


Abstract
The lysophospholipids, lysophosphatidic acid (LPA) and sphingosine 1-phosphate (S1P), are now recognized as important extracellular signaling molecules. These lipid mediators are pleiotropic; among the most common cellular responses are mitogenesis, cell survival (anti-apoptosis), inhibition of adenylyl cyclase and calcium mobilization. Physiologic events associated with these mediators include platelet aggregation, vasopressor activity, wound healing, immune modulation, and angiogenesis. Many of the actions of LPA and S1P are mediated through a set of eight G protein-coupled receptors. Five of these are S1P-prefering while the remaining three are LPA receptors. These receptors are expressed widely and in aggregate signal through a variety of heterotrimeric G proteins. The lysophospholipid receptor family is referred to commonly as the “Edg” group (e.g., Edg-1, Edg-2, etc.). Herein, the molecular pharmacology of the lysophospholipid receptors is reviewed briefly, and a rational nomenclature for LPA and S1P receptors that is consistent with the International Union of Pharmacology guidelines is proposed.
I. Introduction
Lysophosphatidic acid (LPA1) and sphingosine 1-phosphate (S1P) are now known to be pleiotropic extracellular signaling molecules. The first report of lysophosphatidic acid-evoked responses from tissue described its activity as a vasopressor (Tokumura et al., 1978). Subsequently, LPA was found to be proliferative for cultured cells (van Corven et al., 1989)—an observation that spurred much subsequent investigation. S1P has also been shown to be proliferative for cultured cells (Zhang et al., 1991). Investigations into numerous aspects of lysophospholipid (LPL) signaling led to the realization that many LPL-evoked events are mediated by heterotrimeric G proteins, thus predicting the existence of LPL G protein-coupled receptors (GPCRs).
II. Discovery of Lysophospholipid Receptors
The receptor cluster that contains the eight LPL receptors has the colloquial name “Edg” (an acronym for endothelialdifferentiation gene). The name Edg (or EDG) was coined in 1990 by Hla and Maciag to describe a set of immediate early response gene products cloned from human umbilical vein endothelial cells; Edg-1 was found to be a rhodopsin family GPCR (Hla and Maciag, 1990). A GPCR that was 50% identical was reported in 1993 (“AGR16”; Okazaki et al., 1993) and independently the following year (“H218”; MacLennan et al., 1994). The Edg name appeared again in 1995 (Edg-2) to describe a third, more distantly related, GPCR (Masana et al., 1995). The orthologous (species homolog) mouse GPCR was then described (“rec1.3”; Macrae et al., 1996). The other five members of the cluster were named in order of their appearance as Edg-3, Edg-4, Edg-6, Edg-7, and Edg-8 and eventually AGR16/H218 became commonly known as Edg-5.
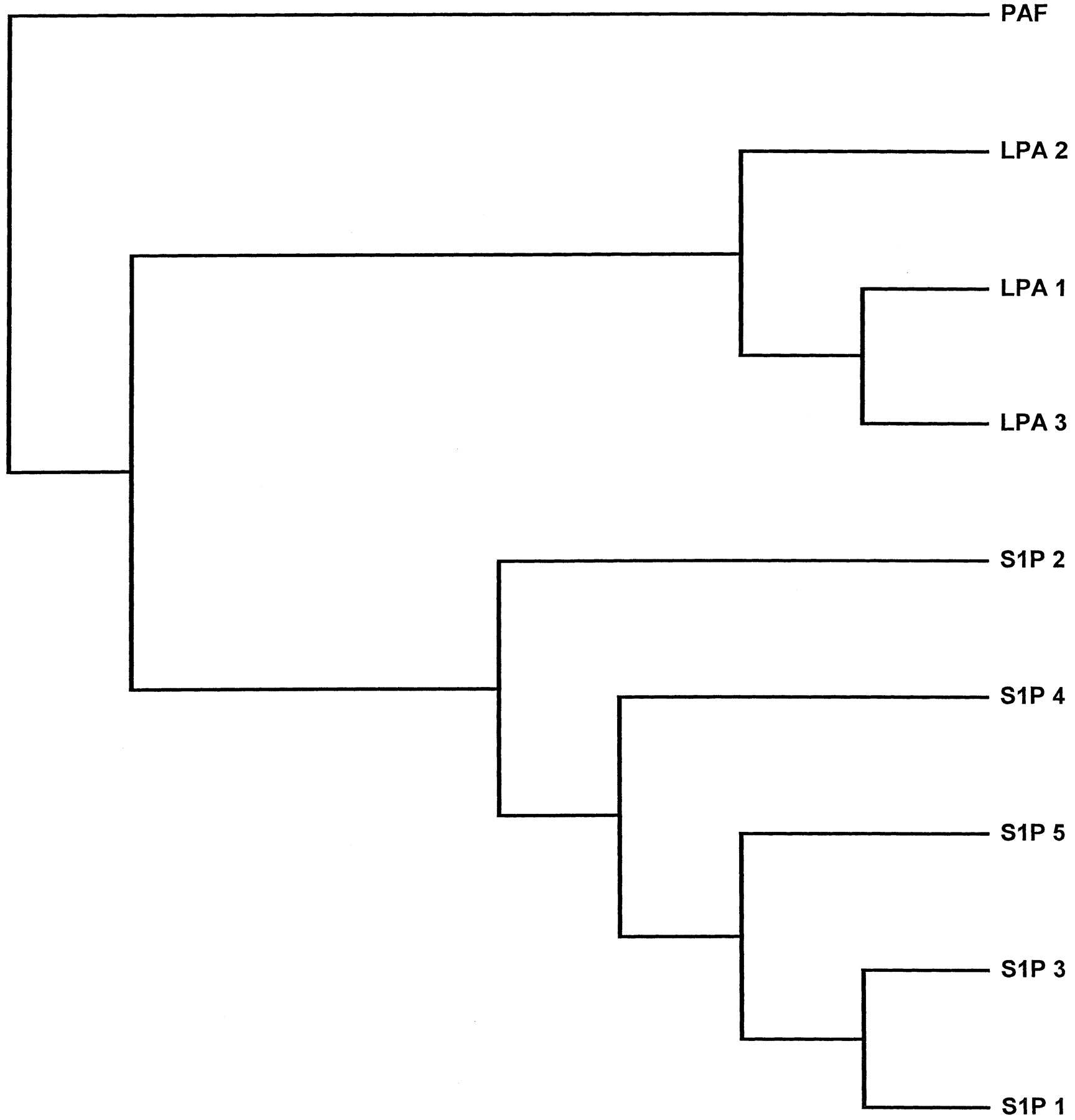
A seminal event in the LPL field occurred when Chun and colleagues discovered that mouse Edg-2 (they called it “vzg-1”) is an LPA receptor (Hecht et al., 1996). This was followed quickly by three reports that Edg-3 and Edg-1 are S1P-preferring receptors (An et al., 1997; Lee et al., 1998; Zondag et al., 1998). Within several years, reports from a number of groups established that there are eight Edg receptor genes in the human genome. Five of these encode S1P receptors (Edg-1, -3, -5, -6, and -8) whereas the remaining three encode LPA receptors (Edg-2, -4, and -7). The S1P receptors share about 50% identical amino acids whereas the LPA receptors have about 55% sequence identity. The subclusters are about 35% identical. A maximum parsimony tree showing graphically the relationship among the LPL receptors and the distantly related platelet-activating factor receptor is shown as the Fig. 1. Different aspects of LPL biochemistry, physiology, and cell biology are discussed in a variety of recent review articles (Hla, 2001; Tigyi, 2002) (also for review, see Chun et al., 1999; Moolenaar, 1999; Pyne and Pyne, 2000;Fukushima et al., 2001; Hla et al., 2001).

The single maximum parsimony tree of the human LPL receptor amino acid sequences and the human platelet-activating factor receptor sequence. This unrooted tree was built with PROTPARS (available at www.evolution.genetics.washington.edu) in PHYLIP package (5 runs, 10–23 jumbles per run).
III. Receptor Nomenclature
Although the Edg acronym proved prescient for Edg-1 (Hla et al., 2001), this name has little relevance to the other seven receptors in the family. The names AGR16, H218, vzg-1, rec1.3, etc. are trivial also. Furthermore, the Edg name has been applied to unrelated proteins and thus is a possible source of confusion (Hla et al., 1997). A rational, alternate nomenclature has been proposed (Chun et al., 1999), but this scheme has not been embraced, and it is not consistent with IUPHAR guidelines (Humphrey et al., 2000; Ruffolo et al., 2000). According to those guidelines, a receptor is to be named with the abbreviation for the natural agonist with the highest potency, followed by a subscripted arabic number. Because the NC-IUPHAR Subcommittee on Lysophospholipid Receptors recommends that sphingosine 1-phosphate be abbreviated S1P (rather than SPP or Sph-P), Edg-1 becomes S1P1 and Edg-2 becomes LPA1. The subcommittee decided further that the order of numbering is to reflect the chronology of the publication of receptor sequence (regardless of whether the ligand was known at that time), thus Edg-5/H218/AGR16 becomes S1P2, etc. Table 1 lists recommended LPL receptor names, the IUPHAR Receptor Code (Humphrey et al., 2000), and previous LPL receptor names. LPL receptor splice variants, such as that described for the human LPA3 receptor (Fitzgerald et al., 2000), are not named individually since they have not been shown to have distinct pharmacologic properties. The recommended receptor nomenclature is flexible in that it readily accommodates additional LPL GPCRs, regardless of their similarity to the Edg cluster. Additional LPA receptors (Guo et al., 1996) as well as sphingosylphosphorylcholine (SPC) (Xu et al., 2000) and lysophosphatidylcholine (LPC) (Kabarowski et al., 2001) receptors have been suggested. However, until data verifying these identifications is published independently, the NC-IUPHAR Subcommittee on Lysophospholipid Receptors has decided against including these putative LPL receptors at this juncture.
LPL receptors
IV. Nonhuman Lysophospholipid Receptors
Current evidence suggests the existence of an orthologous set of eight LPL receptor genes in rodents; presumably this will hold true for all mammals. An analysis of the nearly complete Takifugu rubripes (Japanese puffer fish) genome reveals at least ten LPL receptor-like genes—four LPA receptor-like and six S1P receptor-like (K. R. Lynch, unpublished observation). There is little, if any, evidence for LPL receptors in any nonvertebrate species. The IUPHAR nomenclature should be applied to mammalian receptors only because the nonmammalian LPL receptors are so distant as to make identification of orthologs problematic. For example, the zebrafish gene, miles apart (Mil)—a mutation of which results in cardiac bifida (Kupperman et al., 2000)—has been suggested to be orthologous to the human S1P2 receptor because of their 60% identical amino acids. However, this is an insufficient basis to assume the correspondence in function between Mil and S1P2 that is predicted for orthologous genes—and the deletion of the S1P2 gene in mice does not result in detectable abnormalities in cardiac development (MacLennan et al., 2001). The exception to the rule is the LPA1receptor, which shows a high degree of sequence conservation among chicken, fish, amphibians (Xenopus), and mammals. Remarkably, the LPA1 receptor from all these species shares >90% identical amino acids, with most of the mismatch at the amino-terminal regions. The other LPL receptors—and GPCRs in general—share only 40 to 70% identical amino acids when sequences from disparate vertebrate species are compared. The extraordinary sequence conservation of the LPA1 receptor leads one to wonder whether this protein is serving some function beyond binding LPA and signaling heterotrimeric G proteins.
V. Lysophospholipid Receptor Ligands
A synthetic chemistry focused on LPL receptors is currently underway, but there are very few defining ligands at present. For both LPA and S1P, the addition of a head group (e.g., choline) to form a phosphate diester or the replacement of the phosphate with an alcohol result in decreases in potency of several log orders (for review, seeLynch and Macdonald, 2001). Conversely, the degree of saturation of the alkyl moiety of either ligand has little effect on potency, that is dihydro S1P is equipotent to S1P and 16:0 LPA is equipotent to 18:1 LPA—with the important exception of the LPA3receptor, which has a pronounced preference for unsaturated LPAs (Bandoh et al., 1999; Im et al., 2000b). Recently, two LPA receptor antagonists have been described. The first is di-octyl glycerol pyrophosphate, which is a competitive antagonist of the LPA3 receptor with a reportedK i of 100 nM (Fisher et al., 2001). The second, which is an N-oleoyl ethanolamide phosphate that is substituted at the second carbon with a benzyl-4-oxybenzyl moiety, is a competitive antagonist of the LPA1 and LPA3 receptors (K i values 125 and 430 nM, respectively). The opposite enantiomer (R) of the latter compound is about 10-fold more potent in blocking the LPA3 receptor, but is an agonist at the LPA1 site (Heise et al., 2001).
There are currently no available S1P receptor selective agonists or antagonists, but the availability of a radioligand binding assay allows the measurement of ligand affinities at recombinant S1P receptors. Although no one group has yet reported comparative binding analyses of all five S1P receptors, the reportedK D values from different laboratories (obtained by equilibrium binding methods) are in the range of 1 to 60 nM, with the S1P4 receptor having the lowest affinity for S1P (van Brocklyn et al., 2000). A model of the S1P1 receptor ligand binding domain has been proposed (Parrill et al., 2000) with a specific emphasis on the Arg-Glu-Gly motif that is present at the exofacial aspect of the third transmembrane-spanning region of all S1P receptors. An obvious prediction is that side chains of the arginine and glutamate residues interact with the vicinal phosphate and amino groups on S1P. This prediction was tested by mutating the Arg-Glu-Gly motif to that conserved among LPA receptors (Arg-Gln-Gly), and the ligand selectivity of the mutant receptors (S1P1 and LPA1) was found to switch in concert with the mutations (Wang et al., 2001). Thus the hydrophilic “head group” of the lysophospholipids is thought to interact with the amino-terminal aspect of the third transmembrane region. However, the areas of the receptor protein that interact with the hydrophobic “tail” of the LPL ligands are uncertain at present.
VI. Lysophospholipid Receptor Gene Knockouts
The function of the LPL receptors has been explored by germ line ablation of the individual receptor genes. In addition to the zebrafishmiles apart mutation mentioned above, the LPA1, S1P1, S1P2, and S1P3 receptor genes in mice have been “knocked out”. The S1P3 receptor −/− mice are without obvious phenotype (Ishii et al., 2001) whereas some 20% of S1P2 receptor −/− mice were reported to experience at least one epileptic seizure between 3 and 6 weeks of age (MacLennan et al., 2001). The S1P1 receptor −/− mice die about gestational age E13 from failure of the vasculature to become invested with smooth muscle (Liu et al., 2000). Mice lacking a functional LPA1 receptor gene are born but have a defect in their suckling behavior—apparently because of a defect in olfaction—that in turn results in increased neonatal mortality and stunted growth of survivors (Contos et al., 2000). Analyses of the phenotypes of mice with the remaining four LPL receptor genes ablated are not yet published but are underway in several laboratories.
VII. Lysophospholipid Receptor Expression
Finally, the expression of each of the LPL receptor genes has been examined in rodents and humans. The general lack of high quality antibodies and high affinity radiolabeled ligands necessitates inferring receptor expression by measurement of accumulation of the cognate mRNAs. The LPA1 receptor mRNA is restricted largely (in rodents) to myelinating glia including Schwann cells, but before birth the LPA1 mRNA is abundant in developing cortical neurons (Hecht et al., 1996; Weiner et al., 1998). However, the human LPA1 receptor mRNA is reported to be found in the extract of many tissues including heart, brain, colon, small intestine, and prostate but not in extracts of liver, lung, thymus, or leukocytes (An et al., 1998). LPA2 receptor expression is most prominent in leukocytes (An et al., 1998) whereas the LPA3receptor mRNA is found in extracts of kidney, lung, heart, pancreas, and prostate (Bandoh et al., 1999; Im et al., 2000b). In mouse, all three LPA receptor type mRNAs are prominent in testes extracts (Contos and Chun, 2001). Among the S1P receptors, the S1P1 receptor type is expressed ubiquitously as is the S1P3 receptor (Yamaguchi et al., 1996). The S1P2 receptor is expressed in embryonic brain and postnatally in brain, heart, lung, stomach, intestine, and adrenal gland (Okazaki et al., 1993; MacLennan et al., 1994). The S1P4 receptor is unusual in that its expression is confined to lymphoid tissue (Gräler et al., 1998). Finally, the S1P5 receptor mRNA is found in white matter and spleen in rats (Im et al., 2000a).
VIII. Conclusion
Although there is little likelihood that additional Edg cluster receptor genes will be found, their identification represents only the end of the beginning. The importance of the LPL mediators is evidenced by the steady increase of publications focused on these molecules. The application of selective ligands under development that mimic or block LPLs at their receptors will surely reveal a rich pathophysiology controlled by this signaling system.
Footnotes
Address correspondence to: Dr. Kevin R. Lynch, Subcommittee Chair, Pharmacology Box 800735, University of Virginia Health System, 1300 Jefferson Park Avenue, Charlottesville, VA 22908.
Center for Vascular Biology, Department of Physiology, University of Connecticut, 263 Farmington Ave., Farmington, CT 06030; Yasuyuki Igarashi, Department of Biomembrane and Biofunctional Chemistry, Hokkaido University, Nishi 6, Kita 12, Kita-ku, Sapporo 060-0812, Japan; Kevin R. Lynch (Chair), Department of Pharmacology, Box 800735, University of Virginia, 1300 Jefferson Park Ave., Charlottesville, VA 22908-0735; Wouter H. Moolenaar, Division of Cellular Biochemistry, Netherlands Cancer Institute, Plesmanlaan 121, 1066 CX Amsterdam, The Netherlands; Susan Pyne, Department of Physiology and Pharmacology, University of Strathclyde, 27 Taylor St., Glasgow G4 ONR, Scotland; and Gabor Tigyi, Department of Physiology, University of Tennessee, 894 Union Ave., Memphis, TN 38163.
Composition of the NC-IUPHAR Subcommittee on Lysophospholipid Receptors: Jerold Chun, Merck Research Laboratories, 505 Coast Boulevard South, La Jolla, CA 92037; Edward J. Goetzl, Department of Medicine, University of California, U88B Box 0711, 533 Parnassus Ave., San Francisco, CA 94143-0711; Timothy Hla,
Abbreviations
- LPA
- lysophosphatidic acid
- S1P
- sphingosine 1-phosphate
- LPL
- lysophospholipid
- GPCR
- G protein-coupled receptor
- Edg
- endothelial differentiation gene
- NC-IUPHAR
- International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification
- SPC
- sphingosylphosphorylcholine
- LPC
- lysophosphatidylcholine
- The American Society for Pharmacology and Experimental Therapeutics
References
In this issue




{kind=link}
Jump to section
- Article
- Abstract
- I. Introduction
- II. Discovery of Lysophospholipid Receptors
- III. Receptor Nomenclature
- IV. Nonhuman Lysophospholipid Receptors
- V. Lysophospholipid Receptor Ligands
- VI. Lysophospholipid Receptor Gene Knockouts
- VII. Lysophospholipid Receptor Expression
- VIII. Conclusion
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters