


Abstract
In this work, we examined the ability of gp120, a human immunodeficiency virus-1 (HIV-1) viral envelope glycoprotein, to trigger the innate immune response in astrocytes, an HIV-1 brain cellular target, and we investigated the functional expression of the ATP-binding cassette membrane transporter P-glycoprotein (P-gp) in primary cultures of rat astrocytes treated with gp120 or cytokines [tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and IL-6]. Standard 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium and d-mannitol uptake assays confirmed that HIV-196ZM651 gp120 treatment did not alter cell viability or membrane permeability. Semiquantitative reverse-transcriptase polymerase chain reaction analysis and enzyme-linked immunosorbent assay demonstrated increased TNF-α, IL-1β, and IL-6 mRNA and protein expression in cultures treated with HIV-196ZM651 gp120, suggesting in vitro activation of immune responses. Cytokine secretion was detected when CXCR4 but not CCR5 was inhibited with a specific antibody, implying that cytokine secretion is primarily mediated via CCR5 in astrocytes triggered with HIV-196ZM651 gp120. P-gp protein expression was increased in astrocyte cultures exposed to TNF-α (2.9-fold) or IL-1β (1.6-fold) but was decreased profoundly in the presence of IL-6 (8.9-fold), suggesting that IL-6 is primarily involved in modulating P-gp expression. In parallel, after HIV-196ZM651 gp120 treatment, immunoblotting analysis showed a significant decrease in P-gp expression (4.7-fold). Furthermore, the accumulation of two P-gp substrates, digoxin and saquinavir (an HIV-1 protease inhibitor), was enhanced (1.5- to 1.8-fold) in HIV-196ZM651 gp120-treated astrocyte monolayers but was not altered by P-gp inhibitors [e.g., valspodar (PSC833) and elacridar (GF120918)], suggesting a loss of transport activity. Taken together, these data imply that HIV-196ZM651 gp120 or cytokine treatment modulate P-gp functional expression in astrocytes, which may lead to complex drug-transporter interactions during HIV-1 encephalitis-associated immune responses.
Immunological responses in the brain have been observed in various disease pathologies, including human immunodeficiency virus type-1 (HIV-1) encephalitis (HIVE). Increased production and secretion of proinflammatory cytokines (e.g., TNF-α, IL-1β, and IL-6) during HIVE is well-established (Persidsky and Gendelman, 2003; Kaul et al., 2005) and is believed to result from persistent cytopathic infection within perivascular and blood-derived parenchymal brain macrophages and microglia (Kaul et al., 2005). These infected cells release soluble viral proteins (e.g., HIV-1 envelope glycoprotein gp120) into the brain parenchyma (Kaul et al., 2005). Recent in vivo studies have demonstrated that gp120 treatment increases cytokine secretion (Holguin et al., 2004; Corasaniti et al., 2005). For example, Holguin et al. (2004) observed increased expression of TNF-α, IL-1β, and IL-6 in spinal cord samples isolated from rats injected intrathecally with gp120. Corasaniti et al. (2005) demonstrated elevated IL-1β expression in rat neocortex after gp120 administration, suggesting a relationship between gp120 exposure and increased brain concentrations of cytokines. Studies in cultured astrocytes suggest that gp120 binding to cell surface chemokine receptors (e.g., CXCR4 and CCR5) may mediate this inflammatory response (Wu et al., 1997; Koller et al., 2002).
Effective pharmacological treatment of central nervous system disease requires that drugs achieve therapeutic concentrations within the brain. Despite the presence of several specific uptake systems at the blood-brain barrier (BBB) and blood-cerebrospinal fluid barrier, many compounds do not attain therapeutic levels in the brain (Lee and Bendayan, 2004). One potential mechanism for these low brain concentrations is the expression of ATP-binding cassette (ABC) efflux drug transporters [e.g., P-glycoprotein (P-gp), multidrug resistance-associated proteins (MRP in humans; Mrp in rodents), and breast cancer resistance protein (also known as ABCG2)] in cellular compartments of the central nervous system (e.g., brain microvessel endothelial cells, astrocytes, and microglia). P-gp, a 170-kDa membrane protein, was originally identified in colchicine-resistant Chinese hamster ovary cells (Juliano and Ling, 1976). P-gp has been shown to be an ATP-dependent efflux transporter capable of extruding from cells several structurally unrelated drugs, including chemotherapeutic agents (e.g., doxorubicin, vincristine) (DiDiodato and Sharom, 1997) and HIV-1 protease inhibitors (e.g., saquinavir, indinavir) (Lee et al., 1998; Ronaldson et al., 2004b). P-gp is encoded by the multidrug resistance (MDR) gene, which has two isoforms in humans (MDR1 and MDR2) and three isoforms in rodents (mdr1a, mdr1b, and mdr2). Although MDR2/mdr2 is primarily involved in hepatic phosphatidylcholine transport, overexpression of human MDR1 or rodent mdr1a/mdr1b confers the MDR phenotype (Gottesman et al., 1995). The MDR phenotype is defined as the simultaneous resistance to several structurally unrelated compounds. MDR does not result from independent genetic mutations, which confers resistance to a single xenobiotic (Gottesman et al., 1995).
In the central nervous system, P-gp expression has been localized to the apical surface of the choroid plexus and at the brain microvascular endothelium (Beaulieu et al., 1997; Rao et al., 1999). Our laboratory has also characterized P-gp functional expression in a rat microglia cell line (MLS-9) (Lee et al., 2001), cultured rat astrocytes (Ronaldson et al., 2004a), and in an immortalized rat brain microvessel endothelial cell line (RBE4) (Bendayan et al., 2002).
Recent studies have suggested that the brain innate immune response may alter the functional expression of P-gp (Goralski et al., 2003; Rivest, 2003; Hartz et al., 2006). Exogenous stimuli such as bacterial and viral infection, trauma, and cell stress can trigger immunological responses that are characterized by cytokine secretion (i.e., TNF-α, IL-1β, and IL-6) (Rivest, 2003). For example, Goralski et al. (2003) observed decreased mdr1a mRNA levels in whole brain tissue isolated from Sprague-Dawley rats treated with 5 mg/ml lipopolysaccharide (LPS), a bacterial endotoxin and potent activator of the brain's innate immune system. Increased brain accumulation of digoxin, a P-gp substrate, in rats exposed to LPS has also been reported, which implies impaired P-gp activity during an immunological response (Goralski et al., 2003). Similar findings were observed by Hartz et al. (2006), who demonstrated increased accumulation of NBD-CSA, a fluorescent cyclosporine A analog and established P-gp substrate, in isolated rat brain capillaries treated with LPS. Furthermore, this same study demonstrated decreased P-gp activity in isolated capillaries exposed to TNF-α, which implies that P-gp-mediated transport may be altered in the presence of cytokines.
In the present study, we characterized cytokine secretion in an in vitro glial model of gp120-mediated inflammatory response, evaluated P-gp functional expression in cultured astrocytes treated with HIV-196ZM651 gp120, and investigated the effects of TNF-α, IL-1β, and IL-6 on P-gp protein expression.
Materials and Methods
Materials. The following reagents were obtained from the National Institutes of Health AIDS Research and Reference Reagent Program, Division of AIDS, National Institute of Allergy and Infectious Diseases, National Institutes of Health (Bethesda, MD): HIV-196ZM651 gp120 full-length protein (derived from subtype C HIV-1) and the murine monoclonal CCR5 antibody 3A9. [14C]Saquinavir (3.2 × 10-4 μCi/mmol) was a gift from Roche Products Ltd. (Hertfordshire, UK). PSC833 (valspodar) and GF120918 (elacridar) were kindly provided by Novartis Pharma (Basel, Switzerland) and GlaxoSmithKline (Research Triangle Park, NC), respectively. [3H]Digoxin (23.5 Ci/mmol) was purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). MRK16 was obtained from BIOMOL Research Laboratories (Plymouth Meeting, PA). Rat recombinant TNF-α and the murine monoclonal actin antibody AC-40 were purchased from Sigma-Aldrich (Oakville, ON, Canada). Rat recombinant IL-1β, rat recombinant IL-6, rat recombinant MIP-1α, human recombinant SDF-1α, the murine monoclonal TNF-α-neutralizing antibody, and the murine monoclonal IL-1β-neutralizing antibody were purchased from Chemicon Inc. (Temecula, CA). The rat monoclonal IL-6-neutralizing antibody was purchased from R & D Systems (Minneapolis, MN). The rabbit polyclonal anti-CXCR4 antibody was purchased from eBioscience (San Diego, CA), and the murine monoclonal P-gp antibody C219 was purchased from ID Labs (London, ON, Canada).
Cell Culture. Primary cultures of rat astrocytes were prepared as described previously (Ronaldson et al., 2004a). All procedures were carried out in accordance with the University of Toronto Animal Care Committee and the Province of Ontario Animals for Research Act. In brief, postnatal (1- to 3-day-old) Wistar rats (Charles River Laboratories, St. Constant, PQ, Canada) were killed by cervical dislocation, and the whole brains were isolated. Cerebral cortices were dissected and subjected to enzymatic digestion for 30 min in serum-free minimum essential medium containing 2.0 mg/ml porcine pancreatic trypsin (Sigma-Aldrich) and 0.005% DNase I (Roche Applied Science, Laval, PQ, Canada). The tissue was then mechanically disaggregated using a cell dissociation kit (Sigma-Aldrich) to yield a mixed glial cell suspension. The cell suspension was then centrifuged for 10 min at 100g and resuspended in primary glial culture medium, which consisted of minimum essential medium supplemented with 5% horse serum, 5% fetal bovine serum, and 50 μg/ml gentamicin. The cells were plated onto 75-cm2 polystyrene tissue culture flasks (Sarstedt, St. Leonard, PQ, Canada) and incubated in fresh medium at 37°C, in 5% CO2/95% air overnight for 7 to 10 days until confluence was attained. The cells were then placed on an orbital shaker at 120 rpm for 6 h to remove contaminating oligodendrocytes, microglia, progenitor cells, and neurons. The cells were harvested with 0.1% trypsin/EDTA in Hanks' balanced salt solution and plated at a density of 5 × 104 cells/well on 48-well polystyrene plates (BD Biosciences, San Jose, CA). The astrocytic nature of the cells in culture and their purity were assessed previously by morphological analysis and immunostaining for standard biochemical markers (e.g., glial fibrillary acidic protein) (Ronaldson et al., 2004a).
The Madin-Darby Canine Kidney MDR1 (MDCK-MDR1) cell line was kindly provided by Dr. Piet Borst (Netherlands Cancer Institute, Amsterdam, The Netherlands). The MDCK-MDR1 cell line was created by retroviral transfection using the plasmid pCMV-Neo-MDR1. The cells were grown as monolayers on 75-cm2 tissue culture flasks at 37°C in 5% CO2/95% air. The cultures were fed every 2 days with Dulbecco's modified Eagle's medium with GlutaMAX supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. Confluent cultures were subcultured with 0.25% trypsin-EDTA.
gp120/Cytokine/Chemokine Cell Treatment. All treatments were performed on monolayers of primary cultures of rat astrocytes grown in 75-cm2 tissue culture flasks. At the beginning of each experiment, culture medium was aspirated, and fresh medium containing 10% fetal bovine serum and 1.0 nM HIV-196ZM651 gp120 was added. HIV-196ZM651 gp120 is R5-tropic (also known as macrophagetropic) and is derived from a subtype C viral isolate. HIV-1 subtype C predominates globally and is estimated to have caused 47.2% of new HIV-1 infections in 2000 (Julg and Goebel, 2005). In addition, R5-tropic viruses are the most prevalent strains of HIV-1 in the brain (Gabuzda and Wang, 2000). All experiments were conducted at 37°C in 5% CO2/95% air. Control (i.e., untreated) cultures were composed of untreated cells in fresh culture medium. At 6, 12, and 24 h, culture medium was collected for enzyme-linked immunosorbent assay (ELISA) analysis, and the cells were collected and prepared for RT-PCR or immunoblot analysis as described below.
To elucidate the role of chemokine receptors in the gp120 inflammatory response, astrocyte cultures were treated with CXCR4- and/or CCR5-neutralizing antibodies in the presence or absence of HIV-196ZM651 gp120, SDF-1α, or MIP-1α. SDF-1α and MIP-1α are natural ligands for CXCR4 and CCR5, respectively. Primary cultures of rat astrocytes were incubated for 1 h in the presence of 1.0 μg/ml anti-CXCR4 polyclonal antibody and/or 1.0 μg/ml 3A9 monoclonal CCR5 antibody. In our hands, these antibody concentrations blocked the biological activity (i.e., secretion of cytokines) of SDF-1α and MIP-1α at their respective receptor as determined by ELISA analysis. After preincubation with the neutralizing antibodies, 1.0 nM HIV-196ZM651 gp120, 1.25 pM SDF-1α, or 1.0 pM MIP-1α was added to each culture flask. At 6, 12, and 24 h, the medium was collected for ELISA analysis, and the cells were collected for immunoblot analysis.
Cytokine exposure experiments were initiated by aspirating the culture medium and adding fresh medium containing 0.5 or 10 ng/ml TNF-α, 0.4 or 10 ng/ml IL-1β, or 0.3 or 10 ng/ml IL-6. These cytokines were selected because their expression is known to be increased during HIV-1-associated immunological responses in the brain (Kaul et al., 2005). Untreated cells in 10% fetal bovine serum containing culture medium were used as control. At 6, 12, and 24 h, the medium was aspirated, and the cells were collected for immunoblot analysis.
Treatment of primary cultures of rat astrocytes with cytokine-neutralizing antibodies and HIV-196ZM651 gp120 was conducted by aspirating culture medium and replacing it with fresh medium containing the cytokine-neutralizing antibody and 1.0 nM HIV-196ZM651 gp120. At 6, 12, and 24 h, the medium was aspirated, and the cells were collected for immunoblot analysis. Concentrations for the neutralizing antibodies were determined from cytokine-activity curves provided by the manufacturer. For these experiments, the following concentrations were selected because they completely neutralized the biological activity of their respective cytokine: 0.2 μg/ml TNF-α-neutralizing antibody, 0.5 μg/ml IL-1β-neutralizing antibody, and 0.5 μg/ml IL-6-neutralizing antibody.
Cell Viability Assay. Cell viability was assessed in primary cultures of rat astrocytes treated with HIV-196ZM651 gp120 using a standard MTT assay (Denizot and Lang, 1986) with a few modifications. In brief, cells were plated in a 96-well assay plate at a density of 105 cells/well. After 24 h, the medium was aspirated and replaced with fresh medium containing various concentrations of HIV-196ZM651 gp120 (0.5, 1.0, 10, 25, and 100 nM). The cultures were then incubated for 20 h at 37°C in 5% CO2/95% humidity, at which time 10 μl of MTT (5.0 mg/ml; Sigma-Aldrich) was added to each well. After an additional4hof incubation, 100 μl of extraction buffer, pH 4.7, which contained 20% (w/v) SDS, 50% (v/v) dimethylformamide, 2.0% (v/v) acetic acid, and 0.025 N HCl, was added to each well. The formazan content of each well was determined by UV spectrophotometry (570 nm) using a SpectraMax 384 microplate reader (Molecular Devices, Sunnyvale, CA).
Semiquantitative RT-PCR Analysis. Primary cultures of rat astrocytes and microglia were lysed using TRIzol reagent (Invitrogen Canada Inc., Burlington, ON, Canada). Cell lysates were incubated for 3 min at room temperature with chloroform/isoamyl alcohol (49:1) to eliminate contamination of genomic DNA. The lysates were then centrifuged at 11,500g (4°C, 15 min) to separate the organic and the aqueous phases. The aqueous layer was removed, treated with icecold isopropyl alcohol, and centrifuged at 11,500g (4°C, 15 min) to isolate cellular RNA. RT was performed by preparing a reaction mixture of 0.5 μg of oligo(dt)(12-18) primer and 5 μg of RNA in a 20-μl solution containing 20 mM Tris-HCl, pH 8.4, 50 mM KCl, 5 mM MgCl2, 5 mM dithiothreitol, 0.5 mM dNTP,2Uof DNase I, and 200 U of SuperScript II reverse transcriptase (Invitrogen). PCR was conducted with 1.5 mM MgCl2, 0.5 mM dNTP, 0.5 μM forward and reverse primers,1Uof platinum Taq DNA polymerase (Invitrogen), and 10% cDNA (i.e., RT product) using a GeneAmp 2400 Thermocycler (Applied Biosystems, Foster City, CA) as described in our previous studies (Lee et al., 2001; Ronaldson et al., 2004a). Primer sequences used for these experiments are presented in Table 1. Primer sequences for mdr1a, mdr1b, and β-actin have been described in our previous publications (Lee et al., 2001; Ronaldson et al., 2004a). All other primer sets were designed using GeneFisher Interactive Primer Design Software (F. Meyer and C. Schleiermacher, 1995-1999), available online at http://bibiserv.techfak.uni-bielefeld.de/genefisher. Primers were always chosen according to the following parameters: length between 18 and 25 bases, optimal 20 to 22 bases, and melting temperature between 54 and 65°C. In addition, the forward and reverse primers were designed to span different exons, ensuring that the cDNA amplification product would be a different length from that of contaminant genomic DNA sequences. The PCR program consisted of an initial denaturation at 95°C for 5 min followed by 30 cycles of three phases: denaturing phase (30-60 s at 94°C), annealing (30-60 s, see Table 1 for temperature conditions), and extension (60 s at 72°C). After 11 to 14 cycles, primers for the appropriate housekeeping gene (e.g., β-actin and glyceraldehyde-3-phosphate dehydrogenase) were added to each reaction tube, and the PCR was continued until completion as described previously (Wong et al., 1994). Amplified cDNA products were resolved on 1.7% agarose gels, stained with 1.0 mg/ml ethidium bromide, and visualized by UV transilluminescence. Densitometric analysis of ethidium bromide-stained gels was performed using ImageQuant 5.2 densitometric software (Molecular Dynamics, Sunnyvale, CA). The ratio between the target mRNA and the appropriate housekeeping gene was calculated to obtain the relative mRNA expression of the particular gene of interest. Results are expressed as mean ± S.D. after normalization to the appropriate housekeeping gene.
List of primers and conditions for RT-PCR analysis
Western Blotting. Crude membranes from primary cultures of rat astrocytes and MDCK-MDR1 were prepared by centrifuging the cell suspension at 400g (4°C, 10 min). The supernatant was discarded, and the pellet was lysed for 30 min at 4°C in 250 mM sucrose buffer containing 1.0 mM EDTA and 0.1% (v/v) protease inhibitor cocktail (Sigma-Aldrich). The cell suspension was incubated for 15 min at 4°C and then homogenized with a Dounce homogenizer at 10,000 rpm for three cycles of 15 s each. Homogenates were collected and centrifuged at 3000g for 10 min to remove cellular debris. The supernatant was collected and centrifuged at 100,000g for 1 h at 4°C. The resultant pellet was resuspended in 10 mM Tris buffer, pH 8.8, and frozen at -20°C until further use. Protein concentration of the crude membrane preparations was determined with Bradford's protein assay.
For immunoblotting, 35-μg aliquots of crude membrane proteins were mixed in Laemmli buffer and resolved on a 10% SDS-polyacrylamide gel. The gel was then electrotransferred onto a polyvinylidene difluoride (PVDF) membrane. Protein transfer was verified by Ponceau S staining. The membranes were blocked overnight at 4°C in Tris-buffered saline (15 mM Tris-HCl and 150 mM NaCl, pH 7.6) containing 0.05% (v/v) Tween 20 and 5% (mass per volume) dry skim milk powder. After six washes (5 min each) with Tris-buffered saline/Tween 20, the membrane was incubated with the appropriate primary antibody for4 h at room temperature. P-gp protein expression was assessed using the monoclonal C219 antibody, which recognizes a conserved intracellular epitope on mdr1a, mdr1b, and mdr2. Actin expression was detected using the monoclonal AC-40 antibody, which, according to the manufacturer's data sheet, recognizes a conserved C-terminal epitope on all actin isoforms. After a second wash, the membranes were incubated for 1.5 h in the presence of anti-mouse (Serotec Inc., Raleigh, NC) or anti-rabbit (Sigma-Aldrich) horseradish peroxidase-conjugated secondary antibodies (1:3500 dilution) in 5% milk at room temperature. Protein bands were detected by enhanced chemiluminescence and exposed to X-ray film for 1 min. The MDCK-MDR1 cell line was used as a positive control for the P-gp blots.
ELISA. Ultrasensitive ELISA kits for the detection of rat TNF-α, IL-1β, and IL-6 were purchased from Pierce Biotechnology (Rockford, IL) and were used to determine the secretion of cytokines from primary cultures of rat astrocytes triggered with HIV-196ZM651 gp120 in the presence or absence of blocking antibodies for CXCR4 and CCR5. In addition, these ELISA kits were used to measure cytokine secretion from cultured rat astrocytes triggered with endogenous ligands for CXCR4 (i.e., SDF-1α) or CCR5 (i.e., MIP-1α). Standard curves for TNF-α (0-2500 pg/ml), IL-1β (0-2500 pg/ml), and IL-6 (0-2000 pg/ml) were generated using the appropriate recombinant rat cytokines, and the assays were performed according to the manufacturer's instructions. Absorbance was read at 450 nm using a SpectraMax Plus384 microplate spectrophotometer (Molecular Devices). The concentration of secreted TNF-α, IL-1β, and IL-6 is expressed as picograms per milliliter. All experiments reflect eight separate measurements obtained from different cell cultures on different days.
Functional Studies. These studies were performed on confluent monolayers of rat astrocytes grown on 48-well polystyrene plates (Becton-Dickinson) at a density of approximately 8 × 104 cells/well. The uptake/accumulation of [3H]digoxin and [14C]saquinavir were determined as described previously in our laboratory (Ronaldson et al., 2004a,b). In brief, the cells were washed and incubated at 37°C for 30 min in Earle's balanced saline solution, pH 7.4, containing 1.8 mM CaCl2, 5.4 mM KCl, 0.8 mM MgSO4, 138 mM NaCl, 1.0 mM Na2HPO4, 5.5 mM d-glucose, and 20 mM HEPES. The cells were then incubated for the desired time with [3H]digoxin (100 nM) or [14C]saquinavir (0.78 μM) in the presence or absence of P-gp inhibitors (1.0 μM PSC833, 1.0 μM GF120918) or an Mrp1 inhibitor (5.0 μM MK571). At the end of each time point, the incubation medium was aspirated, and the reaction was terminated with 750 μl of ice-cold 0.16 M NaCl solution. The cells were then solubilized with 375 μl of 1.0 M NaOH for 30 min and transferred to 7.0-ml scintillation vials containing 187.5 μl of HCl (2.0 M) to neutralize the NaOH. [3H]Digoxin or [14C]saquinavir cellular accumulation was measured by a Beckman liquid scintillation counter with automated quench correction. All samples were corrected for “zero time” and background radioactivity. Uptake/accumulation of [3H]digoxin or [14C]saquinavir was standardized to the protein concentration (milligrams per milliliter) determined by the Bradford colorimetric method using bovine serum albumin (Sigma-Aldrich) as the standard. Cellular accumulation of digoxin was expressed as picomoles per milligram of protein, whereas cellular accumulation of saquinavir was expressed as micromoles per milligram of protein. At the concentrations used, digoxin, saquinavir, and the various P-gp/Mrp1 inhibitors did not compromise cell viability as determined by an MTT assay. In addition, the distribution of the radiolabeled probe in the extracellular space in both untreated cells and in cells treated with 1.0 nM HIV-196ZM651 gp120, as measured by [14C]d-mannitol (100 nM), was low (i.e., less than 1%), indicating the presence of a confluent cell monolayer and minimal leakage of digoxin or saquinavir.
Data Analysis. Each set of experiments was repeated at least three times in cells pertaining to different isolations. In an individual experiment, each data point represents quadruplicate trials. Results are reported as a mean ± S.D. from at least three separate experiments. To determine the significance of transport inhibition, the Student's t test was used for unpaired experimental data. For multiple comparisons, the test of repeated-measured analysis of variance and the post hoc multiple-comparison Bonferroni t test was used. A value of p < 0.05 was considered to be statistically significant.

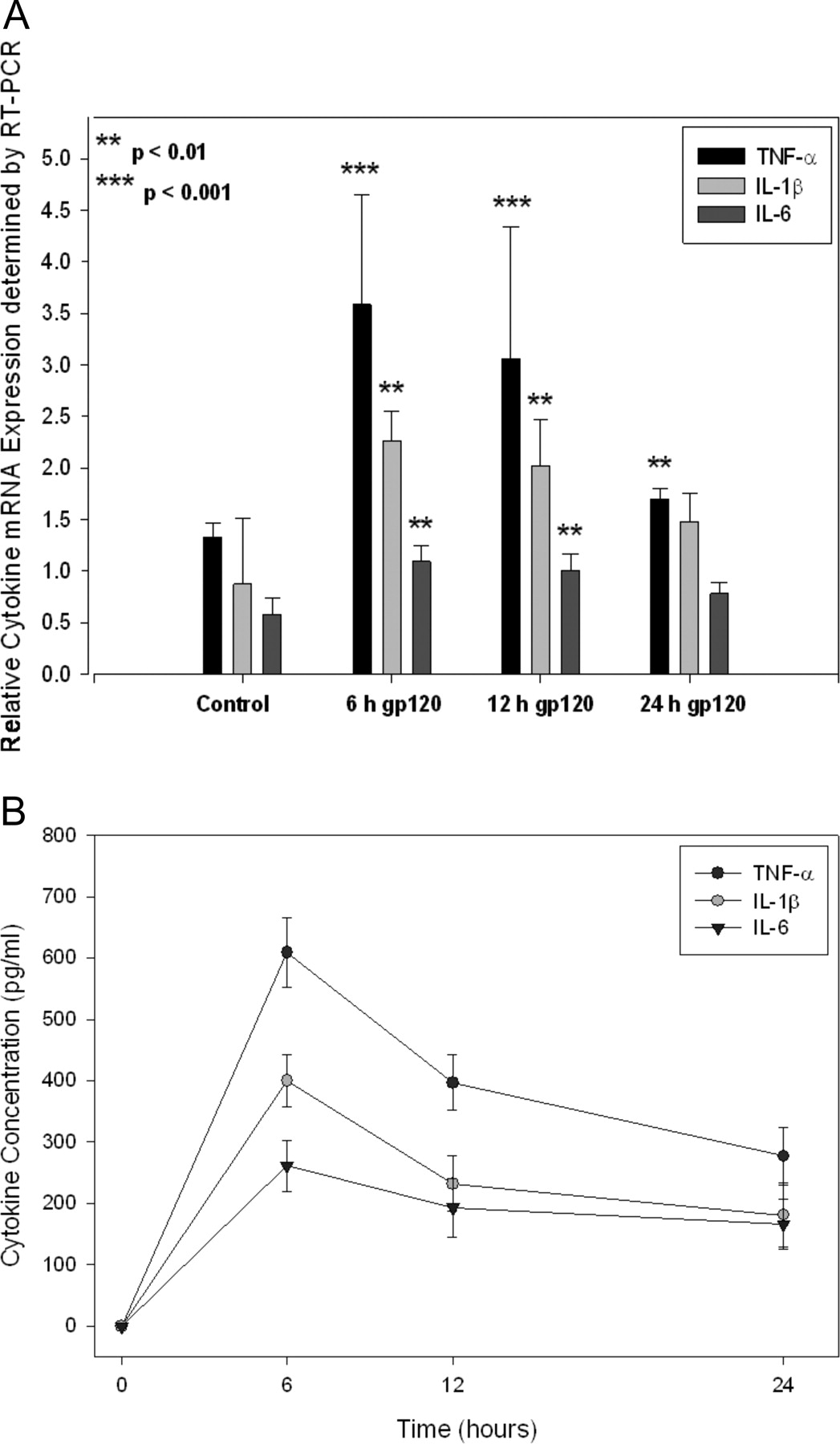
Expression of HIVE markers (i.e., proinflammatory cytokines) in primary cultures of rat astrocytes triggered with the HIV-196ZM651 gp120. A, densitometric analysis of TNF-α, IL-1β, and IL-6 mRNA expression in primary cultures of rat astrocytes either not treated (i.e., control) or treated with 1.0 nM HIV-196ZM651 gp120 for 6, 12, and 24 h. Levels of mRNA were evaluated by semiquantitative RT-PCR. Results are expressed as mean ± S.D. of three separate experiments. *, data points that are significantly different from control. B, ELISA analysis of TNF-α, IL-1β, and IL-6 protein expression in primary cultures of rat astrocytes treated with the HIV-196ZM651 gp120. Cells were treated with HIV-196ZM651 gp120 for 6, 12, and 24 h, and protein levels were examined using ELISA. Data points are expressed as mean ± S.D. of eight separate measurements obtained from different cultures on different days.
Results
Chemokine Receptor Gene and Protein Expression. Previous studies have shown that the chemokine receptors CXCR4 and CCR5 are expressed in cultured rat astrocytes (Bajetto et al., 1999; Koller et al., 2002). In the present study, we confirmed the expression of both CXCR4 and CCR5 in our primary cultures of rat astrocytes. Using RT-PCR analysis, we detected both CXCR4 (∼224 bp) and CCR5 (∼431 bp) mRNA transcripts (data not shown). Immunoblot analysis using the polyclonal anti-CXCR4 antibody and the 3A9 monoclonal CCR5 antibody detected single bands for CXCR4 and CCR5 at the appropriate molecular weights (data not shown). Taken together, these data provide evidence for the molecular (i.e., gene and protein) expression of both CXCR4 and CCR5 in primary cultures of rat astrocytes and suggest that treatment with HIV-196ZM651 gp120 could elicit an immunological response in this cell culture system.
Cell Viability. We performed an MTT assay in primary cultures of rat astrocytes to determine a working concentration of HIV-196ZM651 gp120 that did not significantly alter cell proliferation and viability. At HIV-196ZM651 gp120 concentrations of 0.5 and 1.0 nM, cell viability was not significantly different from control (i.e., untreated) cultures (data not shown). However, cell viability was significantly (p < 0.01) compromised (i.e., more than 25% of cells were dead) at concentrations of 10 nM or greater (data not shown). Based on these data, an HIV-196ZM651 gp120 concentration of 1.0 nM was used in our experiments.
Production/Secretion of Cytokines. Previous studies have demonstrated that gp120 concentrations in excess of 1.0 nM can induce cytokine secretion in cultured astrocytes and microglia (Kong et al., 1996). Therefore, to verify the cytokine response in cultured astrocytes, we measured the gene expression of TNF-α, IL-1β, and IL-6 using semiquantitative RT-PCR analysis. Using gene-specific primers (Table 1), we observed increased (p < 0.01) expression of TNF-α, IL-1β, and IL-6 mRNA in primary cultures of rat astrocytes triggered with 1.0 nM HIV-196ZM651 gp120 (Fig. 1A) compared with untreated cells (i.e., control). In this experiment, the greatest increase in mRNA expression was detected for TNF-α, followed by IL-1β and IL-6. Ultrasensitive ELISA analysis (Fig. 1B) detected significantly increased TNF-α, IL-1β, and IL-6 protein in cell culture supernatants at all time points examined (i.e., 6, 12, and 24 h). Similar to the RT-PCR experiments, TNF-α was present in the cell culture supernatant at the greatest concentrations, followed by IL-1β and IL-6. TNF-α, IL-1β, and IL-6 secretion in untreated cells and in cultures treated with 1.0 nM denatured (i.e., heat-inactivated) HIV-196ZM651 gp120 were lower than the detection limit of the assay (data not shown). Taken together, these data provide evidence for increased cytokine production in cultured rat astrocytes triggered with HIV-196ZM651gp120.
Effect of CXCR4/CCR5 on Cytokine Production. To determine whether HIV-196ZM651 gp120 triggered cytokine secretion was mediated through binding to chemokine receptors, we examined the effect of CXCR4 and CCR5 inhibitory antibodies on the secretion of TNF-α, IL-1β, and IL-6 using ultrasensitive ELISA analysis. In the presence of 1.0 mg/ml polyclonal anti-CXCR4 antibody and 1.0 mg/ml 3A9 anti-CCR5 antibody, 1.0 nM HIV-196ZM651 gp120 did not induce the secretion of any of the cytokines examined (Table 2). Secretion of TNF-α, IL-1β, and IL-6 was significantly enhanced (p < 0.001) in cultured astrocytes treated with HIV-196ZM651 gp120 and the anti-CXCR4 antibody but was inhibited when the cultures were treated with HIV-196ZM651 gp120 and the 3A9 antibody, suggesting that CCR5 is primarily involved in regulating HIV-196ZM651 gp120-mediated cytokine release.
ELISA analysis of cytokine secretion in cultured astrocytes treated with HIV-196ZM651 gp120
Results are expressed as mean ± S.D. of eight separate measurements obtained from different cultures on different days. All values are compared with untreated controls. Cytokine values are expressed as picograms per milliliter.
To further confirm that CXCR4 and CCR5 were involved in the secretion of cytokines in vitro, we treated primary cultures of rat astrocytes with natural endogenous ligands for CXCR4 (i.e., SDF-1α) and CCR5 (i.e., MIP-1α). In the presence of 1.25 pM SDF-1α, ultrasensitive ELISA analysis detected a significant increase (p < 0.001) in the secretion of TNF-α and IL-1β but not for IL-6 (Table 3). Treatment with 1.0 pM MIP-1α induced the secretion TNF-α, IL-1β, and IL-6, implying that IL-6 secretion was mediated by CCR5 and not by CXCR4. These data were further verified by treating the cultures with SDF-1α or MIP-1α and the chemokine receptor inhibitory antibodies. In SDF-1α-treated cultures, cytokine secretion was negated in the presence of the anti-CXCR4 antibody whereas the 3A9 antibody blocked MIP-1α-mediated cytokine secretion (Table 3). When the cultures were treated with SDF-1α and the 3A9 antibody, the secretion of TNF-α and IL-1β, but not IL-6, was significantly increased (p < 0.001). In the presence of MIP-1α and the anti-CXCR4 antibody, the secretion of all three cytokines examined (i.e., TNF-α, IL-1β, and IL-6) was significantly increased (p < 0.001).
ELISA analysis of cytokine secretion in cultured astrocytes treated with SDF-1α or MIP-1a
Results are expressed as mean ± S.D. of eight separate measurements obtained from different cultures on different days. All values are compared with untreated controls. Cytokine values are expressed as picograms per milliliter.
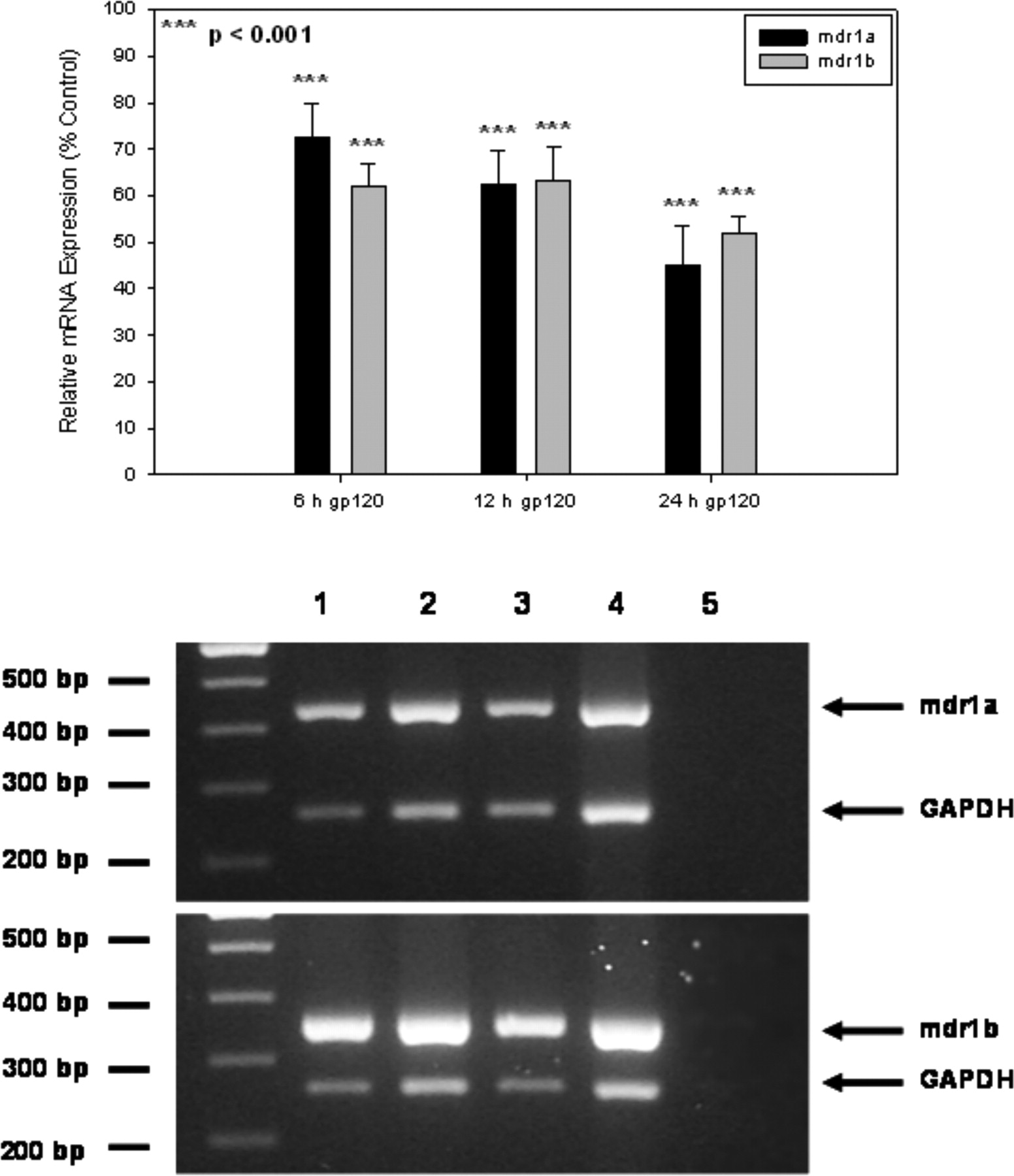
P-gp mRNA and Protein Expression. Semiquantitative RT-PCR analysis was used to assess the expression of mdr1a and mdr1b mRNA in primary cultures of rat astrocytes treated with HIV-196ZM651 gp120. We detected both mdr1 isoforms in control (i.e., untreated) cultures and in cultures treated with HIV-196ZM651 gp120 for 6, 12, and 24 h (Fig. 2). As expected, no bands were detected in the negative control, which consisted of RT product prepared in the absence of mRNA. Densitometric analysis revealed that these decreases were up to 2.2- and 1.9-fold for mdr1a and mdr1b, respectively (Fig. 2).
Immunoblot analysis with the C219 monoclonal antibody was used to detect P-gp in primary cultures of rat astrocytes triggered with HIV-196ZM651 gp120 (Fig. 3). This antibody recognizes a common intracellular epitope of mdr1a, mdr1b, and mdr2. As expected, in the MDCK-MDR1 cell line (the positive control), a single band was observed at approximately 170 to 180 kDa, a size reported previously for P-gp (Gottesman et al., 1995). A single band at approximately 43 kDa corresponding to actin was detected in each lane, which verified appropriate sample loading. In primary cultures of rat astrocytes exposed to 1.0 nM HIV-196ZM651 gp120, P-gp protein expression was significantly decreased compared with control, with maximal suppression (4.7-fold) observed after 24 h of treatment (Fig. 3). P-gp protein expression was not changed in cultures treated with 1.0 nM denatured HIV-196ZM651 gp120, implying that a cellular response specific for the native conformation of HIV-196ZM651 gp120 is involved in the suppression of P-gp protein expression.
Functional Studies. To investigate the functional activity of P-gp in primary cultures of rat astrocytes treated with 1.0 nM HIV-196ZM651 gp120, we measured the uptake/accumulation of [3H]digoxin, a cardiac glycoside drug, and [14C]saquinavir, an HIV-1 protease inhibitor. Both of these compounds are established P-gp substrates (DiDiodato and Sharom, 1997). We confirmed that this dose of HIV-196ZM651 gp120 did not alter membrane permeability with the use of the extracellular marker [14C]d-mannitol (data not shown). In these experiments, cells were grown as monolayers and were incubated in the presence or absence of 1.0 nM HIV-196ZM651 gp120 for 24 h. The time course of digoxin (100 nM) accumulation at 37°C (Fig. 4A) shows increasing accumulation until a plateau is reached at approximately 1 h under control conditions, which corroborates data from our previous publication (Ronaldson et al., 2004a). In cultured rat astrocytes treated with 1.0 nM HIV-196ZM651 gp120 for 24 h, digoxin accumulation at 1 h was significantly enhanced (1.8-fold), suggesting a decrease in P-gp functional activity (Fig. 4A). Digoxin accumulation was also examined in the presence and absence of two standard P-gp inhibitors (i.e., PSC833 and GF120918). As expected, under control conditions, the accumulation of digoxin was significantly enhanced in the presence of 1.0 μM PSC833 (1.9-fold) and 1.0 μM GF120918 (1.8-fold) (Fig. 4A). However, in rat astrocyte cultures exposed to 1.0 nM HIV-196ZM651 gp120 for 24 h, neither PSC833 nor GF120918 significantly enhanced digoxin accumulation, suggesting a loss of transport activity. Likewise, saquinavir (0.78 μM) accumulation at 30 min, 1 h, and 2 h was significantly enhanced in cultures treated with HIV-196ZM651 gp120 compared with controls (Fig. 4B). Because saquinavir is also a known Mrp1 substrate (Dallas et al., 2004) and Mrp1 functional expression has been demonstrated in rat astrocyte cultures (Decleves et al., 2000), we also measured its uptake/accumulation in the presence of 5.0 μM MK571, a leukotriene D4 receptor antagonist and potent Mrp1 inhibitor. As expected, in cultures treated with 5.0 μM MK571 in the absence of HIV-196ZM651 gp120, saquinavir accumulation was significantly enhanced (Fig. 4B), which implies that Mrp1 is involved in saquinavir efflux from cultured rat astrocytes. However, MK571 did not significantly enhance saquinavir accumulation in cultures treated with HIV-196ZM651 gp120, suggesting that P-gp may be involved in saquinavir transport to a greater degree than Mrp1 during the immune response in cultured rat astrocytes.

Semiquantitative RT-PCR analysis of mdr1a and mdr1b isoforms in primary cultures of rat astrocytes treated with HIV-196ZM651 gp120. A, a representative ethidium bromide-stained gel shows amplification of specific bands for mdr1a (∼440 bp) and mdr1b (∼355 bp) from primary cultures of rat astrocytes in the absence of HIV-196ZM651 gp120 (lane 1) or treated with HIV-196ZM651 gp120 for 6 (lane 2), 12 (lane 3), or 24 h (lane 4). No bands were observed in the negative controls, which contained water in place of cDNA (lane 5). Densitometric analysis of mdr1a and mdr1b isoforms in cultured rat astrocytes treated with HIV-196ZM651 gp120 is shown in the inset. Results are expressed as mean ± S.D. of three separate experiments. *, data points that are significantly different from control.

P-gp expression in primary cultures of rat astrocytes treated with gp120. Immunoblot analysis of primary cultures of rat astrocytes triggered with 1.0 nM HIV-196ZM651 gp120 (6, 12, and 24 h). Crude membrane preparations (35 μg) from primary cultures of rat astrocytes and MDCK-MDR1 cells were resolved on a 10% SDS-polyacrylamide gel and transferred to a PVDF membrane. P-gp was detected using the monoclonal antibody C219 (1:500 dilution). Densitometric analysis of P-gp protein expression in primary cultures of rat astrocytes treated with 1.0 nM HIV-196ZM651 gp120 and 1.0 nM denatured HIV-196ZM651 gp120 is shown in the inset. Results are expressed as mean ± S.D. of three separate experiments. *, data points that are significantly different from control.
Effect of Cytokines on P-gp Protein Expression. Because cytokine secretion was increased in the presence of HIV-196ZM651 gp120, we explored their role in the regulation of P-gp expression. Immunoblot analysis of primary cultures of rat astrocytes treated with TNF-α or IL-1β or IL-6 was performed. Increased P-gp expression was observed in cultures treated with TNF-α (2.9-fold) or IL-1β (1.6-fold) (Fig. 5, A and B). In contrast, P-gp expression was substantially decreased (8.9-fold) in cells treated with IL-6 (Fig. 5, A and B). Appropriate loading of each sample was confirmed by the detection of a single band at approximately 43 kDa, which corresponds to actin.
To further elucidate the role of cytokines in altering P-gp expression, we measured P-gp protein expression in primary cultures of rat astrocytes treated with HIV-196ZM651 gp120 and cytokine-neutralizing antibodies. In the presence of the three cytokine-neutralizing antibodies only, P-gp protein expression was not significantly altered in our rat astrocyte cultures (Fig. 6, A and B). However, immunoblot analysis demonstrated decreased P-gp expression when cultures were treated with HIV-196ZM651 gp120 and the TNF-α or IL-1β neutralizing antibody. In contrast, P-gp protein expression was not significantly altered in primary cultures of rat astrocytes treated with HIV-196ZM651 gp120 and the IL-6-neutralizing antibody, suggesting that IL-6 but not TNF-α or IL-1β is involved in the down-regulation of P-gp protein expression.

Accumulation of P-gp substrates in cortical rat astrocytes treated with HIV-196ZM651 gp120. A, effect of 24-h HIV-196ZM651 gp120 treatment on the accumulation of digoxin by cortical rat astrocytes monolayer cells. [3H]Digoxin (100 nM) accumulation was measured at 37°C in the presence of 1.0 nM HIV-196ZM651 gp120. Results are expressed as mean ± S.D. of three separate experiments, with each data point in an individual experiment representing quadruplicate measurements. *, data points that are significantly different from control. Effect of P-glycoprotein inhibitors on the accumulation of digoxin by cortical rat astrocyte monolayer cells treated with HIV-196ZM651 gp120 for 24 h is shown in the inset. The accumulation of [3H]digoxin (100 nM) was measured at 37°C in the presence of 1.0 μM PSC833 or 1.0 μM GF120918. Results are expressed as mean ± S.D. of three separate experiments, with each data point in an individual experiment representing quadruplicate measurements. *, data points that are significantly different from control. B, effect of 24-h HIV-196ZM651 gp120 on the accumulation of saquinavir by cortical rat astrocyte monolayer cells. [14C]Saquinavir was measured at 37°C in the presence of 1.0 nM HIV-196ZM651 gp120 in the presence or absence of 5.0 μM MK571. Results are expressed as mean ± S.D. of three separate experiments, with each data point in an individual experiment representing quadruplicate measurements. *, data points that are significantly different from control.
Discussion
Astrocytes, the most abundant central nervous system cell type, are involved in several homeostatic functions, such as regulating neurotransmitter concentrations (i.e., glutamate, GABA), secreting neurotrophic factors, and maintaining BBB integrity (Speth et al., 2005). Because astrocytes can also synthesize messenger molecules and form intercellular networks, they also participate in brain immune responses (Speth et al., 2005). Astrocytes produce cytokines and chemokines, enabling the regulation of immune signaling in both normal and diseased brain (Speth et al., 2005). In vitro studies have demonstrated that astrocytes, when triggered by exogenous mediators [e.g., LPS, poly(inosinic acid)/poly-(cytidylic acid)], secrete cytokines (e.g., TNF-α, IL-6) (Carpentier et al., 2005; Jack et al., 2005). However, little is known about these immune responses in the presence of HIV-1 viral proteins.
In the present work, we examined the ability of gp120, the HIV-1 viral envelope glycoprotein, to trigger the innate immune response in primary cultures of rat astrocytes. HIV-1 cellular entry is initiated by an interaction between gp120 and CD4, followed by gp120 binding to HIV-1 coreceptors (e.g., CXCR4, CCR5). In addition, shed gp120 from HIV-1-infected macrophages and microglia can activate intracellular signaling cascades in CD4-negative cell types by binding directly to CXCR4 and CCR5 (Persidsky and Gendelman, 2003; Kaul et al., 2005). Although astrocytes do not express CD4 (Liu et al., 2004), studies have reported expression of both coreceptors in these cells (Bajetto et al., 1999; Koller et al., 2002). It is interesting that Kong et al. (1996) reported cytokine secretion (i.e., TNF-α, IL-1β, and IL-6) in cultured murine astrocytes triggered with subtype B gp120 isoforms. In addition, it has been demonstrated that gp120 may interact with rodent CXCR4 and CCR5 orthologs (Wu et al., 1997; Koller et al., 2002). Therefore, the secretion of cytokines may be facilitated by interactions between shed gp120 and CXCR4/CCR5. For all of our experiments, we used an HIV-196ZM651 gp120 concentration of 1.0 nM because results from our present study as well as others (Kanmogne et al., 2002; Iskander et al., 2004) have shown that gp120 concentrations in excess of 5 nM are cytotoxic. When our astrocyte cultures were treated with HIV-196ZM651 gp120, an increase in both mRNA transcript expression and secretion of TNF-α, IL-1β, and IL-6 was observed, suggesting that HIV-196ZM651 gp120 elicits an immune response. Furthermore, cytokines were not secreted when astrocyte cultures were treated with denatured HIV-196ZM651 gp120, implying that secretion of TNF-α, IL-1β, and IL-6 is not a generalized response to foreign protein.

Effect of cytokines on P-gp protein expression in primary cultures of rat astrocytes. A, primary cultures of rat astrocytes were treated with TNF-α (0.5 and 10 ng/ml), IL-1β (0.4 and 10 ng/ml), and IL-6 (0.3 and 10 ng/ml) for 6, 12, and 24 h, and P-gp expression was assessed by immunoblot analysis. Crude membrane preparations of primary cultures of rat astrocytes and MDCK-MDR1 cells (35 μg) were resolved on a 10% SDS-polyacrylamide gel and transferred to a PVDF membrane. The blots were incubated with the monoclonal P-gp antibody C219 (1:500 dilution). Equal sample loading was confirmed by the detection of actin using the monoclonal antibody AC40 (1:500 dilution). Primary cultures of rat astrocytes not exposed to cytokines were used as a control. B, densitometric analysis of P-gp protein in cultured rat astrocytes treated with TNF-α, IL-1β, or IL-6. Results (percentage of control) are expressed as mean ± S.D. of three separate experiments. *, data points that are significantly different from control.
Based on these observations, we hypothesized that HIV-196ZM651 gp120 mediates cytokine secretion by binding to CXCR4 and/or CCR5. Therefore, we examined the ability of these receptors to trigger cytokine secretion by treating primary cultures of rat astrocytes with endogenous ligands for CXCR4 (i.e., SDF-1α) or CCR5 (i.e., MIP-1α). Previous studies in cultured murine astrocytes (Han et al., 2001) have shown that both ligands induce TNF-α, IL-1β, and IL-6 secretion. In our hands, secretion of both TNF-α and IL-1β was increased in the presence of SDF-1α or MIP-1α, implying that cytokine secretion may result from direct stimulation of CXCR4 and CCR5. It is interesting that IL-6 secretion was enhanced only by treatment with MIP-1α, suggesting that CCR5 signaling can mediate IL-6 secretion in astrocyte cultures.

Effect of cytokine-neutralizing antibodies on P-gp protein expression in primary cultures of rat astrocytes treated with HIV-196ZM651 gp120. A, primary cultures of rat astrocytes were treated with neutralizing antibodies for TNF-α (0.2 ng/ml), IL-1β (0.5 ng/ml), and IL-6 (0.5 ng/ml) in the presence or absence of 1.0 nM HIV-196ZM651 gp120 for 6, 12, and 24 h, and P-gp expression was assessed by immunoblot analysis. Crude membrane preparations of primary cultures of rat astrocytes and MDCK-MDR1 cells (35 μg) were resolved on a 10% SDS-polyacrylamide gel and transferred to a PVDF membrane. The blots were incubated with the monoclonal P-gp antibody C219 (1:500 dilution). Equal sample loading was confirmed by the detection of actin using the monoclonal antibody AC40 (1:500 dilution). Primary cultures of rat astrocytes not exposed to HIV-196ZM651 gp120 or to the cytokine-neutralizing antibodies were used as a control. B, densitometric analysis of P-gp protein in cultured rat astrocytes treated with HIV-196ZM651 gp120 and various cytokine neutralizing antibodies. Results (percentage of control) are expressed as mean ± S.D. of three separate experiments. *, data points that are significantly different from control. NAb, neutralizing antibody.
HIV-196ZM651 gp120 is derived from an R5-tropic viral strain, which specifically interacts with CCR5 before host cell infection. Therefore, we expected that the immune response observed in rat astrocyte cultures treated with HIV-196ZM651 gp120 was CCR5-mediated. We confirmed this hypothesis by treating these cultures with HIV-196ZM651 gp120 and specific inhibitory antibodies for CXCR4 and/or CCR5. In cultures treated with HIV-196ZM651 gp120 and both antibodies, cytokine secretion was not detected. Because negligible cytokine secretion was observed in the presence of the CCR5 inhibitory antibody but not with the CXCR4 inhibitory antibody, these data imply that cytokine release by HIV-196ZM651 gp120 is primarily mediated by CCR5.
P-gp is an important determinant of drug permeation across the BBB and brain cellular compartments (e.g., astrocytes, microglia). This is of particular significance in the treatment of HIV-1 infection because many antiretroviral agents, including HIV-1 protease inhibitors, are known P-gp substrates (Lee et al., 1998; Ronaldson et al., 2004b). We have described previously P-gp functional expression in healthy (i.e., nonpathological) cultures of rat astrocytes (Ronaldson et al., 2004a). To date, the effect of gp120-triggered innate immune responses on the molecular expression and functional activity of P-gp in astrocytes has not been examined. In the present study, we report decreased mdr1a (2.2-fold) and mdr1b (1.9-fold) mRNA expression and decreased P-gp protein expression (4.7-fold) in HIV-196ZM651 gp120-treated cultures.
To determine whether decreased P-gp gene and protein expression was correlated with decreased functional activity, we used digoxin and saquinavir, two well-known P-gp substrates (DiDiodato and Sharom, 1997). Our studies showed that HIV-196ZM651 gp120 treatment significantly enhanced digoxin accumulation at various time points by rat astrocyte monolayers without an alteration of the membrane permeability at any of the time points examined. In addition, P-gp inhibitors (i.e., PSC833 and GF120918) did not enhance digoxin accumulation in monolayers of cortical rat astrocytes treated with HIV-196ZM651 gp120, suggesting a loss of P-gp-mediated transport activity. Our saquinavir experiments also showed significantly increased accumulation at 30 min, 1 h, and 2 h in HIV-196ZM651 gp120-treated rat astrocyte monolayers. Because saquinavir is also a known Mrp1 substrate (Dallas et al., 2004), we measured saquinavir accumulation in the presence of MK571, a leukotriene D4 antagonist and potent MRP inhibitor. Under control conditions, MK571 significantly enhanced saquinavir accumulation in rat astrocyte monolayers. In contrast, MK571 did not significantly enhance saquinavir accumulation in HIV-196ZM651 gp120-treated astrocyte monolayers, suggesting that P-gp is more prominently involved during the brain immune response.
Previous studies have shown that cytokines (e.g., TNF-α, IL-1β, and IL-6) can alter P-gp molecular expression and functional activity (Sukhai et al., 2001; Hartz et al., 2006). In vitro studies using isolated rat hepatocytes observed reduced P-gp expression and activity in cells treated with IL-6 (Sukhai et al., 2001). Recent studies in isolated rat brain capillaries demonstrated that TNF-α treatment abolished NBD-CSA accumulation in the capillary lumen without altering P-gp protein expression, suggesting a loss of P-gp efflux activity (Hartz et al., 2006). It seems that this effect occurs via activation of the endothelin-B receptor, leading to increased nitric-oxide synthase activity and protein kinase C activation (Hartz et al., 2004, 2006). Although primary cultures of rat astrocytes express the endothelin-B receptor (Koyama et al., 2003), it is unknown whether this signaling pathway is involved in the regulation of P-gp activity in these cells. Because our data demonstrated that primary cultures of rat astrocytes treated with HIV-196ZM651 gp120 secrete TNF-α, IL-1β, and IL-6, we used immunoblot analysis to investigate the P-gp protein expression in cultured astrocytes treated with these cytokines. Although we observed increases in P-gp protein expression in cultures treated with TNF-α (2.9-fold) and IL-1β (1.6-fold), protein expression was profoundly decreased in the presence of IL-6 (8.9-fold). Taken together, these data imply that cytokines, particularly IL-6, are involved in altering P-gp protein expression during activation of the brain's innate immune response.
We further examined the role of cytokines on P-gp protein expression by treating primary cultures of rat astrocytes with HIV-196ZM651 gp120 in the presence of TNF-α-, IL-1β-, or IL-6-neutralizing antibodies. Our results demonstrate that P-gp expression was not altered in cultures treated with the IL-6 neutralizing antibody but was significantly decreased when the TNF-α- or IL-1β-neutralizing antibodies were used. These data further confirm that IL-6 secretion is the principal cytokine involved in the down-regulation of P-gp expression in HIV-196ZM651 gp120-treated astrocytes.
At present, the intracellular signaling mechanisms by which HIV-196ZM651 gp120 and cytokines alter P-gp functional expression remain unclear. Several transcription factors, including nuclear factor-κB, c-Jun, and the pregnane-X-receptor (PXR in rodents; SXR in humans), have been shown to be involved in the regulation of MDR1/mdr1a/mdr1b gene expression (Miao and Ding, 2003; Chen et al., 2005; Hayashi et al., 2005). For example, nuclear factor-κB, a well-characterized transcription factor, has been shown to regulate mdr1a gene expression in rat brain microvessel endothelial cells treated with the HIV-1 transactivator protein Tat (Hayashi et al., 2005). In contrast, studies in a doxorubicin-resistant human leukemia cell line (MDR K562/A02) have shown that c-Jun activation decreases MDR1 mRNA expression (Miao and Ding, 2003). A recent in vivo study observed that LPS treatment downregulates both PXR and mdr1a mRNA expression in mouse placenta (Chen et al., 2005). However, it was not determined whether the observed decrease in mdr1a expression was due to decreased PXR expression. Further studies are required to elucidate which, if any, transcription factors are involved in altering P-gp expression during HIV-196ZM651 gp120 and/or IL-6 treatment.
In summary, this study describes the implementation of an in vitro astrocyte model that can be used to examine the effect of HIV-196ZM651 gp120 in triggering an immune response and the regulation of P-gp functional expression by a viral protein. Our data provide evidence for the involvement of astrocytic cytokines in the modulation of P-gp molecular expression and function during activation of the brain's innate immune response by HIV-1 viral envelope protein gp120. Overall, these observations suggest that complex drug-transporter interactions may occur during HIVE-associated immune responses and may contribute to difficulty in predicting brain distribution of antiviral agents, such as HIV-1 protease inhibitors.
Acknowledgments
We thank Dr. Jason Zastre for advice on the manuscript and Manisha Ramaswamy for excellent technical assistance.
Footnotes
-
This work was supported by grants from the Canadian Institutes of Health Research (grant MOP-56976) and the Ontario HIV Treatment Network, Ontario Ministry of Health. P.T.R. is a recipient of an Ontario HIV Treatment Network Studentship Award.
-
ABBREVIATIONS: HIV-1, human immunodeficiency virus type-1; BBB, blood-brain barrier; ELISA, enzyme-linked immunosorbent assay; HIVE, HIV-1 encephalitis; MDCK-MDR1, Madin-Darby canine kidney MDR1 cell line; MDR, multidrug resistance; MRP, multidrug resistance-associated protein; MTT, 3-(4,5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide; P-gp, P-glycoprotein; PVDF, polyvinylidene diflouride; TNF-α, tumor necrosis factor-α; IL, interleukin; RT-PCR, reverse transcriptase-polymerase chain reaction; LPS, lipopolysaccharide; PCR, polymerase chain reaction; RT, reverse transcriptase; bp, base pair(s); PSC833, valspodar; GF120918, elacridar; SDF-1α, stromal-derived factor-1α; MIP-1α, macrophage inflammatory protein-1α; MK571, 3-[[3-[2-(7-chloroquinolin-2-yl)vinyl]phenyl]-(2-dimethylcarbamoylethylsulfanyl)methylsulfanyl] propionic acid.
- Received April 24, 2006.
- Accepted June 21, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}