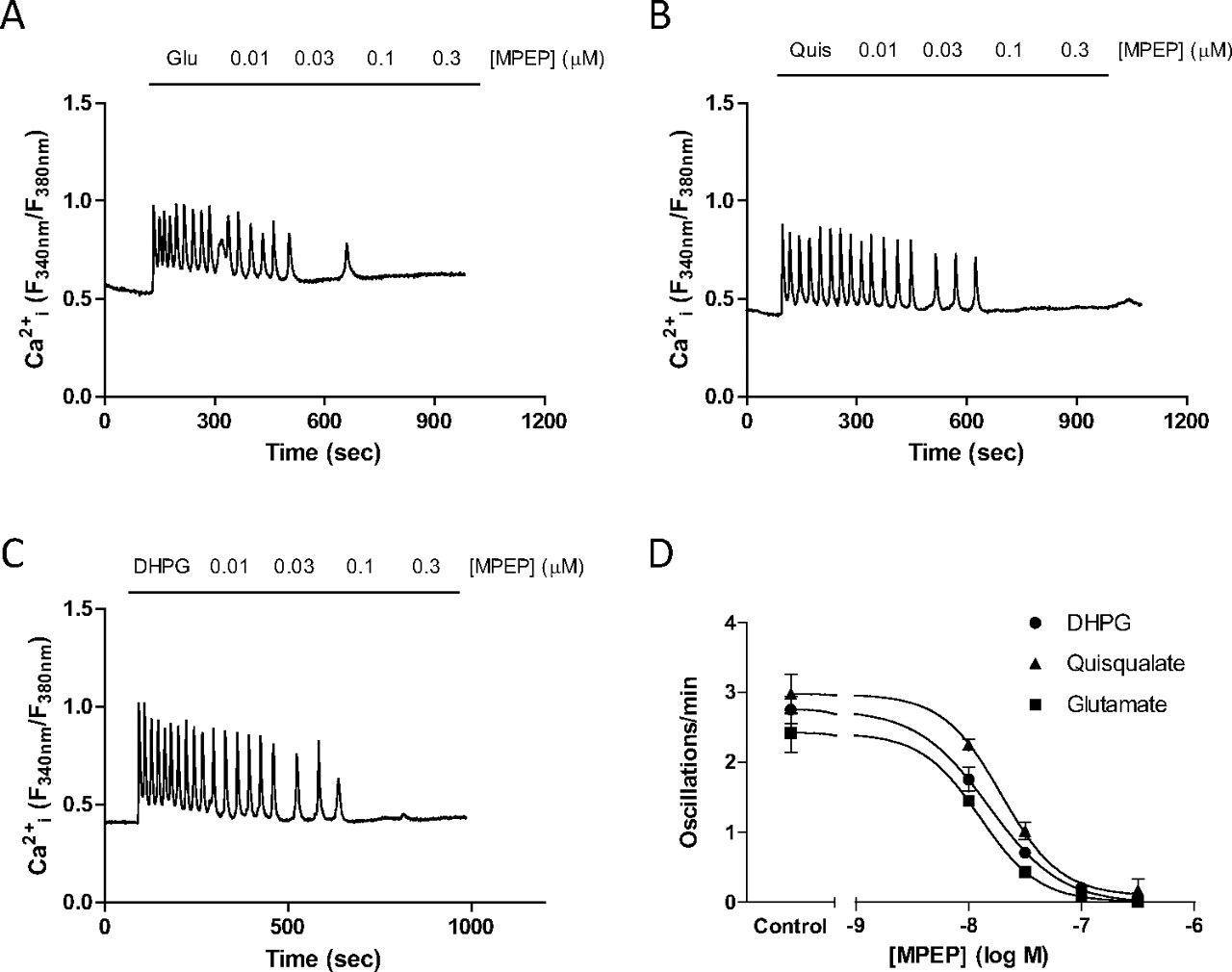
Concentration-dependent effects of MPEP on Ca2+ oscillations stimulated by glutamate, quisqualate, or DHPG in rat cerebrocortical astrocytes. Representative traces showing the effects of incrementally increasing concentrations of MPEP (0.01–0.3 μM) on Ca2+ oscillations elicited by the continuous presence of glutamate (100 μM; A), quisqualate (10 μM; B), or DHPG (10 μM; C). Summary data are also shown for the concentration-dependent suppression of Ca2+ oscillations by MPEP when cells were stimulated by glutamate, quisqualate, or DHPG (D). Mean pIC50 (M) values for inhibition of glutamate-, quisqualate- or DHPG-stimulated Ca2+ oscillation frequency by MPEP were 7.90 ± 0.06, 7.71 ± 0.07 and 7.82 ± 0.06, respectively. Data are shown as means ± S.E.M. from at least 50 individual cells recorded over at least three separate experiments.
Comparison of potency (pIC50) and efficacy (Rmax) indices for mGlu5 receptor-mediated [3H]IPx and Ca2+ responses
pEC50 values are given as −log molar values, and Rmax values are given as a percentage of the maximal response to quisqualate.
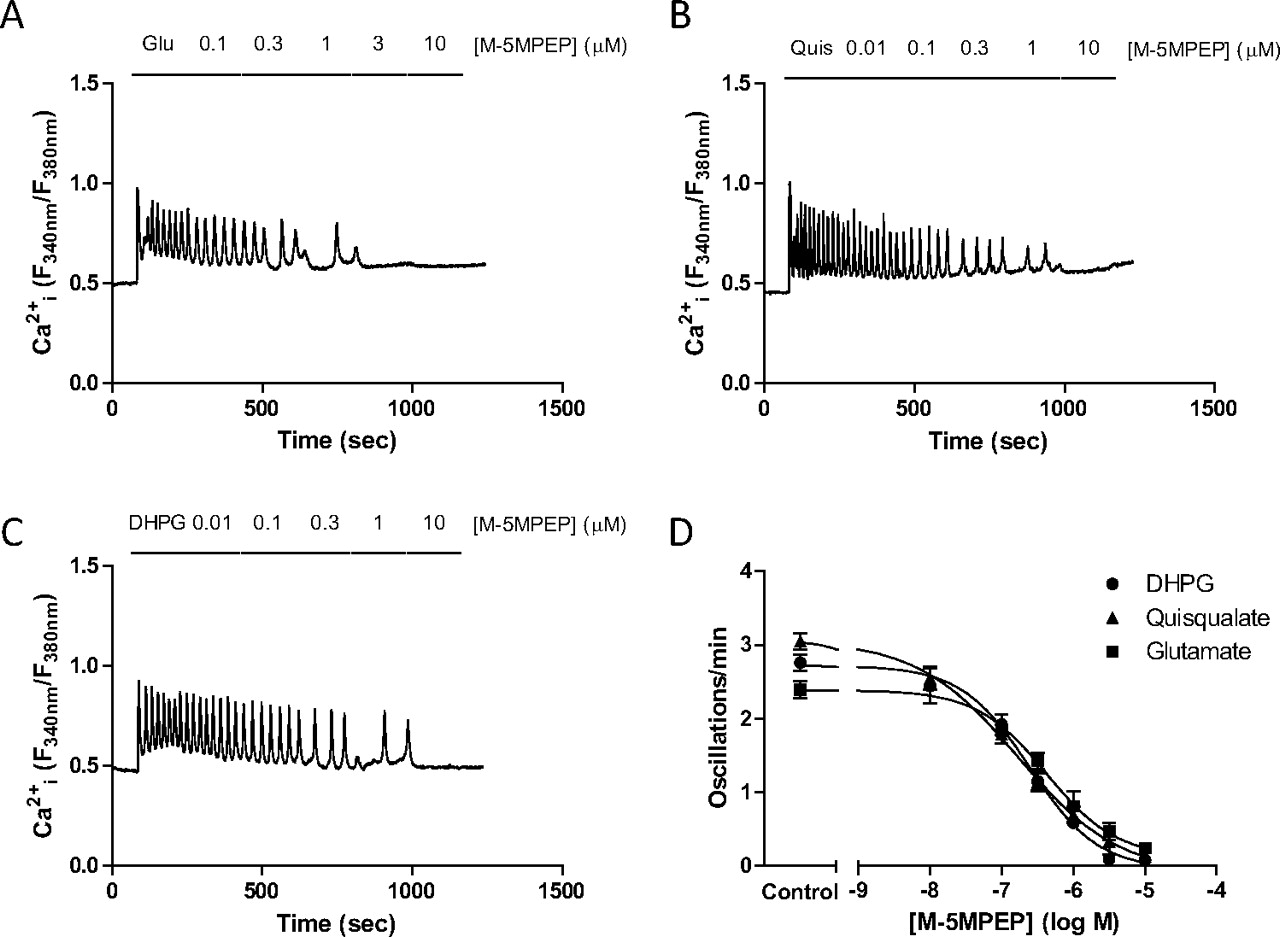
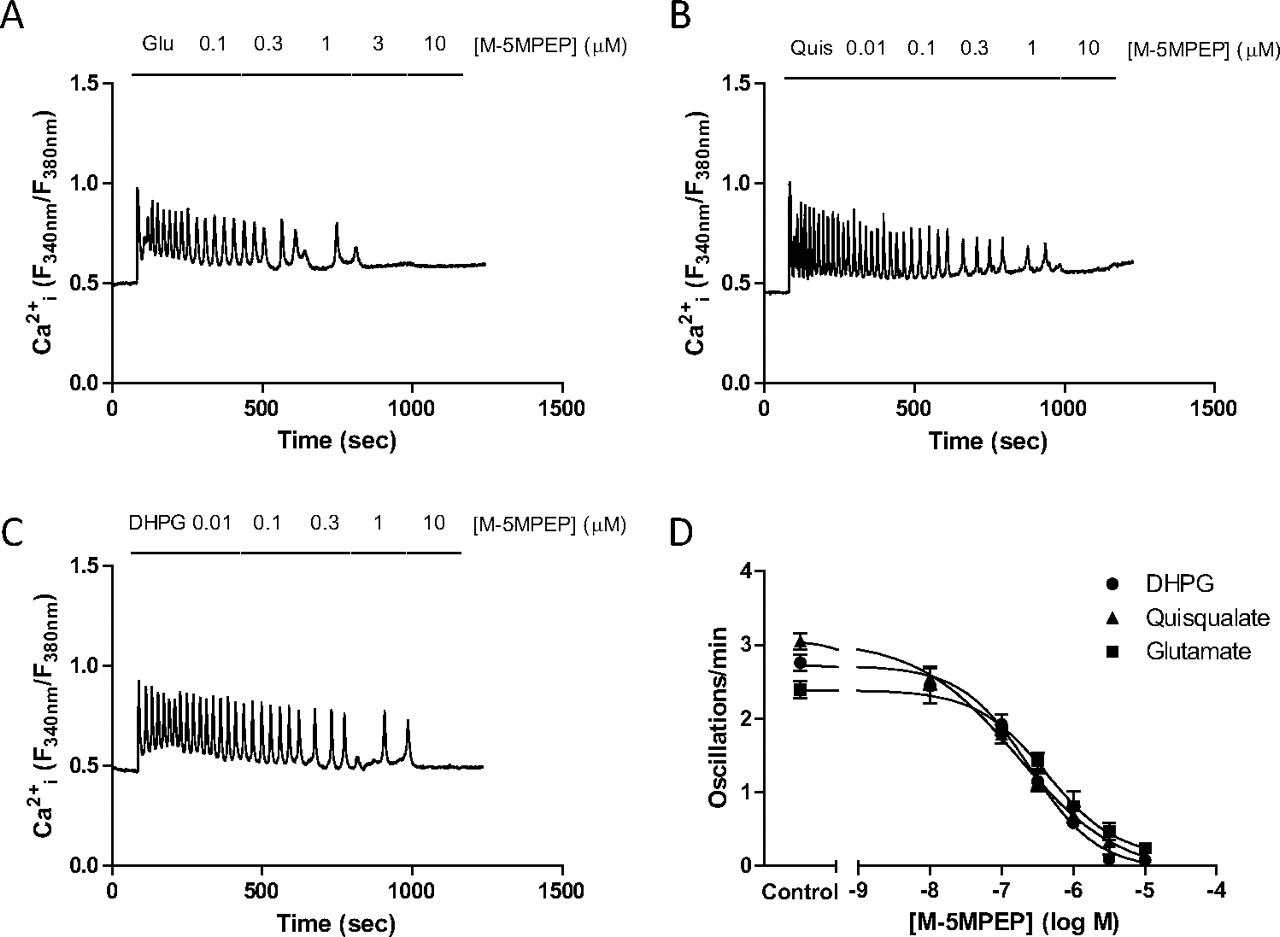
Concentration-dependent effects of M-5MPEP on Ca2+ oscillation frequency stimulated by glutamate, quisqualate, or DHPG in rat cerebrocortical astrocytes. Representative traces showing the effects of incrementally increasing concentrations of M-5MPEP (0.01–10 μM) on Ca2+ oscillations elicited by the continuous presence of glutamate (100 μM; A), quisqualate (10 μM; B), or DHPG (10 μM; C). Summary data are also shown for the concentration-dependent suppression of Ca2+ oscillations by M-5MPEP when cells were stimulated by glutamate, quisqualate, or DHPG (D). Mean pIC50 (M) values for inhibition of glutamate-, quisqualate-, or DHPG-stimulated Ca2+ oscillation frequency by M-5MPEP were 6.61 ± 0.08, 6.77 ± 0.13, and 6.38 ± 0.15, respectively. Data are shown as means ± S.E.M. from at least 50 individual cells recorded over at least 3 separate experiments.
Effects of NAMs on Phosphoinositide Hydrolysis in Astrocytes.
In the presence of Li+, the addition of quisqualate for 20 min stimulated an approximately 6-fold increase in [3H]IPx accumulation [basal, 74,899 ± 1389; +quisqualate (30 μM), 447,086 ± 16,090 dpm/mg of protein; pEC50 (M) = 6.80 ± 0.02]. The mGlu5 receptor NAMs MPEP and M-5MPEP decreased the [3H]IPx accumulation stimulated by quisqualate (10 μM) in a concentration-dependent manner (Fig. 7; Table 4), whereas 5-MPEP was without effect (data not shown). The pIC50 values for MPEP-mediated inhibition of mGlu5 receptor-mediated [3H]IPx accumulation compared well with pIC50 values derived for the inhibition of Ca2+ oscillations (Table 4), whereas lower IC50 values were observed for inhibitory effects on Ca2+ oscillations relative to [3H]IPx accumulation for M-5MPEP (Table 4). More strikingly, whereas MPEP was able to inhibit fully agonist-stimulated [3H]IPx accumulation, M-5MPEP only partially inhibited this response (Fig. 7, A and B), contrasting with the ability of this NAM to inhibit completely the Ca2+ response. This “partial NAM” effect of M-5MPEP was also observed with respect to the [3H]IPx accumulation stimulated by DHPG (Fig. 7B). Using an alternative experimental design, it could be shown that increasing concentrations of MPEP progressively suppress quisqualate-stimulated [3H]IPx accumulation in astrocytes with only an approximately 3-fold change in apparent quisqualate potency (Fig. 7C; pEC50 (M): −MPEP, 6.72 ± 0.09; +1 μM MPEP, 6.24 ± 0.11). Qualitatively the effect of MPEP is highly likely to be mediated by changes in quisqualate efficacy, as the ligands exhibit neutral affinity cooperativity with respect to one another (see above). Analysis of the dataset according to the operational model of allosterism (Leach et al., 2007; eq. 5 under Materials and Methods) indicates a high degree of negative efficacy cooperativity between quisqualate and MPEP (β = 0.01), reflecting the full inhibition of quisqualate-stimulated [3H]IPx accumulation. These data are quantitatively similar to those seen for MPEP in the Ca2+ oscillation assay. In contrast, increasing concentrations of M-5MPEP (up to 30 μM) resulted in only a partial, concentration-dependent suppression of quisqualate-stimulated [3H]IPx accumulation (Fig. 7D). Quantitative analysis of the interaction between M-5MPEP and quisqualate suggests a weak degree of negative efficacy cooperativity (β = 0.37) between the two ligands in the [3H]IPx accumulation assay, in marked contrast to the full inhibitory effect (β approaches 0) observed in the Ca2+ oscillation assay.

Effects of MPEP and M-5MPEP on agonist-stimulated [3H]IPx accumulation in rat cortical astrocytes. Increasing concentrations of mGlu5 receptor NAMs were preincubated for 10 min before orthosteric agonist addition. Data are expressed as the percentage of maximal [3H]IPx accumulation on stimulation with quisqualate (10 μM; A) or DHPG (10 μM; B). Data shown are means ± S.E.M. for four to seven separate experiments performed in duplicate. Also shown are quisqualate-stimulated concentration-response curves performed in the absence or presence of 0.03, 0.1, 0.3, or 1 μM MPEP (C) or 1, 3, 10, or 30 μM M-5MPEP (D). Either MPEP or M-5MPEP was added 10 min before the addition of quisqualate at the concentrations indicated. Data are shown as means ± S.E.M. for three separate experiments performed in duplicate.
Discussion
The discovery of compounds that alter mGlu receptor activity through binding to allosteric sites within the transmembrane domain has been rapidly followed-up by the generation of an array of compounds that show good mGlu receptor subtype selectivity, as well as additional pharmacologically desirable pharmacokinetic/pharmacodynamic properties (Conn et al., 2009; Niswender and Conn, 2010). Positive or negative allosteric modulation of the mGlu5 receptor has been indicated as a fruitful therapeutic approach in a number of neuropsychiatric disorders (Swanson et al., 2005; Marino and Conn, 2006; Dölen and Bear, 2008). Whereas mGlu5 receptor PAMs and NAMs can exert direct agonist or inverse agonist activity, for example, at a mutant mGlu5 receptor in which the N-terminal orthosteric ligand binding domain has been removed (Goudet et al., 2004), the majority of the NAMs and PAMs reported to date (and those investigated here) are considered to be true modulators in that they alter the ability of orthosteric agonists to effect mGlu5 receptor activity. This allosteric modulatory action can be brought about by altering the affinity and/or the efficacy of the orthosteric agonist at the receptor (Langmead and Christopoulos, 2006). Surprisingly, little information is currently available defining the mode of mGlu5 receptor allosteric modulation, and a key objective of the present study has been to provide a more quantitative analysis of the actions of prototypic NAMs and PAMs at native mGlu5 receptors.
To achieve this, we first examined the interaction of NAMs and PAMs at the MPEP site on the mGlu5 receptor using [3H]MPEP binding. The density of [3H]MPEP binding sites in G5 supplement-differentiated rat cortical astrocytes approximated that found in adult rat cerebral cortex; however, it should be noted that the major splice variant expressed in the former is the mGlu5a (Biber et al., 1999), whereas mGlu5b is likely to predominate in the latter (Minakami et al., 1995). The NAMs MPEP and M-5MPEP, and the neutral allosteric modulator 5-MPEP, all competed for the [3H]MPEP binding site in membrane preparations from rat cortical astrocytes and cerebral cortex and defined identical levels of specific binding. Competition isotherms were completely unaffected by the absence or presence of quisqualate at a concentration expected to occupy fully the orthosteric binding site. In contrast, PAMs inhibited [3H]MPEP binding to different extents. Previous work has demonstrated that DFB (O'Brien et al., 2003), CDPPB (Kinney et al., 2005; Chen et al., 2007), and ADX47273 (Liu et al., 2008) bind to a similar (or substantially overlapping) allosteric binding site to MPEP, whereas CPPHA binds to a distinct locus within the mGlu5 receptor 7-transmembrane domain (O'Brien et al., 2004; Chen et al., 2008). Indeed, a number of studies have provided information on the amino acid residues that contribute to common NAM/PAM-mGlu5 receptor interactions (Gasparini et al., 1999; Mühlemann et al., 2006). Whereas apparently incomplete inhibition of specific [3H]MPEP binding has been observed previously for DFB (versus the MPEP analog [3H]methoxyPEPy; O'Brien et al., 2003), another study has reported that high concentrations of CDPPB can inhibit [3H]methoxyPEPy binding to levels similar to those in MPEP (Kinney et al., 2005). Therefore, although it might be tempting to speculate on possible allosteric interactions of these PAMs with respect to [3H]MPEP binding it is more likely that technical difficulties (e.g., low affinity, compound solubility problems, etc.) underlie the [3H]MPEP competition binding isotherms observed for DFB and CDPPB. Notwithstanding these concerns, it is clear that agonist occupancy of the mGlu5 orthosteric binding site alters the apparent affinity of DFB, CDPPB, and ADX47273 at the [3H]MPEP binding site, with respective 1.5-, 3.2-, and 11-fold leftward shifts (α values) being seen in astrocyte membranes. In contrast, agonist occupancy of the orthosteric site did not affect the apparent affinity of CPPHA for the [3H]MPEP binding site, but analysis using the allosteric ternary complex model revealed the degree to which negative cooperativity was increased. These latter data confirm and extend previous studies on the pharmacological properties of CPPHA (O'Brien et al., 2004; Chen et al., 2008).
To investigate efficacy-related effects of PAMs, we adopted a protocol used previously (O'Brien et al., 2003, 2004; Kinney et al., 2005; Chen et al., 2008; Liu et al., 2008); however, [3H]IPx accumulation rather than a population Ca2+ response was used here as a readout. Quisqualate stimulated a ∼6-fold increase in [3H]IPx accumulation in astrocytes in the presence of Li+. In the presence of an approximate EC20 concentration (50 nM) of the orthosteric agonist, each of the PAMs caused an enhancement of the quisqualate-stimulated [3H]IPx accumulation, but to different extents. Qualitatively the maximal response ranking order was ADX47273 ≈ CDPPB > CPPHA > DFB with maximal potentiation of the 50 nM quisqualate response varying from 4.7-fold down to 1.5-fold. Further analysis using a modified form of the operational model of allosterism (Leach et al., 2007; see Materials and Methods) yielded net affinity/efficacy cooperativity parameters (αβ; Table 2), which varied from 30 (ADX47273) to 2.1 (DFB). By using the previously determined affinity cooperativity (α) parameters, it was possible to resolve affinity and efficacy components for each PAM. Strikingly, we have demonstrated that ADX47273 and CDPPB differ substantially in the ways that they exert their PAM actions. Whereas CDPPB is primarily an affinity modulator (i.e., exerts it PAM activity through increasing the apparent mGlu5 receptor binding affinity for orthosteric agonists), ADX47273 exerts a much greater part of its PAM activity through efficacy modulation. Thus, through the application of quantitative pharmacological analyses, it is possible to reveal marked mechanistic differences between allosteric modulators that are not apparent from more conventional analysis. That two PAMs bind to a common site to exert mechanistically different allosteric effects is not inconsistent with our current understanding of allostery at GPCRs (Kenakin and Miller, 2010). Indeed, it would be interesting to perform mutagenesis studies to establish whether, within the mGlu5 receptor MPEP binding pocket, the amino acid residues critical for the actions of each PAM diverge and thus provide an empirical basis for the differing receptor ensemble conformational selectivity of the two PAMs that might be hypothesized. These new findings also raise the issue of whether the mechanism of positive allosteric modulation is of potential importance from a therapeutic perspective (i.e., with respect to the therapeutic deployment of mGlu5 receptor PAMs in neuropsychiatric disorders): is an affinity- or efficacy-driven modulation more desirable?
Competition analyses for MPEP and M-5MPEP versus [3H]MPEP binding in astrocyte and adult cerebral cortex membranes indicated similar definitions of specific (displaceable) binding and a complete lack of effect of agonist occupation of the orthosteric site on respective competition isotherms, strongly indicating that these agents exert their negative modulatory activity through efficacy rather than affinity effects. A mechanism for efficacy-only modulation is unclear and these effects could be mediated via changes in receptor-G protein interaction. The effects of the NAMs are unlikely to be due to steric hindrance between G protein and receptor, because previous site-directed mutagenesis studies have clearly indicated a transmembrane domain binding locus. Therefore, it could be that there is negative affinity cooperativity between the G protein and NAM but not between orthosteric agonist(s) and the NAM. This manifests as apparent “negative efficacy cooperativity” when considering only the ternary complex of orthosteric agonist, receptor, and NAM. However, it is possible to argue that if a quaternary complex, incorporating G protein, is considered, then the differential affinity cooperativity of PAMs and NAMs (with either orthosteric agonist or G protein) might become apparent; however, the necessary tools are not presently available to test this possibility.
To further compare MPEP and M-5MPEP, we assessed their respective abilities to inhibit orthosteric agonist-stimulated [3H]IPx accumulation and single-cell Ca2+ oscillations in astrocytes. Although MPEP fully inhibited both functional responses with pIC50 values consistent with previous studies (Gasparini et al., 1999; Nash et al., 2002; O'Brien et al., 2003; Rodriguez et al., 2005; Chen et al., 2008), M-5MPEP only partially inhibited the [3H]IPx response (irrespective of whether it was stimulated by quisqualate or DHPG) but fully inhibited Ca2+ oscillations stimulated by l-glutamate, quisqualate, or DHPG. In the original report on M-5MPEP, this compound was described as a “partial antagonist” based on its ability only partially to inhibit glutamate-stimulated changes in [Ca2+]i in populations of astrocytes (Rodriguez et al., 2005). Here, we have reproduced this original finding with respect to the [3H]IPx response, but, as reported previously, in CHO-lac-mGlu5a cells (Bradley et al., 2009), M-5MPEP also seems to possess a greater inverse efficacy with respect to Ca2+ oscillatory responses in astrocytes. It is also noteworthy that the potency of M-5MPEP for the inhibition of Ca2+ oscillations was significantly greater than for the (partial) inhibition of [3H]IPx accumulation. For example, M-5MPEP displayed a 17-fold greater potency for inhibiting quisqualate-stimulated Ca2+ oscillations versus [3H]IPx responses. As discussed previously, mGlu5 receptor-mediated Ca2+ oscillations are driven by phosphorylation/dephosphorylation of the receptor itself (Nash et al., 2002; Bradley et al., 2009) driven by diacylglycerol (and Ca2+)-dependent protein kinase C isoenzymes. Therefore, although it is difficult to view the [3H]IPx and Ca2+ oscillatory responses as independent readouts, it is nevertheless tempting to speculate that M-5MPEP may be a NAM that can display permissive antagonism (Kenakin, 2005).
Authorship Contributions
Participated in research design: Bradley, Langmead, Watson, and Challiss.
Conducted experiments: Bradley.
Performed data analysis: Bradley, Langmead, and Challiss.
Wrote or contributed to the writing of the manuscript: Bradley, Langmead, and Challiss.
Footnotes
This work was supported by a PhD CASE studentship from the Biotechnology and Biological Sciences Research Council of Great Britain (to S.J.B.).
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
doi:10.1124/mol.110.068882.
-
ABBREVIATIONS:
- mGlu
- metabotropic glutamate
- PAM
- positive allosteric modulator
- NAM
- negative allosteric modulator
- MPEP
- 2-methyl-6-(phenylethynyl)-pyridine
- 5MPEP
- 5-methyl-2-(phenylethynyl)pyridine
- M-5MPEP
- 2-(2-(3-methoxyphenyl)ethynyl)-5-methylpyridine
- DHPG
- (S)-3,5-dihydroxyphenylglycine
- ADX47273
- S-(4-fluoro-phenyl)-{3-[3-(4-fluoro-phenyl)-[1,2,4]oxadiazol-5-yl]-piperidinl-1-yl}-methanone
- DFB
- 3, 3′-difluorobenzaldazine
- CDPPB
- 3-cyano-N-(1,3-diphenyl-1H-pyrazol-5-yl)benzamide
- CPPHA
- N-{4-chloro-2-[(1,3-dioxo-1,3-dihydro-2H-isoindol-2-yl)methyl]phenyl}-2-hydroxybenzamide
- CHO
- Chinese hamster ovary
- [3H]IPx
- [3H]inositol mono, bis-, and trisphosphate fraction.
- Received September 13, 2010.
- Accepted February 1, 2011.
- Copyright © 2011 The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}